
Abstract
Halophiles are among the best model microbes to study cellular adaptation. Haloferax, Haloarcula, Haloquadratum, Halobacteriales and many other halophiles living in the Great Salt Lake can survive extremely high salt concentrations. Many of them cannot survive if moved from hypersaline to non-saline habitats, and die from immediate cell lysis. The unique biological survivability of these bacteria in the saline condition is dictated by the phylogenetic dexterity at the given environmental constraints. Microbial adaptation has played a major role when bacteria in all forms branched out into different environmental niches, arising from the hypersaline conditions of the primordial sea. Their pervasive nature, created by ever-changing environmental conditions, was acquired by microbes from millions of years of making. The adaptation of enzymes during the course of their evolutionary development and some metabolic differences helped them expand and achieve environmental diversity. Metabolic processes, like osmoregulation in halotolerant cells, dictate the regulation of the bacterial cell membrane. For example, glycerol metabolism has been linked to osmoregulation in some halophilic microbes. Some mechanisms behind osmoregulation in halophiles are extremely energetic since they live in hostile environments. Pathways of such regulations, including de novo synthesis of solutes anabolically and/or extraction of solutes environmentally, help them to produce secondary metabolites like poly-β-hydroxyalkanoates. How halophiles maintain high metabolic similarity to other non-saline bacteria while showing different survivability under heavy salt stress remains an important question. This review attempts to outline progress in our understanding of their environmental diversity and biological survivability.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
6.1 Introduction
In recent years, modern tools and techniques have provided insight into the selection and optimization of extremophiles for commercial uses (Tango et al. 2002). The potential application of halophiles in bioconversion processes has emerged as a promising field for future biofuel conversion technology (Oren 2010). The breakdown of biomass materials to gain energy, and the cleanup of contaminated salty lands and aqueous sites are particularly important potential uses as reported (Rodriguez-Valera 1991). The exploration of their biodiversity in extreme salty environments has opened a new era for biotechnologists to use various techniques including genomics and proteomics. For example, in the last 10 years genomic sequencing and proteomic analysis have vastly expanded since the genomics studies of Halobacterium NRC-1 was first published in the year 2000 (Ng et al. 2000). Since then, the complete genomic sequences of many halophiles and halotolerant bacteria were published, including Haloarcula marismortui (Baliga et al. 2004), Natronomonas pharaonis (Falb et al. 2005), Haloquadratum walsbyi (Bolhuis et al. 2006), Halomicrobium mukohataei (Tindall et al. 2008), Halorhabdus utahensis (Bakke et al. 2009), Halogeometricum borinquense (Malfatti et al. 2009), Haloterrigena turkmenica (Saunders et al. 2009), and Haloferax volcanii (Hartman et al. 2010).
Halophiles are salt‐loving microbes and multiply quickly in saline environments. They are predominantly concentrated in bacterial group while some fungi (such as Wallemia ichthyophaga) and others algae (Dunaliella salina). Among the prokaryotes, the domain Archaea has several extremophilic genera of halophiles, like Haloferax, Haloarcula, Haloquadratum, Halobacteriales etc. The bacterial domain also has halophiles like Salinibacter, but they are less common when compared to archaea. Halophilic bacteria come in many sizes and shapes and live in variable salt stress conditions. For example, the bacterium Thiomargarita namibiensis, found in the ocean sediments of the continental shelf of Namibia, is the largest bacteria ever discovered. The second largest bacterium, Epulopiscium fishelsoni, was found in the Red Sea within the marine surgeonfish (Angert et al. 1993) (Fig. 6.1a). This bacterium is a million times larger than E. coli, and one can see it with the naked eye (Fig. 6.1b).

(a, b) The largest bacterium Epulopiscium fishelsoni found in the Red Sea within the marine surgeonfish was isolated by D. A. Angert, and reported in 1993 (a) This bacterium is a million times larger than E. coli, as shown in the picture* as a dot, and one can see E. fishelsoni with the naked eye (b) (Picture permission was obtained from Dr. D. Angert, Cornell University, Ithaca, New York)
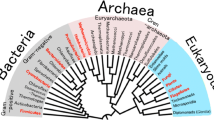
The family Halobacteriaceae, which contains 15 genera, is a member of the domain Archaea and comprises the majority of the prokaryotic extremophiles (Oren 2002a). Due to the presence of halophiles like Salinibacter, Haloquadratum, Halobacteriales, and Dunaliella salina, many lake and saltern habitats display a pink or reddish color. Hence, most known halophiles such as Halobacterium, Haloferax, Halococcus, Halorubrum, Halogeometricum, Haloterrigena and Haloarcula have become popular models for habitat adaptation studies in the archaeal domain. This review will focus on the exploitation of halophilic bacteria and its environmental adaptability and biological survivability in salt stress conditions.
6.2 Habitat and Environmental Adaptation
Microbial adaptation has played a major role in the existence of Halophiles, especially when bacteria in all forms and shapes branched out into different environmental niches (Rajendran et al. 2008), when they were evolving from the hypersaline primordial sea (Dundas 1998). High salty salterns or lakes represent an exceptional ecosystem in which several bacteria, especially halotolerants, can survive under the high salt pressure. Halophiles can thrive not only in those small salt water bodies such as salterns and lakes but also in large oceans and benthic floors of the sea. Halophilic bacteria are some of the best examples of environmental adaptation. With a heavy salt presence in the surrounding area, these microbes have developed metabolic conditions favorable to their survival. Some may be found within fluid inclusions in salt crystals (Norton and Grant 1988) and others produce compatible solutes to adapt high salt-stress conditions. The requirement for sodium chloride plays a major role in the metabolism of extremely halophilic bacteria, but they also require magnesium. As the growth of halophiles is predominantly dictated by the concentration of salt in the water, most moderate and slight halophilic bacteria do not require magnesium (Grant et al. 2001).
Halophilic bacteria are more abundant than any other groups in high salt conditions such as the Dead Sea (Bodaker et al. 2010), saline lakes in Inner Mongolia (Pagaling et al. 2009), African soda lakes, and deep-sea brines (van der Wielen et al. 2005) etc. The water salinity (Fig. 6.2) for fresh water is measured as less than 0.05 %, brackish water is 0.05–3 %, saline water is 3–5 % and Brine is more than 5 % (Anati 1999). Halophilic bacteria multiply better at the temperature of 28–37 °C with a pH range of 7.0–8.0 at 5–20 % sodium chloride concentration. Based on their salinity habitat and extent of their halotolerance, they were grouped into extremely (15–32 % w/v), moderately (3–15 % w/v), and slightly (1–3 % w/v) halophilic (Oren 2013). The slight halophiles generally prefer 0.3–0.8 M NaCl (Sodium chloride 1.8–4.7 % – seawater is 0.6 M or 3.5 %), moderate halophiles 0.8–3.4 M (4.7–20 %), and extreme halophiles 3.4–5.1 M (20–30 %). Some of the regular halophiles are halotolerant, meaning they do not require high NaCl condition, but can grow under saline conditions (Bowers et al. 2009).

Four different saline aqueous environments such as Freshwater, Brackish water, Saline water and Brine water were pictured with examples. Their salinity contents were expressed in percentage as well as in the traditional parts per thousand units
Halophiles can be aerobic or anaerobic (Ollivier et al. 1994). Their cellular uniqueness, adaptation and survivability are dictated by the phylogenetic dexterity at given environmental constraints. For example, the osmotic stress in the surrounding space makes the saline water enter into the cytoplasm through the membrane and non-halophiles may not survive under this stress. However, halophiles and halotolerant microbes maintain turgor pressure at least equal to the ambient pressure (Mustkhimov et al. 2010). In that case, anaerobic halophilic bacteria Haloanaerobiales, acetogenic anaerobes such as Halobacterioides, Sporohalobacter, and Acetohalobium can all thrive well in those high salt water bodies. Those that maintain aerobic life are also able to tolerate low salt pressure like Halobacteriaceae, and depend on other forms of nutrients and become phototrophic, fermentative, sulfate-reducing, homoacetogenic, methanogens etc. (Oren 2002a). Some show heterotrophic, phototrophic, photosynthetic, methenogenic or litotrophic nature. Analysis of microbial diversity in lakes and salterns using metagenomic studies (Bodaker et al. 2010) are useful in order to show biological adaptability and nutritional habits.
There are many hypersaline environments found around the globe and different types of halophilic bacteria survive in different salt stresses as presented to them by the environment (Gunde-Cimerman et al. 2005). Within each ecosystem, well-adapted microfloras flourish and in some cases possess unique resident microbiota within themselves. Some live conveniently on the surfaces of large marine ecosystems and some in the far more complex saline systems including the various trophic levels of ocean, deep sea, mangrove saline sites, arid, islands and coastal area, underground salt mines and saline caves. Some reservoirs of natural salt sources such as the Great Salt Lake (Utah), Owens Lake (CA), halite cores from Saline Valley (CA), Thane Papke (Storrs, CT), deep-brines of the Red Sea (Antunes et al. 2003), Xinjiang salt lake, Lake Assal in Djibouti (French Somaliland), Chinese salt mine, Goa salterns in India, salterns of Israel, the Salar de Atacama, Chile, Turkish salt mine, Iranian salt lakes hypersaline lake in Argentina, La Malá saltern near Granada, salterns of Spain, Western Australian hypersaline lake (Lake O’Grady North), Quidam Basin Quaternary sediments, Gabara in the Wadi Natrun, Egypt, brine wells in southwestern China, The South China Sea, a Korean salt flat, Venere Lake, Pantelleria Island Italy, the Yellow Sea, Mongolian salt and soda lakes and south Siberian hypersaline lakes, Mexican soda environments, salted hides), benthic floor of the Mediterranean Sea, the Gulf of Mexico, and soda lakes of the Kulunda Steppe (Sorokin et al. 2010) are ecologically significant as they are considered rich in halophiles.
6.3 Cellular Adaptation and Biological Survivability
Marine microorganisms and halophiles in particular play a major role in the atmospheric oxygen stability. Halophiles perform a recycling function similar to other diverse non-halophiles (Kastritis et al. 2007) in spite of their salt-stress cellular adaptations. The higher rate of oxygen production comes from a significant marine bacterium, Cyanobacterium, which predominantly occupies the surface of global saltwater. This individual species, C. prochlorococcus is very diverse due to its adaptability and shows 96 variations of strains sampled from southeast of Bermuda in the Atlantic Ocean. Such distinct variable markers within a single species indicates that this species would have originated millions of years ago and diverted into ecologically distinct groups because of the genomic diversity with hypervariable genes (Kashtan 2014). Others, like Haloferax, Haloarcula, Haloquadratum, Halobacteriales and many more can survive extremely high salt concentrations and also produce a high rate of oxygen.
Many halophiles cannot show optimal growth if moved from hypersaline to less-saline habitats (Mustkhimov et al. 2010). During the course of evolutionary development in several millions of years, the enzymes and other biomolecules of halotolerant microbes have been modified to function efficiently at high intracellular salt concentrations. Hence they may die from immediate lysis of cell if the salt conditions are changed drastically. For example, in Haloarcula marismortui, the behavior of membrane proteins is highly active in high-salt environments in support of ribosome and molecular adaptations of malate dehydrogenase and other enzymes (Madern et al. 2000). The acidic surface of the macromolecule allows protein-salt interactions that avoid water or salt enrichment at the surface of the protein and preserve its solubility (Ebel and Zaccai 2004). In a situation where the concentration of salinity increases, the number of diversified groups of halotolerant bacteria living in the vicinity of such hypersalinity conditions decreases. Since high salinity represents an extreme condition, relatively few microbes can survive in those conditions (Gunde-Cimmerman et al. 2005). Even if the rate of salinity is inconsistent in the aqueous environment, they can often survive due to their defensive mechanisms in order to prevent a desiccation type of dehydration.
The osmosis process is the most common method of preventing water loss among halophiles in a hypersaline stress condition (Mustkhimove et al. 2010). In this way, organic compounds known as osmoprotectants or compatible solutes such as amino acids, sugars, ectoines, betaines etc. are accumulated in the cytoplasm to prevent the desiccation. Some groups of halophiles such as Salinibacter ruber of the Halanerobiales group adopt another mechanism, using an influx of potassium ions into cytoplasm. Both mechanisms increase the internal osmo-regularity condition of the microbial cell. Such adaptation of cells helps most halophiles to sustain their viability in their native high salt conditions. If they were removed and left in a freshwater environment, their cells often burst and may not survive due to the change of osmotic pressure. The osmoregulatory mechanism with the help of osmoprotectant or compatible solutes is the most adapted cellular mechanism from an evolutionary point of view. Compared to the mechanism of K+ ion influx into the cytoplasm, the compatible solute mechanism helps halophiles gain more ATPs for other uses. More structural protein molecules are needed in the mechanism of influx of K+ ions into the cytoplasm. Since the compatible solutes often act as high salt stress protectants, most halophiles follow this method of osmoregulation (Santos et al. 2002).
6.4 Pathways of Regulation and Evolutionary Adaptability
The ubiquitous nature of halophiles in the salty environment was acquired by microbes from millions of years of making. It occurred due to the ever-changing environmental conditions due to the climate variations and is dictated by the regulation of the bacterial cell membrane like osmoregulation in hypersaline natural habitats. Some mechanisms behind osmoregulation in halophiles are extremely energetic (Saum and Müller 2008), and adaptation of enzymes during the course of their evolutionary development, made some differences in environmental diversity (McGenity et al. 2000), and helped halophilic cells to develop a metabolic variation. In order to adapt to stress and damage to proteins, halophiles possess more acidic residues especially glutamate (Kuntz 1971). Glutamate has a water binding nature higher than any other amino acid (Saenger 1987). Such evolutionary development helps halophiles to adapt to the salt stress by binding more compactly with water molecules (Lanyi 1974; Bolhuis et al. 2008). Beyond this, extreme halophiles have increased the number of acidic glutamic and aspartic acids (Kennedy et al. 2001), and charged amino acids (Fukuchi et al. 2003) on the surface of active proteins (Tadeo et al. 2009). Since glutamate residues have a superior water binding capacity over all other amino acids, they are generally found in excess on the surface of halophilic proteins.
The catabolic ability of a microbe depends on the environment (Hough and Danson 1999) where it tries to survive, by overcoming the chemical rejections or autointoxication. For example, Thermus aquaticus is a bacterium that lives in hot springs and hydrothermal vents, and Taq polymerase was identified (Chien et al. 1976) as an enzyme able to withstand the protein-denaturing conditions (high temperature) required during PCR (Saiki 1988). In salty conditions extremophiles develop evolutionary variations to have specific hyperenzymatic reactions against given substrates such as highly concentrated sodium chloride. Such ability leads extremophiles to competitively survive better than any other group of microbes in a saltwater body, such as the ocean. Since one third of the biosphere is covered with salty water, it is obvious that halophiles are a dominant group (Gomes and Steiner 2004). Protein hydration needs free water molecules, but in the ocean, water molecules are sequestered in hydrated ionic structures (Danson and Hough 1997). The evolutionary adaptation of halophilic cells to high concentrated salt condition depends on the ability of the catalytic protein dehydrogenase (Britton et al. 2006) or synthetase which they harbor. Such enzymes provide a cover to halophiles or an ability to adapt the influx of salt to regulate the osmotic pressure (DasSarma et al. 2010). The three dimensional structural features of the family of halophilic proteins (Frolow et al. 1996), and their functional stabilities against salt stress supports their survivability (Dym et al. 1995)
6.5 Exploitation of Halophilic Bacteria
6.5.1 Secondary Metabolic Products
Moderate halophiles accumulate high cytoplasmic concentrations of organic compounds to cope with the osmotic stress and to maintain positive turgor pressure (Graf et al. 2008). The natural ability of halophiles to produce and accumulate high concentrations of these low-molecular-weight compounds makes moderate halophiles useful for the biotechnological production of these osmolytes (Santos and da Costa 2002) Some these compatible solutes, especially glycine, betaine and ectoines have gained considerable attention in the recent past. They are good stabilizers of enzymes, nucleic acids, membranes and whole cells (Louis et al. 1994). They are used as stress protectants against high salinity, thermal denaturation, desiccation, and freezing. The industrial uses of these compounds in enzyme technology are used in biosensors and PCR technologies as well as in pharmaceuticals and cosmetics (Vendosa 1995). Salt labile enzymes such as lactate dehydrogenase and phosphofructokinase can be protected by using several compatible solutes such as betaine, trehalose, glycerol, glycine proline, ectoines and hydroxyectoine (Louis et al. 1994). Ectoine is as an active ingredient for many epidermal applications. This medically exploitable product commercially manufactured by a German company BitopAG from Halomonas elongata is one the most successful bacterial resources for Ectoine (Graf et al. 2008). Currently ectoine becomes part of compounding industries in medicinal product preparations (Graf et al. 2008). The derivatives of ectoine are used in Microarray and PCR technology as biomolecular protectants (Schnoor et al. 2004). Many of them were used as building blocks for some pharmaceuticals. Novel antibiotics were isolated from Halophilic Actinomycetes. Nocardiopsis, Saccharomonospora, and Streptomonospora were also tested in the context of cytotoxic effect against a range of cancer cell lines.
Some commercially significant products are obtained from halophiles such as β-carotene from Dunaliella, bacteriorhodopsin from Halobacterium, and ectoine from Halomonas (Oren 2010). For example, Halomonas elongata is used to produce ectoine, the active ingredient of many cosmetics and skin care products. Ectoine and hydroxyectoine biosynthesis is widely found in halophilic and halotolerant microorganisms, and can only be produced biologically. The production of ectoine for industrial purposes by using H. halodenitrificans is known since 1996. The halophilic cells were grown in an anaerobic fed-batch fermentation process, in a synthetic medium at high concentration with glycerol as the carbon source. Once the cell density reached a certain threshold, ectoine was extracted by “bacterial milking”, a process previously developed for Holomonas elongata, in which an osmotic down-shock applied from 10 to 2 % NaCl, results in excretion of about 80 % of the intracellular ectoine to the surrounding medium (Van-Thuoc et al. 2010). Subsequent exposure of the cells to a hyperosmotic shock from 2 to 10 % NaCl restored the original level of ectoine in a period of 10 h. Thus, a yield of 2 g ectoine per liter of medium per day was obtained. Ectoine and hydroxyectoine can also be produced by the Gram-positive moderate halophiles like Marinococcus strain M52. A maximum yield of 35.3 g/l cell dry weight was achieved as reported (Frings et al. 1995). The use of a dialysis reactor, in which cells were grown in an inner chamber fed with fish peptone and glucose, resulted in a dramatic increase in yield up to 132 g of cell dry weight with about 20 % hydroxyectoine obtained (Krahe et al. 1996). Another organism used for the production of ectoine is the actinomycetes Nocardiopsis lucentensis A5-1 (Yassin et al. 1993), isolated from a saline soil near Alicante, Spain (Lippert and Galinski 1992).
Halophiles, while surviving under high salt stress, can also produce secondary metabolites like poly-β-hydroxyalkanoates or bioplastics, halophilic enzymes, and biofuel. Some halophilic bacteria such as Halomonas boliviensis and members of genus Haloferax produce poly-β-hydroxyalkanoates commonly known as bioplastic (Quillaguaman et al. 2010). Many useful enzymes obtained commercially from halophilic microbes such as Salinibacter are applied in the detergent and the textile industries. Some enzymes such as proteases, cellulases, lipases, amylases, and mannanases are produced from various halophiles (Min et al. 1993) including but not limited to the group of Haloarchaea. A novel halophilic dehydrogenase was created using site directed mutagenesis from Halobacterium salinarum glutamate dehydrogenase (GDH). Due to their high salt and heat tolerance ability, some of them such as Haloferax volcanii (Large et al. 2007), are used as industrial biocatalysts and production of alcohol from ketones. Some halophiles like Alcanivorax dieselolei, Marinobacter and Halomonas sp., are able to degrade various polyaromatic hydrocarbons (Fig. 6.2). Another process in which halophiles may contribute in the future is the production of biofuel. Some halophilic bacteria have shown that they can break down cellulosic biomass via fermentation. The yield of ethanol and hydrogen could be a good source of biofuel, which opens up a new line of commercial uses for halophiles.
In recent years the poly-β-hydroxyalkanoate-based bioplastics has stimulated higher expectations for biotechnologists as ingredients for many house-hold products. Such biodegradable polymers are produced from Halomonas boliviensis (Quillaguaman et al. 2010). A successful attempt was made by mixing the biopolymer from H. boliviensis and the compatible solute ectoine from H. elongata with hydroxyectoine to gain a synthetic high-value product (Van-Thuoc 2010). A commercial halophilic strain of Haloferax was used to produce Tirel, a bioplastic product by Metabolix. The genus Haloferax (H. volcanii and H. mediterranei) was tested (Han et al. 2009) for its polyhydroxyalkanoate (PHA), and Haloarcula hispanica was also compared (Lu et al. 2008) for its poly (3-hydroxybutyrate-co-3-hydroxyvalerate) synthase.
6.5.2 Halophilic Remediation
Biological remediation is well known for its ability to take up and concentrate contaminants in the biological tissues without destroying the environment. Microbial remediation in particular has been developed from a conceptual methodology to a viable technology for contaminant clean-up (Rajendran et al. 2013). The potential applications of halophiles as remediators of toxic chemicals and biological wastes were studied well in the past (Ventosa and Nieto 1995). The halophiles are relatively less exploited and applied in the commercial sectors, compared to other extremophiles with the notable exceptions of β-carotene, bacteriorhodopsin, and ectoine (Oren 2010). Some of those commercially successful products from halophiles are employed as surfactants in olive oil industries to recover hypersaline waste brines, and some were used in fur-curing leather industries. The halophilic bacteria can be used to remove phosphate from saline environments and it is known to be a cost-effective method (Ramos-Coremzama et al. 1991).
Many toxicants persist hundreds of years in large and small bodies of water such as seas and lakes not only because of natural disasters but also due to man-made wastes. These wastes can arise from smelter and mining sites, manufactured gas plants, ammunition factories, run-over contaminants from fertilized farmland, sludge from agriculture, and pollutants from industrial and municipal waste-dumping grounds. These wastes are now reaching the oceans (Rajendran et al. 2008). Many chemical contaminants, especially hydrocarbon-based petroleum contaminants, are very problematic for the environment (Schwab et al. 1999). Remediation of such contaminants from the land and aqueous sites, using non halophiles, is a very difficult and slow process. Only 25 % of petroleum-contaminated land sites were being bioremediated using Pseudomonas (Holden et al. 2002). Beyond this, using those microbes in high salt environments is difficult due the effect of salt on the cell membrane and for other metabolic reasons. The application of halophiles as remediator in petroleum and other contaminants land and aqueous environment will have an impact on cleaning up our chemical toxic sites and will become increasingly important in the cleanup of aquifers and large plumes.
In halophilic remediation process, toxic heavy metals and organic pollutants are the major targets for a potential remediation (Roberts 1987). Many chemicals like polycyclic aromatic hydrocarbon (PAH) cause many health problems (Pradhan et al. 1998). Enhancement of PAH dissipation in soil and aqueous environment is often suggested to be a result of human impact. For example, PAHs is one of the dominant contaminants found in the superfund sites and resources conservation and recovery act (RCRA) sites (Kevein et al. 2006). One of the methods that can be effectively applied for cleaning-up many toxicants like PAHs is using microbes. Other major toxic chemicals and its species are Polychlorinated biphenyls, Chlorinated benzoic acid, Mercury (Hg), Hexachlorobiphenyl, 2,4,6-trinitrotoluene (TNT), Naphthalene, Pyrene, Chloroacetamide herbicides, Benzo(a)pyrene, 3,3′4,4-tetra chloroazobenzene (TCAB), Arsenic (As), metals such as Nickel (Ni), Zinc (Zn), Copper (Cu), Rubidium (Rb), Cesium (Cs), Manganese (Mn), Iron (Fe), Selenium (Se), Chromium (Cr), Cadmium (Cd), Lead (Pb), or radioactive isotopes such as Uranium, Cesium-137, Strontium or Cobalt (Rajendran et al. 2013).
Microbial species uptake, metabolize and accumulate toxic chemicals and metals. Environmental cleaning of such chemical contaminants by bacteria is varied from bacterial species to species, and based on the chemical contaminant in question. Some species act as powerful remediators of the primary chemical contaminants but others remediate only the chemical species of the primary contaminants. For example, uptake of Selenium, Trichloroethylene (TCE) Tetrachloroethylene (PCE) and its speciation such as Trichloroethanol, and Trichloroacetic acid as well as Nitroaromatic compounds such as Aminodinitrotoluene (ADNT), Diaminonitrotoluene (DANT), Hexahydro-1,3,5-trinitro-1,3,5-triazine (RDX) and Octahydro-1,3,5,7-tetranitro-1,3,5,7,-tetraazocine (HMX), Brominated compounds such as Ethylene dibromide, 2,4,5-Trichlorophenoxyacitic acid (2,4,5-T), Carbon Tetrachloride and Tetrachloroethlene, and Dibromochloropropane as well as nonhalogenated compounds such as methyl-t-butyl ether (MTBE) are significantly present in many contaminated sites and aquifers (Rajendran et al. 2013).
Many bacteria can be directly used to remediate toxic wastes including PCBs (Roberts 1987). As the result, the accumulated biomass, similar to that of “commercial ore” can be recycled. It can also be decomposed into manure or disposed-off into a traditional landfill (Rajendran et al. 2008). The association of halophilic microbes and toxic contaminants (Table 6.1) therefore offer a viable means of accomplishing the remediation of contaminated aquifer and other sites like deep hypersaline anoxic basins (van der Wielen 2005). Such bioremediation processes have many advantages over physical and chemical means of remediation at the open surface of ocean and other large bodies of water such as lakes. They are environmentally safe (Philip et al. 2005), biologically feasible, and economically cheaper to remove contaminants from the crude oil wastes and oil spillage or a variety of other contaminants such as pesticides, solvents. At high risk sites of contaminants and benthic zones, halophilic remediation can be used as supportive method to remove contaminants (Ramos-Cormenzana 1991). Because of the ability of specific halophiles which can reach specific depths and clean-up the last remains of contaminants trapped in the benthic floor halophilic remediation could become a truly viable option.
6.6 Future Application of Halophiles
Problems of chemical contamination and its salvage will continue because of the continuous use of the metals, chemicals and its derivatives in our everyday life in one way or other. The tremendous use of chemicals today will have a profound effect on the environment tomorrow. Due to the synergistic chemical effect on public health (Barbosa et al. 1998), which poses a significant threat to our economy, application of halophilic microbes or its products will dictate the alternative approaches to solve such emerging risks. The other future application of halophiles is the use of halotolerant bacteria in breakdown of biomass materials to gain the energy in the form of ethanol as biofuel (Tango and Islam 2002). The current progress towards achieving an outcome-based approach as studied by many marine microbiologists to replace fresh-water algae expected to reduce high consumption of fresh water (as in the case of fresh-water algae) and to speed up the entire biofuel conversion process (Fathepure 2014). Diverting the established algal conversion technologies and accessory approaches towards halophilic bacterial biofuel conversion will not only reduce energy consumption for quick recovery of products but also offer a cost-effective way for making commercial products. The commercial success of ectoine (Bestvater et al. 2008) achieved through the biotechnology and genetic engineering synthesis has opened an array of products from halophiles to be commercialized using fermentation technology. Cloning of ectoine genes made it possible so that 1 day salt tolerance genes could be cloned in stable foods such wheat to make them grow in salty soils or use sea water to grow rice in paddy fields (Min-Yu et al. 1993).
6.7 Conclusion
Halophilic bacteria and their abilities to survive in the extreme salt condition have stretched our understanding not only of how metabolic processes played crucial roles in survivability but also in the intermediate processes involved in the evolutionary adaptability. The current proteomic and genomic studies on the stability, structural and functional analysis aided by modern molecular and genetic engineering approaches paved a way for scientists to develop a family of enzymes and products from Halophiles for commercial purposes. However, how halophiles maintain high metabolic similarities to other non-hypersaline bacteria, but show different survivability under heavy salt stress, remains an important question to be further studied.
References
Anati DA (1999) The salinity of hypersaline brines: concepts and misconceptions. Int J Salt Lake Res 8:55–70
Angert ER, Clements KD, Pace NR (1993) The largest bacterium. Nature 362(6417):239–241
Antunes A, Eder W, Fareleira P, Santos H, Huber R (2003) Salinisphaera shabanensis gen. nov., sp. nov., a novel, moderately halophilic bacterium from the brine-seawater interface of the Shaban Deep, Red Sea. Extremophiles 7:29–34
Bakke P, Carney N, Deloache W, Gearing M, Ingvorsen K, Lotz M, McNair J, Penumetcha P, Simpson S, Voss L, Win M, Heyer LJ, Campbell AM (2009) Evaluation of three automated genome annotations for Halorhabdus utahensis. PLoS One 4:e6291
Baliga NS, Bonneau R, Facciotti MT, Pan M, Glusman G, Deutsch EW, Shannon P, Chiu Y, Weng RS, Gan RR, Hung P, Date SV, Marcotte E, Hood L, Ng WV (2004) Genome sequence of Haloarcula marismortui: a halophilic archaeon from the Dead Sea. Genome Res 14:2221–2234
Barbosa AC, Silva SR, Dorea JG (1998) Concentration of mercury in hair of indigenous mothers and infants from the Amazon basin. Arch Environ Contam Toxicol 34:100–105
Bestvater T, Louis P, Galinski EA (2008) Heterologous ectoine production in Escherichia coli: by-passing the metabolic bottle-neck. Saline Syst 4:12
Bodaker I, Sharon I, Suzuki MT, Reingersch R, Shmoish M, Andreishcheva E, Sogin ML, Rosenberg M, Belkin S, Oren A, Béjà O (2010) The dying Dead Sea: comparative community genomics in an increasingly extreme environment. ISME J 4:399–407
Bolhuis H, Palm P, Wende A, Farb M, Rampp M, Rodriguez-Valera F, Pfeiffer F, Oesterhelt D (2006) The genome of the square archae on Haloquadratum walsbyi: life at the limits of water activity. BMC Genomics 7:169
Bolhuis A, Kwan D, Thomas TR (2008) Halophilic adaptations of proteins. In: Siddiqui KS, Thomas T (eds) Protein adaptation in extremophiles. Nova Science, New York, pp 71–104
Bowers KJ, Mesbah NM, Wiegel J (2009) Biodiversity of poly-extremophilic bacteria: does combining the extremes of high salt, alkaline pH and elevated temperature approach a physico-chemical boundary for life? Saline Syst 5:9
Britton KL, Baker PJ, Fisher M, Ruzheinikov S, Gilmour DJ, Bonete MJ, Ferrer J, Pire C, Esclapez J, Rice DW (2006) Analysis of protein solvent interactions in glucose dehydrogenase from the extreme halophile Haloferax mediterranei. Proc Natl Acad Sci U S A 2006(103):4846–4851
Chien A, Edgar DB, Trela JM (1976) Deoxyribonucleic acid polymerase from the extreme thermophile Thermus aquaticus. J Bacteriol 127(3):1550–1557
Danson MJ, Hough DW (1997) The structural basis of protein halophilicity. Comp Biochem Physiol A Physiol 117:307–312
DasSarma P, Coker JA, Huse V, DasSarma S (2010) Halophiles. Biotechnology. In: Flickinger MC (ed) Encyclopedia of industrial biotechnology, bioprocess, bioseparation, and cell technology. Wiley, Hoboken, pp 2769–2777
Dundas I (1998) Was the environment for primordial life hypersaline? Extremophiles 2:375–377
Dym O, Mevarech M, Sussman JL (1995) Structural features that stabilize halophilic malate dehydrogenase from an archaebacterium. Science 267:1344–1346
Ebel C, Zaccai G (2004) Crowding in extremophiles: linkage between solvation and weak protein-protein interactions, stability and dynamics provides insight into molecular adaptation. J Mol Recognit 17:382–389
Falb M, Pfeiffer F, Palm P, Rodewald K, Hickmann V, Tittor J, Oesterhelt D (2005) Living with two extremes: conclusions from the genome sequence of Natronomonas pharaonis. Genome Res 15:1336–1343
Fathepure ZB (2014) Recent studies in microbial degradation of petroleum hydrocarbons in hypersaline environment. Front Microbiol 5:173
Frings E, Sauer T, Galinski EA (1995) Production of hydroxyectoine: high cell-density cultivation and osmotic down shock of Marinococcus strain M52. J Biotechnol 43:53–61
Frolow F, Harel M, Sussman JL, Mevarech M, Shoham M (1996) Insights into protein adaptation to a saturated salt environment from the crystal structure of a halophilic 2Fe-2S ferredoxin. Nat Struct Biol 3:452–458
Fukuchi S, Yoshimune K, Wakayama M, Moriguchi M, Nishikawa K (2003) Unique amino acid composition of proteins in halophilic bacteria. J Mol Biol 327:347–357
Gomes J, Steiner W (2004) The biocatalytic potential of extremophiles and extremozymes. Food Technol Biotechnol 42:223–235
Graf R, Anzali S, Bünger J, Pflücker F, Driller H (2008) The multifunctional role of ectoine as a natural cell protectant. Clin Dermatol 26:326–333
Gunde-Cimerman N, Oren A, Plemenitas A (eds) (2005) Adaptation to life at high salt concentrations in archaea, bacteria, and eukarya. Springer, Dordrecht
Han J, Lu Q, Zhou L, Liu H, Xiang H (2009) Identification of the polyhydroxyalkanoate (PHA)-specific acetoacetyl coenzyme A reductase among multiple FabG paralogs in Haloarcula hispanica and reconstruction of the PHA biosynthetic pathway in Haloferax volcanii. Appl Environ Microbiol 75:6168–6175
Hartman AL, Norais C, Badger JH, Delmas S, Haldenby S, Madupu R, Robinson J, Khouri H, Ren Q, Lowe TM, Maupin-Furlow J, Pohlschröder M, Daniels C, Pfeiffer F, Allers T, Eisen JA (2010) The complete genome sequence of Haloferax volcanii DS2, a model archaeon. PLoS One 5:e9605
He Z, Zhou A, Baidoo E, He Q, Joachimiak MP, Benke P, Phan R, Mukhopadhyay A, Hemme CL, Huang K, Alm EJ, Fields MW, Wall J, Stahl D, Hazen TC, Keasling JD, Arkin AP, Zhou J (2010) Global transcriptional, physiological, and metabolite analyses of the responses of Desulfovibrio vulgaris Hildenborough to salt adaptation. Appl Environ Microbiol 76:1574–1586
Holden PA, LaMontagne MG, Bruce AK, Miller WG, Lindow SE (2002) Assessing the role of Pseudomonas aeruginosa surface-active gene expression in hexadecane biodegradation in sand. Appl Environ Microbiol 68:2509–2518
Hough DW, Danson MJ (1999) Extremozymes. Curr Opin Chem Biol 1:39–46
Kashtan N (2014) Single-cell genomics reveals hundreds of coexisting subpopulations in wild Prochlorococcus. Science 344:416–420
Kastritis PL, Papandreou NC, Hamodrakas SJ (2007) Haloadaptation: insights from comparative modeling studies of halophilic archaeal DHFRs. Int J Biol Macromol 41(4):447–453
Kennedy SP, Ng WV, Salzberg SL, Hood L, DasSarma S (2001) Understanding the adaptation of Halobacterium species NRC-1 to its extreme environment through computational analysis of its genome sequence. Genome Res 11:1641–1650
Krahe M, Antranikian G, Markl H (1996) Fermentation of extremophilic microorganisms. FEMS Microbiol Rev 18:271–285
Kuntz ID (1971) Hydration of macromolecules IV: polypeptide conformation in frozen solutions. J Am Chem Soc 93:516–518
Lanyi JK (1974) Salt dependent properties of proteins from extremely halophilic bacteria. Bacteriol Rev 38:272–290
Large A, Stamme C, Lange C, Duan Z, Allers T, Soppa J, Lund PA (2007) Characterization of a tightly controlled promoter of the halophilic Haloferax volcanii and its use in the analysis of the essential cct1 gene. Mol Microbiol 66:1092–1106
Lippert K, Galinski EA (1992) Enzyme stabilization by ectoine-type compatible solutes: protection against heating, freezing and drying. Appl Microbiol Biotechnol 37:61–65
Louis P, Trüper HG, Galinski EA (1994) Survival of Escherichia coli during drying and storage in the presence of compatible solutes. Appl Microbiol Biotechnol 41:648–688
Lu Q, Han J, Zhou L, Zhou J, Xiang H (2008) Genetic and biochemical characterization of the poly(3-hydroxybutyrate-co-3-hydroxyvalerate) synthase in Haloferax mediterranei. J Bacteriol 190:4173–4180
Madern D, Ebel C, Zaccai G (2000) Halophilic adaptation of enzymes. Extremophiles 4:91–98
Malfatti S, Tindall BJ, Schneider S, Faehnrich R, Lapidus A, LaButti K, Copeland A, Glavina Del Rio T, Nolan M, Chen F, Lucas S, Tice H, Cheng J-F, Bruce D, Goodwin L, Pitluck S, Anderson IJ, Pati A, Ivanova N, Mavromatis K, Chen A, Palaniappan K, D’Haeseleer P, Goeker M, Bristow J, Eisen JA, Markowitz V, Hugenholtz P, Kyrpides NC, Klenk H-P, Chain P (2009) Complete genome sequence of Halogeometricum borinquense type strain (PR3T). Stand Genomic Sci 1:150–158
McGenity TJ, Gemmell RT, Grant WD, Stan-Lotter H (2000) Origins of halophilic microorganisms in ancient salt deposits. Environ Microbiol 2:243–250
Min-Yu L, Ono H, Takano M (1993) Gene cloning of ectoine synthase from Halomonas sp. Annu Rep Int Cent Coop Res Biotechnol Jpn 16:193–200
Mueller EK, Shann JR (2006) PAH dissipation in spiked soil: impact of bioavailability, microbial activity, and trees. Chemosphere 64:1006–1014
Mustakhimov I, Reshetnikov I, Khmelenina AS, Trotsenko VN, Yu A (2010) Regulatory aspects of Ectoine biosynthesis in Halophilic bacteria. Microbiology 79:583–592
Ng WV, Kennedy SP, Mahairas GG, Berquist B, Pan M, Shukla HD, Lasky SR, Baliga NS, Thorsson V, Sbrogna J, Swartzell S, Weir D, Hall J, Dahl TA, Welti R, Goo YA, Leithausen B, Keller K, Cruz R, Danson MJ, Hough DW, Maddocks DG, Jablonski PE, Krebs MP, Angevine BM, Dale H, Isenbarger TA, Peck RF, Pohlschröder M, Spudich JL, Jung K-H, Alam M, Freitas T, Hou S, Daniels CJ, Dennis PP, Omer AD, Ebhardt E, Lowe TM, Liang P, Riley M, Hood L, DasSarma S (2000) Genome sequence of Halobacterium species NRC-1. Proc Natl Acad Sci U S A 97:12176–12181
Norton CF, Grant WD (1988) Survival of halobacteria within fluid inclusions in salt crystals. J Gen Microbiol 134:1365–1373
Ollivier B, Caumette P, Garcia J-L, Mah R (1994) Anaerobic bacteria from hypersaline environments. Microbiol Rev 58(1):27–38
Oren A (2002a) Diversity of halophilic microorganisms: environments, phylogeny, physiology and applications. J Ind Microbiol Biotechnol 28:56–63
Oren A (2002b) Molecular ecology of extremely halophilic Archaea and Bacteria. FEMS Microbiol Ecol 39:1–7
Oren A (2010) Industrial and environmental applications of halophilic microorganisms. Environ Technol 31:825–834
Oren A (2013) Life at high salt concentrations. In: Rosenberg E, Delong EF, Lory S, Stackebrandt E, Thompson F (eds) The prokaryotes – prokaryotic communities and ecophysiology. Springer, Berlin, pp 421–440
Pagaling E, Wang HZ, Venables M, Wallace A, Grant WD, Cowan DA, Jones BE, Ma Y, Ventosa A, Heaphy S (2009) Microbial biogeography of six salt lakes in Inner Mongolia, China, and in Argentina. Appl Environ Microbiol 75:5750–5760
Philip JC, Bamforth SM, Singleton I, Atlas RM (2005) Environmental pollution and restoration: a role for bioremediation. In: Atlas RM, Philip J (eds) Bioremediation: applied microbial solutions for real-world environmental cleanup. ASM Press, Washington, DC, pp 1–48
Quillaguaman J, Guzman H, Van-Thuoc D, Hatti-Kaul R (2010) Synthesis and production of polyhydroxyalkanoates by halophiles: current potential and future prospects. Appl Microbiol Biotechnol 85:1687–1696
Rajendran N (2013) The state of the rhizoinhabitants in bridging the gap between plant productivity and persuasiveness during remediation. In: Maheshwari DK (ed) Bacteria in agronomy: crop productivity. Springer, Berlin, pp 463–496
Rajendran N, Saraf M, Tank N (2008) Rhizomediation in soil for sustainable agriculture. In: Maheshwari DK, Dubey RC (eds) Potential microorganisms for sustainable agriculture: a techno-commercial perspective, 19th edn. I. K. International Publishing House, New Delhi, pp 364–393
Ramos-Cormenzana A (1991) Halophilic organisms and the environment. In: Rodriguez-Valera F (ed) General and applied aspects of halophilic microorganisms. Plenum Press, New York, pp 15–24
Roberts L (1987) Discovering microbes with a taste for PCBs. Science 237:975–977
Rodriguez-Valera F (ed) (1991) General and applied aspects of halophilic microorganisms. Plenum Press, New York, pp 40–44
Saenger W (1987) Structure and dynamics of water surrounding biomolecules. Annu Rev Biophys Biophys Chem 16:93–114
Saiki RK (1988) Primer-directed enzymatic amplification of DNA with thermostable DNA polymerase. Science 239(4839):487–491
Santos H, da Costa MS (2002) Compatible solutes of organisms that live in hot saline environments. Environ Microbiol 4:501–509
Saum SH, Müller V (2008) Regulation of osmoadaptation in the moderate halophile Halobacillus halophilus: chloride, glutamate and switching osmolyte strategies. Saline Syst 4:4–6
Saunders E, Tindall BJ, Fahnrich R, Lapidus A, Copeland A, Glavina Del Rio T, Lucas S, Chen F, Tice H, Cheng J-F, Han C, Detter JC, Bruce D, Goodwin L, Chain P, Pitluck S, Pati A, Ivanova N, Mavromatis K, Chen A, Palaniappan K, Land M, Land M, Hauser l, Chang Y-J, Jeffries CD, Brettin T, Rohde M, Goker M, Bristow J, Eisen JA, Markowitz V, Hugenholtz P, Klenk H-P, Kyrpides NC (2009) Complete genome sequence of Haloterrigena turkmenica type strain (4kT). Stand Genomic Sci 2:107–116
Schnoor M, Voß P, Cullen P, Böking T, Galla HJ, Galinski EA, Lorkowski S (2004) Characterization of the synthetic compatible solute homoectoine as a potent PCR enhancer. Biochem Biophys Res Commun 322:867–872
Schwab AP, Su J, Wetzel S, Pekarek S, Banks MK (1999) Extraction of petroleum hydrocarbons from soil by mechanical shaking. Environ Sci Technol 33:1940–1945
Sorokin DY, Rusanov II, Pimenov NV, Tourova TP, Abbas B, Muyzer G (2010) Sulfidogenesis at extremely haloalkaline conditions in soda lakes of Kulunda Steppe (Altai, Russia). FEMS Microbiol Ecol 73:278–290
Tadeo X, Lopez-Mendez B, Trigueros T, Lain A, Castano D, Millet O (2009) Structural basis for the amino acid composition of proteins from halophilic archaea. PLoS Biol 7
Tango MSA, Islam MR (2002) Potential for extremophiles for biotechnological and petroleum applications. Energy Sources 24:543–559
Tindall BJ, Schneider S, Lapidus A, Copeland A, Glavina del Rio T, Nolan M, Lucas S, Chen F, Tice H, Cheng J-F, Saunders E, Bruce D, Goodwin L, Pitluck S, Mikhailova N, Pati A, Mavromatis K, Ivanova N, Chen A, Palaniappan K, Chain P, Land M, Hauser L, Chang Y-J, Jeffries CD, Brettin T, Han C, Rohde M, Goeker M, Bristow J, Eisen JA, Markowitz V, Hugenholtz P, Klenk H-P, Kyrpides NC, Detter JC (2009) Complete genome sequence of Halomicrobium mukohataei type strain (arg-2T). Stand Genomic Sci 1:270–277
van der Wielen PW, Bolhuis H, Borin S, Daffonchio D, Corselli C, Giuliano L, D’Auria G, de Lange GJ, Huebner A, Varnavas SP, Thomson J, Tamburini C, Marty D, McGenity TJ, Timmis KN, BioDeep Scientific Party (2005) The enigma of prokaryotic life in deep hypersaline anoxic basins. Nature 307:121–123
Van-Thuoc D, Guzman H, Quillaguaman J, Hatti-Kaul R (2010) High productivity of ectoines by Halomonas boliviensis using a combined two-step fed-batch culture and milking process. J Biotechnol 147:46–51
Ventosa A, Nieto JJ (1995) Biotechnological applications and potentialities of halophilic microorganisms. World J Microbiol Biotechnol 11:85–94
Yassin AF, Galinski EA, Wohlfarth A, Jahnke K-D, Schaal KP, Trüper HG (1993) A new actinomycete species, Nocardiopsis lucentensis sp. nov. Int J Syst Bacteriol 43:266–271
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Rajendran, N. (2015). Environmental Diversity and Biological Survivability of Halophilic Bacteria. In: Maheshwari, D., Saraf, M. (eds) Halophiles. Sustainable Development and Biodiversity, vol 6. Springer, Cham. https://doi.org/10.1007/978-3-319-14595-2_6
Download citation
DOI: https://doi.org/10.1007/978-3-319-14595-2_6
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-14594-5
Online ISBN: 978-3-319-14595-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)