
Abstract
This chapter gives a basic introduction to the anatomy of the eye. This background is critical to understanding the physiology of the eye and in particular aspects of visual perception which are discussed elsewhere in this handbook.
Access provided by Autonomous University of Puebla. Download reference work entry PDF
Similar content being viewed by others



Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
Introduction
The human eye is the basic organ of sight. The mechanism of sight and visual perception involves a set of structures (each of which has a definite function). The eye is housed in a protective framework of bones and connective tissue, and this is called the orbit . The eyelids contain glands that produce the tear film layer over the anterior (front surface) of the eye, as well as protecting the anterior surface. The muscles that are attached to the eyeball and control the movement of the eyes are called extraocular muscles . In addition, the muscles are coordinated between the two eyes, a necessary condition for binocular vision. A complex network of blood vessels and neurons provide nutrients as well as sensory and motor innervations to the eye. The crystalline lens of the eye plays a major role in focusing the light rays through a process called accommodation controlled via the ciliary muscles. The retina, the innermost of various layers, contains the light-absorbing rod and cone photoreceptors, as well as a neural network to process and transmit the electrical signals via the optic nerve to the visual cortex in the brain via the lateral geniculate body. Figure 1 illustrates the main features of the human eye. It also shows the eye in its bony orbit. Additional details on the optical elements and functional and physiological properties of the eye can be found, for example, in the books by Grosvenor (1989), Oyster (1990), Remington (2005), and Hart (1992). Detailed description of the optics of the eye can be found in the book by Atchison and Smith (2000).

(a) The human eye in its bony orbit, showing the extraocular muscles (b) Anatomic cross section of the eye (From Walls 1942)
Basic Dimensions of the Eye
The eye is a spheroid structure that rests in the orbit on the frontal surface of the skull. The dimensions of the human eye are reasonably constant in adults, varying by only about a millimeter or so. The sagittal diameter (the vertical) is about 24 mm and is usually less than the transverse diameter which is about 24.5–25 mm. The adult human eye weighs approximately 7.5 g.
Eye Formation and Growth
Eye formation begins during the end of the third week of development when outgrowths of brain neural tissue, called the optic vesicles , form at the sides of the forebrain region. The major structures of the eye are initially formed by the fifth month of fetal development. The eye structures enlarge, mature, and form increasingly complex neural networks prenatally. At birth, infant eyes are about two-thirds the size of an adult eye. Until the first month of life, infants lack complete retinal development, with consequent effects on development of visual functions (i.e., visual acuity, contrast sensitivity, motion, etc.). From the second year until puberty, eye growth progressively slows. It should be noted that infants are born hyperopic (too much positive power), and a process of emmetropization occurs. The mechanism and development of emmetropia (as well as myopia development in children) is a major area of current research (e.g., Charman and Radhakrishnan 2010; Pang et al. 2006).
Layers of the Eye
The thick wall of the eye contains three layers: the sclera, the choroid, and the retina. The sclera , the white part of the eye, is the outermost layer. This is a thick layer and gives structural stability and shape to the eye. A delicate membrane called the conjunctiva covers the visible portion of the eye. A study (Pang et al. 2006) of 50 eye-bank eyes found that the mean scleral thickness ± SD was 0.53 ± 0.14 mm at the corneoscleral limbus (where the cornea and sclera meet), significantly decreasing to 0.39 ± 0.17 mm near the equator and increasing to 0.9–1.0 mm near the optic nerve. The mean total scleral surface area by surface area was found to be about 17.0 cm2. The sclera is optically opaque and the transparent cornea allows light rays to enter the eye.
Underneath the sclera is the second layer of tissue, the choroid, composed of a dense pigment and blood vessels that nourish the tissues. The vascular layer is also called the uvea . The choroid is a dense vascular network and supplies nutrients to the retinal layers. The choroid is thought to play a role in eye growth. Recent studies show that the choroid is about 426 μm and the thickness undergoes diurnal fluctuations (Brown et al. 2009). Optical coherence tomography (OCT) results reveal that thickness decreases with increasing axial length of the eye (Esmaeelpour et al. 2010). Near the center of the visible portion of the eye, the choroid layer forms the ciliary body, which contains the muscles used to change the shape of the lens (accommodation). The ciliary body in turn merges with the iris, a diaphragm that regulates the size of the pupil. The iris is the area of the eye where the pigmentation of the choroid layer, usually brown or blue, is visible because it is not covered by the sclera. The pupil is the round opening in the center of the iris; it is dilated and contracted by muscular action of the iris, thus regulating the amount of light that enters the eye, and is therefore like the aperture stop of the optical system of the eye. The change in pupil size not only controls the amount of light incident on the retina but can also affect the retinal image quality that is due to diffraction and depth of focus. The diameter of the eye can change because of factors such as illumination, age, accommodation, and drugs. Behind the iris is the lens, a transparent, elastic, but solid ellipsoid body that focuses the light on the retina, the third and innermost layer of tissue. Accommodation is the process by which the eye lens changes its shape (and increases its power) to bring a near object to focus. The mechanism of accommodation is discussed, for example, in Muller and Strobel (2007), Charman (2008), and Koretz (2000).
The retina is a network of nerve cells, notably the rods and cones, and nerve fibers that fan out over the choroid from the optic nerve as it enters the rear of the eyeball from the brain. Unlike the two outer layers of the eye, the retina does not extend to the front of the eyeball. The retina will be discussed in greater detail later in this chapter.
Accessory Structures
These include the lachrymal gland and its ducts in the upper lid, which bathe the eye with tears. The tear film layer is about a quarter of a wavelength thick and keeps the cornea moist and clean. The drainage ducts carry the excess moisture to the interior of the nose. The eye is protected from dust and dirt by the eyelashes, eyelid, and eyebrows.
In addition, there are six muscles which extend from the eye socket to the eyeball, enabling it to move in various directions. The extraocular muscles and their actions are shown in Fig. 2. There are two types of movement: conjugate (both eyes move in the same direction) and disjunctive (the eyes move in opposite directions). There is an antagonist-agonist reciprocal innervation for eye muscles. More details on oculomotor mechanisms and characteristics can be found in the book by Ciuffreda and Tannen (1995).

The extraocular muscles and their actions (Reprinted from Simon and Calhoun 1998)
The Humors of the Eye
The space between the cornea and iris known as the anterior chamber is filled with a thin watery liquid, which is optically clear and slightly alkaline, called the aqueous humor . The aqueous humor is secreted into the chamber by the ciliary body and is drained by the trabecular meshwork. The aqueous humor inflates the globe of the eye, maintains the intraocular pressure, and provides nutrients to the avascular structures of the eye, namely, the posterior cornea, lens, etc. The refractive index of the aqueous humor is taken to be about 1.336 (for sodium D line), and the anterior chamber has a volume of about 151 μm3 (Behndig and Markstrom 2007).
The space between the back of the lens and retina, called the posterior chamber , is filled with vitreous humor, a jellylike substance. The vitreous is colorless, transparent, and gelatinous. The viscosity of the material is about two to four times that of pure water. It too has a refractive index of about 1.336. The vitreous in contact with the retina helps to keep it in place by pressing against the choroid. In addition, unlike the aqueous, the vitreous is stagnant – it does not get replaced. Therefore, if blood, cells, or other debris get into the vitreous, they will cast shadows, diffraction effects, etc., on the retina and produce floaters. The depth of the posterior chamber is approximately 16.03 mm.
The Cornea
The cornea is the major structure that optically aids in retinal image formation. If the unaccommodated eye has a power of about 60 D (i.e., the eye is looking at an object at optical infinity), then the cornea contributes roughly 42 D of this effective power. The cornea is the first refractive surface element that light comes in contact with. The cornea has an anterior radius of curvature of about 7.8 mm. The cornea is roughly 11.5 mm in diameter, has a thickness of about 0.5–0.6 mm in the center and about 0.8 mm in the periphery, and takes about one-sixth of the globe. It is slightly raised from the sclera at the limbus. Even though a single anterior radius of curvature is given, in reality, the cornea is spherical only in the central 1–3 mm zone. In the paracentral zone, approximately 3–4 to 7–8 mm, it is approximately a prolate spheroid; in the periphery, with the outer zone diameter of about 11 mm, there is greatest asphericity and flattening. At the limbus, the cornea steepens. The front surface of the cornea can be roughly modeled as an ellipsoid, with an eccentricity factor of about 0.6–0.8 (Applegate and Howland 1995). Because of the asphericity, the cornea minimizes spherical aberration and coma in the retinal image. Young’s modulus of elasticity of the cornea is 0.45–1.0 MPa and the Poisson ratio is about 0.49.
There are five layers of the human cornea. The cornea is completely transparent and hence has no blood vessels. There is also a systematic arrangement of collagen fibrils in a lattice formation. It gets its oxygen through direct diffusion from the air. The refractive index of the cornea is approximately 1.376. Because the change in refractive index between the cornea (posterior) and the aqueous is not very big, it contributes about 6 D to the overall refraction.
The Crystalline Lens
The lens, which is part of the anterior segment of the eye, is behind the iris of the eye. The lens is suspended in place by the zonular fibers which attach to the lens near its equatorial line and connect the lens to ciliary body. Posterior to the lens is the vitreous. The lens contributes about 18 D to the overall effective power of the eye. The lens has an ellipsoid, biconvex shape. The anterior surface is less curved than the posterior. In the adult, the lens is typically 10 mm in diameter and has an axial length of about 4 mm, though it is important to note that the size and shape can change due to accommodation and because the lens continues to grow throughout a person’s lifetime. During accommodation, for near objects, the ciliary muscle contracts, zonule fibers loosen and relax, and the lens thickens, resulting in a rounder shape and thus increasing refractive power. Changing focus to an object at a greater distance requires the relaxation of the ciliary muscle, which in turn increases the tension on the zonules, thereby flattening the lens and thus decreasing the lens. It should be noted that with age, this ability to accommodate decreases, resulting in a condition called presbyopia which affects people typically in their 40s. This decrease of accommodation with age is called Donder’s curve (Stark et al. 1985). The refractive index of the lens varies from approximately 1.406 in the central layers down to 1.386 in less dense cortex of the lens. The lens has a gradient index nature (Pierscionek 2010), which helps to reduce various optical aberrations. The lens grows throughout life; as a result, the gradient index (GRIN) structure is formed with the highest index in the lens nucleus and the lowest index in the capsule of the lens.
Even though the lens is completely transparent, it is made up of three layers: the lens capsule, the lens epithelium, and lens fibers. The lens capsule completely covers the lens and contributes to a higher curvature on the anterior side than the posterior side. The lens fibers form the majority of the lens material. Opacities in the lens are called cataracts , and if they are large enough to interfere with vision, they are surgically removed, and an artificial intraocular lens is substituted.
The Retina
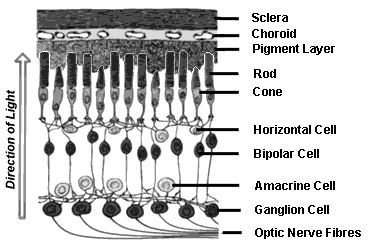
The retina is the fundamental sensory layer of the eye. The cross section of the retina is shown in Fig. 3. Note that light comes in from below in the diagram. It contains about 200 million photoreceptors, both rods and cones. These photoreceptors absorb visible light and convert the absorbed light into nerve impulses that are sent out to the brain via the optic nerve. The central retina contains the macula, which is a specialized area for perceiving fine detail and color. The center of the macula is called the fovea . The fovea contains only cone photoreceptors and is completely devoid of rods. The density of cones decreases as we go toward the periphery of the retina (Fig. 4; Osterberg 1935; Curcio et al. 1990). When a photoreceptor absorbs a photon, there is a series of photochemical events that result in a hyperpolarization of the photoreceptor cell. This electrical signal is transmitted down through the cell layers till they reach the optic nerve head/optic disk, a region devoid of photoreceptors. The region of the optic nerve head is called the blind spot . The optic nerve is made up of millions of nerve fibers that collect information from the eye and send it to the visual cortex of the brain (Rodieck 1998; Dowling 1987).

The structure of the retina (Reprinted from http://www.catalase.com/retina.gif)
{kind=link}

Distribution of rods and cones in the retina (Data from Osterberg 1935)
The human retina is approximately 0.2 mm thick and has an area of approximately 1,100 mm2 (about the size of a silver dollar). The various layers of the retina are as follows:
-
1.
Inner limiting membrane (ILM) is the boundary between the vitreous humor in the posterior chamber and the retina itself.
-
2.
Ganglion cell layer comprises the cell bodies and axons of ganglion cells.
-
3.
Inner plexiform layer (IPL) contains the synapses made between bipolar, amacrine, and ganglion cells. The thickness of this layer varies considerably across species, where “simpler” organisms (such as frogs, pigeons, and squirrels) possess thicker IPLs than “higher” organisms like primates. The thicker IPL indicates that these retinas perform more peripheral and specialized image processing.
-
4.
Inner nuclear layer (INL) contains bipolar cell and horizontal and amacrine cell bodies.
-
5.
Outer plexiform layer (OPL) contains bipolar cell, horizontal cell, and receptor synapses.
-
6.
Outer nuclear layer (ONL) contains the nuclei of photoreceptors.
-
7.
Outer limiting membrane (OLM) interfaces with the base of inner segments of photoreceptors.
-
8.
Photoreceptor layer contains the inner and outer segments of rod and cone photoreceptors. The photoreceptors contain the opsins (the various proteins) and the chromophores (the photon-catching molecules; consisting of retinal, an aldehyde of vitamin A). The absorption of a photon by the chromophore results in a conformational change of the chromophore, which in turn results in a photochemical cascade. The isomerization of 11-cis retinal to all-trans begins the process of phototransduction. The exact chain of events is that isomerization of photopigment breaks apart a molecule called transducin , which activates an enzyme called phosphodiesterase . Phosphodiesterase, in turn, breaks cGMP into its inactive form, which causes Na+ channels (which are open in the resting state) to close. Closing Na+ channels hyperpolarizes the neuron (unlike in other neuronal systems). Light stimulation thus causes fewer transmitters to be released at the synapse. The hyperpolarization of the outer segment spreads to the inner segment by electrotonic conduction. Since receptors are small, the receptor potential is still large at the axon terminal in the inner segment. Thus, most retinal neurons transmit information using only graded potentials. Some amacrine cells and all ganglion cells use action potentials. It is of interest to note that photoreceptors act as classical fiber optic devices and guide light to sites of absorption (Lakshminarayanan and Enoch 2010). Photoreceptors are found to be oriented toward the center of the exit pupil of the eye (Enoch and Lakshminarayann 1991).
-
9.
Pigment epitheliums (PEs) are darkly pigmented cells which absorb light not captured by photoreceptors, thus reducing scattering; they also play a role in “trimming” photoreceptors which follows a diurnal cycle. Diurnal species (active in bright-light environments) typically possess dark PEs; nocturnal species (active in dim-light environments) possess an adaptation called a tapetum . The tapetum is a mirrorlike layer behind the photoreceptors which reflects photons not captured by the photoreceptors back out the eye, thus giving the receptors a “second chance” to capture them. Sensitivity to light in these animals is thus increased by approximately twofold. The dominant wavelength of light reflected by the tapetum is usually close to the absorbance peak of rhodopsin (the photopigment contained by rods). People lack a tapetum lucidum; shining light into the eye while the pupil is dilated illuminates the blood-rich choroid behind the retina, thus the red eye seen in flash photos.
There are three types of cones, thus giving rise to trichromatic vision. The first responds maximally to light of long wavelengths, peaking in the yellow region (564–580 nm); this type is designated L for long wavelength-sensitive cones or red cones. The second type responds most to light of medium wavelength, peaking at green (534–545 nm), and is known as M cones or green cones. The third type responds most to short-wavelength light, of a violet color, the S cones (or blue cones), and has a peak of absorption at 420–440 nm (see Fig. 5). The packing arrangements of these three cone types were recently studied using adaptive optics techniques (Roorda and Williams 1999). The packing arrangements are important because of sampling issues and aliasing that can occur in human vision (Lakshminarayanan and Nygaard 1992). The absence of one (or more) of these cone types leads to various color deficiencies (see chapter “Color Vision Deficiencies” on color vision deficiencies). The rods on the other hand are more sensitive for low light levels and peak around 420 nm. The sensitivity curves for rods and cones are shown in Fig. 5.

Rod and cone sensitivity curves
As noted previously, when light falls on a receptor, it sends a proportional response synaptically to bipolar cells which in turn signal the retinal ganglion cells. The receptors are also “cross-linked” by horizontal cells and amacrine cells, which modify the synaptic signal before the ganglion cells. Rod and cone signals are intermixed and combine. Despite the fact that all are nerve cells, only the retinal ganglion cells and few amacrine cells create action potentials. Although there are more than 120 million photoreceptors, there are only about 1.2 million fibers in the optic nerve; a large amount of preprocessing is performed within the retina. The retina spatially encodes (compresses) the image to fit the limited capacity of the optic nerve. The retina does so by encoding the incoming images in a suitable manner. These operations are carried out by the center-surround receptive field structures as implemented by the bipolar and ganglion cells. These center-surround structures are functional and not anatomical. These center-surround structures of the ganglion and bipolar cells encode the information by performing edge detection and other tasks. The fovea produces the most accurate information. Despite occupying about 0.01 % of the visual field (less than 2° of visual angle), about 10 % of axons in the optic nerve are devoted to the fovea. That is, there is an almost 1:1 connection between each cone cell in the fovea and a ganglion cell. As a result, there is great spatial resolution in the fovea. However, in the periphery, there is a multiplexing of approximately ten rod cells to one ganglion cell, and hence, resolution falls off considerably (see chapter “Visual Acuity”). The information capacity of the retina is estimated at 500,000 bits per second without color or around 600,000 bits per second with color coding.
Summary
In this chapter, a brief summary of ocular anatomy is given. Emphasis has been given to topics dealing with visual optics and retinal function.
Abbreviations
- GRIN:
-
Gradient index
- ILM:
-
Inner limiting membrane
- INL:
-
Inner nuclear layer
- IPL:
-
Inner plexiform layer
- OCT:
-
Optical coherence tomography
- OLM:
-
Outer limiting membrane
- ONL:
-
Outer nuclear layer
- PEs:
-
Pigment epitheliums
Further Reading
Applegate RA, Howland HC (1995) Non-invasive measurement of corneal topography. IEEE Eng Med Biol 14:30–42
Atchison DA, Smith G (2000) Optics of the human eye. Butterworth Heinemann, St. Louis
Behndig A, Markstrom K (2007) Determination of aqueous humor volume by 3-D mapping of the anterior chamber. Ophthalmic Res 37:13–16
Brown JS, Flitcroft D, Ying G, Frances EL et al (2009) In vivo human choroidal thickness measurements: evidence for diurnal fluctuations. Invest Ophthalmol Vis Sci 50:5–12
Chalupa L, Werner J (2000) The visual neurosciences, vol 2. MIT Press, Cambridge, MA
Charman WN (2008) The eye in focus: accommodation and presbyopia. Clin Exp Optom 91:207–225
Charman WN, Radhakrishnan H (2010) Peripheral refraction and the development of refractive error: a review. Ophthalmic Physiol Opt 30:321–338
Ciuffreda K, Tannen B (1995) Eye movement basics for the clinician. C.V.Mosby, St. Louis
Curcio CA, Sloan KR, Kalina RE, Hendrickson AE (1990) Human photoreceptor topography. J Comp Neurol 292:497–523
Davson H (1972) The physiology of the eye, 3rd edn. Academic, New York
Dowling J (1987) The retina: an approachable part of the brain. Belknap, Cambridge, MA
Enoch JM, Lakshminarayann V (1991) Retinal fiber optics. In: Charman N (ed) Vision and visual dysfunction, vol I. McMillan Press, London, pp 280–309
Esmaeelpour M, Povazay B, Hermann B, Hofer B et al (2010) Three dimensional 1060 nm OCT: choroidal thickness maps in normal subjects and improved posterior segment visualization in cataract patients. Invest Ophthalmol Vis Sci 51:5260–5266
Fatt I, Weissman BA (1992) Physiology of the eye: an introduction to vegetative functions. Butterworth-Heinemann, St. Louis
Grosvenor TP (1989) Primary care optometry. Professional Press, New York
Hart WM (1992) Adler’s physiology of the eye, 9th edn. Mosby, St. Louis
Koretz JF (2000) Development and aging of human visual focusing mechanisms. In: Lakshminarayanan V (ed) Vision science and its applications, vol 35, Trends in optics and photonics series. Optical Society of America, Washington, DC, pp 246–258
Lakshminarayanan V, Enoch JM (2010) Biological waveguides, chapter 8. In: Bass M, DeCusatis C, Enoch JM, Lakshminarayanan V et al (eds) Handbook of optics, vol III. McGraw Hill, New York
Lakshminarayanan V, Nygaard RW (1992) Aliasing in the human visual system. Concepts Neurosci 3:201–212
Muller M, Strobel J (2007) Mechanism of accommodation of human eye – some new aspects. Klin Monbl Augenheilkd 224:653–658
Osterberg G (1935) Topography of the layers of the rods and cones in the human retina. Acta Ophthalmol Suppl 13:1–102
Oyster CW (1990) The human eye. Sinauer, Sunderland
Palmer SE (1999) Vision science. Photons to phenomenology. MIT Press, Cambridge, MA
Pang Y, Maino DM, Zhang G, Lu F (2006) Myopia: can its progression be controlled? Optom Vis Dev 37:75–79
Pierscionek B (2010) Gradient index optics in the eye, chapter 18. In: Bass M, De Cusatis C, Enoch JM, Lakshminarayanan V et al (eds) Handbook of optics, 3rd edn. McGraw Hill, New York
Remington LA (2005) Clinical anatomy of the visual system, 2nd edn. Butterwoth Heinemann, St. Louis
Rodieck RW (1998) The first steps in seeing. Sinauer, Sunderland
Rolls ET, Deco G (2004) The computational neuroscience of vision. Oxford University Press, Oxford
Roorda A, Williams DW (1999) The arrangement of the three cone classes in the living human eye. Nature 397:520–522
Simon JW, Calhoun JH (1998) A child’s eyes: a guide to pediatric primary care. Triad Publishing, Gainesville
Stark L, Sun F, Lakshminarayanan V, Wong J, Nguyen A, Muller E (1985) Presbyopia in the light of accommodation. Work reports on the 3rd international symposium on presbyopia, vol 2. Essilor International, Paris, pp 340–352
Toyoda J, Murakami M, Kaneko A, Saito T (1999) The retinal basis of vision. Elsevier, Amsterdam
Walls GL (1942) The vertebrate eye and its adaptive radiations. Cranbook Institute of Science, Bloomfield Hills
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this entry
Cite this entry
Garhart, C., Lakshminarayanan, V. (2016). Anatomy of the Eye. In: Chen, J., Cranton, W., Fihn, M. (eds) Handbook of Visual Display Technology. Springer, Cham. https://doi.org/10.1007/978-3-319-14346-0_4
Download citation
DOI: https://doi.org/10.1007/978-3-319-14346-0_4
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-14345-3
Online ISBN: 978-3-319-14346-0
eBook Packages: EngineeringReference Module Computer Science and Engineering