
Abstract
Sitosterolemia is a rare genetic lipid disorder, mainly characterized by the accumulation of dietary xenosterols in plasma and tissues. It is caused by inactivating mutations in either ABCG5 or ABCG8 subunits, a subfamily-G ATP-binding cassette (ABCG) transporters. ABCG5/G8 encodes a pair of ABC half transporters that form a heterodimer (G5G8). This heterodimeric ATP-binding cassette (ABC) sterol transporter, ABCG5/G8, is responsible for the hepatobiliary and transintestinal secretion of cholesterol and dietary plant sterols to the surface of hepatocytes and enterocytes, promoting the secretion of cholesterol and xenosterols into the bile and the intestinal lumen. In this way, ABCG5/G8 function in the reverse cholesterol transport pathway and mediate the efflux of cholesterol and xenosterols to high-density lipoprotein and bile salt micelles, respectively. Here, we review the biological characteristics and function of ABCG5/G8, and how the mutations of ABCG5/G8 can cause sitosterolemia, a loss-of-function disorder characterized by plant sterol accumulation and premature atherosclerosis, among other features.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
Sitosterolemia is a rare genetic recessive disease in which an individual is unable to excrete xenosterols.
In 1974, Bhattacharyya and Conner first described a new syndrome, a lipid storage disorder, in two sisters who presented with tendon and tuberous xanthomas with normal plasma cholesterol levels and elevated plasma levels of plant sterols (phytosterols), such as sitosterol, campesterol, and stigmasterol. The disease was named—sitosterolemia—after the most abundant dietary xenosterol, sitosterol. In fact, some think that perhaps a more appropriate name for the disease would be xenosterolemia (Maguire et al. 2001). Anyway, this discovery initiated fundamental studies on how dietary sterols traffic and are eliminated by the body (Williams et al. 2021).
It took another 26 years before the sitosterolemia locus was mapped to chromosome 2p21 and the discovery that the genetic locus whose dysfunction leads to sitosterolemia encodes two genes, abcg5 and abcg8, whose proteins (ABCG5 and ABCG8) function as obligate heterodimers (Berge et al. 2000; Lu et al. 2001).
ABCG5 and ABCG8 are expressed only in hepatocytes, gallbladder epithelium, and enterocytes and are responsible for excretion of sterols, with xenosterols preferred over cholesterol (Patel et al. 2018). In fact, the naming of the disease has led to a bias: a vast range of other sterols, not just sitosterol, are accumulating in the body (Gregg et al. 1986). According to some researchers, a better name should be xenosterolemia.
Several missense mutations on either genes are the causative gene defect that lead to loss of function of the ABCG5/G8 transporter, which is associated with lipid phenotypes (Miettinen 1980; Salen et al. 1985; Berge et al. 2000; Lee et al. 2001; Lu et al. 2001; Brown and Yu 2010; Williams et al. 2021). Subsequent case reports established the recessive genetics of the disease and greatly expanded its potential clinical presentation.
2 Sterol Transport
All living cells depend on their ability to transfer molecules such as nutrients, hormones, metabolites, and across their membranes. Cell membrane is the natural barrier for intracellular constituents and the checkpoint of molecules and signals from the extracellular milieu. Lipids are the primary component of mammalian cell membranes, with cholesterol being a key component. Cholesterol accounts for ∼40–50% of the total lipid content in the plasma membrane (Steck and Lange 2018). Cholesterol serves as the precursor molecule for steroid hormones that modulate gene regulation, for bile acids that enable for nutrient absorption, and for vitamin D which are vital for body health (Rezaei et al. 2023).
Translocation of cholesterol molecules on biological membranes plays an essential role in maintaining cholesterol homeostasis. However, relatively little is known about the mechanisms that control the sterol shuttling across lipid-bilayer membranes. Anyway, lipid-transport membrane proteins have been shown to be essential for the translocation of sterols and phospholipids to maintain lipid homeostasis, cellular functions, and the structural integrity of mosaic lipid bilayers (Abumrad et al. 2000; Sharom 2011; López-Marqués et al. 2015).
Cholesterol can be obtained by de novo cell biosynthesis or via dietary uptake in the intestine. In normal diets, the levels of cholesterol and non-cholesterol sterols from plants (xenosterols) or other dietary sources are usually equal. However, 50–60% of dietary cholesterol is absorbed, while xenosterols exhibit poor bioavailability, with <5% absorption (Salen et al. 1970). When more plant sterols are ingested, they compete with the bulk cholesterol for solubilization, thereby reducing dietary absorption of cholesterol and lowering plasma cholesterol.
Elimination of excess cholesterol is vital for life. Abnormal elevations in plasma cholesterol contribute to hyperlipidemia, a critical factor leading to cardiovascular diseases and other metabolic disorders (Salen et al. 1970). However, not all imported cholesterol is metabolized in the cells. In fact, few cells have this capacity. Therefore, elimination of the excess amount of cholesterol has to be cleared from cells and tissues via two metabolic pathways that are essential to maintain homeostasis: reverse cholesterol transport through sterol acceptors in the circulation or direct cholesterol excretion through biliary and intestinal secretion (Vrins et al. 2012; Ouimet et al. 2019). Xenosterols have efficient biliary elimination (Salen et al. 1970).
In more advanced life forms, functions such as nutrient intake and the exchange of compounds between cellular organelles or tissues often take place against concentration gradients across cellular membranes (Schumacher and Benndorf 2017). It is therefore not surprising that in simple life forms like bacteria, almost 10% of the entire genome is dedicated to proteins that are involved in transport processes in the form of membrane-bound or soluble proteins (Blattner et al. 1997). Transport processes against chemical gradients always require free energy which is derived from either by simultaneous use of an opposing electrochemical potential difference (secondary active transport) or a coupled enzymatic reaction exploiting the chemical energy of adenosine triphosphate (ATP) hydrolysis (primary active transport) (Blattner et al. 1997; Schumacher and Benndorf 2017). This ATP-driven transport, which act through the activity of energy-dependent unidirectional, membrane-bound, compound-efflux transporter proteins, comprise a large superfamily, the ABC (ATP-binding cassette) transporters.
3 ABC Transporters
The ABC transporter superfamily comprises one of the largest families of evolutionarily conserved membrane proteins and is ubiquitously expressed in all domains of life, from nearly all prokaryotes to virtually all types of eukaryotic cells (Higgins 1992; Dean et al. 2001a, b; Dean and Annilo 2005; Locher 2016; Bilsing et al. 2023). ABC transporters are most abundantly expressed in organs with high metabolic rates and in endothelial cells that isolate organs like the blood–brain barrier and blood–testis barrier (Schumacher and Benndorf 2017).
These transporters are involved in a broad range of cellular processes, therein actively transporting a wide range of different substrates across the plasma membrane (Schumacher and Benndorf 2017). ABC transporters use the energy from ATP hydrolysis to drive the passage or flipping of various moieties across the bilayer membrane, from small inorganic and organic molecules to larger organic compounds (Schumacher and Benndorf 2017), including both hydrophilic and hydrophobic molecules such as sugars, peptides, drugs, phospholipids, and sterols (Dean and Allikmets 1995; Linton and Higgins 1998; Dean et al. 2001a, b; Hwang et al. 2016; Plummer et al. 2021).
ABC transporters are divided into three subclasses, two groups of importers and one group of exporters, according to their functional and architectural characteristics (Schumacher and Benndorf 2017). ABC importers are predominantly found in prokaryotes where they manage the nutrient and ion intake (Ferreira and de Sá-Nogueira 2010; Gisin et al. 2010); they only sparsely occur in eukaryotes. The vast majority of ABC transporters expressed in eukaryotes are ABC exporters, promoting functions such as secreting dietary substances and metabolites or even transport signaling molecules (Schumacher and Benndorf 2017). Also, ABC transporters are major sterol exporters responsible for both cholesterol efflux from peripheral cells and the elimination of excess cholesterol and dietary sterols such as sitosterols (Borst and Elferink 2002; Xavier et al. 2019).
There are currently 49 different genes known to encode ABC transporters in humans, which categorize subfamilies of ABC transporter proteins based on sequence similarity, sequence divergence and structural arrangement (Plummer et al. 2021; Huang and Ecker 2023). Since several of those genes are alternatively spliced during transcription, each of those 49 genes not only encode one single protein, but instead often a multiple of different ABC protein variants (Dean et al. 2001a, b). The subfamilies are named ABCA–ABCG. Five distinct families (A, B, C, D, and G) display a wide array of substrate specificities and functionalities (Alam and Locher 2023). Of all ABC transporters that have been described so far, the three members ABCB1, ABCC1, and ABCG2 are less organ specific (Zhang et al. 2015).
The first human ABC transporter described, P-glycoprotein (ABCB1) (170 kDa) was discovered in 1976 by Juliano and Ling in Chinese hamster ovary (CHO) cells, selected for resistance to colchicine. These cells displayed pleiotropic cross-resistance to a wide range of amphiphilic drugs. Because the glycoprotein altered the membrane permeability (P), it was called P-glycoprotein (P-gp, encoded by the MDR-1/abcb1 gene) (Huang and Ecker 2023). ABCB1 is the first mammalian member of the large family of ABC transporters present in prokaryote (Davidson et al. 2008) and eukaryotes, from plants (Theodoulou 2000) to humans (Gottesman and Ambudkar 2001).
The division of ABC transportes in subfamilies is based mainly on similarity in gene structure, e.g., half vs. full transporters, and on sequence homology in the nucleotide-binding domains (NBDs) and transmembrane domains (TMD) (Schumacher and Benndorf 2017).
ABC transporters are organized as two symmetric halves that are expressed either (a) as separate subunits (half-transporters) that assemble as homodimers or heterodimers or (b) as monomers containing two nonidentical halves within a single polypeptide (full transporter) (Alam and Locher 2023). Each half comprises, at minimum, a NBD that is responsible for ATP binding and hydrolysis and a TMD that facilitates substrate export (away from the NBDs) or import (toward the NBDs) (Alam and Locher 2023). The human ABC transporters can be either full or half-transporters. In principle, full ABC transporter transcripts, such as members in the A- and C-subfamilies, comprise four domains within one polypeptide chain, namely 2 TMDs embedded in the lipid bilayer, and 2 NBDs facing the cytoplasmatic space (Plummer et al. 2021; Huang and Ecker 2023). The motifs are arranged as N-TMD-NBD-TMD-NBD-C, whereas the half ones have only one TMD and one NBD (Plummer et al. 2021). Hence, the half-transporters should form homodimers or heterodimers to perform their function (Huang and Ecker 2023). Among ABC transporter families, NBDs exhibit high sequence homology, with several canonical motifs. The transport function is generally believed to be driven by the NBD dimerization, in which ATP is bound and hydrolyzed. TMDs are structurally highly diverse, suggesting distinctive transport mechanisms for individual transporters (Ford and Beis 2019).
4 Main Diseases Related to ABC Transporters
Active in nearly all cells and tissues, ABC transporters play vital physiological roles ranging from lipid homeostasis to transport of diverse endogenous and exogenous compounds (Alam and Locher 2023). Several diseases result directly from dysfunction of these transporters, making them important targets for therapeutic intervention. Therefore, human ABC transporters hold tremendous biomedical and pharmacological relevance (Dean et al. 2001a, b; Borst and Elferink 2002; Leonard et al. 2003).
A major obstacle in cancer treatment is the development of cancer resistance to several structurally dissimilar cytotoxic substances (Huang and Ecker 2023). This phenomenon is termed as multidrug resistance (MDR), which renders the cancer cells ineffective in accumulating drugs, preventing their death. ABCB1, ABCC1, and ABCG2 were frequently observed with enhanced overexpression in multiple cancer types (Zhang et al. 2015). In fact, ABCB1 transporter is the most studied ABC regarding chemotherapy against cancer (Hwang et al. 2016; Seelig and Li-Blatter 2023). Subsequent to the discovery of P-gp, studies of cancer cells revealed other phenotypes, which showed multidrug resistance related characteristics. These multidrug resistance related proteins (MRPs) were later classified as the ABCC subfamily (Cole et al. 1992). Simultaneously, a novel half transporter member of the ABC superfamily was identified from a resistant breast cancer cell line (Doyle et al. 1998), hence named as breast cancer resistance protein (BCRP), encoded by the ABCG2 gene. Additionally, other members of ABC transporter were reported to export at least one anticancer agent. For instance, ABCA2, ABCC2, ABCC3, ABCC4, ABCC5, ABCC6, and ABCC11 (Hwang et al. 2016). Others, such as ABCB11, also known as Spgp (sister of P-glycoprotein) or BSEP (bile salt exporter protein), which is predominantly expressed in liver, has the capacity to confer resistance to cytotoxic substrates like taxol and vinblastine (Childs et al. 1995, 1998).
With at least 20 human ABC transporters being related to the transport of lipids or lipid-like compounds, it is not surprising that some of these transporters have been linked to the pathogenesis of atherosclerotic vascular diseases (Schumacher and Benndorf 2017). Moreover, ABC transporters have also been associated with vascular endothelial homeostasis and blood pressure regulation, as well as platelet production and aggregation (Schumacher and Benndorf 2017).
Dysfunction of ABCA1 can lead to Tangier’s disease (Alam and Locher 2023). ABCA2 dysfunction has been associated with intellectual and developmental deficiency, and also to amyloid homeostasis, thereby pointing to a potential role in Alzheimer’s disease (Alam and Locher 2023). Genetic variations in the ABCA3 gene, involved in phospholipid transport from the cytoplasm to the lumen of lamellar bodies, can cause pulmonary surfactant metabolism dysfunction 3, a severe respiratory disorder (Alam and Locher 2023).
5 ABCG5/G8 Transporter
ATP-binding cassette subfamily G (ABCG) sterol transporters maintain the homeostasis of endogenous and exogenous sterol. A substantial part of exogenous sterols are undigestible phytosterols, which can lead to complications when accumulated. ABCG5/G8 is the main functioning protein to remove ingested plant sterols providing protection from their toxic effects, although the structural features behind substrate binding in ABCG5/G8 remain relatively poorly resolved.
5.1 ABCG5/G8 Transporter Genes
The ABCG subfamily comprises five genes encoding half-transporters. Both of the two genes abcg5 and abcg8 are located on chromosome 2p21, adjacent to each other in a head-to-head fashion, on opposite strands, and are separated by only 374 base pairs (Berge et al. 2000).
Due to their proximity and opposite orientation, these two genes are regulated by a bidirectional intergenic promotor, which contains binding sites for hepatocyte nuclear factor 4α, GATA 4/6 (Sumi et al. 2007), and a liver receptor homolog 1 (Freeman et al. 2004). In addition, agonists for either liver X receptor or farnesoid X receptor regulate ABCG5/G8 mRNA levels (Repa et al. 2002).
Each ABC transporter (ABCG5/G8) comprises two nucleotide-binding composite sites, where the Walker A motif of one NBD is paired with the ABC signature motif of the other NBD. Therefore, one of the ATP-binding sites presents a degenerate motif, while the other presents a conserved motif, which is the only one able to support ATP hydrolysis.
5.2 Characteristics of the ABCG5/G8 Transporter Structure
The members of the G-subfamily are half transporters with only one NBD and one TMD. The ABCG transporters are also characteristic. They are the only human ABC transporters with their inverted domain topology that contain an N-terminal NBD followed by a C-terminal TMD (Alam and Locher 2023).
G subfamily members must form homo- or heterodimers for functionality (Dean et al. 2001a, b). Although two transportes, ABCG1 and ABCG4, have been proposed to also function as heterodimers, most studies support the notion that both are homodimers (Cserepes et al. 2004; Hegyi and Homolya 2016). In contrast, ABCG5 and ABCG8 form obligate heterodimers (Lee et al. 2001; Wang et al. 2006; Brown and Yu 2010).
Each gene ABCG5 and ABCG8 encodes a “half-transporter” protein that is nonfunctional in the monomeric state (Brown and Yu 2010). However, assembly of an ABCG5/G8 heterodimer, driven by the adipocyte-derived hormone leptin, leads to the formation of the fully functional transporter ABCG5/G8 (Brown and Yu 2010).
Structural information of the ABCG5/G8 heterodimer was revealed by X-ray crystallography, establishing a new molecular framework toward a mechanistic understanding of ABC sterol transporters (Graf et al. 2002, 2003). This structure contained an asymmetric unit with two heterodimers that interact through their TMDs with NBDs at opposite sides of the membrane. Without bound nucleotide or lipid substrate inside the transporter, the ABCG5/G8 structure exhibits an inward-facing conformation (Plummer et al. 2021). Three helices from both ABCG5 and ABCG8, form a three-helix bundle, which bridges the TMD and NBD.
In addition, a sterol-binding site was postulated at the membrane-transporter interface based on the crystal structure of ABCG5/G8 (Farhat et al. 2022), solving the crystal structure of ABCG5/G8 in complex with cholesterol. The structure shows that an orthogonal cholesterol molecule fitting horizontally in front of A540, a conserved ABCG5 residue at this orthogonal sterol-binding site.
5.3 Observations on ABCG5/G8 Mutations
Mutations present in some sitosterolemia patients impair heterodimer trafficking (Graf et al. 2004), suggesting that these mutations disrupt ABCG5/G8 cellular localization rather than reducing ABCG5/G8 transport activity (Plummer et al. 2021). However, this seems not to be a rule. The Ala540Phe mutant in ABCG5, a residue that putatively binds cholesterol, resulted in reduced biliary cholesterol transport (Lee et al. 2016).
On the triple-helical bundle or the transmembrane polar relay, several residues have been shown to bear disease-causing missense mutations from patients with sitosterolemia or other lipid metabolic disorders. Notably, several disease-causing mutations are clustered in the membrane-spanning region or at the NBD–TMD interface. This suggests the roles of these structural motifs in regulating the ABCG5/G8 function.
ABCG5/G8 has also been studied through overexpression of either wild-type or mutant ABCG5/G8 in abcg5/abcg8 KO mice (Plummer et al. 2021). Mice lacking abcg5, abcg8, or both show increased plasma levels of sterols and reduction of sterol secretion into the bile. In contrast, overexpression of ABCG5/G8 showed the opposite effect, with reduced sterol absorption and increased biliary sterol levels (Yu et al. 2002a, b). These effects are dependent on ABCG5/G8 expression levels. Moreover, using mdr2 KO mice, a lack of secretion was observed, suggesting that the function of ABCG5/G8 is dependent on a functional ABCB4 transporter (Plummer et al. 2021).
Homodimers of either ABCG5 or ABCG8 are likely nonfunctional, evident from low ATPase activity relative to the native heterodimer (Wang et al. 2006), impaired trafficking (Graf et al. 2003), and low biliary cholesterol transfer in KO mice (Graf et al. 2003; Zhang et al. 2006). Surprisingly, abcg8 KO in mice results in continued expression of ABCG5 at the apical membrane and secretion of sitosterol into the bile, suggesting that the ABCG5 homodimer may be functional or that an alternative pathway of sterol secretion into the bile exists (Klett et al. 2004).
5.4 Function and Regulation of ABCG5/G8
Physiologically, ABCG5/G8 plays an essential role in controlling sterol homeostasis in our bodies.
The ABCG5/G8 transporter is predominantly expressed on the apical surface of hepatocytes along the canalicular membranes of the bile ducts in the liver and on the brush-border membranes of enterocytes in the small intestines (Patel et al. 1998; Berge et al. 2000; Graf et al. 2002, 2003; Yu et al. 2002a, b; Hirata et al. 2009; Brown and Yu 2010; Zein et al. 2019).
ABCG5/G8 is the primary transporter and sterol-efflux pump that selectively exports excess cholesterol, noncholesterol sterols, and dietary plant sterols from hepatocytes into bile canaliculi and in the intestine back to the intestinal lumen (Lee et al. 2001, Wang et al. 2006; Rezaei et al. 2023), by translocating sterols within the plasma membrane and in endosomes (Sano et al. 2014; Pandzic et al. 2017; Xavier et al. 2020). Expression level of ABCG5 and ABCG8 is further modulated by bile acid levels in both the liver (Dean et al. 2001a, b) and intestine (Kamisako et al. 2007).
One proposed mechanistic mode for the sterol transfer to bile acid suggests that ABCG5/G8 translocates sterol across the bilayer membrane, functioning as a liftase (Small 2003; Lee et al. 2016). However, there is little experimental evidence to support this hypothesis.
Another model suggests that ABCG5/G8 functions only as a sterol floppase, increasing the sterol concentration in the outer leaflet and allowing for extraction of sterol by the bile salt micelle (Kosters et al. 2006). This hypothesis is supported by the observation that the function of ABCG5/G8 depends on ABCB4, a phosphatidylcholine transporter, and sterol is potentially extracted from the outer leaflet via sterol-phospholipid vesicles (Crawford et al. 1997). Multidrug-resistant protein 3 (MDR3), also known as ABCB4, is a phospholipid translocase embedded in the canalicular membrane. Although it actively flips inner leaflet phospholipids and sterols to the outer leaflet, there is evidence supporting its subsequent role in substrate secretion toward the bile. Furthermore, MDR3 is found to be essential for the proper function of ABCG5/G8, which believed to be caused by its involvement in the formation of mixed micelles.
Finally, ABCG5/G8 may directly transfer sterols to bile salt micelles following sterol flipping across the plasma membrane (Plummer et al. 2021). Spontaneous cholesterol flipping between leaflets is a common event in the plasma membrane, capable of undertaking intermediate horizontal orientation within the membrane core. It is thus possible that ABCG5/G8 catalyzes cholesterol flipping from inner to outer leaflets, peripherally through its exterior surface.
In addition, ABCG5/G8 exhibits cholesterol efflux activity in the presence of bile acid micelles (Vrins et al. 2007). Mixed micelles are very charged, small aggregates of phospholipids, cholesterol and bile salts, and these micelles form the basis of currently known ABCG5/G8 acceptor particles. Accordingly, it has been suggested that mixed micelles of bile acids dock onto ABCG5/G8 to induce conformational changes and stimulate ATPase for the transport of cholesterol (Johnson et al. 2010). Acceptor particles will then intake the exported lipids from ABCG5/G8.
Differently from homodimeric ABCG1, the heterodimeric ABCG5/G8 carries out selective sterol excretion. ABCG5/G8 is unique in its capability of preferential efflux for dietary plant sterols over cholesterol, preventing the abnormal accumulation of plant sterols in human body (Berge et al. 2000; Lee et al. 2001; Graf et al. 2003; Yang et al. 2004; Schumacher and Benndorf 2017). Intestinal ABCG5/G8 acts as a first-pass gate, pumping xenosterols back into the intestinal lumen, whereas liver ABCG5/G8 pumps xenosterols into the bile (Patel et al. 2018). There are numerous reports that ABCG5/G8 exhibits a preference for the transport of noncholesterol sterols over cholesterol (Yu et al. 2004; Plummer et al. 2021). The preference for noncholesterol sterols in either intestine- or liver-localized abcg5/abcg8 KO mice was slightly reduced compared to whole-animal abcg5/abcg8 KO models, suggesting that expression of ABCG5/G8 in both liver and intestine is required for full selectivity (Wang et al. 2015). The mechanisms that govern such substrate selectivity within this protein subfamily, however, remain elusive.
6 Sitosterolemia
This very rare autosomal recessive disorder is characterized by drastically elevated plasma and tissue levels of plant sterols (Schumacher and Benndorf 2017). As stated before, missense mutations in both genes, ABCG5 and ABCG8, are related to sitosterolemia (Berge et al. 2000; Hubacek et al. 2001; Lu et al. 2001). ABCG5/G8 exhibits substrate specificity, particularly for plant sterols (e.g., sitosterol and campesterol) over cholesterol (Matsumura et al. 2007; Michaki et al. 2012). In fact, sitosterolemia patients exhibit increased absorption of numerous plant xenosterols, not just sitosterol (Morita and Imanaka 2012).
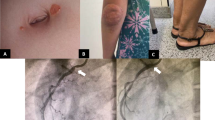
The clinical presentation may include elevated low-density lipoprotein (LDL) cholesterol and tuberous tendon xanthomas. Hypercholesterolemia-induced premature atherosclerosis was particularly observed to affect male patients at a young age, leading to CVD-like angina pectoris, myocardial infarctions, and sudden cardiac death (Kwiterovich et al. 1981; Brown and Yu 2010). Hematologic manifestations (hemolytic anemia, macrothrombocytopenia, splenomegaly, and bleeding disorders) can result from the accumulation of plant sterols in platelet membranes, producing hypertrophic and hyperplasic dysfunctional platelets. Adrenal dysfunction, arthritis, elevated liver function tests, and cirrhosis (in rare cases hepatic failure) are other features (Shulman et al. 1976; Miettinen 1980; Kwiterovich et al. 1981; Lin et al. 1983; Salen et al. 1985; Beaty et al. 1986; Nguyen et al. 1990; Bhattacharyya et al. 1991; Rees et al. 2005; Mushtaq et al. 2007; Wang et al. 2014; Bazerbachi et al. 2016). Clinical studies in individuals with sitosterolemia revealed reductions in cholesterol synthesis, biliary cholesterol secretion, plasma clearance, and fecal elimination of neutral sterols (Salen et al. 1989; Nguyen et al. 1990; Bhattacharyya et al. 1991; Cobb et al. 1997). Due to similar clinical presentations, sitosterolemia has been, in many cases, inaccurately diagnosed as familial hypercholesterolemia or idiopathic liver cirrhosis.
The ABCG5/G8 loss of function in animal models, as well as in humans, shows that accumulation of xenosterols leads to dramatic phenotypes, such as macrothrombocytopenia and platelet dysfunction, liver disease, appearance of gallstones, elevation of low-density lipoprotein cholesterol levels and cholesterol accumulation with xanthoma formation and atherosclerosis (Grass et al. 1995; Yu et al. 2002a, b, 2005; Kajinami et al. 2004; Acalovschi et al. 2006; Buch et al. 2007; Wang et al. 2007; Chen et al. 2008; Kuo et al. 2008; Katsika et al. 2010; Patel and Salen 2010; Srivastava et al. 2010; Renner et al. 2013; Von Kampen et al. 2013; Jiang et al. 2014). In mouse models (but not humans), infertility, immune dysfunction, and cardiomyopathy have been reported (McDaniel et al. 2013; Solca et al. 2013; Wilson et al. 2013). This begs the question whether a lifetime of low-level exposure to dietary bioactive xenosterols, whose levels of entry and retention may be altered by polymorphisms in ABCG5 and ABCG8, may have biological consequences.
Despite the absorptive phenotype and metabolism of phytosterols to bile acids, the clinical management of these patients with low sterol diets and bile acid-binding resins resulted in modest and inconsistent reductions in plasma phytosterols (Lin et al. 1983; Nguyen et al. 1991). Treatment of sitosterolemia involves a diet that is low in plant lipids and the administration of the drug ezetimibe, which acts as an inhibitor of Niemann-Pick C1-like protein 1 (NPC1L1), that functions in intestinal sterol absorption Morita et al. 2007).
In conclusion, although sitosterolemia is a rare genetic disease, this disorder should be considered in the differential diagnosis, due to its early clinical manifestations and relatively difficult treatment.
Abbreviations
- ABC:
-
ATP-binding cassette
- ABCB1:
-
ATP-binding cassette subfamily B1
- ABCG5:
-
ATP-binding cassette subfamily G5
- ABCG8:
-
ATP-binding cassette subfamily G8
- ATP:
-
Adenosine triphosphate
- BSEP:
-
Bile salt exporter protein
- KO mice:
-
Knock out mice
- MDR:
-
Multidrug resistance
- NBD:
-
Nucleotide-binding domain
- P-gp:
-
P-glycoprotein
- TMD:
-
Transmembrane domain
References
Abumrad NA, Sfeir Z, Connelly MA et al (2000) Lipid transporters: membrane transport systems for cholesterol and fatty acids. Curr Opin Clin Nutri Metab Care 3:255–262
Acalovschi M, Ciocan A, Mostean O et al (2006) Are plasma lipid levels related to ABCG5/ABCG8 polymorphisms? A preliminary study in siblings with gallstones. Eur J Intern Med 17:490–494
Alam A, Locher KP (2023) Structure and mechanism of human ABC transporters. Annu Rev Biophys 52:275–300
Bazerbachi F, Conboy EE, Mounajjed T et al (2016) Cryptogenic cirrhosis and Sitosterolemia: a treatable disease if identified but fatal if missed. Ann Hepatol 16:970–978
Beaty TH, Kwiterovich PO, Khoury MJ et al (1986) Genetic analysis of plasma sitosterol, apoprotein B, and lipoproteins in a large Amish pedigree with sitosterolemia. Am J Hum Genet 38:492–504
Berge KE, Tian H, Graf GA et al (2000) Accumulation of dietary cholesterol in sitosterolemia caused by mutations in adjacent ABC transporters. Science 290:1771–1775
Bhattacharyya AK, Connor WE (1974) Beta-sitosterolemia and xanthomatosis. A newly described lipid storage disease in two sisters. J Clin Invest 53:1033–1043
Bhattacharyya AK, Connor WE, Lin DS et al (1991) Sluggish sitosterol turnover and hepatic failure to excrete sitosterol into bile cause expansion of body pool of sitosterol in patients with sitosterolemia and xanthomatosis. Arterioscler Thromb AJ Vasc Biol/Am Heart Assoc 11:1287–1294
Bilsing FL, Anlauf MT, Hachani E et al (2023) ABC transporters in bacterial nanomachineries. Int J Mol Sci 24:6227
Blattner FR, Plunkett G, Bloch CA et al (1997) The complete genome sequence of Escherichia coli K-12. Science 277:1453–1462
Borst P, Elferink RO (2002) Mammalian ABC transporters in health and disease. Annu Rev Biochem 71:537–592
Brown JM, Yu L (2010) Protein mediators of sterol transport across intestinal brush border membrane. Subcell Biochem 51:337–380
Buch S, Schafmayer C, Völzke H et al (2007) A genome-wide association scan identifies the hepatic cholesterol transporter ABCG8 as a susceptibility factor for human gallstone disease. Nat Genet 39:995–999
Chen ZC, Shin SJ, Kuo KK et al (2008) Significant association of ABCG8:D19H gene polymorphism with hypercholesterolemia and insulin resistance. J Hum Genet 53:757–763
Childs S, Yeh RL, Georges E et al (1995) Identification of a sister gene to P-glycoprotein. Cancer Res 55:2029–2034
Childs S, Yeh RL, Hui D et al (1998) Taxol resistance mediated by transfection of the liver-specific sister gene of P-glycoprotein. Cancer Res 58:4160–4167
Cobb MM, Salen G, Tint GS (1997) Comparative effect of dietary sitosterol on plasma sterols and cholesterol and bile acid synthesis in a sitosterolemic homozygote and heterozygote subject. J Am Coll Nutr 16:605–613
Cole S, Bhardwaj G, Gerlach J et al (1992) Overexpression of a transporter gene in a multidrug-resistant human lung cancer cell line. Science 258:1650–1654
Crawford AR, Smith AJ, Hatch VC et al (1997) Hepatic secretion of phospholipid vesicles in the mouse critically depends on mdr2 or MDR3 P-glycoprotein expression. Visualization by electron microscopy. J Clin Investig 100:2562–2567
Cserepes J, Szentpetery Z, Seres L et al (2004) Functional expression and characterization of the human ABCG1 and ABCG4 proteins: indications for heterodimerization. Biochem Biophys Res Commun 320:860–867
Davidson AL, Dassa E, Orelle C, Chen J (2008) Structure, function, and evolution of bacterial ATP-binding cassette systems. Microbiol Mol Biol Rev 72:317–364
Dean M, Allikmets R (1995) Evolution of ATP-binding cassette transporter genes. Curr Opin Genet Dev 5:779–785
Dean M, Annilo T (2005) Evolution of the ATP-binding cassette (ABC) transporter superfamily in vertebrates. Annu Rev Genom Hum Genet 6:123–142
Dean M, Hamon Y, Chimini G (2001a) The human ATP-binding cassette (ABC) transporter superfamily. J Lipid Res 42:1007–1017
Dean M, Rzhetsky A, Allikmets R (2001b) The human ATP-binding cassette (ABC) transporter superfamily. Genome Res 11:1156–1166
Doyle LA, Yang W, Abruzzo LV et al (1998) A multidrug resistance transporter from human MCF-7 breast cancer cells. Proc Natl Acad Sci U S A 95:15665–15670
Farhat D, Rezaei F, Ristovski M et al (2022) Structural analysis of cholesterol binding and sterol selectivity by ABCG5/G8. J Mol Biolo 434:167795
Ferreira MJ, de Sá-Nogueira I (2010) A multitask ATPase serving different ABC-type sugar importers in Bacillus subtilis. J Bacteriol 192:5312–5318
Ford RC, Beis K (2019) Learning the ABCs one at a time: structure and mechanism of ABC transporters. Biochem Soc Trans 47:23–36
Freeman LA, Kennedy A, Wu J et al (2004) The orphan nuclear receptor LRH-1 activates the ABCG5/ABCG8 intergenic promoter. J Lipid Res 45:1197–1206
Gisin J, Müller A, Pfänder Y et al (2010) A Rhodobacter capsulatus member of a universal permease family imports molybdate and other oxyanions. J Bacteriol 192:5943–5952
Gottesman MM, Ambudkar SV (2001) Overview: ABC transporters and human disease. J Bioenerg Biomembr 33:453–458
Graf GA, Li W-P, Gerard RD et al (2002) Coexpression of ATP-binding cassette proteins ABCG5 and ABCG8 permits their transport to the apical surface. J Clin Investig 110:659–669
Graf GA, Yu L, Li W et al (2003) ABCG5 and ABCG8 are obligate heterodimers for protein trafficking and biliary cholesterol excretion. J Biol Chem 278:48275–48282
Graf GA, Cohen JC, Hobbs HH (2004) Missense mutations in ABCG5 and ABCG8 disrupt heterodimerization and trafficking. J Biol Chem 279:24881–24888
Grass DS, Saini U, Felkner RH et al (1995) Transgenic mice expressing both human apolipoprotein B and human CETP have a lipoprotein cholesterol distribution similar to that of normolipidemic humans. J Lipid Res 36:1082–1091
Gregg RE, Connor WE, Lin DS et al (1986) Abnormal metabolism of shellfish sterols in a patient with sitosterolemia and xanthomatosis. J Clin Inves 77:1864–1872
Hegyi Z, Homolya L (2016) Functional cooperativity between ABCG4 and ABCG1 isoforms. PLoS One 11:e0156516
Higgins CF (1992) ABC transporters: from microorganisms to man. Annu Rev Cell Biol 8:67–113
Hirata T, Okabe M, Kobayashi A et al (2009) Molecular mechanisms of subcellular localization of ABCG5 and ABCG8. Biosci Biotechnol Biochem 73:619–626
Huang J, Ecker GF (2023) A structure-based view on ABC-transporter linked to multidrug resistance. Molecules 28:495
Hubacek JA, Berge KE, Cohen JC et al (2001) Mutations in ATP-cassette binding proteins G5 (ABCG5) and G8 (ABCG8) causing sitosterolemia. Hum Mutat 18:359–360
Hwang JU, Song WY, Hong D et al (2016) Plant ABC transporters enable many unique aspects of a terrestrial plant’s lifestyle. Mol Plant 9:338–355
Jiang ZY, Cai Q, Chen EZ (2014) Association of three common single nucleotide polymorphisms of ATP binding cassette G8 gene with gallstone disease: a meta-analysis. PLoS One 9:e87200
Johnson BJ, Lee JY, Pickert A et al (2010) Bile acids stimulate ATP hydrolysis in the purified cholesterol transporter ABCG5/G8. Biochemistry 49:3403–3411
Juliano R, Ling V (1976) A surface glycoprotein modulating drug permeability in Chinese hamster ovary cell mutants. Biochim Biophys Acta Biomembr 455:152–162
Kajinami K, Brousseau ME, Ordovas JM et al (2004) Interactions between common genetic polymorphisms in ABCG5/G8 and CYP7A1 on LDL cholesterol-lowering response to atorvastatin. Atherosclerosis 175:287–293
Kamisako T, Ogawa H, Yamamoto K (2007) Effect of cholesterol, cholic acid and cholestyramine administration on the intestinal mRNA expressions related to cholesterol and bile acid metabolism in the rat. J Gastroenterol Hepatol 22:1832–1837
Katsika D, Magnusson P, Krawczyk M et al (2010) Gallstone disease in Swedish twins: risk is associated with ABCG8 D19H genotype. J Intern Med 268:279–285
Klett EL, Lu K, Kosters A et al (2004) A mouse model of sitosterolemia: absence of Abcg8/sterolin-2 results in failure to secrete biliary cholesterol. BMC Med 2:5
Kosters A, Kunne C, Looije N et al (2006) The mechanism of ABCG5/ABCG8 in biliary cholesterol secretion in mice. J Lipid Res 47:1959–1956
Kuo K-K, Shin S-J, Chen Z-C (2008) Significant association of ABCG5 604Q and ABCG8 D19H polymorphisms with gallstone disease. Br J Surg 95:1005–1011
Kwiterovich PO, Bachorik PS, Smith HH et al (1981) Hyperapobetalipoproteinaemia in two families with xanthomas and phytosterolaemia. Lancet 1:466–469
Lee M-H, Lu K, Hazard S et al (2001) Identification of a gene, ABCG5, important in the regulation of dietary cholesterol absorption. Nat Genet 27:79–83
Lee J-Y, Kinch LN, Borek DM et al (2016) Crystal structure of the human sterol transporter ABCG5/ABCG8. Nature 533:561–564
Leonard GD, Fojo T, Bates SE (2003) The role of ABC transporters in clinical practice. Oncologist 8:411–424
Lin HJ, Wang C, Salen G et al (1983) Sitosterol and cholesterol metabolism in a patient with coexisting phytosterolemia and cholestanolemia. Metab Clin Exp 32:126–133
Linton KJ, Higgins CF (1998) The Escherichia coli ATP-binding cassette (ABC) proteins. Mol Microbiol 28:5–13
Locher KP (2016) Mechanistic diversity in ATP-binding cassette (ABC) transporters. Nat Struct Mol Biol 23:487–493
López-Marqués RL, Poulsen LR, Bailly A et al (2015) Structure and mechanism of ATP-dependent phospholipid transporters. Biochim Biophys Acta 1850:461–475
Lu K, Lee MH, Hazard S et al (2001) Two genes that map to the STSL locus cause sitosterolemia: genomic structure and spectrum of mutations involving sterolin-1 and sterolin-2, encoded by ABCG5 and ABCG8, respectively. Am J Hum Genet 69:278–290
Maguire A, Hellier K, Hammans S et al (2001) X-linked cerebellar ataxia and sideroblastic anaemia associated with a missense mutation in the ABC7 gene predicting V411L. Br J Haematol 115:910–917
Matsumura Y, Sakai H, Sasaki M et al (2007) ABCA3-mediated choline-phospholipids uptake into intracellular vesicles in A549 cells. FEBS Lett 581:3139–3144
McDaniel AL, Alger HM, Sawyer JK et al (2013) Phytosterol feeding causes toxicity in ABCG5/G8 knockout mice. Am J Pathol 182:1131–1138
Michaki V, Guix FX, Vennekens K et al (2012) Down-regulation of the ATPbinding cassette transporter 2 (Abca2) reduces amyloid-beta production by altering Nicastrin maturation and intracellular localization. J Biol Chem 287:1100–1111
Miettinen TA (1980) Phytosterolaemia, xanthomatosis and premature atherosclerotic arterial disease: a case with high plant sterol absorption, impaired sterol elimination and low cholesterol synthesis. Eur J Clin Investig 10:27–35
Morita M, Imanaka T (2012) Peroxisomal ABC transporters: structure, function and role in disease. Biochim Biophys Acta 1822:1387–1396
Morita SY, Kobayashi A, Takanezawa Y et al (2007) Bile salt–dependent efflux of cellular phospholipids mediated by ATP binding cassette protein B4. Hepatology 46:188–199
Mushtaq T, Wales JK, Wright NP (2007) Adrenal insufficiency in phytosterolaemia. Eur J Endocrinol 157(Suppl 1):S61–S65
Nguyen LB, Salen G, Shefer S et al (1990) Decreased cholesterol biosynthesis in sitosterolemia with xanthomatosis: diminished mononuclear leukocyte 3-hydroxy-3-methylglutaryl coenzyme A reductase activity and enzyme protein associated with increased low-density lipoprotein receptor function. Metab Clin Exp 39:436–443
Nguyen LB, Cobb M, Shefer S et al (1991) Regulation of cholesterol biosynthesis in sitosterolemia: effects of lovastatin, cholestyramine, and dietary sterol restriction. J Lipid Res 32:1941–1948
Ouimet M, Barrett TJ, Fisher EA (2019) HDL and reverse cholesterol transport: basic mechanisms and their roles in vascular health and disease. Circ Res 124:1505–1518
Pandzic E, Gelissen IC, Whan R et al (2017) The ATP binding cassette transporter, ABCG1, localizes to cortical actin filaments. Sci Rep 7:42025
Patel SB, Salen G (2010) Sitosterolemia: xenophobia for the body. In: Vissers MN, Kastelein JJP, Stroes ES (eds) Evidence-based management of lipid disorders. TFM Publishing Ltd., Shrewsbury, pp 217–230
Patel SB, Honda A, Salen G (1998) Sitosterolemia: exclusion of genes involved in reduced cholesterol biosynthesis. J Lipid Res 39:1055–1061
Patel SB, Graf GA, Temel RE (2018) ABCG5 and ABCG8: more than a defense against xenosterols. J Lipid Res 59:1103–1113
Plummer AM, Alan T, Lial M (2021) The ABCs of sterol transport. Annu Rev Physiol 83:153–181
Rees DC, Iolascon A, Carella M et al (2005) Stomatocytic haemolysis and macrothrombocytopenia (Mediterranean stomatocytosis/macrothrombocytopenia) is the haematological presentation of phytosterolaemia. Br J Haematol 130:297–309
Renner O, Lutjohann D, Richter D et al (2013) Role of the ABCG8 19H risk allele in cholesterol absorption and gallstone disease. BMC Gastroenterol 13:30
Repa JJ, Berge KE, Pomajzl C et al (2002) Regulation of ATPbinding cassette sterol transporters ABCG5 and ABCG8 by the liver X receptors α and β. J Biol Chem 277:18793–18800
Rezaei F, Farhat D, Gursu G et al (2023) Snapshots of ABCG1 and ABCG5/G8: a Sterol’s journey to cross the cellular membranes. Int J Mol Sci 24:484
Salen G, Ahrens EH Jr, Grundy SM (1970) Metabolism of beta sitosterol in man. J Clin Invest 49:952–967
Salen G, Horak I, Rothkopf M et al (1985) Lethal atherosclerosis associated with abnormal plasma and tissue sterol composition in sitosterolemia with xanthomatosis. J Lipid Res 26:1126–1133
Salen G, Shore V, Tint GS et al (1989) Increased sitosterol absorption, decreased removal, and expanded body pools compensate for reduced cholesterol synthesis in sitosterolemia with xanthomatosis. J Lipid Res 30:1319–1330
Sano O, Ito S, Kato R et al (2014) ABCA1, ABCG1, and ABCG4 are distributed to distinct membrane meso-domains and disturb detergent-resistant domains on the plasma membrane. PLoS One 9:e109886
Schumacher T, Benndorf RA (2017) ABC transport proteins in cardiovascular disease—a brief summary. Molecules 22:589
Seelig A, Li-Blatter X (2023) P-glycoprotein (ABCB1)—weak dipolar interactions provide the key to understanding allocrite recognition, binding, and transport. Cancer Drug Resist 6:1–29
Sharom FJ (2011) Flipping and flopping–lipids on the move. IUBMB Life 63:736–746
Shulman RS, Bhattacharyya AK, Connor WE et al (1976) Beta-sitosterolemia and xanthomatosis. N Engl J Med 294:482–483
Small DM (2003) Role of ABC transporters in secretion of cholesterol from liver into bile. PNAS 100:4–6
Solca C, Tint GS, Patel SB (2013) Dietary xenosterols lead to infertility and loss of abdominal adipose tissue in sterolindeficient mice. J Lipid Res 54:397–409
Srivastava A, Srivastava A, Srivastava K et al (2010) Role of ABCG8 D19H (rs11887534) variant in gallstone susceptibility in northern India. J Gastroenterol Hepatol 25:1758–1762
Steck TL, Lange Y (2018) Transverse distribution of plasma membrane bilayer cholesterol: picking sides. Traffic 19:750–760
Sumi K, Tanaka T, Uchida A et al (2007) Cooperative interaction between hepatocyte nuclear factor 4α and GATA transcription factors regulates ATP-binding cassette sterol transporters ABCG5 and ABCG8. Mol Cell Biol 27:4248–4260
Theodoulou FL (2000) Plant ABC transporters. Biochim Biophys Acta Biomembr 1465:79–103
Von Kampen O, Buch S, Nothnagel M et al (2013) Genetic and functional identification of the likely causative variant for cholesterol gallstone disease at the ABCG5/8 lithogenic locus. Hepatology 57:2407–2417
Vrins C, Vink E, Vandenberghe KE et al (2007) The sterol transporting heterodimer ABCG5/ABCG8 requires bile salts to mediate cholesterol efflux. FEBS Lett 581:4616–4620
Vrins CL, Ottenhoff R, van den Oever K et al (2012) Trans-intestinal cholesterol efflux is not mediated through high density lipoprotein. J Lipid Res 53:2017–2023
Wang Z, Stalcup LD, Harvey BJ et al (2006) Purification and ATP hydrolysis of the putative cholesterol transporters ABCG5 and ABCG8. Biochemistry 45:9929–9939
Wang Y, Jiang ZY, Fei J et al (2007) ATP binding cassette G8 T400K polymorphism may affect the risk of gallstone disease among Chinese males. Clin Chim Acta 384:80–85
Wang Z, Cao L, Su Y et al (2014) Specific macrothrombocytopenia/hemolytic anemia associated with sitosterolemia. Am J Hematol 89:320–324
Wang J, Mitsche MA, Lutjohann D et al (2015) Relative roles of ABCG5/ABCG8 in liver and intestine. J Lipid Res 56:319–330
Williams K, Segard A, Graf GA (2021) Sitosterolemia: twenty years of discovery of the function of ABCG5 ABCG8. Int J Mol Sci 22:2641
Wilson DW, Oslund KL, Lyons B et al (2013) Inflammatory dilated cardiomyopathy in Abcg5-deficient mice. Toxicol Pathol 41:880–892
Xavier BM, Jennings WJ, Zein AA et al (2019) Structural snapshot of the cholesterol-transport ATP-binding cassette proteins. Biochem Cell Biol 97:224–233
Xavier BM, Zein AA, Venes A et al (2020) Transmembrane polar relay drives the allosteric regulation for ABCG5/G8 sterol transporter. Int J Mol Sci 21:8747
Yang C, Yu L, Li W et al (2004) Disruption of cholesterol homeostasis by plant sterols. J Clin Invest 114:813–822
Yu L, Hammer RE, Li-Hawkins J (2002a) Disruption of Abcg5 and Abcg8 in mice reveals their crucial role in biliary cholesterol secretion. Proc Natl Acad Sci U S A 99:16237–16242
Yu L, Li-Hawkins J, Hammer RE et al (2002b) Overexpression of ABCG5 and ABCG8 promotes biliary cholesterol secretion and reduces fractional absorption of dietary cholesterol. J Clin Investig 110:671–680
Yu L, von Bergmann K, Lutjohann D et al (2004) Selective sterol accumulation in ABCG5/ABCG8-deficient mice. J Lipid Res 45:301–307
Yu L, Gupta S, Xu F et al (2005) Expression of ABCG5 and ABCG8 is required for regulation of biliary cholesterol secretion. J Biol Chem 280:8742–8747
Zein AA, Kaur R, Hussein TOK et al (2019) ABCG5/G8: a structural view to pathophysiology of the hepatobiliary cholesterol secretion. Biochem Soc Trans 47:1259–1268
Zhang DW, Graf GA, Gerard RD et al (2006) Functional asymmetry of nucleotidebinding domains in ABCG5 and ABCG8. J Biol Chem 281:4507–4516
Zhang Y-K, Wang Y-J, Gupta P et al (2015) Multidrug resistance proteins (MRPs) and cancer therapy. AAPS J 17:802–812
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2024 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Bydlowski, S.P., Levy, D. (2024). Association of ABCG5 and ABCG8 Transporters with Sitosterolemia. In: Lizard, G. (eds) Implication of Oxysterols and Phytosterols in Aging and Human Diseases. Advances in Experimental Medicine and Biology, vol 1440. Springer, Cham. https://doi.org/10.1007/978-3-031-43883-7_2
Download citation
DOI: https://doi.org/10.1007/978-3-031-43883-7_2
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-43882-0
Online ISBN: 978-3-031-43883-7
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)