
Abstract
Primary headaches are highly prevalent conditions that may burden individuals significantly. Their biological underpinnings are still not fully unveiled, yet the autonomic nervous system (ANS) arguably plays a significant role in the development and progression of these disorders.
In this chapter, we will review the complex role of the ANS in migraine and cluster headache in the attack generation, clinical manifestations, interictal conditions, and the potential treatment effects of its iatrogenic modulation. Finally, we will discuss the evolutionary significance of primary headaches to provide a comprehensive bio-behavioural view to explain pain meaning and the distinctive autonomic-behavioural responses observed during migraine and cluster headache attacks.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
- Migraine
- Cluster headache
- Trigeminal autonomic reflex (TAR)
- Trigeminal autonomic cephalalgias
- Facial pain
- Trigeminovascular system (TVS)
7.1 Introduction
Headache is the symptom of head, face, or neck pain and represents the most common neurological symptom, experienced at least once in a lifetime almost universally. Headache disorders are classified as either primary or secondary. Primary headaches are highly prevalent conditions with no underlying cause, yet they can significantly burden a subgroup of patients. Conversely, secondary headaches are less common yet may herald life-threatening conditions and require urgent appropriate diagnostic investigation [1].
Primary headaches are a heterogeneous group, including migraine, tension-type headaches, and trigeminal autonomic cephalalgias (TACs). The exact pathophysiology of primary headaches is not fully understood, yet arguably involves a complex interplay of genetic, environmental, and biochemical factors. Migraine, for example, is thought to be caused by a combination of genetic susceptibility and environmental triggers, leading to changes in the brainstem and the trigeminal nerve that cause pain and other symptoms [2]. Tension-type headache may be related to muscle contractions and changes in neurotransmitter levels [3]. In contrast, cluster headache may involve alterations in the hypothalamus and its regulation of the circadian rhythm [4].
The autonomic nervous system (ANS) plays a role in the development and progression of primary headaches, particularly in migraine and TACs. Understanding the complex role of the ANS in primary headaches may inform their pathophysiology and treatment [5,6,7].
In this chapter, we will discuss the role of the ANS from different aspects in migraine and cluster headache (prototype of TACs): (i) pathophysiology, (ii) clinical autonomic manifestations, (iii) interictal autonomic dysfunction, (iv) autonomic modulation as a treatment strategy, and (v) interpretation of primary headaches in a bio-behavioural view.
7.2 Migraine Pathophysiology and the Autonomic Nervous System
Migraine is a complex neurological disorder characterized by recurrent headache attacks that last 4–72 h, typically causing severe pain on one side of the head, with accompanying symptoms such as nausea, vomiting, light and sound sensitivity, and intensifying with physical activity [2]. The migraine attack comprises distinctive consequential phases: prodrome symptoms, aura, headache phase, and postdrome symptoms. The presence of an aura preceding or concomitant to the headache phase distinguishes migraine into two main subtypes: migraine with aura and without aura. Migraine is highly prevalent in the general population, especially young women, and has a significant genetic component [8]. In the past, migraine was believed to be a cerebrovascular disorder; however, current research suggests a much more complex pathophysiology. The exact cause is still not fully understood, but it is thought to involve a primary brain dysfunction leading to activation and sensitization of the trigeminovascular system (TVS) [9]. TVS is composed of trigeminal and upper cervical dorsal root ganglions (first-order neurons), the trigeminocervical complex (second-order neurons), and ascending axonal projections to the midbrain, thalami, and hypothalamus (third-order neurons) [2].
The meninges and the pial cerebral vessels are the main pain-sensitive structures of the head and are innervated by trigeminal terminal ends that contain several neuropeptides, including calcitonin gene-related peptide (CGRP), pituitary adenylate cyclase-activating polypeptide (PACAP) and neuropeptide substance P (SP). Activation of the TVS promotes the blood release of these neuropeptides that are pivotal in migraine pain, as also suggested by the therapeutic efficacy of their inhibitors [10, 11].
Autonomic control of the central autonomic network (CAN) is exerted at every level of the TVS mechanisms [12]. Specifically, connections with the superior salivatory nucleus and the sphenopalatine ganglion are responsible for the cranial autonomic symptoms during migraine attacks, while TVS third-order neurons orchestrate autonomic-behaviour responses. Furthermore, the presence of non-noxious prodromes, such as somnolence, changes in fluid balance, hyperphagia, or food rejection, argues for a primary role of the CAN in the migraine attack generator [2, 8].
7.3 Cluster Headache Pathophysiology and the Autonomic Nervous System
TACs comprise a group of headache disorders characterized by recurrent attacks of strictly unilateral facial pain with predominant accompanying cranial autonomic symptoms [4, 13].
These disorders are distinguished based on clinical features, attack duration and frequency, responsiveness to therapy, and typical triggers (Fig. 7.1) [4, 13]. Cluster headache is the predominant subtype, accounting for more than 90% of the TAC spectrum. The other disorders are hemicrania continua, paroxysmal hemicrania, and short-lasting unilateral neuralgiform headache attack syndromes (SUNHA) [1]. Cluster headache patients experience bouts of headache attacks with circadian and circannual periodicity. The attacks are usually excruciating, accompanied by agitation and restlessness, and last 15–180 min. The underlying pathophysiology of cluster headache, and TACs at large, is complex yet more unveiled than other primary headaches. Interestingly, the autonomic nervous system plays a central role in these disorders. A dysfunctional hypothalamus is the well-known primum movens of cluster headache, as suggested by associated neuroendocrine changes, clinical periodicity, functional neuroimaging studies, and response to neuromodulation devices [4, 14,15,16]. Several hypothalamic nuclei are involved in the pathogenesis, such as the paraventricular nuclei that project to the superior salivatory nucleus (SSN) and the suprachiasmatic nucleus that is responsible for circadian periodicity. The dysfunctional hypothalamus arguably acts as the primary pacemaker and orchestrates the autonomic-behavioural responses of cluster headache attacks by activating the trigeminovascular system (TVS) and trigeminal autonomic reflex (TAR) that ultimately influence each other in a vicious circle [4].

Clinical characteristics of headache attacks frequency and duration in trigeminal autonomic cephalalgias. SUNHA short-lasting unilateral neuralgiform headache attack syndromes
7.4 Autonomic Manifestations during Migraine and Cluster Headache Attacks
In primary headaches, trigeminal autonomic symptoms secondary to an enhanced cranial parasympathetic outflow are frequently observed during headache attacks [5, 6]. These autonomic symptoms comprise conjunctival injection, lacrimation, periorbital swelling, ptosis, miosis, nasal congestion, rhinorrhea, and facial sweating. These autonomic manifestations are strictly unilateral and very prominent in all TACs and, consistently, are an essential part of the diagnostic criteria [1]. Nonetheless, these manifestations are frequently observed also in migraine attacks, yet usually milder and bilaterally. Notably, almost half of the migraine patients may display unilateral autonomic symptoms, usually accompanied by more severe pain intensity, potentially representing a distinctive migraine endophenotype with pathophysiological and treatment implications [17].
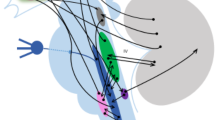
The trigeminal autonomic reflex (TAR) accounts for these cranial autonomic manifestations observed in primary headaches [5]. The TAR is a physiological reflex with a protective function engendered by any nociceptive stimulation of the trigeminal nerve endings (Fig. 7.2). This reflex protects mainly the eyes and sensitive facial skin areas but is also activated by intracranial structures such as the meninges. The information is transmitted to the trigeminal cervical complex, the thalamic, and cortical areas. Then, the efferents parasympathetic branches arise from the superior salivatory nucleus synapse with the sphenopalatine ganglion (SPG), where post-ganglionic fibres innervate the effector structures, such as lacrimal glands and Dural vessels [5]. The intensity of the autonomic manifestations during headache attacks usually corresponds to the severity of the pain experienced by the patient, further suggesting the protective role of the TAR. While activation of the TAR alone may not cause a primary headache attack, it may contribute to the escalation of headache pain during an attack by triggering centrally-mediated parasympathetic manifestations [18, 19]. Accordingly, several acute and preventive headache treatments target the TAR (see beyond).

Graphical view of the trigeminal autonomic reflex (TAR). Afferents and efferents components of the reflex are illustrated in blue and red, respectively. Th thalamus, SSN superior salivatory nucleus, TCC trigeminal cervical complex, SPG sphenopalatine ganglion
7.5 Interictal Autonomic Dysfunction in Migraine and Cluster Headache
Studies on interictal (between headache attacks) autonomic dysfunction in migraine have used various techniques, such as measuring cerebrovascular reactivity, pupil reactivity, cardiovascular reflexes, and biochemical and pharmacological responses. However, these studies have produced inconsistent results with no clear pattern of autonomic dysfunction observed, including hypo- and hyperfunction, as well as normal findings [20]. Therefore, there is currently no apparent autonomic deficit considered intrinsic to migraine headache.
Interictal autonomic dysfunction has also been investigated in cluster headache, mainly during the active cluster phase. A recent review focusing on cardiovascular autonomic changes found no significant or consistent interictal dysfunction except for a moderately increased sympathetic tone in cluster headache patients compared to control subjects [21]. Whereas, a following study demonstrated a prolonged latency of skin response on the affected side of the face, suggesting post-ganglionic hypofunction [22].
Collectively, studies investigating ANS dysfunction interictal in both migraine and cluster headache did not reveal significant alterations. Nonetheless, these studies did not consider modulation that headache attacks may involve integrated behavioural responses, where pain perception and modulation are a behavioural motif, and autonomic responses may serve that behaviour [7].
7.6 Primary Headache Treatment and Modulation of the ANS
Modulation of TAR may play a role in treating primary headache disorders such as migraine, cluster headache, and other TACs. Specific therapies, such as indomethacin, oxygen, and neurostimulation of the hypothalamus, sphenopalatine ganglion (SPG), and vagal nerve, have shown promise in impacting TAR and potentially treating these headache disorders [5]. However, more research is still needed to fully understand the relationship between TAR and primary headache disorders and determine the best treatment approach.
Indomethacin is considered an effective treatment for specific primary headaches, such as paroxysmal hemicrania and hemicrania continua [1], due to its ability to inhibit nitrogen oxide-induced Dural vasodilation [23]. This unique property, compared to other non-steroidal anti-inflammatory drugs (NSAIDs) [23], suggests that indomethacin may modulate TAR and its function in primary headache disorders [5]. Further research is needed to fully understand the mechanism behind indomethacin’s efficacy in treating these conditions.
Oxygen therapy effectively treats cluster headache attacks [24] and, to a lesser extent, migraine attacks with autonomic symptoms [25]. Preclinical studies have suggested that the brainstem plays a critical role in sensory and autonomic symptoms and that the parasympathetic outflow to the cranial vasculature may be a target for oxygen [26]. The fact that oxygen therapy is effective in both cluster headache and migraine with autonomic symptoms support the idea that it may modulate TAR.
High-frequency stimulation of the SPG is highly effective in treating cluster headache patients who have not responded to other treatments [27]. It works by blocking the parasympathetic discharge. Interestingly, low-frequency stimulation of the SPG, which was thought to trigger cluster headache attacks, was ineffective [18]. This reinforced the concept that the TAR’s peripheral activation is insufficient to trigger headache attacks.
Non-invasive vagal nerve stimulation (nVNS) was first attempted on refractory epilepsy [28], yet a concomitant reduction of migraine attacks frequency in epileptic patients was observed. After that, nVNS was tested in migraine and cluster headache and proved consistently effective [29, 30]. The underlying mechanism is not fully unveiled, yet a reduction of autonomic symptoms was observed, thus, modulation of the TAR is possible.
7.7 Primary Headaches as a Reflection of Genetic Darwinian Adaptive Autonomic-Behavioural Responses
Primary headaches are considered painful episodic disorders, yet pain does not represent the full spectrum of clinical manifestations observed during attacks and may even be missing in some rare headaches (e.g., migraine aura without headache) [1]. Therefore, primary headaches can only be fully understood by a comprehensive behavioural view [7]. In this model, the multifaceted phenomena of primary headaches are unified, and the autonomic nervous system’s significant role is highlighted not only for the pain but also for the non-headache (prodrome and postdrome) symptoms characteristics of the attacks.
7.7.1 Behaviours Engendered during Primary Headaches
During a migraine attack, patients exhibit a characteristic behaviour where they seek to become as immobile as possible and rest, avoiding physical and mental activity and trying to be in an environment with minimal stimulation [2].
Conversely, during a cluster headache attack, patients exhibit quite the opposite behaviour, characterized by restlessness, psychomotor agitation, aggressiveness, and even self-hurting behaviours [4].
7.7.2 The Behavioural Meaning of Pain
Headache, and pain at large, could be envisaged within the frame of its behavioural significance and utility as a powerful homeostatic emotion. The subjects can perceive pain as either escapable or inescapable and, therefore, is accompanied by opposite autonomic-behaviour responses (Fig. 7.3) [7, 31, 32]. Each type of pain is conveyed by different sensory modalities and pathways. Escapable pain is typically conveyed by A-delta fibres from the skin, whereas inescapable pain is conveyed by C fibres mainly from deep viscera, serving as visceral homeostatic pain [31, 32]. They also activate different pathways in the central nervous system, specifically in regions such as the periaqueductal Gray (PAG), hypothalamus, amygdala, and forebrain. Escapable pain-activated A-delta fibres project to the dorsolateral/lateral PAG and posterior hypothalamus that coordinate a sympathoexcitation autonomic-behavioural response, namely fight-or-flight defensive response [33]. This manifests with increased production of catecholamines, arterial hypertension, tachycardia, hyperventilation, increased blood flow to the skeletal muscles, aggressiveness, and restlessness. All these features represent an adaptive integrated innate response optimized to fight stressors or flight from potentially threatening situations (escapable pain).

Schematic representation of the different neuronal networks underlying autonomic-behavioural responses of escapable and inescapable pain. PAG periaqueductal gray
This leads to a fight-or-flight defensive response, with the subjects ready to confront the stressor or run from potentially threatening situations.
Conversely, inescapable pain-activated C fibres project to the ventrolateral PAG that coordinates a sympathoinhibition autonomic-behaviour response, namely sickness behaviour [32]. This leads to hypotension, bradycardia, motor quiescence, lethargy, and disengagement from the environment. These features represent an adaptive integrated innate response optimized to confront stressors in an unavoidable engagement (inescapable pain).
Collectively, these emotional and autonomic responses associated with pain have an evolutionary basis and are part of a defined behavioural programme. Darwin first applied an evolutionary perspective to emotions in animals and humans and identified patterns of emotional behaviours that had evolutionary significance.
7.7.3 The Behavioural Meaning of Migraine and Cluster Headache Attacks
The clinical manifestations observed during primary headache attacks remarkably overlap with the autonomic-behavioural responses shared by all mammalians, suggesting they are evolutionary conserved adaptive strategies [7].
The passive behaviour frequently observed during migraine attacks is likely related to a brain homeostatic imbalance signalled by trigeminal C fibres that help the brain to recover to the homeostasis of the brain itself. This bio-behavioural model of migraine explains the interictal traits distinctive of the migraine brain, such as the deficient brain energy metabolism and deficient habituation in information processing [34]. Therefore, this model views migraine pain as serving as a protective function, as a visceral homeostatic emotion like thirst and hunger that monitors the internal bodily world with an adaptive role. Hints of this fundamental role may come from the persistence of migraine as a frequent genetic trait in the population of reproductive age, potentially underlying an evolutionary advantage of migraineurs [35].
The active behaviour engendered during cluster headache attacks resembles the fight-or-flight defensive response and is arguably related to the activation of trigeminal A-delta fibres.
Therefore, behaviours during migraine and cluster headache attacks should be considered evolutionarily conserved adaptive responses. Nonetheless, even though the behaviours may have evolved for a specific purpose, they may become inappropriate and maladaptive in certain circumstances, such as aggressive or anxiety behaviours [7].
7.8 Conclusions
The ANS plays a pivotal role in primary headaches underlying pathophysiology and its involvement explains several clinical manifestations observed during headache attacks. Additionally, its iatrogenic modulation has important therapeutic implications. The bio-behavioural theory views primary headache attacks as adaptive behavioural responses engendered by a network of pattern generators in the ANS to maintain brain homeostasis. Therefore, to investigate the role of the ANS, it is futile to study autonomic functions during headache attacks separately from the bio-behavioural responses they contribute to.
References
Headache classification Committee of the International Headache Society (IHS) the international classification of headache disorders, 3rd edition. Cephalalgia 2018;38:1–211.
Ferrari MD, Goadsby PJ, Burstein R, et al. Migraine. Nat Rev Dis Primers. 2022;8:2.
Steel SJ, Robertson CE, Whealy MA. Current understanding of the pathophysiology and approach to tension-type headache. Curr Neurol Neurosci Rep. 2021;21:56.
Wei DY, Goadsby PJ. Cluster headache pathophysiology - insights from current and emerging treatments. Nat Rev Neurol. 2021;17:308–24.
Moller M, May A. The unique role of the trigeminal autonomic reflex and its modulation in primary headache disorders. Curr Opin Neurol. 2019;32:438–42.
Gevirtz R. The role of the autonomic nervous system in headache: biomarkers and treatment. Curr Pain Headache Rep. 2022;26:767–74.
Montagna P, Pierangeli G, Cortelli P. The primary headaches as a reflection of genetic darwinian adaptive behavioral responses. Headache. 2010;50:273–89.
Ashina M. Migraine. N Engl J Med. 2020;383:1866–76.
Ashina M, Hansen JM, Do TP, Melo-Carrillo A, Burstein R, Moskowitz MA. Migraine and the trigeminovascular system-40 years and counting. Lancet Neurol. 2019;18:795–804.
Do TP, Al-Saoudi A, Ashina M. Future prophylactic treatments in migraine: beyond anti-CGRP monoclonal antibodies and gepants. Rev Neurol (Paris). 2021;177:827–33.
Christensen CE, Ashina M, Amin FM. Calcitonin gene-related peptide (CGRP) and pituitary adenylate cyclase-activating polypeptide (PACAP) in migraine pathogenesis. Pharmaceuticals (Basel). 2022;15:15.
Benarroch EE. Pain-autonomic interactions. Neurol Sci. 2006;27(Suppl 2):S130–3.
Nahas SJ. Cluster headache and other trigeminal autonomic Cephalalgias. Continuum (Minneap Minn). 2021;27:633–51.
Leone M, Bussone G. A review of hormonal findings in cluster headache. Evidence for hypothalamic involvement. Cephalalgia. 1993;13:309–17.
May A, Bahra A, Buchel C, Frackowiak RS, Goadsby PJ. Hypothalamic activation in cluster headache attacks. Lancet. 1998;352:275–8.
Leone M, Franzini A, Felisati G, et al. Deep brain stimulation and cluster headache. Neurol Sci. 2005;26(Suppl 2):s138–9.
Barbanti P, Aurilia C, Dall’Armi V, Egeo G, Fofi L, Bonassi S. The phenotype of migraine with unilateral cranial autonomic symptoms documents increased peripheral and central trigeminal sensitization. A case series of 757 patients. Cephalalgia. 2016;36:1334–40.
Moller M, Haji AA, Hoffmann J, May A. Peripheral provocation of cranial autonomic symptoms is not sufficient to trigger cluster headache attacks. Cephalalgia. 2018;38:1498–502.
Guo S, Petersen AS, Schytz HW, et al. Cranial parasympathetic activation induces autonomic symptoms but no cluster headache attacks. Cephalalgia. 2018;38:1418–28.
Miglis MG. Migraine and autonomic dysfunction: which is the horse and which is the jockey? Curr Pain Headache Rep. 2018;22:19.
Barloese MC. A review of cardiovascular autonomic control in cluster headache. Headache. 2016;56:225–39.
Altiokka O, Mutluay B, Koksal A, et al. Evaluation of interictal autonomic function during attack and remission periods in cluster headaches. Cephalalgia. 2016;36:37–43.
Summ O, Andreou AP, Akerman S, Goadsby PJ. A potential nitrergic mechanism of action for indomethacin, but not of other COX inhibitors: relevance to indomethacin-sensitive headaches. J Headache Pain. 2010;11:477–83.
Cohen AS, Burns B, Goadsby PJ. High-flow oxygen for treatment of cluster headache: a randomized trial. JAMA. 2009;302:2451–7.
Jürgens TP, Schulte LH, May A. Oxygen treatment is effective in migraine with autonomic symptoms. Cephalalgia. 2013;33:65–7.
Akerman S, Holland PR, Summ O, Lasalandra MP, Goadsby PJ. A translational in vivo model of trigeminal autonomic cephalalgias: therapeutic characterization. Brain. 2012;135:3664–75.
Jürgens TP, Barloese M, May A, et al. Long-term effectiveness of sphenopalatine ganglion stimulation for cluster headache. Cephalalgia. 2017;37:423–34.
Oliveira T, Francisco AN, Demartini ZJ, Stebel SL. The role of vagus nerve stimulation in refractory epilepsy. Arq Neuropsiquiatr. 2017;75:657–66.
Gaul C, Diener HC, Silver N, et al. Non-invasive vagus nerve stimulation for PREVention and acute treatment of chronic cluster headache (PREVA): a randomised controlled study. Cephalalgia. 2016;36:534–46.
Tassorelli C, Grazzi L, de Tommaso M, et al. Noninvasive vagus nerve stimulation as acute therapy for migraine: the randomized PRESTO study. Neurology. 2018;91:e364–e73.
Keay KA, Bandler R. Distinct central representations of inescapable and escapable pain: observations and speculation. Exp Physiol. 2002;87:275–9.
Parry DM, Macmillan FM, Koutsikou S, McMullan S, Lumb BM. Separation of A- versus C-nociceptive inputs into spinal-brainstem circuits. Neuroscience. 2008;152:1076–85.
Lumb BM. Hypothalamic and midbrain circuitry that distinguishes between escapable and inescapable pain. News Physiol Sci. 2004;19:22–6.
Gross EC, Lisicki M, Fischer D, Sandor PS, Schoenen J. The metabolic face of migraine - from pathophysiology to treatment. Nat Rev Neurol. 2019;15:627–43.
Montagna P. Migraine genetics. Expert Rev Neurother. 2008;8:1321–30.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2023 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Cortelli, P., Pensato, U. (2023). Primary Headaches and the Autonomic Nervous System. In: Micieli, G., Hilz, M., Cortelli, P. (eds) Autonomic Disorders in Clinical Practice. Springer, Cham. https://doi.org/10.1007/978-3-031-43036-7_7
Download citation
DOI: https://doi.org/10.1007/978-3-031-43036-7_7
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-43035-0
Online ISBN: 978-3-031-43036-7
eBook Packages: MedicineMedicine (R0)