
Abstract
Today’s Campos Sulinos have witnessed important paleogeographic, paleoclimatic, and paleoenvironmental shifts in the last 300 million years. When this history is evaluated through the perspective of fossil-bearing sedimentary rocks, it can be divided into four major events: Permian deglaciation and evolution to an alluvial plain in Pangean condition; Triassic recovery of a massive extinction and development in alluvial plains in Pangean circumstances; establishment of an aeolian system under desertic conditions during the Jurassic-Cretaceous; and the Pleistocene onset of megafauna and dominance of grassland vegetation responding to glacial and interglacial cycles. Permian biotas range from marine parareptiles, such as mesosaurs, to fully terrestrial fauna, including large herbivores such as provelosaurids, a saber-toothed herbivore, and large carnivores. Terrestrial environments were dominated by a Glossopteris Flora, with large pteridophytes and primitive gymnosperms. Triassic biota vary from near-the-water parareptiles, temnospondyls, and archosauromorphs, right after the most impressive mass extinction on our planet, to the onset of dinosaurs and the evolution of cynodonts. The Triassic also witnessed one of the main vegetation transitions in Earth’s history: environments once dominated by a Glossopteris Flora gradually became occupied by Dicroidium and Coniferous Floras. The Jurassic faunas are mostly represented by dinosaur footprints from a desert-dominated environment prior to the South America/Africa breakup. Late Pleistocene biota includes large-bodied mammals and reptiles, representatives of the so-called Pleistocene Megafauna. In addition, extant taxa are also present in the fossil record. The Late Pleistocene paleobotanic fossil record demonstrates widespread grasslands that were only slowly subject to the expansion of forests.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
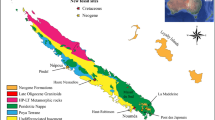
The Campos Sulinos, i.e., the grasslands that dominate natural landscapes in the three southernmost states of Brazil, Rio Grande do Sul (SC), Santa Catarina (SC, and Paraná (PR), extend over different geological grounds in southern Brazil: igneous and metamorphic rock from the crystalline basement, fossiliferous sedimentary rocks from the Paraná Basin, volcanic rocks from the top of Serra Geral, and fluvial and coastal sedimentary deposits (Silva and Vaine 2001; Wildner et al. 2008, 2014; see also Menegat 2023, Chap. 2, this volume). This region witnessed important paleogeographic, paleoclimatic, and paleobiotic shifts in the last 300 million years, which will be summarized here (Fig. 3.1). Four major moments can be envisaged in this scenario in southern Brazil. The Permian started with a Gondwanic glaciation, but subsequent deglaciation led to sea level rise, the formation of an internal sea, and evolution to an alluvial plain in Pangean conditions. The Triassic witnessed the recovery of a massive extinction and development in alluvial plains in Pangean circumstances. The Late Jurassic and Early Cretaceous are known for their desert environments, prior to the breakup of South America and Africa. Finally, the glacial and interglacial cycles of the Pleistocene led to the onset of megamammals and large reptiles, later extinct between the latest Pleistocene and early Holocene, and the establishment of “campos”–forest mosaics.

Geological time scale and positioning of “Campos Sulinos” fossiliferous rocks in Permian (280 million years, Ma), Triassic (220 Ma), and Pleistocene (Last Maximum Glacial). (Global reconstructions after Ron Blakey)
2 Paleogeographic and Paleoenvironmental Reconstruction
Here we present the geobiological transformation over the last 300 million years (Ma), depicting four different periods: the Permian, Triassic, Jurassic-Cretaceous periods, and the Pleistocene Epoch (Fig. 3.1).
2.1 Permian
During the Permian, all land masses were gathered in a single, “C”-shaped supercontinent called Pangaea (see Fig. 3.1). This term was coined by Alfred Wegener in 1912 to designate the geological and paleobiological coalescence of present continents into a single supercontinent (Wegener 1912). According to current knowledge, the Pangaea was formed from the convergence of Laurentia, Baltica, and Gondwana during the late Carboniferous (Scotese and Golonka 1997). This supercontinent was bordered by two oceans, the Panthalassa and Tethys.
The Paraná Basin was part of the Gondwanan realm of Pangaea. Its geological history goes from glaciation at the beginning of the Permian period (Itararé Group) and later deglaciation, which led to sea level rise, coastal shift (Rio Bonito and Palermo Formations), and formation of an epicontinental sea (Irati Formation) in the mid-Permian (Holz et al. 2010). During the late Permian, continentalization started, leading to the formation of shallow lakes (Teresina Formation) and alluvial plains (Rio do Rasto Formation). The rocks from the Itararé Group are the record of glacial and postglacial environments, such as glaciomarine or glaciolacustrine settings (Vesely 2007; Weinschütz and Castro 2006). The record of Gondwanan glaciation in Brazil is preserved in some isolate sites, such as the Witmarsun and Salto glacial striations and Mafra dropstones (Santa Catarina), ruiniform relief at Vila Velha State Park (Paraná), and Itu Varvite (São Paulo) (Rocha-Campos 2002a, b; Melo et al. 2002; Pérez-Aguilar et al. 2008). Some outcrops are also known from Rio Grande do Sul.
The Rio Bonito Formation records lagoon-barriers deposited under sea level rise conditions, right after deglaciation. These more humid conditions led to the deposition of peat environments and coal formation (Lavina and Lopes 1987; Castro et al. 1999; Maahs et al. 2019). These rocks include leaf imprints of the Glossopteris Flora, which Alfred Wegener used as evidence of continental drift (Iannuzzi 2010; Jasper et al. 2009). The Palermo Formation is formed by mudstones and shales, due to a more prominent sea level rise, only recording rare marine microfossils (Simões and Rocha-Campos 1993). Outcrops are known from São Paulo, Paraná, Santa Catarina, and Rio Grande do Sul.
The Irati Formation records an epicontinental sea with bituminous shales (Chahud et al. 2012). Its fossiliferous content, mainly characterized by crustaceans and mesosaurs, are worthy of mention, as they were also used as evidence of continental drift by Alfred Wegener (Araújo-Barberena et al. 2002; Cerri et al. 2020; Oleofsen and Araújo 1983a, b, 1987a, b; Ricardi-Branco et al. 2009). Silicified logs are also present (Merlotti 2009). The preservation of almost complete skeletons of mesosaurs is related to winter storms and sea floor erosion (Lavina et al. 1991; Xavier et al. 2018). Outcrops are known from Paraná and Rio Grande do Sul states.
The Serra Alta Formation is composed of regressive facies of dark gray mudstones (Warren et al. 2015), recording bivalves (Matos et al. 2017a, b), ostracods (Bergue et al. 2020), and silicified logs (Merlotti 2009). The Teresina Formation is characterized by gray siltstones, intercalated with micritic carbonate and stromatolites, representing the transition from marine environments to marine zones with shallow water influenced by storms (Rohn 2001; Silva and Vaine 2001; Callefo et al. 2015), with fossils restricted to palynomorphs (Neregatto et al. 2008), pelecypods (Silva and Vaine 2001), ostracods (Bergue et al. 2020), and microbial mats (Callefo et al. 2015).
The Rio do Rasto Formation facies are diverse, with fluvial, aeolian, and lacustrine environments (Schemiko et al. 2014). Their sedimentary evolution indicates the formation of a large confined water body and the accumulation of a transgressive-regressive sequence at its lower portion, eventually recording storm conditions, while the progradational character of the upper portion led to the colmatation of the water body with deltaic and aeolian deposition (Warren et al. 2008). The Rio do Rasto Formation was considered Lopingian (Malabarba et al. 2003). However, a recent U-Pb age repositioned it to Guadalupian (Francischini et al. 2018).
2.2 Triassic
Still under the overall condition of continental agglutination, the Triassic witnessed a slight northern displacement of Pangea (Fig. 3.1), maintaining the paleoequator at the inflection of the boomerang-like structure (Golonka et al. 2018). At the end of the Triassic (Norian and Rhaetian), a massive bulging took place at that point, leading to the formation of a magmatic province, namely the Central Atlantic Magmatic Province (CAMP; Marzoli et al. 2018), which is related to the opening of the Atlantic Ocean and the extinction of several land vertebrates (Davies et al. 2017).
The Triassic record of the Paraná Basin is restricted to its southern portion, in Rio Grande do Sul. Here, two depositional packages were recognized, the Sanga do Cabral Supersequence, composed of the arenaceous rocks of the homonymous geological formation, and the Santa Maria Supersequence, integrating the Santa Maria and Caturrita formations (Zerfass et al. 2003). More recently, Horn et al. (2014) subdivided the Santa Maria Supersequence into third-order sequences: Pinheiros-Chiniquá (Ladinian?), Santa Cruz (early Carnian), Candelária (late Carnian to Norian), and Mata (Rhaetian?).
The Sanga do Cabral Supersequence records fine sandstones with intercalated intraformational conglomerates, deposited in broad alluvial plains (Da Rosa et al. 2009a, b), while the Santa Maria Supersequence (SMS) presents a more complex organization (Da Rosa 2015). The lower part of the SMS, i.e., the Pinheiros-Chiniquá, Santa Cruz, and the lower portion of the Candelária sequences, records the intercalation of sandstones and fine-grained lithologies, deposited at low energy, high sinuosity rivers, representing channels, levees, overbank deposits, shallow and temporary lakes, loess, and paleosols (Da Rosa et al. 2004a, b, 2005; Horn et al. 2013). On the other hand, the upper portion of the SMS shows a predominantly sandy deposition in high energy, low sinuosity rivers, with channels and overbank deposits (Müller et al. 2015).
2.3 Jurassic/Cretaceous
The time interval between the end of the Jurassic and the beginning of the Cretaceous is characterized by the continuation of the Pangaea breakup, with main interest in the separation of South America and Africa circa 135 Ma ago. In the Paraná Basin, the Guará Formation consists of fine to conglomeratic sandstones of whitish to reddish color, with trough cross-stratification from small to large size and horizontal lamination. The facies associations characterize fluvial to fluvio-aeolian deposits, with a distinctive change throughout its exposure, the northern part consisting of facies associations of braided fluvial systems and the southern part showing a tendency to replace the predominant fluvial facies by aeolian associations (Scherer and Lavina 2005). These fluviatile-desertic sandstones are correlated to the Tacuarembó Formation in Uruguay and placed at the end of the Jurassic based on their faunal content (see Sect. 3.3.3).
2.4 Pleistocene
The main Cenozoic units from Rio Grande do Sul are restricted to the Quaternary Period (2.58 million years ago to the present), more precisely between the Middle Pleistocene and the Holocene. During the Pleistocene, the continents were already organized in their present condition (Fig. 3.1), with rapidly changing environments due to glacial cycles and subsequent sea level fall, the imposition of dry climates, and the shift of river profiles and coastal lines (Golonka 2009). The Pleistocene deposits, mostly formed in fluvial environments, are found in western RS (WRS – Alegrete, Quaraí, and Uruguaiana) (Lopes et al. 2020), central region (Caçapava do Sul, Pantano Grande), and Coastal Plain (CPRS – Santa Vitória do Palmar). In addition to these locations, there are several others with sparse records, such as São Gabriel and Jaguarão. Different methodologies, such as Carbon-14 (14C), Accelerated Mass Spectrometry (AMS), Electron Spin Resonance (ESR), and Optically Stimulated Luminescence (OSL), have been applied to time-calibrate fossils and Pleistocene sediments. Radiometric ages from WRS (Touro Passo Creek, Uruguaiana; Quaraí River, Quaraí; Sanga da Cruz and Ibicuí River, Alegrete) are placed between ~42 and 10 ka (Bombin 1976; Miller 1987; Kotzian et al. 2005; Milder 2000; Ribeiro and Scherer 2009; Kerber et al. 2011a, b, c), while deposits from the CPRS include older sediments. Chuí Creek sedimentary strata (Santa Vitória Formation, Santa Vitória do Palmar) were deposited between ~190 and ~30 ka, with most fossils dated between 50 and 30 ka (Lopes et al. 2014a, b, 2019, 2020). The fossils from onshore deposits found in Balneario Hermenegildo (Santa Vitória do Palmar) are even older, dated between 650 and 18 ka (Lopes et al. 2010). Concerning the record of tetrapods, the fossils found in these locations show a certain taxonomic uniformity (although some peculiarities are observed; see Sect. 3.4) and mostly represent fossils of large mammals (Mammalia) known as the Pleistocene megafauna. However, other small vertebrates, such as lizards, birds, and fishes, are also recorded.
3 Biodiversity and Paleoecological Reconstruction of Fauna and Flora
Here, we summarize the faunistic (tetrapods) and floristic biodiversity and paleoecological inferences of the studied time intervals, i.e., Permian, Triassic, Jurassic/Cretaceous, and Pleistocene.
3.1 Permian
Almost all Permian rocks in the Campos Sulinos region have a fossil record restricted to fish, invertebrates and/or plant fossils. However, exceptions are the Irati and Rio do Rasto formations, which present tetrapods in a more complex food chain. Few fossils are known from the Itararé Group, and they consist of invertebrates (Matos et al. 2017a; Neves et al. 2014; Taboada et al. 2016), fish fragments and ichnofossils, such as annelid and trilobite trackways (Netto et al. 2009; Lima et al. 2015; Wilner et al. 2016; Noll and Netto 2018). The Irati Formation is remarkable for its exceptional record of mesosaurs, i.e., parareptilles adapted to an aquatic way of life, such as thickened bones (especially ribs) and characteristic teeth. Three mesosaur species were proposed for the Irati Formation, Mesosaurus brasiliensis, “Stereosternum tumidum”, and “Brazilosaurus saopaolensis”). Piñeiro et al. (2021), however, recognized only the former as valid. Mesosaurs seem to have fed on fish and pygocephalomorph crustaceans in the Irati-Whitehill Sea.
A complex and diversified fauna inhabited the aquatic environments associated with the deposition of the Rio do Rasto Formation. The unit already yielded abundant actinopterygian remains, even though those are often represented by fragmentary bones and isolated “paleonisciform” scales. Exceptions to these highly transported, mainly uninformative materials come from the Serrinha Member, known for its representativeness of low-energy depositional environments (e.g., Milani et al. 2007). Perhaps the best-preserved actinopterygian so far recovered from the Rio do Rasto Formation is the bizarre Paranaichthys longianalis, a laterally flattened fish with flamboyant anal fins (Dias 2012). Paranaichthys is known solely from its holotype, a superbly preserved specimen recovered in Paraná state. Rubidus pascoalensis, another Rio do Rasto actinopterygian known from a complete specimen, had a more elongated body, displaying several rows of diamond-shaped scales (Richter 2002).
The diagnostic nature of isolated shark teeth and fin spines made it possible to recognize three nominal shark species for the Rio do Rasto Formation, all known by fragmentary specimens. Two of these, Xenacanthus ragonhai and Triodus richterae (Pauliv et al. 2014, 2017), belong to the Xenacanthiformes, eel-like sharks with long and low dorsal fins and strong denticulated fin spines. Xenacanthiform teeth are easily recognized by their tricuspid crowns, with long lateral cusps flanking a smaller central one. Recent findings suggest that xenacanthiforms may be at least partially responsible for producing the exceptional coprolite concentrations found in Rio do Rasto outcrops (e.g., Dentzien-Dias et al. 2012a, b). A third shark species, Sphenacanthus riorastoensis, belongs to the enigmatic Sphenacanthidae, and is only known from isolated fin spines (Pauliv et al. 2012).
Sharks were not the apex predators in mid- and late-Permian freshwater bodies. At that time, this role was played by the Temnospondyli, an extraordinarily diverse clade of early-diverging tetrapods commonly referred to as “amphibians” (Schoch 2013). With a wide range of sizes and an obligate carnivorous diet, temnospondyls may have played an ecological role similar to that of modern-day crocodilians, even though several representatives were adapted to feed in terrestrial environments (Clack 2012). Several temnospondyl nominal species are known from the Rio do Rasto Formation, this being by far the best-represented group in this sedimentary unit. Like modern caimans, some Rio do Rasto temnospondyls, such as Rastosuchus hammeri (Dias et al. 2020) and Parapytanga catarinensis (Strapasson et al. 2015) had short and broad snouts, while the bizarre Australerpeton cosgriffi and Bageherpeton longignathus had exceptionally long and slender snouts, completely filled with small (but sharp) needle-shaped teeth (Dias and Barberena 2001; Pacheco et al. 2016; Azevedo et al. 2017).
Synapsids, the mammal forerunners, were abundant during the Permian period, dominating most land ecosystems and occupying several ecological roles that ranged from primary consumers to apex predators. In the Rio do Rasto Formation, herbivore synapsids belong to the clade Anomodontia. Anomodonts were diverse forms of mainly tusked herbivores that prospered during most of the late Permian and Triassic times. Early-diverging anomodonts are rare in the fossil record. Among these, the most striking is certainly Tiarajudens eccentricus, a saber-toothed pig-sized animal known by an almost complete skull and some postcranial remains from a Rio do Rasto Formation outcrop. Besides long canines, Tiarajudens had strong molariforms adapted to macerate vegetation (Cisneros et al. 2011, 2015). The saber-like canines of Tiarajudens likely evolved as display structures, perhaps also being used during intraspecific agonistic interactions, similar to what is seen today in musk deers (Cisneros et al. 2015).
Dicynodontia, the most diverse anomodont subclade, has two representatives in the Permian of Brazil. Among these, the best-known is Rastodon procurvidens, a diminutive (~40 cm long) dicynodont that lacked teeth except for two small forward-directed banana-shaped tusks (Boos et al. 2016; Simão-Oliveira et al. 2019). Rastodon is known from a single, superbly preserved specimen, including an almost complete skull and several postcranial bones. As can be extrapolated from other Permian dicynodonts, Rastodon was probably gregarious and excavated burrows for protection (Simão-Oliveira et al. 2019). The second Rio do Rasto Formation dicynodont, Endothiodon sp., is known only from a crushed and incomplete skull from Paraná (Barberena and Araújo 1975; Boos et al. 2013). Despite fragmentary, Endothiodon is a key taxon to understanding Rio do Rasto Formation geochronology, allowing a direct correlation with the better-studied South African Karoo Basin.
The remaining synapsids recovered from the Rio do Rasto Formation belong to the clade Dinocephalia. Medium to large animals, dinocephalians apparently had a wide range of feeding habits and were first reported from the Rio do Rasto Formation through isolated teeth and jaw fragments (Langer 2000). Only in 2012 the first reasonably complete specimen was introduced, the complete (albeit crushed) skull of the medium-sized carnivore Pampaphoneus biccai (Cisneros et al. 2012). This specimen so far represents the largest land predator of the Brazilian Permian, probably occupying an apex position in its food chains. A second, more complete Pampaphoneus specimen was recently recovered from the same outcrop where the holotype was found (Costa Santos et al. 2023). This new specimen contributes with information on the anatomy of this carnivorous dinocephalian. Still unpublished, this specimen will contribute with information on the anatomy of this carnivorous dinocephalian. The Rio do Rasto Formation also yielded a single herbivorous dinocephalian specimen (Boos et al. 2015). Albeit not sufficiently complete to allow a specific attribution, the material confirms the presence of the dome-headed tapinocephalid dinocephalians in the Brazilian Permian. A summarized scenery of the Rio do Rasto Formation is available in Fig. 3.2.

Permian. Faunistic elements depicted, from front to back: Australerpeton, Bageherpeton, Rastodon, Tiarajudens, Provelosaurus, Pampaphoneus, and indeterminate tapinocephalian. (Floristic records: Equisetales and Cycadaceae. Artwork: Voltaire Dutra Paes Neto)
The taphoflora preserved in the Rio do Rasto Formation deposits is characterized by the latest records of the Glossopteris Flora, which dominated most environments during the Permian Period, before the formation of desertic landscapes at the end of the Paleozoic. The Glossopteris Flora represents one of the most important paleofloras preserved over geological time. In addition to its wide paleogeographic distribution, which attests to the continental drift proposed by Alfred Wegener in 1912, this flora also witnessed the most important environmental and climatic changes that occurred in the course of the Permian Period, from the transition of the icehouse to the greenhouse stage and the aridization of previously moist environments, culminating in the Earth’s greatest mass extinction.
The main floristic elements that characterize the Glossopteris Flora were diagnosed mainly through impressions of leaves and stems of Glossopteris, Gangamopteris, Rubidgea, and Buriadia (Guerra-Sommer and Cazzulo-Klepzig 2000a, b). Other groups still present in today’s environments, such as the lycophytes Cyclodendron sp. (Herbst 1986) and Lycopodiopsis sp. (e.g., Martins and Sena-Sobrinho 1951; Ferraz et al. 2021), the sphenophyte Shenophylum sp. (e.g., Rohn and Rösler 1990; Iannuzzi 2010), and the pteridophytes Pecopteris sp. (e.g., Cazzulo-Klepzig 1978; Rohn and Rösler 1986; Ferraz et al. 2021), Schizoneura sp. (e.g., Cazzulo-Klepzig 1978; Cazzulo-Klepzig and Correia 1981), Dichophyllites sp. (Bortoluzzi 1975; Rohn 1988), and Paracalamites sp. (Bortoluzzi 1975; Rösler and Rohn 1984), have also been documented.
Although vegetation records in the Rio do Rasto Formation predominantly indicate hygrophilic and mesophilic habits (Rohn 1988), the slow process of aridization during this period was documented through the reduction of floristic diversity. Records of pteridophytes become rarer, lycophytes are represented only by arborescent and shrubby forms, and Glossopteris species show changes in their leaf structure, with a reduction in limb dimensions and secondary venation of leaves. These signs suggest a warm and semi-arid climate, with long dry seasons alternating with rainy seasons of moderate rainfall (Guerra-Sommer and Cazzulo-Klepzig 2000a, b).
Due to environmental disturbances in the Permian (Guadalupian), including forest fires (Manfroi et al. 2015), there was a significant change in the floristic landscape, also affecting the cohabiting fauna. For example, evidence of insect–plant interactions in these deposits was seen in Glossopteris sp. with damaged leaf edges, indicating phytophagy and climatic cyclicity (Rohn 1988). Likewise, herbivorous vertebrates that fed on the existing vegetation on the edges of Permian lakes suffered from the reduction of available plant biomass, such as recorded for the Equisetales. This may have caused a disturbance in the entire food chain, which, along with other factors, later resulted in the end of the Paleozoic Era.
3.2 Triassic
The tetrapod biodiversity of the Brazilian Triassic is summarized into five assemblage zones (AZs) of biostratigraphic relevance (Schultz et al. 2020). Each AZ can be regarded as a different fauna. The AZs are organized in time, so that their succession represents the evolution of land communities from the devastated, postextinction Early Triassic environments toward the mature, stable Late Triassic ecosystems in which the first dinosaurs dwelt.
The diverse ecomorphological adaptations present in vertebrates from each AZ suggest the presence of complex ecological interactions and food chains during the time intervals recorded in the southern Brazilian fossil record (see evidence of trophic relationships in Schwanke and Kellner 2009). The earliest Brazilian Triassic assemblage zone is the Procolophon AZ. Albeit still poorly sampled, this AZ has recently been shown to hold an important and relatively diverse tetrapod fauna. The archosauromorph Teyujagua paradoxa and temnospondyl amphibians (e.g., Sangaia lavinai and Tomeia witecki) played their roles as carnivores in Early Triassic times (Pinheiro et al. 2016, 2019; Eltink et al. 2016). At the same time, the parareptiles Procolophon trigoniceps and Oryporan insolitus were probably omnivore/herbivore animals (Pinheiro et al. 2021). The proterosuchid archosauriforms and the tanystropheid Elessaurus gondwanaoccidens were probably specialized faunivores (De-Oliveira et al. 2020, 2022). Although inferring Early Triassic food chains is not an easy task, the increasing number of specimens recovered will certainly improve our knowledge of ecosystem dynamics in this period.
The second, younger Brazilian Triassic assemblage zone is the Dinodontosaurus AZ (Schultz et al. 2020). This AZ records a more complex ecosystem, where huge archosaurs such as Prestosuchus chiniquensis (~12 m long) were the apex predators. In contrast, smaller archosaurs (e.g., Decuriasuchus quartacolonia, Pagosvenator candelariensis, Archeopelta arborensis, and indeterminate proterochampsids and rauisuchians) may have played a secondary role as carnivores. In this AZ, non-mammaliaform cynodonts become very diversified in the fossil record of Rio Grande do Sul. The herbivore/omnivore cynognathians are represented by several traversodontids: Massetognathus ochagaviae and M. pascuali, Traversodon stahleckeri, Protuberum cabralense, Luangwa sudamericana, and Scalenodon ribeiroae (von Huene 1936; Abdala and Teixeira 2004; Liu et al. 2008; Reichel et al. 2009; Melo et al. 2017). The last two taxa have coeval species recorded in African Triassic strata. Probainognathians (mostly carnivore/insectivore cynodonts) include Chiniquodon theotonicus, Protheriodon estudianti, Candelariodon barberenai, Aleodon cromptoni, and Bonacynodon schultzi (von Huene 1936; Bonaparte et al. 2006; Oliveira et al. 2011; Martinelli et al. 2016, 2017a). Dicynodonts were herbivores of large (Stahleckeria potens) and medium size (Dinodontosaurus) (von Huene 1935; Schultz et al. 2020). The owennetid parareptile Candelaria barbouri seems to have been faunivorous, probably insectivorous.
The next assemblage zone (Santacruzodon AZ) has a still scarce fossil record, which can be attributed to sampling bias or a possible co-occurrence with the older Dinodontosaurus AZ. This biozone has produced a number of traversodontid and probainognathian cynodonts, but rauisuchian and proterochampsid archosaurs are also present. The traversodontid Santacruzodon hopsoni, which names the AZ, is the most abundant cynodont (Abdala and Ribeiro 2003), but there are also records of Massetognathus ochagaviae, Menadon besairiei, and the probainognathians Santacruzgnathus abdalai and Chiniquodon sp. (Melo et al. 2015; Martinelli et al. 2016; Schmitt et al. 2019). Interestingly, the Malagasy Menadon besairiei is a Triassic cynodont with hypsodont postcanines (indicating the presence of specialized adaptations in herbivores as a result of the arid Triassic environments), previously described from the ‘Isalo II’ group of Madagascar (Melo et al. 2015, 2019). Aside from indeterminate proterochampsids (Hsiou et al. 2002; Raugust et al. 2013; Ezcurra et al. 2015), a single large-sized predator archosaur was recorded for the Santacruzodon AZ. Dagasuchus santacruzensis, known solely from a fragmentary pelvis (Lacerda et al. 2015), was a medium-sized rauisuchian, probably very similar to its “cousin” Prestosuchus chiniquensis.
The Hyperodapedon AZ was probably the most diverse assemblage zone, in which several carnivores disputed the same geographic area (Garcia et al. 2019). Archosaurs such as proterochampsids, rauisuchians, silesaurids, and early dinosaurs (herrerasaurids and basal sauropodomorphs) were the main predators. The most common herbivores were rhynchosaurs, but cynodonts were also present, as well as clevosaurid sphenodontians. Procolophonians were never recorded from this time interval, but a new taxon is currently under study (Silva-Neves in prep). Regarding the cynodonts, the cynognathians are represented by the traversodontid Exaeretodon riograndensis, one of the most abundant fossils of this association, Gomphodontosuchus brasiliensis, represented by few remains, and the recently described Santagnathus mariensis (von Huene 1928; Hopson 1985; Abdala et al. 2002; Schultz et al. 2020; Schmitt et al. 2023). Probainognathians are more diverse and include the enigmatic Charruodon tetracuspidatus, the large body-sized ecteniid Trucidocynodon riograndensis, and the small prozostrodontians, probably insectivorous or carnivores, Prozostrodon brasiliensis, Therioherpeton cargnini, and Alemoatherium huebneri (Barberena et al. 1987; Abdala and Ribeiro 2000; Oliveira et al. 2010; Martinelli et al. 2017b; Kerber et al. 2020, 2022; Stefanello et al. 2023). In addition, two other cynodonts may be included in this AZ, or alternatively in the younger Riograndia AZ, or even at an as-yet-unknown interval: the traversodontid Siriusgnathus niemeyerorum and the probainognathian Agudotherium gassenae, both recovered from outcrops in Agudo municipality in central RS, but without index fossils so far (Pavanatto et al. 2018; Miron et al. 2020; Martinelli et al. 2020). The fossil content of these two localities appears to be endemic when compared to other Brazilian Triassic outcrops.
Archosauromorph diapsids were also a major component of the Hyperodapedon AZ. As mentioned before, the most abundant herbivores were the rhynchosaurs, bizarre pig-sized early archosauromorphs bearing beak-like projections on their jaws. Two rhynchosaur taxa are known for this AZ: the widespread genus Hyperodapedon and the smaller, more recently described species Teyumbaita sulcognathus (Montefeltro et al. 2010). The bulky aetosaurs were armored omnivore archosaurs, represented by the taxa Aetosauroides scagliai and Aetobarbakinoides brasiliensis (e.g., Desojo et al. 2012; Brust et al. 2018; Paes-Neto et al. 2021). Aside from these species, Hyperodapedon AZ archosauromorphs are also represented by the proterochampsids Rhadinosuchus gracilis (von Huene 1938), Cerritosaurus binsfeldi (Price 1946) and Proterochampsa nodosa (Barberena 1982), as well as the rauisuchian Rauisuchus tiradentes (von Huene 1938). Most interestingly, the Hyperodapedon AZ preserves a wealth of early dinosaurs, most of which being forerunners of the long-necked sauropods, such as the sauropodomorphs Buriolestes schultzi (Cabreira et al. 2016), Saturnalia tupiniquim (Langer et al. 1999), Bagualosaurus agudoensis (Pretto et al. 2018), and Pampadromaeus barberenai (Cabreira et al. 2011). Typical carnivores are represented by the Herrerasauridae, medium-sized apex predators bearing sharp teeth and hooked hand claws (Staurikosaurus pricei and Gnathovorax cabreirai) (Colbert 1970; Pacheco et al. 2019). Finally, Ixalerpeton polesinensis was a small dinosaur-related gracile archosaur, recently proven to be akin to the pterosaurs (Cabreira et al. 2016; Ezcurra et al. 2020). Figure 3.3 illustrates some taxa of the Hyperodapedon AZ.

Triassic. Faunistic elements, from front to back: Hyperodapedon, Exaeretodon, Gnathovorax, and Dynamosuchus. (Floristic records: Dicroidium, Cycadaceae. Artwork: Márcio L. Castro)
Unlike the Carnian strata, the Norian fossil record of synapsids in the subsequent Riograndia AZ is characterized by the return of large-sized dicynodonts to the Brazilian fossil record. The only known taxon is the stahleckeriid Jachaleria candelariensis (Kannemeyeriiformes), which represents one of the last South American dicynodonts (Araújo and Gonzaga 1980; Martinelli et al. 2020). Although almost all fossils point to fully terrestrial forms, two aquatic vertebrates are present: a fragmentary interclavicle of a Stereospondyli (Dias-da-Silva et al. 2009) and part of a snout of an indeterminate phytosaur (Kischlat and Lucas 2003). On the other hand, except for putative records of traversodontids (Ribeiro et al. 2011) plus the records mentioned above (pending further geochronological studies), the confident fossil record of cynodonts is exclusively of small prozostrodonts, which include the “ictidosaur” Riograndia guaibensis, and the tritheledontids Irajatherium hernandezi, Botucaraitherium belarminoi, and Brasilodon quadrangularis, the sister-group to Mammaliaformes (Bonaparte et al. 2003, 2005; Martinelli et al. 2005, 2017c; Soares et al. 2011, 2014; Kerber et al. 2022). These forms are important for understanding the origin and early evolution of Mammaliaformes and, consequently, provide evidence for the reconstruction of mammalian history.
Dinosaurs were also present in the Riograndia AZ strata, albeit were less diversified than in the older Hyperodapedon AZ. Three dinosaur taxa are currently known, two of them belonging to the Sauropodomorpha (Macrocollum itaquii and Unaysaurus tolentinoi) (Leal et al. 2004; Muller et al. 2018), while the third species (Guaibasaurus candelariensis) has disputed affinities (Bonaparte et al. 1999; Langer et al. 2010). Other Riograndia AZ archosauromorphs include the pterosauromorphs Faxinalipterus minima and Maehary bonapartei (Bonaparte et al. 2010; Kellner et al. 2022) and the dinosaur-like silesaurid Sacisaurus agudoensis (Ferigolo and Langer 2021).
The Triassic period has witnessed one of the main vegetation transitions in the Earth’s history, with environments dominated by the Dicroidium Flora gradually replacing the Glossopteris Flora, abundant during the Permian. The Dicroidium Flora was composed of plants better adapted to the new climatic conditions, significantly warmer and drier than in the previous period. Among them are the Corystospermales (Xylopteris, Dicroidium, and Jonstonia), which, despite being morphologically very similar to ferns (pteridophytes), produced seeds (spermatophytes) as the gymnosperms (Guerra-Sommer et al. 1999; Pankaj et al. 2014).
In RS deposits, remarkable fossiliferous assemblages that reveal the taphoflora, mainly of conifers of the group Araucarioxylon, are associated with the last fossil records of Triassic tetrapods. The herbivores at the time probably fed on shrubs and small arboreal plants of the Dicroidium Flora. The changes that occurred in the composition of the flora due to lower rainfall and higher temperatures are pointed out as one of the reasons for the substitution of the cynodont and dicynodont fauna, which did not adapt to the new vegetation, thus leaving room for the domain of rhynchosaurs and first dinosaurs (Guerra-Sommer et al. 1998; Guerra-Sommer and Cazzulo-Klepzig 2007; Langer et al. 2007; Schultz and Langer 2007; Horn et al. 2014).
The Triassic plains in western Gondwana (where southern Brazil is located today) were filled not only by plant groups that are now extinct, such as Corystospermales, but also by plants that still have extant representatives, e.g., the phyllicophytes (Todites, Clapophlebis, and Asterotheca), ginkgophytes (Ginkoites, Baiera, and Stenorachis), sphenophytes (Neocalamites), conifers (Araucarioxylon, Podozamites, and Rissikia), and cycads (Pseudoctenisi, Otozamites, and Taeniopteris). This taphoflora is found in the fossil record through the impression of fronds, leaves, and stems in clayey sediments and silicified stems in clayey and sandy sediments (e.g., Guerra-Sommer et al. 1999; Da Rosa et al. 2009a, b; Barboni and Dutra 2015). These distinct taphonomic and depositional processes have been the subject of debate up to the present. The so-called Dicroidium Flora, preserved through normally articulated impressions, is associated with a low-energy depositional environment. On the other hand, the Coniferous Flora, or “Araucarioxylon Flora”, is represented by phytofossils of silicified stems associated with high energy deposition. The Coniferous Flora was composed of floristic elements with overall small leaves, such as the Araucariaceae, Taxodiaceae, and Cheiropodiaceae. The difficulty in performing the biostratigraphy insertion of this palaeoflora results from the fact that the fossiliferous record is rarely found in situ. However, Guerra-Sommer et al. (1999) suggest that the Dicroidium Flora and the Coniferous Flora preserved in RS deposits were contemporary floristic communities and belonged to the same depositional system, but preserved in different faciologies. The Dicroidium Flora is related to a depositional system proximal to the fluvial plain, due to its hygromesophilic features, and the Araucarioxylon Flora, with mesoxerophilic features, is associated with distal deposits of the flooding plain.
Regarding the conifer taphoflora in the Triassic deposits of RS, there are very few records of leaf branches of Araucariaceae (Crisafulli and Dutra 2009; Barboni and Dutra 2013), and all other records of this conifer flora are thus far only constituted by silicified wood, with no other records of associated macro- or microfossils (Horn et al. 2014). This assembly of silicified woods of Araucarioxylon preserved in conglomeratic sandstones that characterize the Mata Sequence represents one of the most important paleobotanical sites in South America and, although its deposition is evidently associated with high-energy fluvial events, being an allochthonous taphocoenosis, the analysis of the anatomical structures of these woods corroborates a climate with low seasonality as inferred for the Triassic in these latitudes (Pires and Guerra-Sommer 2004; Pires et al. 2005; Dos Santos et al. 2023).
3.3 Juro-Cretaceous
The Brazilian Jurassic is predominantly represented by aeolian sedimentary units, so the scarcity of body fossils is remarkable. As such, most of what we know about Jurassic faunas that thrived in southern Brazil comes from ichnological evidence – especially footprints and burrows (e.g., Dentzien-Dias et al. 2007, 2012a, b; Francischini et al. 2015, 2017). These have been recovered from the Guará Formation (Upper Jurassic), a geographically restricted unit with a predominance of aeolian and fluvial deposition (Scherer and Lavina 2005). The Guará Formation crops out in the southwestern portion of Rio Grande do Sul, with aeolian exposures concentrated toward the south (Scherer and Lavina 2005). However, it is noteworthy that the Guará Formation has a lateral correlation with the Uruguayan Tacuarembó Formation, from which a large number of somatic (i.e., body fossils) occurrences have already been reported (e.g., Perea et al. 2009; Soto et al. 2021).
Dinosaurs are the main components of the Guará Formation ichnoassemblage. Their footprints are found in both sand dune and sand sheet deposits (Dentzien-Dias et al. 2012a, b), especially within the boundaries of Santana do Livramento and Rosário do Sul municipalities. The first Guará Formation dinosaur footprints were formally reported by Scherer and Lavina (2005). Subsequently, Dentzien-Dias et al. (2007) conducted the first detailed assessment of dinosaur tracks and small-sized vertebrate burrows. These authors recognized a diverse assemblage composed of sauropods, theropods, and ornithopods. In a later contribution, Dentzien-Dias et al. (2012a, b) quantified Guará Formation occurrences, recognizing the presence of 60 dinosaur tracks. Footprints from which a conclusive identification can be made reveal an ichnoassemblage dominated by sauropods (n = 22) and theropods (n = 15), with a smaller contribution of ornithopods (n = 6).
The most recent contribution to the knowledge of the southern Brazilian Late Jurassic faunas was the recognition by Francischini et al. (2017) of the presence of ankylosaur tracks in a Guará Formation outcrop. Besides the scarcity of ankylosaur fossils in South America, the Guará Formation occurrence represents the oldest record of this clade in the western portion of Gondwana. The discovery of this new and unexpected ichnotaxon highlights the underexplored potential of the Guará Formation in revealing clues from the elusive Brazilian Jurassic faunas.
3.4 Pleistocene
The fossil record of Pleistocene tetrapods, especially mammals, shows a mixture of taxa from lineages with different biogeographic origins: mainly forms that evolved in South America during the Paleogene (in addition to caviomorphs, of African origin) and Neogene, and tetrapods that immigrated from the northern hemisphere (Central and North America) during the Great American Biotic Interchange (GABI) (see Woodburne 2010; Goin et al. 2012; Forasiepi et al. 2014; Carrillo et al. 2015; O’Dea et al. 2016). This biogeographic event, which is perhaps primarily responsible for the emergence of the modern fauna of the South American continent (Carrillo et al. 2015), was the biotic interchange between South America and North and Central America. It began during the Late Miocene (~10–7 Ma), with the arrival of the first immigrants (i.e., procyonids and cricetid rodents, also called “island hoppers”, in reference to the fact that the isthmus was not fully formed), and was intensified between the end of the Pliocene (2.7 million years) and the Pleistocene (Woodburne 2010; Goin et al. 2012; O’Dea et al. 2016). Before the Pliocene, terrestrial organisms could not pass freely between these continents because the Isthmus of Panama had not yet been fully formed. As a result, South America was virtually isolated from the northern hemisphere by land routes during part of the Cenozoic. After the formation of this terrestrial connection, several organisms migrated from one region to another, significantly modifying the biotic composition of the continent’s ecosystems. This mixture of organisms resulted in the so-called South America Pleistocene Megafauna (Fig. 3.4), characterized by the presence of several large-sized mammals (>300 kg), such as giant sloths, glyptodonts, pampatheres, toxodonts, macrauchenias, horses, cervids, large carnivores, and mastodonts, among others. Apart from a few exceptions, megamammals disappeared at the end of the Pleistocene. So far, all mammals found in RS are placentals, as marsupials are still unknown in the fossil record. These placentals represent four large groups: Xenarthra, Laurasiatheria, Euarchontoglires, and Afrotheria (Asher et al. 2009). Some iconic fossils from the Pleistocene Megafauna are shown in Fig. 3.4.
Concerning autochthonous South American mammals present in the Pleistocene of RS, a first group is Xenarthra the only placental lineage with extant species originating in South America. This is one of the most emblematic groups of the Cenozoic of this continent because of their great taxonomic diversity, abundant fossil record, and endemism (Gaudin and Croft 2015). During the Pleistocene of RS, xenarthrans were represented by two groups: pilosans (ground sloths) and cingulates (glyptodonts, armadillos, and pampatheres). Both megatheriid and mylodontid ground sloths have been found in RS Pleistocene deposits (Oliveira 1992, 1996). Megatheriid records include the co-occurrence of Megatherium americanum and Eremotherium laurillardi in at least two localities: Arroio Seival, Caçapava do Sul (Oliveira et al. 2002) and Chui Creek, Santa Vitória do Palmar (Lopes et al. 2019). This is the only Brazilian territory with records of both megatheriids, because M. americanum had an austral distribution and E. laurillardi had an intertropical distribution. Hence, RS was possibly a geographic boundary in the distribution of both species. The large mylodontid Lestodon armatus was reported from Caçapava do Sul and Chuí Creek (Vargas-Peixoto et al. 2021). Other ground sloths from the coastal plain of RS (CPRS) include the mylodontids Mylodon darwinii, the only record of this species in Brazil, Glossotherium robustum, and the scelidotheriine mylodontid Catonyx cf. C. cuvieri (Oliveira 1992; Lopes and Pereira 2010; Pitana et al. 2013). In western RS (WRS), ground sloths are rarer than in CPRS. Only records of the mylodontid Glossotherium robustum have been confidently reported (Pitana et al. 2013). Cingulates, especially glyptodonts, are common in RS Pleistocene deposits. However, most records are based on isolated osteoderms. The glyptodonts Glyptodon clavipes and Panochthus are present in most fossiliferous assemblages (Kerber and Oliveira 2008a; Pereira et al. 2012; Kerber et al. 2014a, b). Recently, large portions of carapaces, probably of G. clavipes, were found in the Ibicuí River, Alegrete, and Jaguarão, but the material is still undescribed (LK, pers. obs.). Panochthus from CPRS is represented by two species: P. tuberculatus and Panochthus cf. P. greslebini (Ferreira et al. 2015). Doedicurus is present only in deposits of the CPRS (Pereira et al. 2012), and rare material of Neuryurus was recorded from Garupá Creek, Quaraí (Kerber and Oliveira 2008b). The record of the enigmatic cingulate Pachyarmatherium is only based on isolated osteoderms from Balneário Hermenegildo (Lopes and Buchmann 2010). The pampatheres Pampatherium humboldti and Holmesina paulacoutoi have been documented from several Pleistocene deposits (Oliveira and Pereira 2009; Ferreira et al. 2018), although the species of Pampatherium of WRS is unknown (Kerber et al. 2014a, b). Dasypodids are the rarest cingulates, probably because of their smaller size, which makes it difficult to find their fossils. Propraopus grandis fossils have been mentioned for several localities (Pitana and Ribeiro 2007) and Dasypus sp. for the Chuí Creek (Oliveira and Pereira 2009).
Other iconic Cenozoic South American mammals are the native ungulates (Croft et al. 2020). These animals, possibly related to perissodactyls (horses, tapirs, rhinos, etc.; Laurasiatheria) (Buckley 2015; Croft et al. 2020), have five major lineages. The native ungulates from the Pleistocene of RS belong to two groups: notoungulates (toxodontids) and litopterns (protherotheriids and macraucheniids). Toxodontids are large herbivores, hippo-like mammals, and are represented by Toxodon platensis found in most Pleistocene localities of RS (Pereira et al. 2012; Kerber et al. 2014a, b). Proterotheriids are litopterns with more plesiomorphic morphological traits and are smaller than macraucheniids, which are characterized by elongated necks and nasal retraction. Fossils of the proterotheriid cf. Neolicaphrium recens, the only Quaternary survivor of the lineage (Bond et al. 2001), were found in the Hermenegildo Beach (Santa Vitória do Palmar) (Scherer et al. 2009). Like T. platensis, the macraucheniid Macrauchenia patachonica has also been found in several localities (Scherer et al. 2009).

Late Pleistocene landscape of Rio Grande do Sul, showing from left to right the glyptodont Glyptodon clavipes, the ursid Arctotherium, the toxodontid Toxodon platensis (behind), and the megatheriid Megatherium americanum. The predominance of herbaceous vegetation is also evident, typical of parts of the landscape with a less significant presence of Araucaria Forest. (Artwork Julio Lacerda)
Euarchontoglires is represented by rodents. Two groups are found in Pleistocene sediments of RS: caviomorphs (African origin) and cricetids (Holartic origin). Together, both lineages compose the most diversified lineage of South American mammals (D’Elía et al. 2019). The Pleistocene record of caviomorphs includes the capybara (Hydrochoerus hydrochaeris) and the coypu (Myocastor coypus) in deposits from WRS and CPRS (Kerber and Ribeiro 2011; Kerber et al. 2014a, b). A fossil of the cavy Galea sp. (absent in the RS modern mastofauna) was found in deposits of the Touro Passo Creek (Kerber et al. 2011b). Similarly, in the fossiliferous assemblage from the Chuí Creek, three rodents that currently inhabit drier areas of southern South America have been found: the cavy Microcavia, the vizcacha Lagostomus cf. maximus, and the mara Dolichotinae indet. (Ubilla et al. 2008; Kerber et al. 2011c). The fossil record of cricetids is scarce, represented by remains of Reithrodon auritus from Balneário Hermenegildo (Rodrigues and Ferigolo 2004) and Holochilus brasiliensis from WRS and CPRS (Kerber et al. 2012).
Like cricetids, other mammalian lineages recorded in Pleistocene deposits of RS have a Holarctic origin, and their ancestral lineages arrived during the Plio-Pleistocene GABI (see Woodburne 2010; Goin et al. 2012). Most of them had wide geographic distributions in the South American Pleistocene. Artiodactyla is the most diverse clade of laurasiatherians, with three groups: tayassuids, camelids, and cervids. Tayassuids (peccaries) show records of the extant white-lipped peccary Tayassu pecari at Touro Passo Creek, and of the large extinct Brasiliochoerus stenocephalus in WRS as well as in CPRS (Gasparini et al. 2009; Kerber et al. 2014a, b; Copetti et al. 2021). Camelids, the llamas and guanacos, are represented by three species in several localities: the extant guanaco Lama guanicoe and the vicuña L. aff. gracilis, and extinct Hemiauchenia paradoxa (Scherer 2006; Scherer et al. 2007). Cervids (deers) have records of indeterminate species of the extant genus Mazama in the CPRS, and the extinct genus Morenelaphus, and the large Antifer in several localities (Kerber et al. 2014a, b; Pereira et al. 2012; Fontoura et al. 2020).
Perissodactyls include two species of horses, Equus neogeus and Hippidion principale, and the tapir Tapirus cf. terrestris, which were widely distributed (Pereira et al. 2012; Holanda et al. 2012; Kerber et al. 2014a, b). The fossil record of carnivorans is rare, and most fossils come from the CPRS. Few specimens of the canid Dusicyon cf. D. avus, and the large hypercarnivore forms Protocyon troglodytes and cf. Theriodictis are recorded from the CPRS, as well as the iconic saber-toothed cat Smilodon popularis and the jaguar Panthera onca (Rodrigues et al. 2004; Lopes et al. 2020). Ursids are known from undescribed bones from Quaraí River, an isolated tooth from Pantano Grande, and a canine assigned to cf. Arctotherium from Chuí Creek (Pereira et al. 2012). Afrotherians are represented by proboscids, with records of mastodont Notiomastodon platensis from several localities (Marcon 2008).
Other continental tetrapods are much less abundant and studied. From the Touro Passo Creek, there are some records of chelonians, not described in detail, such as Hydromedusa tectifera (Maciel et al. 1996; Hsiou 2009), the endemic extinct teiid lizard Tupinambis uruguaianensis (Hsiou 2007), and the tropical stork Mycteria cf. M. americana (Ribeiro et al. 1995; Hsiou 2009). Finally, the tropical stork Ciconia is recorded from Chuí Creek (Lopes et al. 2019).
Due to the geographic location of RS, the Pleistocene fossils are in the geographic boundary between two biogeographic regions, the Pampean and Intertropical regions (Oliveira 1999; Oliveira et al. 2002; Oliveira and Pereira 2009; Kerber et al. 2014a, b; Lopes et al. 2019, 2020). For this reason, their taxonomic composition show a mix of taxa predominantly recorded in the southernmost parts of South America (e.g., Doedicurus and Macrauchenia) and forms of intertropical affinities (e.g., Eremotherium). Most large mammals were extinct at the end of the Pleistocene and early Holocene. During the Holocene, the taxonomic composition of the modern faunas originated (see Hadler et al. 2008, 2009, 2016; Lopes et al. 2020 and references therein for a review). Although the causes of this extinction event of continental proportions are still debated, it is inferred that the extinctions were not simultaneous, but occurred over the course of a few thousand years. (Lopes et al. 2020). According to the model proposed by Lopes et al. (2020), the most recent study on RS extinctions, large mammals disappeared in at least two phases, which can be related to climatic changes in the Late Pleistocene. The first phase occurred around 30 ka in the CPRS, predating the arrival of humans into the region by about 25 ka. A second phase eliminated taxa that survived until about 12.7 ka in reduced suitable environments associated with perennial rivers of the WRS. This phase coincides with the transition from glacial to interglacial conditions. Finally, it was suggested that early human occupations may have contributed to environmental modifications. No confident archaeological records of humans’ coexistence or exploitation of extinct mammals have been documented in RS yet, although there are plenty of them in South America (Bampi et al. 2022). However, early inhabitants may have contributed to environmental changes through the use of fire for agricultural purposes (Behling et al. 2009).
It is clear that it was the climatic variation over time that outlined the environmental mosaic of the Campos Sulinos region. Different studies confirm that grassland vegetation is much older than forests in the Pleistocene phytogeography of southern Brazil. During much of the Pleistocene, the landscape that made up these environments was predominantly grassland vegetation, with rivers not accompanied by riparian forests. These open areas were populated by megafauna and had a humid and cold climate for most of the year (Scherer and Da Rosa 2003; Behling et al. 2009). The presence of megafauna also played an important role in vegetation dynamics, and herbivory action guided the process of plant evolution and their morphological patterns. Studies carried out in savannah areas of South America and Africa show that the vegetation coexisting with the past megafauna developed specific protection mechanisms, such as thorns and smaller and rigid leaves, including the production of unpalatable chemical substances to minimize predation (Dantas and Pausas 2022). Thus, large herbivorous mammals also maintained grassland vegetation for a long time during the Pleistocene. After the extinction of the megafauna, an opportunity for the expansion of forest was created (Behling et al. 2009; Dantas and Pausas 2022). The development of vegetation cover in southern Brazil can be traced from the palynological record of peats recovered in different locations in the southern states of Brazil: Paraná (Behling 1997), Santa Catarina (Behling 1995), and Rio Grande do Sul (Roth and Lorscheitter 1993; Behling et al. 2001, 2004), which abundantly preserve pollen from the Pleistocene, including species of Poaceae, Cyperaceae, and Asteraceae, all typical of grassland communities. On the other hand, the presence of forests in the region, especially the Araucaria Forest, and their expansion over grasslands vegetation is only verified from 3 ka AP on, associated with the increase and frequency of rainfall. At first, forests were restricted to areas close to water bodies; later, they advanced over grassland vegetation in areas farther away (Behling et al. 2009). In addition, the presence of charcoal in palynological slides (e.g., Behling et al. 1995, 2005; Ribeiro et al. 2020) is evidence of paleowildfires during the Pleistocene in the Campos Sulinos (Behling et al. 2009). Fire is one of the key modeling elements of past ecosystems (e.g., Scott and Stea 2002; Scott and Damblon 2010; Manfroi et al. 2023), and can be compared to herbivory as a modifier of different biomes over time (Bond and Keeley 2005). In the Campos Sulinos, we thus conclude that both fire and herbivory were instrumental in restricting forest expansion over grasslands during much of the Pleistocene (see also Behling et al. 2023, Chap. 4, in this volume).
4 Summary and Conclusions
4.1 Earlier Ecosystems
Permian ecosystems are directly related to Pangean conditions, ranging from glacial to desert environments, including epicontinental marine forms. In these scenarios, the fossils from early Permian (Sakmarian-Artinskian) of southern Brazil record several ecosystems, from glacial and postglacial highlands and lowlands covered with shrubs and forests, to coastal lagoons and sandy barriers coupled with peat environments, and recurrent wildfires. Middle Permian deposits only record an epicontinental sea and its coastal environments, but no continental succession is known so far. The late Permian sediments record the continentalization of Gondwana, and the formation of complex food chains of amniotes and temnospondyls in flat alluvial plains covered with shrubs and small forests, also subject to wildfires.
Triassic ecosystems also relate to Pangean conditions, but resulted from environmental and biotic recovery after the most massive extinction event, and later the continental breakup forming Laurasia and Gondwana. The early Triassic deposits (Olenekian) gave rise to alluvial plain ecosystems, with fish, temnospondyls, and archosauriforms, in a food chain not yet detailed. Middle and Late Triassic deposits can be related to two different fluvial scenarios: low energy, high sinuosity rivers, and related floodplain environments (Ladinian and Carnian), and high energy rivers with restricted overbank deposits (Norian). There is a dominance of archosaurs and synapsids (cynodonts and dicynodonts) at first, in Dinodontosaurus and Santacruzodon Assemblage Zones (AZ’s), followed by the disappearance of dicynodonts or their substitution by specialized herbivores and the irradiation of more agile carnivores, respectively, rhynchosaurs and dinosaurs, in Hyperodapedon AZ. The Riograndia AZ records the return of dicynodonts, but in ecosystems mainly occupied by cynodonts and sauropodomorph dinosaurs as main herbivores, and a plethora of carnivores, such as cynodonts, dinosaurs, and archosaurs. The twofold scenario is also recorded by the presence of shrubby plants of the Dicroidium Flora that are later replaced by the Conifer Flora, more related to forests.
Late Jurassic land faunas are known only by their ichnological remains, found in association with typically aeolian deposits, a result of arid desertic environments. Despite that, the yet poorly known fossil record of this timespan reveals the presence of an association of medium to large-sized herbivore and carnivore dinosaurs. More data on southern Brazil Jurassic faunas are needed in order to better understand the ecological dynamics of ecosystems.
4.2 Pleistocene and Modern Ecosystems
The present relief of southern Brazil is a post-Cretaceous construction that formed after the breakup of South America and Africa. Most rivers started flowing inward and later toward the Atlantic Ocean, forming the coastal plain and continental platform (Potter 1997). The result of fluvial erosion is recorded in the Pelotas Basin’s Cretaceous, Paleogene, and Neogene succession (Anjos-Zerfass et al. 2008).
Along the last 400 ka, the intercalation of glacial and interglacial cycles led to the formation of a mosaic of forests and grasslands, each cycle favoring a specific environment and associated flora and fauna. The onset of interglacial cycles led to sea level rise, and the continental onward shift of the coastline, preserving lagoons and sandy barriers at the coastal plain of Rio Grande do Sul. In contrast, the glacial cycles shifted the same coastline basinward, i.e., at least 100 km from the present shoreline, depositing fluvial sediments along the submerged continental platform (Lopes et al. 2021). On the continental side, the glacial cycles reinforced fluvial erosion, due to river profile lowering. In contrast, the interglacial cycles favored fluvial deposition, through progressive drowning of the river profile (see references in Maahs et al. 2019).
Some ecological processes are considered important in shaping the Campos Sulinos landscapes, both in the past and today. The action of fire on vegetation, for example, is recognized as an important controller in the maintenance of ecosystems, and evidence of fire exist especially in the more recent past (the last 600 years), despite humid climatic conditions during this period. Likely, occurrence of fire is related to human populations, did not allow for widespread development of forests and keeping grassland vegetation as the predominant system (e.g., Behling et al. 2009; Piperno and Becker 1996; see also Behling et al. 2023, Chap. 4 in this volume). Along with fire, the influence of herbivore animals is likely equally important in the dynamics of the Campos Sulinos landscapes. Whether large herbivore mammals that inhabited the region until 8500 years ago (Scherer and Da Rosa 2003) or cattle introduced by the Jesuits in the seventeenth century (Porto 1954), herbivores consume plants and prevent the expansion of woody vegetation, while also limiting the accumulation of biomass and thus preventing the availability of fuel for larger fires (Behling et al. 2009). Clearly, human occupation has had an influence on natural environments from their earliest beginnings.
The fossil record of the region known today as the Campos Sulinos is extremely rich and dates back to 300 million years ago. These fossils testify important paleogeographic, paleoclimatic, and paleofaunistic shifts, and provide evidence for the origin and evolution of the different ecosystems. It is impossible to study the region’s biodiversity without considering its past, including biotic and abiotic events that occurred during the Cenozoic and probably acted as shaping agents of environments and current biodiversity.
References
Abdala F, Ribeiro AM (2000) A new therioherpetid cynodont from the Santa Maria Formation (middle Late Triassic), southern Brazil. Geodiversitas 22(4):589–596
Abdala F, Ribeiro AM (2003) A new traversodontid cynodont from the Santa Maria Formation (Ladinian-Carnian) of southern Brazil, with a phylogenetic analysis of Gondwanan traversodontids. Zool J Linnean Soc 139:529–545
Abdala F, Sá-Teixeira AM (2004) A traversodontid cynodont of African affinity in the South American Triassic. Palaeontol Afr 40:11–22
Abdala F, Barberena MC, Dornelles J (2002) A new species of the traversodontid cynodont Exaeretodon from the Santa Maria Formation (Middle/Late Triassic) of southern Brazil. J Vertebr Paleontol 22(2):313–325
Anjos-Zerfass GS, Souza PA, Chemale FJ (2008) Biocronoestratigrafia da Bacia de Pelotas: estado atual e aplicação na geologia do petróleo. Rev Bras Geociênc 38(2):47–62
Araújo DC, Gonzaga TD (1980) Uma nova espécie de Jachaleria (Therapsida, Dicynodontia) do Triássico do Brasil. In: Actas del II Congreso Argentino de Paleontología y Bioestratigrafía. I Congreso Latinoamericano de Paleontología, Buenos Aires, pp 159–174
Araújo-Barberena DC, Lacerda FJV, Timm LL (2002) Mesossauro da Serra do Caiapó (Montividiu), GO: Um vertebrado fóssil típico do Paleozóico Superior, importante na história da Deriva Continental. In: Schobbenhaus C, Campos DA, Queiroz ET, Winge M, Berbert-Born MLC (eds) Sítios Geológicos e Paleontológicos do Brasil. DNPM/CPRM – Comissão Brasileira de Sítios Geológicos e Paleobiológicos (SIGEP), Brasília, pp 81–85
Asher RJ, Bennett N, Lehmann T (2009) A new framework for understanding placental mammal evolution. BioEssays 31:853–864
Azevedo KL, Vega CS, Soares MB (2017) A new specimen of Australerpeton cosgriffi Barberena 1998 (Stereospondyli: Rhinesuchidae) from the Middle/Upper Permian Rio do Rasto Formation, Paraná Basin, Brazil. Rev Bras Paleontol 20:333–344
Bampi H, Barberi M, Lima-Ribeiro MS (2022) Megafauna kill sites in South America: a critical review. Quat Sci Rev 298:107851
Barberena MC (1982) Uma nova espécie de Proterochampsa (P. nodosa sp. nov.) do Triássico do Brasil. An Acad Bras Ciênc 54:127–141
Barberena MC, Araújo DC (1975) Tetrápodos fósiles de Sudamérica y deriva continental. In: Congreso Argentino de Paleontologia y Bioestratigrafia, Tucumán, pp 497–504
Barberena MC, Bonaparte JF, Teixeira AMSA (1987) Thrinaxodon brasiliensis. sp. nov. a primeira ocorrência de cinodontes galessauros para o Triássico do Rio Grande do Sul. In: Congresso Brasileiro de Paleontologia, Rio de Janeiro, vol 10, pp 67–76
Barboni R, Dutra TL (2013) New “flower” and leaves of Bennettitales from Southern Brazil and their implication in the age of the Lower Mesozoic deposits. Ameghiniana 50:14–32
Barboni R, Dutra TL (2015) First record of Ginkgo-related fertile organs (Hamshawvia, Stachyopitys) and leaves (Baiera, Sphenobaiera) in the Triassic of Brazil, Santa Maria Formation. J S Am Earth Sci 63:417–435
Behling H (1995) Investigations into the Late Pleistocene and Holocene history of vegetation and climate in Santa Catarina (S Brazil). Veg Hist Archaeobotany 4:127–152
Behling H (1997) Late Quaternary vegetation, climate and fire history in the Araucaria forest and campos region from Serra Campos Gerais (Paraná), S Brazil. Rev Palaeobot Palynol 97:109–121
Behling H, Bauermann SG, Neves PCP (2001) Holocene environmental changes in the São Francisco de Paula region, southern Brazil. J S Am Earth Sci 14:631–639
Behling H, Pillar VD, Orlóci L, Bauermann SG (2004) Late Quaternary Araucaria forest, grassland (Campos), fire and climate dynamics, studied by high-resolution pollen, charcoal and multivariate analysis of the Cambará do Sul core in southern Brazil. Palaeogeogr Palaeoclimatol Palaeoecol 203:277–297
Behling H, Pillar VD, Bauermann, SG (2005) Late Quaternary grassland (Campos), gallery forest, fire and climate dynamics, studied by pollen, charcoal and multivariate analysis of the São Francisco de Assis core in western Rio Grande do Sul (southern Brazil). Review of Palaeobotany and Palynology 133:235–248.
Behling H, Jeske-Pieruschka V, Schüler L, Pillar VD (2009) Dinâmica dos campos no sul do Brasil durante o Quaternário Tardio. In: Pillar VD, Müller SC, Castilhos ZMS, Jacques AVA (eds) Campos Sulinos: Conservação e Uso Sustentável da Biodiversidade, Brasília, pp 13–25
Behling H, Overbeck GE, Pillar VD (2023) Dynamics of South Brazilian grasslands during the Late Quaternary. In: Overbeck GE, Pillar VDP, Müller SC, Bencke GA (eds) South Brazilian grasslands: ecology and conservation of the Campos Sulinos (this volume). Springer, Cham
Bergue CT, Maranhão MSAS, Ng C (2020) The Permian podocopids (Crustacea: Ostracoda) from the Serra Alta and Teresina formations, Paraná Basin, Brazil. Micropaleontology 66(4):301–316
Bombin M (1976) Modelo paleoecológico evolutivo para o Neoquaternário da região da Campanha-Oeste do Rio Grande do Sul (Brasil). A Formaçao Touro Passo, seu conteúdo fossilífero e a pedogênese pós-deposicional: Comunicações do Museu de Ciências da PUCRS 15:1–90
Bonaparte JF, Ferigolo J, Ribeiro AM (1999) A new early Late Triassic saurischian dinosaur from Rio Grande do Sul State, Brazil. Proceedings of the second Gondwanan Dinosaurs symposium. Nat Sci Mus Monogr Tokyo 15:89–109
Bonaparte JF, Martinelli AG, Schultz CL, Rubert R (2003) The sister group of mammals: small cynodonts from the late Triassic of southern Brazil. Rev Bras Palaontol 5:5–27
Bonaparte JF, Martinelli AG, Schultz CL (2005) New information on Brasilodon and Brasilitherium (Cynodontia, Probainognathia) from the late Triassic of southern Brazil. Rev Bras Palaontol 8:25–46
Bonaparte JF, Soares MB, Schultz CL (2006) A new non-mammalian cynodont from the Middle Triassic of southern Brazil and its implications for the ancestry of mammals. Bull N M Mus Nat Hist Sci 37:599–607
Bonaparte JF, Schultz CL, Soares MB (2010) Pterosauria from the Late Triassic of southern Brazil. In: Bandyopadhyay S (ed) New aspects of mesozoic biodiversity. Lect Notes Earth Syst Sci 132:63–71
Bond WJ, Keeley JE (2005) Fire as a global “herbivore”: the ecology and evolution of flammable ecosystems. Trends Ecol Evol 20(7):387–394
Bond M, Perea D, Ubilla M, Tauber AA (2001) Neolicaphrium recens Frenguelli 1921, the only surviving Proterotheriidae (Litopterna, Mammalia) into the South American Pleistocene. Palaeovertebrata 30(1–2):37–50
Boos ADS, Schultz CL, Vega CS, Aumond JJ (2013) On the presence of the Late Permian dicynodont Endothiodon in Brazil. Palaeontology 56:837–848
Boos ADS, Kammerer CF, Schultz CL, Paes Neto VD (2015) A tapinocephalid dinocephalian (Synapsida, Therapsida) from the Rio do Rasto Formation (Paraná Basin, Brazil): taxonomic, ontogenetic and biostratigraphic considerations. J S Am Earth Sci 63:375–384
Boos ADS, Kammerer CF, Schultz CL, Soares MB, Ilha ALR (2016) A new dicynodont (Therapsida: Anomodontia) from the Permian of southern Brazil and its implications for bidentalian origins. PLoS One 11:e0155000
Bortoluzzi CA (1975) Étude de quelques empreintes de la flore gondwanienne du Brésil. In: Cong Nat Soc Savantes, Actes, France, pp 171–187
Brust BAC, Desojo JB, Schultz CL, Paes-Neto VD, Da Rosa AAS (2018) Osteology of the first skull of Aetosauroides scagliai Casamiquela 1960 (Archosauria: Aetosauria) from the Upper Triassic of southern Brazil (Hyperodapedon Assemblage Zone) and its phylogenetic importance. PLoS One 13:e0201450
Buckley M (2015) Ancient collagen reveals evolutionary history of the endemic South American ‘ungulates’. Proc R Soc B 282:20142671
Cabreira SF, Schultz CL, Bittencourt JS, Soares MB, Fortier DC, Silva LR, Langer MC (2011) New stem-sauropodomorph (Dinosauria, Saurischia) from the Triassic of Brazil. Naturwissenschaften 98:1035–1040
Cabreira SF, Kellner AWA, Dias-Da-Silva S, Silva RL, Bronzati M, Marsola J, Almeida C, Müller RT, Bittencourt JS, Batista B, Armando J, Raugust T, Carrilho R, Brodt A, Langer MC (2016) A Unique Late Triassic Dinosauromorph assemblage reveals dinosaur ancestral anatomy and diet. Curr Biol 26:3090–3095
Callefo F, Ricardi-Branco F, Galante D, Rodrigues F (2015) Esteiras microbianas da Formação Teresina, em Taguaí (SP) Grupo Passa Dois, Bacia do Paraná. Terræ 12(1–2):27–35
Carrillo JD, Forasiepi A, Jaramillo C, Sánchez-Villagra MR (2015) Neotropical mammal diversity and the Great American Biotic Interchange: spatial and temporal variation in South America’s fossil record. Front Genet 5:1–11
Castro MRD, Perinotto JADJ, Castro JCD (1999) Fácies, análise estratigráfica e evolução pós-glacial do membro Triunfo/Formação Rio Bonito, na faixa subaflorante do norte catarinense. Rev Bras Geociênc 29(4):533–538
Cazzulo-Klepzig MC (1978) Estudo da tafoflora do Membro Morro Pelado na sua localidade-tipo. Pesquisas 11:2253–2203
Cazzulo-Klepzig MC, Correia NR (1981) Contribuição ao estudo da Tafoflora Permiana no membro Serrinha na Serra do Cadeado, Estado do Paraná, Brasil. Congresso Latinoamericano de Paleontologia 1:233–247
Cerri RI, Luvizotto GL, Tognoli FMW, Warren LV, Okubo J, Morales N (2020) Mechanical stratigraphy and structural control of oil accumulations in fractured carbonates of the Irati Formation, Paraná Basin, Brazil. Braz J Geol 50:1–12
Chahud A, Pacheco MLAF, Meira FVE, Romero GR, Petri S (2012) Paleontology and depositional environments of the Tatuí and Irati formations (Permian) in the Ponte Nova Farm, Ipeúna, state of São Paulo. Rev Bras Geociênc 42(1):1982–1912
Cisneros JC, Abdala F, Rubidbe BS, Dentzien-Dias PC, Bueno AO (2011) Dental occlusion in a 260-million-year-old therapsid with saber canines from the Permian of Brazil. Science 331(6024):1603–1605
Cisneros JC, Abdala F, Atayman-Güven S, Rubidge BS, Celâlsengör AM, Schultz CL (2012) Carnivorous dinocephalian from the Middle Permian of Brazil and tetrapod dispersal in Pangaea. Proc Natl Acad Sci 109(5):1584–1588
Cisneros JC, Marsicano C, Angielczyk KD, Smith RMH, Richter M, Frösbisch J, Kammerer CF, Sadleir RW (2015) New Permian fauna from tropical Gondwana. Nature 6:76–86
Clack JA (2012) The origin and evolution of tetrapods, Gaining ground, 2nd edn. Indiana University Press, Bloomington
Colbert EH (1970) A saurischian dinosaur from the Triassic of Brazil. Am Mus Novit 2405:1–60
Copetti PL, Gasparini GM, Pereira JC, Fontoura E, Lopes RP, Kerber L (2021) A skull of the extinct tayassuid Brasiliochoerus stenocephalus (Lund in Reinhardt, 1880) (Mammalia, Cetartiodactyla) from the Late Pleistocene of southern Brazil: morphology and taxonomy. Hist Biol 33:18981–18910
Costa Santos MA, Paes Neto VD, Schultz CL, Cisneros J, Pierce SE, Pinheiro FL (2023) Cranial osteology of the Brazilian dinocephalian Pampaphoneus biccai (Anteosauridae: Syodontinae). Zool J Linn Soc 199:zlad071. https://doi.org/10.1093/zoolinnean/zlad071
Crisafulli A, Dutra TL (2009) Kaokoxylon zalesskyi (Sahni) Maheswari en los niveles superiores de la Secuencia Santa Maria 3 (Formación Caturrita), Cuenca de Paraná, Brasil. Gaea J Geosci 5:61–69
Croft DC, Gelfo JN, López GM (2020) Splendid innovation: the extinct South American native ungulates. Annu Rev Earth Planet Sci 48(11):1–32
D’Elía G, Fabre PH, Lessa EP (2019) Rodent systematics in an age of discovery: recent advances and prospects. J Mammal 100(3):852–871
Da Rosa AAS (2015) Geological context of the dinosauriform-bearing outcrops from the Triassic of Southern Brazil. J S Am Earth Sci 61:108–119
Da Rosa AAS, Pimentel NLV, Faccini UF (2004a) Paleoalterações e carbonatos em depósitos aluviais na região de Santa Maria, Triássico Médio a Superior do Sul do Brasil. Pesqui em Geociênc 31(1):3–16
Da Rosa AAS, Schwanke C, Cisneros JC, Neto LW, Aurélio PLP, Poitevin M (2004b) “Sítio Cortado” – uma nova assembleia fossilífera para o Triássico Médio do sul do Brasil. Rev Bras Palaontol 7:289–300
Da Rosa AAS, Schwanke C, Aurélio PLP, Poitevin M, Neto LW (2005) Sítio Linha Várzea – uma nova assembleia fossilífera do Triássico Médio do Sul do Brasil. Geociencias 24:115–129
Da Rosa AAS, Guerra-Sommer M, Cazzulo-Klepzig M (2009a) Passo das Tropas, Santa Maria, RS – Marco bioestratigráfico triássico na evolução paleoflorística do Gondwana na Bacia do Paraná. In: Winge M, Schobbenhaus C, Souza CRG, Fernandes ACS, Berbert-Born M, Sallun filho W, Queiroz ET (eds) Sítios Geológicos e Paleontológicos do Brasil. Sítio 084. CPRM, Brasília, pp 332
Da Rosa AAS, Piñeiro G, Dias-da-Silva S, Cisneros JC, Feltrin FF, Neto LW (2009b) Bica São Tomé, um novo sítio fossilífero para o Triássico Inferior do sul do Brasil. Rev Bras Paleontol 12:67–76
Dantas VL, Pausas JG (2022) The legary of the extinct Neotropical megafauna on plants and biomas. Nat Commun 23:129
Davies J, Marzoli A, Bertrand H, Youbi N, Ernesto M, Schaltegger U (2017) End-Triassic mass extinction started by intrusive CAMP activity. Nat Commun 8:15–96
Dentzien-Dias PC, Schultz CL, Scherer CMS, Lavina ELC (2007) The trace fossil record from the Guará Formation (Upper Jurassic?), southern Brazil. Arq do Mus Nac 65:585–600
Dentzien-Dias PC, Figueiredo AEQ, Horn B, Cisneros JC, Schultz CL (2012a) Paleobiology of a unique vertebrate coprolites concentration from Rio do Rasto formation (Middle/Upper Permian), Paraná Basin, Brazil. J S Am Earth Sci 40:53–62
Dentzien-Dias PC, Figueiredo AEQ, Mesa V, Perea D, Schultz CL (2012b). Vertebrate footprints and burrows from the Upper Jurassic of Brazil and Uruguay. In: Netto RG, Carmona NB, Tognoli FMW (eds) Ichnology of Latin America – selected papers. Sociedade Brasileira de Paleontologia, Monografias da Sociedade Brasileira de Paleontologia, Porto Alegre, p 196
De-Oliveira TM, Pinheiro FL, Stock Da-Rosa ÁA, Dias-Da-Silva S, Kerber L (2020) A new archosauromorph from South America provides insights on the early diversification of tanystropheids. PLoS One 15:e0230890
De-Oliveira TM, Kerber L, França MCD, Pinheiro FL (2022) Archosauriform remains from the Lower Triassic Sanga do Cabral Formation of Brazil. J Vertebr Paleontol 41:e2068022
Desojo JB, Ezcurra MD, Kischlat ED (2012) A new aetosaur genus (Archosauria: Pseudosuchia) from the early Late Triassic of southern Brazil. Zootaxa 3166:1–33
Dias EV (2012) A new deep-bodied fóssil fish (Actinopterygii) from the Rio do Rasto Formation, Paraná Basin, Brazil. Zootaxa 3192(1):1–23
Dias EV, Barberena MC (2001) A temnospondyl amphibian from the Rio do Rasto Formation, Upper Permian of Southern Brazil. An Acad Bras Cienc 73(1):135–143
Dias EV, Dias-da-Silva S, Schultz CL (2020) A new short-snouted rhinesuchid from the Permian of southern Brazil. Rev Bras Paleontol 23(2):98–122
Dias-da-Silva S, Dias EV, Schultz CL (2009) First record of stereospondyls (Tetrapoda, Temnospondyli) in the Upper Triassic of Southern Brazil. Gondwana Res 15(1):131–136
Dos Santos ACS, Guerra-Sommer M, Barboza EG, Degani-Schmidt I, Siegloch AM, Vieira CEL, Vieira DT, Bardola TP, Schultz CL (2023) Stressing environmental conditions in the “petrified forest” from the mata sequence in the triassic context of the Paraná basin. J S Am Earth Sci. in press
Eltink E; Da-Rosa AA, Stock ; Dias-Da-Silva, S (2016) A capitosauroid from the Lower Triassic of South America (Sanga do Cabral Supersequence: Paraná Basin), its phylogenetic relationships and biostratigraphic implications. Historical Biology 29:1–12. http://doi.org/10.1080/08912963.2016.1255736
Ezcurra MD, Desojo JB, Rauhut OWM (2015) Redescription and phylogenetic relationships of the proterochampsid Rhadinosuchus gracilis (Diapsida: Archosauriformes) from the early Late Triassic of southern Brazil. Ameghiniana 52(4):391–417
Ezcurra MD, Nesbitt SJ, Bronzati M, Dalla VFM, Agnolin FL, Benson RBJ, Brissón EF, Cabreira SF, Evers SW, Gentil AR, Irmis RB, Martinelli AG, Novas FE, Silva RL, Smith ND, Stocker MR, Turner AH, Langer MC (2020) Enigmatic dinosaur precursors bridge the gap to the origin of Pterosauria. Nature 588:445–449
Felipe LP, Daniel De S-O, Richard JB (2020) Osteology of the archosauromorph Teyujagua paradoxa and the early evolution of the archosauriform skull, Zoological Journal of the Linnean Society 189(1):378–417, https://doi.org/10.1093/zoolinnean/zlz093
Ferigolo J, Langer MC (2021) A Late Triassic dinosauriform from South Brazil and the origin of the ornithischian predentary bone. Hist Biol 19:23–33
Ferraz SJ, Pohlmann BK, Pinheiro FL, Manfroi J (2021) A paleoflora do afloramento Cerro Chato, Formação Rio do Rasto, Rio Grande Do Sul. Anais do Salão Internacional de Ensino, Pesquisa e Extensão, pp 3
Ferreira JD, Zamorano M, Ribeiro AM (2015) On the fossil remains of Panochthus Burmeister, 1866 (Xenarthra, cingulata, Glyptodontidae) from the Pleistocene of southern Brazil. An Acad Bras Ciênc 87(1):15–27
Ferreira TMP, Mayer EL, Ferigolo J, Pereira JC, Ribeiro AM (2018) Pampatherium humboldtii (Lund, 1839) (Xenarthra, Cingulata) of the southern Brazilian Quaternary: cranial anatomy and taxonomic remarks. Rev Bras Palaeontol 21(2):158–174
Fontoura E, Ferreira JD, Bubadué J, Ribeiro AM, Kerber L (2020) Virtual brain endocast of Antifer (Mammalia: Cervidae), an extinct large cervid from South America. J Morphol 281(10):1223–1240
Forasiepi AM, Soibelzon LH, Gomez CS, Sánchez R, Quiroz LI, Jaramillo C, Sánchez-Villagra MR (2014) Carnivorans at the Great American Biotic Interchange: new discoveries from the northern neotropics. Naturwissenschaften 101:965–974
Francischini H, Dentzien-Dias P, Fernandes MA, Schultz CL (2015) Dinosaur ichnofauna of the Upper Jurassic/Lower Cretaceous of the Paraná Basin (Brazil and Uruguay). J S Am Earth Sci 63:180–190
Francischini H, Sales MAF, Dentzien-Dias P, Schultz CL (2017) The presence of ankylosaur tracks in the Guará Formation (Brazil) and remarks on the spatial and temporal distribution of Late Jurassic dinosaurs. Ichnos 25:177–191
Francischini H, Dentzien-Dias P, Guerra-Sommer M, Menegat R, Santos J, Manfroi J, Schultz CL (2018) A middle Permian (Roadian) lungfish aestivation burrow from the Rio do Rasto Formation (Paraná Basin, Brazil) and associated U-Pb dating. PALAIOS 33(2):69–84
Garcia MS, Müller RT, Da Rosa AAS, Dias-Da-Silva S (2019) The oldest known co-occurrence of dinosaurs and their closest relatives: a new lagerpetid from a Carnian (Upper Triassic) bed of Brazil with implications for dinosauromorph biostratigraphy, early diversification and biogeography. J S Am Earth Sci 91:302–319
Gasparini GM, Kerber L, Oliveira EV (2009) Catagonus stenocephalus (Lund in Reinhardt, 1880) (Mammalia, Tayassuidae) in the Touro Passo Formation (Late Pleistocene), Rio Grande do Sul, Brazil. Taxonomic and palaeoenvironmental comments. Neues Jahrb Geol Palaontol Abh 254(3):261–273
Gaudin TJ, Croft DA (2015) Paleogene Xenarthra and the evolution of South American mammals. J Mammal 96:622–634
Goin FJ, Gelfo JN, Chornogubsky L, Woodburne MO, Martin T (2012) Origins, radiations, and distribution of South American mammals: from greenhouse to icehouse worlds. In: Patterson BD, Costa LP (eds) Bones, clones and biomes. The University of Chicago Press, pp 20–49
Golonka J (2009) Phanerozoic paleoenvironment and paleolithofacies maps. Cenozoic Geol 35(4):507–587
Golonka J, Embry A, Krobicki M. (2018) Late Triassic global plate tectonics. In: Tanner L (ed) The late Triassic world. Topics in geobiology, pp 27–57
Guerra-Sommer M, Cazzulo-Klepzig M (2000a) The Triassic Taphoplora from Paraná Basin, Southern Brazil: an overview. Rev Bras Geociênc 30(3):477–481
Guerra-Sommer M, Cazzulo-Klepzig M (2000b) As Floras Gondwânicas do paleozóico Superior do Rio Grande do Sul. In: Paleontologia do Rio Grande do Sul, CIGO/UFRGS, pp 67
Guerra-Sommer M, Cazzulo-Klepzig M (2007) A flora Dicroidium, um importante estágio na sucessão paleofloristica da Bacia do Paraná, Brasil. In: Carvalho IS, Cassab RCT, Schwanke C, Carvalho MA (orgs) Paleontologia: Cenários da Vida, Interciência, Rio de Janeiro, pp 189–201
Guerra-Sommer M, Cazzulo-Klepzig M, Iannuzzi R, Alves LS (1998) A Flora Dicroidium no Rio Grande do Sul: implicações bioestratigráficas. Pesqui em Geocienc 26(1):3–9
Guerra-Sommer M, Cazzulo-Klepzig M, Ianuzzi R (1999) The Triassic taphoflora of the Paraná Basin, southern Brazil: a biostratigraphical approach. J Afr Earth Sci 29:249–255
Hadler P, Verzi DH, Vucetich MG, Ferigolo J, Ribeiro AM (2008) Caviomorphs (Mammalia, Rodentia) from the Holocene of Rio Grande do Sul State, Brazil: systematics and paleoenvironmental context. Rev Bras Paleontol 11(97):116
Hadler P, Goin FJ, Ferigolo J, Ribeiro AM (2009) Environmental change and marsupial assemblages in Holocene successions of Southern Brazil. Mamm Biol 74:87–99
Hadler P, Cherem JJ, Turbay R, Alberti A, Pardinas UFJ (2016) Diversidade de pequenos mamíferos (Didelphimorphia e Rodentia) do Holoceno do Nordeste do Estado do Rio Grande do Sul, Brasil: implicações taxonômicas e paleoambientais. Rev Bras Paleontol 19:126–144
Herbst R (1986) Cyclodendron cf. leslii (sew.) (Lycopodopsidaceae, Lycopsidae) del pérmico de Paraguay y Uruguay. Facena 6:33–36
Holanda EC, Ribeiro AM, Ferigolo J (2012) New material of Tapirus (Perissodactyla: Tapiridae) from the Pleistocene of southern Brazil. Rev Mex de Cienc 29(2):308–318
Holz M, França AB, Souza PA, Iannuzzi R, Rohn R (2010) A stratigraphic chart of the Late Carboniferous/Permian succession of the eastern border of the Paraná Basin, Brazil, South America. J S Am Earth Sci 29(2):381–399
Hopson JA (1985) Morphology and relationships of Gomphodontosuchus brasiliensis von Huene (Synapsida, Cynodontia, Tritylodontoidea) from the Triassic of Brazil. Neues Jahrbuch Für Geologie und Paläontologie – Monatshefte 5:285–299
Horn BLD, Pereira VP, Schultz CL (2013) Calcretes of the Santa Maria supersequence, Middle Triassic, Rio Grande do sul, Brazil: classification, genesis and paleoclimatic implications. Palaeogeogr Palaeoclimatol Palaeoecol 376:39–47
Horn BLD, Melo TM, Schultz CL, Philipp RP, Kloss HP, Goldberg K (2014) A new third order sequence stratigraphic framework applied to the Triassic of the Paraná Basin, Rio Grande do Sul, Brazil, based on structural, stratigraphic and paleontological data. J S Am Earth Sci 55:123–132
Hsiou AS (2007) A new Teiidae species (Squamata, Scincomorpha) from the Late Pleistocene of Rio Grande do Sul State, Brazil. Rev Bras Paleontol 10(3):181–194
Hsiou AS (2009) O registro fóssil de répteis e aves no Pleistoceno final do estado do Rio Grande do Sul, Brasil. In: Ribeiro AM, Bauermann SG, Scherer CS (eds) Quaternário do Rio Grande do Sul – Integrando Conhecimentos: Monografias da Sociedade Brasileira de Paleontologia, Porto Alegre, pp 154
Hsiou AS, Abdala FN, Arcucci AB (2002) Novo registro de proterocampsídeo (Reptilia: Archosauriformes) do Triássico Médio-Superior do Brasil. Rev Bras Paleontol 48:55
Iannuzzi R (2010) The flora of Early Permian coal measures from the Paraná Basin in Brazil: a review. Int J Coal Geol 83:229–247
Jasper A, Guerra-Sommer M, Menegat R, Cazzulo-Klepzig M, Ricardi-Branco FS, Salvi J (2009) Afloramento Quitéria, Encruzilhada do Sul, RS – Sedimentos lagunares com singular associação fitofossilífera da Formação Rio Bonito. In: Winge M, Schobbenhaus C, Souza CRG, Fernandes ACS, Berbert-Born M, Queiroz ET (eds) Sítios Geológicos e Paleontológicos do Brasil. CPRM, Brasília, p 515
Kellner AWA, Holgado B, Grillo O, Pretto FA, Kerber L, Pinheiro FL, Soares MB, Schultz CL, Lopes RT, Araújo O, Müller RT (2022) Reassessment of Faxinalipterus minimus, a purported Triassic pterosaur from southern Brazil with the description of a new taxon. Peer J 10:e13276
Kerber L, Oliveira E (2008a) Fósseis de vertebrados da Formação Touro Passo (Pleistoceno Superior), Rio Grande do Sul, Brasil: atualização dos dados e novas contribuições. J Geosci 4(2):49–64
Kerber L, Oliveira EV (2008b) Presença de Equus (Perissodactyla, Equidae) e Neuryurus (Cingulata, Glyptodontidae) no Quaternário do município de Quaraí, Oeste do Rio Grande do Sul. Caderno de Pesquisa Série Biologia (UNISC) 20:18–25
Kerber L, Ribeiro AM (2011) Capybaras (Rodentia, Hystricognathi, Hydrochoeridae) from the late Pleistocene of southern Brazil: Neues Jahrbuch für Geologie und Paläontologie. Abhandlungen 261(1):11–18
Kerber L, Kinoshita A, José FA, Figueiredo AMG, Oliveira EV, Baffa O (2011a) Electron Spin Resonance dating of southern Brazilian Pleistocene mammals from Touro Passo Formation, and remarks on the geochronology, fauna and palaeoenvironments. Late Quaternary fossil record of Myocastor Kerr, 1792 (Rodentia: Hystricognathi: Caviomorpha) from Brazil with taxonomical and environmental remarks. Quat Int 245:201–208
Kerber L, Ribeiro AM, Oliveira EV (2011b) The first record of Galea Meyen, 1832 (Rodentia, Hystricognathi, Caviidae) in the late Pleistocene of southern Brazil and its paleobiogeographic implications. Alcheringa 35:445–457
Kerber L, Lopes RP, Vucetich MG, Pereira JC (2011c) Chinchillidae and Dolichotinae rodents (Rodentia: Hystricognathi: Caviomorpha) from the late Pleistocene of Southern Brazil. Rev Bras Paleontol 14:229–238
Kerber L, Lopes RP, Oliveira EV, Ribeiro AM, Pereira JC (2012) On the presence of Holochilus brasiliensis (Desmarest, 1819) (Rodentia: Cricetidae: Sigmodontinae) in the late Pleistocene of southern Brazil. J Geosci 8:47–54
Kerber L, Lessa G, Ribeiro AM, Cartelle C (2014a) Late Quaternary fossil record of Myocastor Kerr, 1792 (Rodentia: Hystricognathi: Caviomorpha) from Brazil with taxonomical and environmental remarks. Late Quaternary fossil record of Myocastor Kerr, 1792 (Rodentia: Hystricognathi: Caviomorpha) from Brazil with taxonomical and environmental remarks. Quat Int 352:147–158
Kerber L, Pitana VG, Ribeiro AM, Hsiou AS, Oliveira EV (2014b) Late Pleistocene vertebrates from Touro Passo Creek (Touro Passo Formation), southern Brazil: a review. Rev Mex de Cienc 31:248–259
Kerber L, Martinelli AG, Rodrigues PG, Ribeiro AM, Schultz CL, Soares MB (2020) New record of Prozostrodon brasiliensis (Eucynodontia: Prozostrodontia) from its type-locality (Upper Triassic, Southern Brazil): comments on the endocranial morphology. Rev Bras Paleontol 23:259–269
Kerber L, Martinelli AG, Müller RT, Pretto FA (2022) A new specimen provides insights into the anatomy of Irajatherium hernandezi, a poorly known probainognathian cynodont from the Late Triassic of southern Brazil. Anat Rec 305(11):3113–3132
Kischlat EE, Lucas SG (2003) A phytosaur from the Upper Triassic of Brazil. J Vertebr Paleontol 23(2):464–467
Kotzian CB, Simões MG, Da-Rosa AAS, Milder S (2005) AMS radiocarbon dating of freshwater mollusk shells from the Touro Passo Formation (Pleistocene-Holocene), RS, Brazil. In: 6th Congresso Latino Americano de Paleontologia, Abstracts, Aracajú
Lacerda MB, Schultz CL, Bertoni-Machado C (2015) First ‘Rauisuchian’ archosaur (Pseudosuchia, Loricata) for the Middle Triassic Santacruzodon Assemblage Zone (Santa Maria Supersequence), Rio Grande do Sul State, Brazil. PLoS One 10:e0118563
Langer MC (2000) The first record of dinocephalians in South America: Late Permian (Rio do Rasto Formation) of the Paraná Basin, Brazil. Abhandlungen 215:69–95
Langer MC, Abdala NF, Richter M, Benton MJ (1999) A sauropodomorph dinosaur from the Upper Triassic (Carnian) of southern Brazil. Comptes Rendus de l ‘Academie Bulg des Sci II, A-Sciences de la Terre et des Planetes 329:511–517
Langer MC, Ribeiro AM, Schultz CL, Ferigolo J (2007) The continental tetrapod-bearing Triassic of South Brazil. In: Lucas SG, Spielmann JA (eds) The global Triassic. Museum of Natural History and Science Bulletin, New Mexico, pp 201–218
Langer MC, Bittencourt JS, Schultz CL (2010) A reassessment of the basal dinosaur Guaibasaurus candelariensis, from the Late Triassic Caturrita Formation of South Brazil. Earth Environ Sci Trans R Soc Edinb 101:301–332
Lavina EL, Lopes RC (1987) A Transgressão Marinha do Permiano Inferior e a Evolução Paleogeográfica do Supergrupo Tubarão no Estado do Rio Grande do Sul. Paula-Coutiana, Porto Alegre, pp 51–103
Lavina EL, Barberena ADC, Azevedo SAK (1991) Tempestades de inverno e altas taxas de mortalidade de répteis mesossauros; um exemplo a partir do afloramento Passo São Borja, RS. Pesquisas 18(1):64–70
Leal LA, Azevedo SAK, Kellner AWA, Da Rosa AAS (2004) A new early dinosaur (Sauropodomorpha) from the Caturrita Formation (Late Triassic), Paran Basin, Brazil. Zootaxa, Nova Zelândia 690:1
Lima JHD, Netto RG, Corrêa CG, Lavina ELC (2015) Ichnology of deglaciation deposits from the Upper Carboniferous Rio do Sul Formation (Itararé Group, Paraná Basin) at central-east Santa Catarina State (southern Brazil). J S Am Earth Sci 63:137–148
Liu J, Soares MB, Reichel M (2008) Massetognathus (Cynodontia, traversodontidae) from the Santa Maria Formation of Brazil. Rev Bras Palaontol 11(1):27–36
Lopes RP, Buchmann FSC (2010) Pleistocene mammals from the southern Brazilian continental shelf. J S Am Earth Sci 31:17–27
Lopes RP, Pereira J (2010) Fossils of Scelidotheriinae Ameghino 1904 (Xenarthra, Pilosa) in the Pleistocene deposits of Rio Grande do Sul, Brazil. Gaea 6:44–52
Lopes RP, Oliveira LC, Figueiredo AMG, Kinoshita A, Baffa O, Buchmann FSC (2010) ESR dating of Pleistocene mammal teeth and its implications for the biostratigraphy and geological evolution of the coastal plain, Rio Grande do Sul, southern Brazil. Quat Int 212(2):213–222
Lopes RP, Kinoshita A, Baffa O, Figueiredo AMG, Dillenburg SR, Schultz CL, Pereira JC (2014a) ESR dating of Pleistocene mammals and marine shells from the coastal plain of Rio Grande do Sul state, Southern Brazil. Quat Int 352:124–134
Lopes RP, Dillenburg SR, Schultz CL, Ferigolo J, Ribeiro AM, Pereira JC, Holanda EC, Pitana VG, Kerber L (2014b) The sea-level highstand correlated to marine isotope stage (MIS) 7 in the coastal plain of the state of Rio Grande do Sul, Brazil. An Acad Bras Ciências 86(4):1573–1595
Lopes RP, Pereira JC, Ferigolo J (2019) A late Pleistocene fossil stork (Ciconiiformes: Ciconiidae) from the Santa Vitória Formation, southern Brazil, and its paleoenvironmental significance. Rev Bras Palaontol 22(3):199–216
Lopes RP, Pereira JC, Kerber L, Dillenburg SR (2020) The extinction of the Pleistocene megafauna in the Pampa of southern Brazil. Quat Sci Rev 242:106–428
Lopes RP, Dillenburg SR, Pereira JC, Sial AN (2021) The paleoecology of Pleistocene giant megatheriid sloths: stable isotopes (δ13C, δ18O) of co-occurring Megatherium and Eremotherium from southern Brazil. Rev Bras Paleontol 24(3):245–264
Maahs R, Küchle J, Scherer CMS, Alvarenga RS (2019) Sequence stratigraphy of fluvial to shallow-marine deposits: the case of the early Permian Rio Bonito Formation, Paraná Basin, southernmost Brazil. Braz J Geol 49(4):e20190059
Maciel L, Ribeiro AM, Sedor F (1996) Considerações sobre quelônios fósseis do Quaternário do Rio Grande do Sul, Brasil. Ameghiniana 33(4):467
Malabarba MC, Abdala F, Weiss FE, Perez PA (2003) New data on the Late Permian Vertebrate Fauna of Posto Queimado, Rio do Rasto Formation, Southern Brazil. Rev Bras Paleontol 6:49–54
Manfroi J, Uhl D, Guerra-Sommer M, Francischini H, Martinelli AG, Soares MB, Jasper A (2015) Extending the database of Permian palaeo-wildfire on Gondwana: charcoal remains from the Rio do Rasto formation (Paraná Basin), Middle Permian, Rio Grande do Sul State, Brazil. Palaeogeogr Palaeoclimatol Palaeoecol 436:77–84
Manfroi J, Trevisan C, Dutra TL, Jasper A, Carvalho MDA, Aquino FE, Leppe M (2023) “Antarctic on fire”: paleo-wildfire events associated with volcanic deposits in the Antarctic Peninsula during the Late Cretaceous. Front Earth Sci 11:1048754
Marcon GT (2008) Contribuição ao estudo dos Proboscidea (Mammalia, Gomphotheriidae) do Quaternário do estado do Rio Grande Do Sul. Rev UnG Geociênc 7(1):93–109
Martinelli AG, Bonaparte JF, Schultz CL, Rubert R (2005) A new tritheledontid (Therapsida, Eucynodontia) from the Late Triassic of Rio Grande do Sul (Brazil) and its phylogenetic relationships among carnivorous non-mammalian eucynodonts. Ameghiniana 42:191–208
Martinelli AG, Soares MB, Schwanke C (2016) Two new cynodonts (Therapsida) from the middle-early late Triassic of Brazil and comments on south American probainognathians. PLoS One 11:e0162945
Martinelli AG, Kammerer CF, Melo TP, Paes-Neto VD, Ribeiro AM, Da-Rosa AAS, Schultz CL, Soares MB (2017a) The African cynodont Aleodon (Cynodontia, Probainognathia) in the Triassic of southern Brazil and its biostratigraphic significance. PLoS One 12(6):e0177948
Martinelli AG, Eltink E, Da-Rosa ÁA, Langer MC (2017b) A new cynodont from the Santa Maria formation, South Brazil, improves Late Triassic probainognathian diversity. Pap Palaeontol 3(3):401–423
Martinelli AG, Corfe IJ, Gill PG, Kallonen A, Rayfield EJ, Rodrigues PB, Schultz CL, Soares MB (2017c) Brasilodon quadrangularis, Brasilitherium riograndensis and Minicynodon maieri (Cynodontia): taxonomy, ontogeny and tooth replacement. In: Congresso Brasileiro de Paleontologia, Ribeirão Preto, pp 25, 189
Martinelli AG, Escobar JA, Francischini H, Kerber L, Müller RT, Rubert R, Schultz CL, Da-Rosa AAS (2020) New record of a stahleckeriid dicynodont (Therapsida, Dicynodontia) from the late Triassic of southern Brazil and biostratigraphic remarks on the Riograndia Assemblage Zone. Hist Biol Paleobiol 33:3101–3110
Martins EA, Sena-Sobrinho M (1951) Lycopodiopsis Derby (Renault) e Glossopteris sp. na Estrada Nova (Permo-Triássico) do Rio Grande do Sul. An Acad Bras Ciênc 2:323–326
Marzoli A, Callegaro S, Dal Corso J, Davies J, Chiaradia M, Youbi N, Bertrand H, Reisberg L, Merle R, Jourdan F (2018) The Central Atlantic Magmatic Province (CAMP): a review. In: Tanner L (ed) The Late Triassic world. Topics in geobiology, vol 46. Springer, pp 91–125
Matos SA, Warren LV, Varejao FG, Assine ML, Simoes MG (2017a) Permian endemic bivalves of the Irati anoxic event, Paraná Basin, Brazil: taphonomical, paleogeographical and evolutionary implications. Palaeogeogr Palaeoclimatol Palaeoecol 469:18–33
Matos SA, Warren LV, Fürsich FT, Alessandretti L, Assine ML, Riccomini C, Simôes MG (2017b) Paleoecology and paleoenvironments of Permian bivalves of the Serra Alta Formation, Brazil: ordinary suspension feeders or Late Paleozoic Gondwana seep organisms? J S Am Earth Sci 77:21–41
Melo MS, Bosetti EP, Godoy LC, Pilatti F (2002) Vila Velha, PR – Impressionante relevo ruiniforme. In: Schobbenhaus C, Campos DA, Queiroz ET, Winge M, Berbert-Born MLC (eds) Sítios Geológicos e Paleontológicos do Brasil. DNPM/CPRM – Comissão Brasileira de Sítios Geológicos e Paleobiológicos (SIGEP), Brasília, vol 1, pp 269–277
Melo TP, Abdala F, Soares MB (2015) The Malagasy cynodont Menadon besairiei (Cynodontia; traversodontidae) in the middle–upper Triassic of Brazil. J Vertebr Paleontol 35:e1002562
Melo TP, Martinelli AG, Soares MB (2017) A new gomphodont cynodont (traversodontidae) from the middle-late Triassic Dinodontosaurus assemblage zone of the Santa Maria Supersequence, Brazil. Palaeontology 60(4):571–582
Melo TP, Ribeiro AM, Martinelli AG, Soares MB (2019) Early evidence of molariform hypsodonty in a Triassic stem-mammal. Nat Commun 10:28–41
Menegat R (2023) Geological memories of South Brazilian landscapes: from Precambriam orogensis to the quaternary coastal plain. In: Overbeck GE, Pillar VDP, Müller SC, Bencke GA (eds) South Brazilian grasslands: ecology and conservation of the Campos Sulinos (this volume). Springer, Cham
Merlotti S (2009) Reavaliação taxonômica de lenhos das formações Irati e Serra Alta, Permiano da Bacia do Paraná, Brasil. Pesqui Geociênc 36(1):11–21
Milani EJ, Rangel HD, Bueno GV, Stica JM, Winter WR, Caixeta JM, Neto OCP (2007) Bacias Sedimentares Brasileiras, Cartas Estratigráficas. B Geoci 15(2):183–205
Milder SES (2000) Arqueologia do Sudeste do Rio Grande do Sul: Uma perspectiva Geoarqueológica: São Paulo, Brasil, Museu de Arqueologia e Etnologia, Universidade de São Paulo (USP), 172 pp
Miller ET (1987) Pesquisas arqueológicas paleoindígenas no Brasil Ocidental. Estud Atacameños 8:37–71
Miron LR, Pavanatto AEB, Pretto FA, Müller RT, Dias-da-Silva S, Kerber L (2020) Siriusgnathus niemeyerorum (Eucynodontia: Gomphodontia): the youngest south American traversodontid? J S Am Earth Sci 97:102394
Montefeltro FC, Langer MC, Schultz CL (2010) Cranial anatomy of a new genus of hyperodapedontine rhynchosaur (Diapsida, Archosauromorpha) from the Upper Triassic of southern Brazil. Earth Environ Sci Trans R Soc 101:27–52
Müller RT, Da-Rosa AAS, Silva LR, Aires ASS, Pacheco CP, Pavanatto AEB, Dias-da-Silva S (2015) Wachholz, a new exquisite dinosaur-bearing fossiliferous site from the Upper Triassic of southern Brazil. J S Am Earth Sci 61:120–128
Müller RT, Langer MC, Dias-da-Silva S (2018) An exceptionally preserved association of complete dinosaur skeletons reveals the oldest long-necked sauropodomorphs. Biol Lett 14:06–33
Neregato R, Souza PA, Rohn R (2008) Registros Palinológicos Inéditos nas Formações Teresina e Rio do Rasto (Permiano, Grupo Passa Dois, Bacia do Paraná) – Implicações Biocronoestratigráficas e Paleoambientais. Pesqui Geociênc 35(2):9–21
Netto RG, Balistieri PRMN, Lavina ELC, Silveira DM (2009) Ichnological signatures of shallow freshwater lakes in the glacial Itararé Group (Mafra Formation, Upper Carboniferous Lower Permian of Paraná Basin, S Brazil). Palaeogeogr Palaeoclimatol Palaeoecol 272:240–255
Neves JP, Anelli LE, Simões MG (2014) Early Permian post-glacial bivalve faunas of the Itararé Group, ParanáBasin, Brazil: paleoecology and biocorrelations with South American intraplate basins. J S Am Earth Sci 52:203–233
Noll SH, Netto RG (2018) Microbially induced sedimentary structures in late Pennsylvanian glacial settings: a case study from the Gondwanan Paraná basin. J S Am Earth Sci 88:385–398
O’Dea A, Lessios HA, Coates AG, Eytan RI, Restrepo-Moreno SA, Cione AL, Collins LS, De Queiroz A, Farris DW, Norris RD, Stallard RF, Woodburne MO, Aguilera O, Aubry MP, Bergreen WA, Budd AF, Cozzuol MA, Coppard SE, Duque-Caro H, Gasparini GM, Grossman EL, Jonhson KG, Keigwin LD, Knowlton N, Leight EG, Leonard-Pingel JS, Marco PB, Pyenson ND, Rachello-Domen PG, Soibelzon L, Todd JA, Vermeij GJ, Jackson JBC (2016) Formation of the Isthmus of Panama. Sci Adv 2:e1600883
Oelofsen BW, Araújo DC (1983a) Palaecological implications of the distribution of mesosaurid reptiles in the Permian Irati Sea (Parana Basin), South America. Rev Bras Geociênc 13(1):1–6
Oelofsen BW, Araújo DC (1983b) Palaeoecology implications of the distribution of mesosaurid reptiles in the Permian Irati Sea (Parana Basin). S Am Rev Bras Geoc 13(1):1–6
Oelofsen BW, Araújo DC (1987a) Mesosaurus tenuidens and Stereosternum tumidum from the Permian Gondwana of both Southern Africa and South America. S Afr J Sci 83(6):3701–3372
Oelofsen BW, Araújo DC (1987b) Two Mesosaurid reptiles (Mesosaurus tenuidens and Stereosternum tumidum) in Permian Gondwana of Southern Africa and South America. J Sci Áfr do Sul 83(6):370–372
Oliveira EV (1992) Mamíferos fósseis do Quaternário do Estado do Rio Grande do Sul, Brasil: Programa de Pós Graduação em Geociências, Universidade Federal do Rio Grande do Sul (UFRGS), Porto Alegre, 118 pp
Oliveira EV (1996) Mamiferos Xenarthra (Edentata) do Quaternário do Estado do Rio Grande do Sul, Brasil. Ameghiniana 31(1):65–75
Oliveira EV (1999) Quaternary vertebrates and climates from southern Brazil. In: Rabassa J, Salemme M (eds) Quaternary of South America and Antarctic Peninsula, vol 12, pp 61–73
Oliveira EV, Pereira JC (2009) Intertropical cingulates (Mammalia, Xenarthra) from the Quaternary of southern Brazil: systematic and paleobiogeographical aspect. Rev Bras Paleontol 12(3):167–178
Oliveira EV, Dutra TL, Zelter F (2002) Megaterídeos (Mammalia, Xenarthra) do Quaternário de Caçapava do Sul, Rio Grande do Sul, com considerações sobre a flora associada. Geol Colombiana 27:77–86
Oliveira TV, Soares MB, Schultz CL (2010) Trucidocynodon riograndensis gen nov et sp nov (Eucynodontia), a new cynodont from the Brazilian Upper Triassic (Santa Maria Formation). Zootaxa 2382(1):1–71
Oliveira TV, Soares MB, Schultz CL, Rodrigues P (2011) A new carnivorous cynodont (Synapsida, Therapsida) from the Brazilian Middle Triassic (Santa Maria Formation): Candelariodon barberenai gen. et sp. nov. Zootaxa 3027:19–28
Pacheco CP, Eltink E, Müller RT, Dias-da-Silva S (2016) A new Permian temnospondyl with Russian affinities from South America, the new family Konzhukoviidae, and the phylogenetic status of Archegosauroidea. J Syst Palaeontol 15(3):241–256
Pacheco C, Müller RT, Langer M, Pretto FA, Kerber L, Dias SS (2019) Gnathovorax cabreirai: a new early dinosaur and the origin and initial radiation of predatory dinosaurs. PeerJ 7:e7963
Paes-Neto VD, Desojo JB, Brust ACB, Schultz CL, Da-Rosa AAS, Soares MB (2021) Intraspecific variation in the axial skeleton of Aetosauroides scagliai (Archosauria: Aetosauria) and its implications for the aetosaur diversity of the Late Triassic of Brazil. An Acad Bras Cienc 93:e20201239
Pankaj KP, Ghosh AK, Kar R, Singh RS, Sarkar M, Chatterjee R (2014) Reappraisal of the genus Dicroidium Gothan from the Triassic sediments of India. Palaeobotanist 63:137–155
Pauliv VE, Dias EV, Sedor FA (2012) A new species of Sphenacanthid (Chondrichthyes, Elasmobranchii) from the Rio do Rasto Formation (Paraná Basin), Southern Brazil. Rev Bras Paleontol 15:243–250
Pauliv VE, Dias EV, Sedor FA, Ribeiro AM (2014) A new Xenacanthiformes shark (Chondrichthyes, Elasmobranchii) from the Late Paleozoic Rio do Rasto Formation (Paraná Basin), Southern Brazil. An Acad Bras Ciênc 86:135–145
Pauliv VE, Martinelli AG, Francischini H, Dentzien-Dias P, Soares MB, Schultz CL, Ribeiro AM (2017) The first Western Gondwanan species of Triodus Jordan 1849: a new Xenacanthiformes (Chondrichthyes) from the late Paleozoic of Southern Brazil. J S Am Earth Sci 80:482–493
Pavanatto AEB, Pretto FA, Kerber L, Müller RT, Da-Rosa AAS, Dias-da-Silva S (2018) A new Upper Triassic cynodont-bearing fossiliferous site from southern Brazil, with taphonomic remarks and description of a new traversodontid taxon. J S Am Earth Sci 88:179–196
Perea D, Soto M, Veroslavsky G, Martínez S, Ubilla M (2009) A Late Jurassic fossil assemblage in Gondwana: biostratigraphy and correlations of the Tacuarembó Formation, Paraná Basin, Uruguay. J S Am Earth Sci 28:168–179
Pereira JC, Lopes RP, Kerber L (2012) New remains of late Pleistocene mammals from the Chuí Creek, Southern Brazil. Rev Bras Paleontol 15(2):228–239
Pérez-Aguilar A, Petri S, Hypólito R, Ezaki S, Souza PA, Juliani C, Monteiro LVS, Azevedo SJM, Moschini F (2008) Pavimento Estriado Guaraú, Salto, SP – Marcas de geleira neopaleozóica no sudeste brasileiro. In: Winge M, Schobbenhaus C, Souza CRG, Fernandes ACS, Berbert-Born M, Queiroz ET (eds) Sítios Geológicos e Paleontológicos do Brasil, pp 10
Piñeiro G, Ferigolo J, Mones A, Núñez DP (2021) Mesosaur taxonomy reappraisal: are Stereosternum and Brazilosaurus valid taxa? Rev Bras Paleontol 24(3):205–235
Pinheiro FL, França MAG, Lacerda MB, Butler RJ, Schultz CL (2016) An exceptional fossil skull from South America and the origins of the archosauriform radiation. Scientific Reports 6:22817
Pinheiro FL, Silva’Neves E, Da’rosa, Átila AS (2021). An early-diverging procolophonid from the lowermost Triassic of South America and the origins of herbivory in Procolophonoidea. Papers in Palaeontology 7(3):1601–1612. https://doi.org/10.1002/spp2.1355
Piperno DR, Becker P (1996) Vegetational history of a site in the Central Amazon Basin derived from phytolith and charcoal records from natural soils. Quat Res 45(2):202–209
Pires EF, Guerra-Sommer M (2004) Sommerxylon spiralosus from Upper Triassic in southernmost Paraná Basin (Brazil): a new taxon with taxacean affinity. An Acad Bras Cienc (Impresso) 76(3):595–609
Pires EF, Guerra-Sommer M, Scherer CMMS (2005) Late Triassic climate in southernmost Parana Basin (Brazil): evidence from dendrochronological data. J S Am Earth Sci 18:213–221
Pitana VG, Ribeiro AM (2007) Novos materiais de Propraopus Ameghino, 1881 (Mammalia, Xenarthra, Cingulata) do Pleistoceno final, Rio Grande do Sul, Brasil. J Geosci 3(2):61–67
Pitana VG, Esteba GI, Ribeiro AM, Cartelle C (2013) Cranial and dental studies of Glossotherium robustum (Owen, 1842) (Xenarthra: Pilosa: Mylodontidae) from the Pleistocene of southern Brazil. Alcheringa 37(2):147–162
Price LI (1946). “Sobre um novo pseudosuquio do Triássico Superior do Rio Grande do Sul”. Boletim Divisão de Geología e Mineralogía. 120: 1–38.
Porto A (1954) História das Missões Orientais do Uruguai. Livraria Selbach, Porto Alegre
Potter PE (1997) The Mesozoic and Cenozoic paleodrainage of South America: a natural history. J S Am Earth Sci 10(5–6):331–344
Pretto FA, Langer MC, Schultz CL (2018) A new dinosaur (Saurischia: Sauropodomorpha) from the Late Triassic of Brazil provides insights on the evolution of sauropodomorph body plan. Zool J Linnean Soc 20:1–29
Raugust T, Lacerda M, Schultz CL (2013) The first occurrence of Chanaresuchus bonapartei Romer 1971 (archosauriformes, proterochampsia) of the Middle Triassic of Brazil from the Santacruzodon Assemblage Zone, Santa Maria Formation (Parana Basin). Geol Soc Spec Publ 379:01–16
Reichel M, Schultz CL, Soares MB (2009) A new traversodontid cynodont (Therapsida, Eucynodontia) from the middle Triassic Santa Maria Formation of Rio Grande do sul. Palaeontology 52(1):229–250
Ribeiro AM, Scherer CS (2009) Mamíferos do Pleistoceno do Rio Grande do Sul. In: Ribeiro AM, Bauermann SG, Scherer CS (eds) Quaternário do Rio Grande do Sul – Integrando Conhecimentos: Monografias da Sociedade Brasileira de Paleontologia, Porto Alegre, pp 171–191
Ribeiro AM, Alvarenga HM, Rosenau M (1995) Primeiro registro de ave fóssil para a Formação Touro Passo (Pleistoceno superior-Holoceno inferior) do Rio Grande do Sul. In: 14th Congresso Brasileiro de Paleontologia, Atas: Uberaba, pp 107
Ribeiro AM, Abdala F, Bertoni RS (2011) Traversodontid cynodonts (Therapsida–Eucynodontia) from two upper Triassic localities of the Paraná Basin, southern Brazil. Ameghiniana 48:111
Ribeiro V, Guerra-Sommer M, Menegat R, Kuhn LA, Simões JC, Braga RB, Bauermann SG, Silva WG (2020) Fire events and vegetation dynamics during the Late Pleistocebe-Meghalayan interval in the southernmost Brazilian coastal plain. Rev Bras Paleontol 23(4):234–250
Ricardi-Branco F, Caires ET, Silva AM (2009) Campo de Estromatólitos Gigantes de Santa Rosa de Viterbo, SP – Excelente registro do litoral do mar permiano Irati, Bacia do Paraná, Brasil. In: Winge M, Schobbenhaus C, Berbert-Born M, Queiroz ET, Campos DA, Souza CRG, Fernandes ACS (eds) Sítios Geológicos e Paleontológicos do Brasil, pp 515
Richter M (2002) A ray-finned fish (Osteichthyes) from the Late Permian of the State of Santa Catarina (Paraná Basin), Southern Brazil. Rev Bras Paleontol 3:56–61
Rocha-Campos AC (2002a) Varvito de Itu, SP – Registro clássico da glaciação neopaleozóica. In: Schobbenhaus C, Campos DA, Queiroz ET, Winge M, Berbert-Born MLC (eds) Sítios Geológicos e Paleontológicos do Brasil. DNPM/CPRM – Comissão Brasileira de Sítios Geológicos e Paleobiológicos (SIGEP), Brasília, vol 1, pp 147–154
Rocha-Campos AC (2002b) Rocha moutonnée de Salto, SP – Típico registro de abrasão glacial do Neopaleozóico. In: Schobbenhaus C, Campos DA, Queiroz ET, Winge M, Berbert-Born MLC (eds) Sítios Geológicos e Paleontológicos do Brasil. DNPM/CPRM – Comissão Brasileira de Sítios Geológicos e Paleobiológicos (SIGEP), Brasília, vol 1, pp 155–159
Rodrigues PH, Ferigolo J (2004) Roedores pleistocênicos da planície costeira do estado do Rio Grande do Sul, Brasil. Rev Bras Paleontol 7(2):231–238
Rodrigues PH, Prevosti FJ, Ferigolo J, Ribeiro AM (2004) Novos materiais de Carnivora do Pleistoceno do estado do Rio Grande do Sul, Brasil. Rev Bras Paleontol 7(1):77–86
Rohn RO (1988) Bioestratigrafia e paleoambientes da Formação Rio do Rasto na borda leste da Bacia do Paraná (Permiano Superior, estado do Paraná). USP, São Paulo, pp 111
Rohn R (2001) A estratigrafia da Formação Teresina (Permiano, Bacia do Paraná) de acordo com furos de sondagem entre Anhembi (SP) e Ortigueira (PR). Ciênc Téc Pet, Seção Exploração de Petróleo 20:2092–2018
Rohn RO, Rösler R (1986) Pteridófitas pecopteróides da Formação Rio do Rasto no estado do Paraná e da Formação Estrada Nova de São Paulo (Bacia do Paraná, Permiano Superior). Bol IG-USP 17:57–76
Rohn R, Rösler O (1990) Conchostráceos da Formação Rio do Rasto (Bacia do Paraná, Permiano Superior): bioestratigrafia e implicações paleoambientais. Rev Bras Geociênc 9:486–493
Rösler R, Rohn RO (1984) Sphenophyllum paranaense (Sphenophyta) da Formação Rio do Rastro de Dorizon, Estado do Paraná. Bol IG-USP 15:97–104
Roth L, Lorscheitter ML (1993) Palynology of a bog in Parque Nacional de Aparados da Serra, East Plateau of Rio Grande do Sul, Brazil. J S Am Sci 8:39–69
Schemiko DCB, Veserly FF, Fernandes LA, Sowek GA (2014) Distinção dos elementos deposicionais fluviais, eólicos e lacustres do Membro Morro Pelado, Permiano Superior da Bacia do Paraná. Geol USP Ser Cient 14(3):29–46
Scherer CSS (2006) Estudo dos Camelidae (Mammalia, Artiodactyla) do Quaternário do Estado do Rio Grande do Sul: Porto Alegre, Brasil, Programa de Pós Graduação em Geociências, Universidade Federal do Rio Grande do Sul (UFRGS), pp 176
Scherer CS, Da Rosa AAS (2003) Um eqüídeo fóssil do Pleistoceno de Alegrete, RS, Brasil. Pesqui em Geociênc 30:33–38
Scherer CM, Lavina ELC (2005) Sedimentary cycles and facies architecture of aeolian-fluvial strata of the Upper Jurassic Guará Formation, southern Brazil. Sedimentology 52:1323–1341
Scherer CSS, Ferigolo J, Ribeiro AM, Cartelle CC (2007) Contribution to the knowledge of Hemiauchenia paradoxa (Artiodactyla, Camelidae) from de Pleistocene of Southern Brazil. Rev Bras Paleontol 10(1):35–52
Scherer CS, Pitana VG, Ribeiro AM (2009) Proterotheriidae and Macrauchenidae (Liptoterna, Mammalia) from the Pleistocene of Rio Grande do Sul State, Brazil. Rev Bras Paleontol 12(3):231–246
Schmitt MR, Martinelli AG, Melo TP, Soares MB (2019) On the occurrence of the traversodontid Massetognathus ochagaviae (Synapsida, Cynodontia) in the early late Triassic Santacruzodon assemblage zone (Santa Maria Supersequence, southern Brazil): taxonomic and biostratigraphic implications. J S Am Earth Sci 93:36–50
Schmitt MR, Martinelli AG, Kaiuca JFL, Schultz CL, Soares MB (2023) Old fossil findings in the Upper Triassic rocks of southern Brazil improve diversity of traversodontid cynodonts (Therapsida, Cynodontia). Anat Rec. in press
Schoch RR (2013) The major clades of temnospondyls: an inclusive phylogenetic analysis. J Syst Palaeontol 11:673–705
Schultz CL, Langer MC (2007) Triassic Tetrapods from Rio Grande do Sul, Brazil. In: Paleontologia: Cenários de Vida, Interciência, pp 269–282
Schultz CL, Martinelli AG, Soares MB, Pinheiro FL, Kerber L, Horn LDB, Pretto FA, Müller RT, Melo T (2020) Triassic faunal successions of the Paraná Basin, southern Brazil. J S Am Earth Sci 104:102846
Schwanke C, Kellner AWA (2009) Interações ecológicas no Triássico. In: Átila Augusto Stock Da-Rosa. (Org.). Vertebrados Fósseis de Santa Maria e região. Santa Maria: Gráfica Editora Pallotti 1:279–301.
Scotese CR, Golonka J (1997) Paleogeographic atlas. PALEOMAP Project, University of Texas at Arlington
Scott AC, Damblon F (2010) Charcoal: taphonomy and significance in geology, botany and archaeology. Palaeogeogr Palaeoclimatol Palaeoecol 291:1–10
Scott AC, Stea R (2002) Fires Sweep across the Mid-Cretaceous landscape of Nova Scotia. Geoscientist 12(1):4–6
Silva NE (in prep) Procolophonidae no Sul do Brasil: Ocorrências inéditas para o Triássico médio da supersequência Santa Maria e suas implicações filogenéticas, paleobiogeográficas e bioestratigráficas. Programa de Pós-Graduação em Biodiversidade Animal, Universidade Federal de Santa Maria
Silva DC, Vaine MEE (2001) Atlas geológico do Estado do Paraná. Mineropar. http://www.iat.pr.gov.br/Pagina/Atlas-Geologico-do-Estado-do-Parana#. Accessed 20 June 2021
Simão-Oliveira DS, Kerber L, Pinheiro FL (2019) Endocranial morphology of the Brazilian Permian dicynodont Rastodon procurvidens (Therapsida: Anomodontia). J Anat 236:384–397
Simões MG, Rocha-Campos AC (1993) Systematics and paleoecology of the pelecypods of the Palermo Formation (Permian), Parana Basin, Brazil. An Acad Bras Cienc 65(3):324–326
Soares MB, Schultz CL, Horn BLD (2011) New information on Riograndia guaibensis Bonaparte, Ferigolo, Ribeiro 2001 (Eucynodontia, Tritheledontidae) from the late Triassic of southern Brazil: anatomical and biostratigraphic implications. An Acad Bras Cienc 83(1):329–354
Soares MB, Martinelli AG, Oliveira TV (2014) A new prozostrodontian cynodont (Therapsida) from the late Triassic Riograndia assemblage zone (Santa Maria Supersequence) of southern Brazil. An Acad Bras Cienc 86:1673–1691
Soto M, Montenegro F, Toriño P, Mesa V, Perea D (2021) A new ctenochasmatid (Pterosauria, Pterodactyloidea) from the late Jurassic of Uruguay. J S Am Earth Sci 111:103472
Stefanello M, Martinelli AG, Müller RT, Dias-da-Silva S, Kerber L (2023) A complete skull of a stem mammal from the Late Triassic of Brazil illuminates the early evolution of prozostrodonts. J Mamm Evol. Online first
Strapasson A, Pinheiro FL, Soares M (2015) On a new stereospondylomorpha temnospondyl from the Middle-Late Permian of Southern Brazil. Acta Palaeontol Pol 6(4):843–855
Taboada AC, Neves JP, Weinschütz LC, Pagani MA, Simões MG (2016) Eurydesma–Lyonia fauna (Early Permian) from the Itararé group, Paraná Basin (Brazil): a paleobiogeographic W–E trans-Gondwanan marine connection. Palaeogeogr Palaeoclimatol Palaeoecol 449:431–454
Ubilla M, Oliveira EV, Rinderknecht A, Pereira JC (2008) The hystricognath rodent Microcavia in the late Pleistocene of Brazil (Rio Grande do Sul, South America) (Mammalia: Caviidae): biogeographic and paleoenvironmental implications. Abhandlungen 247:15–21
Vargas-Peixoto D, Colusso CS, Da-Rosa AAS, Kerber L (2021) A new record of Lestodon armatus Gervais 1855 (Xenarthra, Mylodontidae) from the Quaternary of southern Brazil and remarks on its postcranial anatomy. Hist Biol 33(2):159–175
Vesely FF (2007) Sistemas subaquosos alimentados por fluxos hiperpicnais glaciogênicos: modelo deposicional para arenitos do Grupo Itararé, Permocarbonífero da Bacia do Paraná. In: Boletim de Geociências da Petrobras, Rio de Janeiro, pp 7–25
von Huene F (1928) Ein cynodontier aus des Trias Brasiliens. Centralblatt Für Mineralogie, Geologie Und Paläontolgie 1928B:251–270
von Huene F (1935) Lieferung 1. Anomodontia. Die Fossilen Reptilien des Südamerikanischen Gondwanalandes. Ergebnisse der Sauriergrabungen. In: CH Beck’sche Verlagsbuchhandlung Südbrasilien 1928/29, München, pp 1–82
von Huene F (1936) Lieferung 2. Cynodontia. Die Fossilen Reptilien des Südamerikanischen Gondwanalandes. Ergebnisse der Sauriergrabungen. In: CH Beck’sche Verlagsbuchhandlung Südbrasilien 1928/29, München, pp 83–160
von Huene F (1938) Die fossilen Reptilien des südamerikanischen Gondwanalandes. Neues Jahrb Geol Palaontol Abh Referate 3:142–151
Warren LV, Almeida RP, Hachiro J, Machado R, Roldan LF, Steiner SS, Chamani MAC (2008) Evolução sedimentar da Formação Rio do Rasto (Permo-Triássico da Bacia do Paraná) na porção centro sul do estado de Santa Catarina, Brasil. Rev Bras Geociênc 38(2):213–227
Warren LV, Assine ML, Simões MG, Riccomini C, Anelli LE (2015) A Formação Serra Alta, Permiano, no centro-leste do Estado de São Paulo, Bacia do Paraná, Brasil. Braz J Geol 45:109–126
Wegener A (1912) Die Entstehung der Kontinente. Geol Rundsch 3(4):276–292
Weinschütz LC, Castro JC (2006) Sequências Deposicionais da Formação Taciba (Grupo Itararé, Neocarbonífero a Eopermiano) na Região de Mafra (SC), Bacia do Paraná. Rev Bras Geociênc 36:243–252
Wildner W, Ramgrab GE, Lopes RC, Iglesias CMF (2008) Mapa Geológico do Estado do Rio Grande do Sul, Escala 1:750.000. CPRM – Serviço Geológico do Brasil. http://www.cprm.gov.br/publique/media/mapa_rio_grande_sul.pdf. Accessed 03 Mar 2021
Wildner W, Camozzato E, Toniolo JA, Binoto RB, Iglesias CMF, Laux JH (2014) Mapa geológico do Estado de Santa Catarina, Escala 1:500.00. Serviço Geológico do Brasil. http://www.cprm.gov.br/publique/media/mapa_geologico_santa_catarina.pd. Accessed 27 Out 2021
Wilner E, Lemos VB, Scomazzon AK (2016) Associações naturais de conodontes Mesogondolella spp., Grupo Itararé, Cisuraliano da Bacia do Paraná. J Geosci 9:30–36
Woodburne MO (2010) The great American biotic interchange: dispersals, tectonics, climate, sea level and holding pens. J Mamm Evol 17:245–264
Xavier PLA, Aurélio F, Soares MB, Horn BLD, Schultz CL (2018) Sequence stratigraphy control on fossil occurrence and concentration in the epeiric mixed carbonate-siliciclastic ramp of the Early Permian Irati Formation of southern Brazil. J S Am Earth Sci 88:157–178
Zerfass H, Lavina EL, Schultz CL, Garcia AGV, Faccini UF, Chemale JF (2003) Sequence stratigraphy of continental Triassic strata of southernmost Brazil: a contribution to Southwestern Gondwana palaeogeography and palaeoclimate. Sediment Geol 161:85–105
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2024 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
da Rosa, Á.A.S., Kerber, L., Pinheiro, F.L., Manfroi, J. (2024). A Look into the Past: Fossils from the Campos Sulinos Region. In: Overbeck, G.E., Pillar, V.D.P., Müller, S.C., Bencke, G.A. (eds) South Brazilian Grasslands. Springer, Cham. https://doi.org/10.1007/978-3-031-42580-6_3
Download citation
DOI: https://doi.org/10.1007/978-3-031-42580-6_3
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-42579-0
Online ISBN: 978-3-031-42580-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)