
Abstract
The stapedial artery (SA) is an embryological artery that allows the development of orbital and dural arteries as well as maxillary branches. Its complex embryological development explains numerous anatomical variations of the middle meningeal artery (MMA) and orbital arteries. A few anatomists dissected a human middle ear that bore a persistent SA describing the origin and course of this artery [1–7]. ENT surgeons reported cases discovered during middle ear surgeries explaining the technical difficulties of the surgery in the presence of a persistent SA [8–20]. More recently, neuroradiologists paid particular attention to this anatomical variant [21–26]. These studies, associated with the comparative anatomy, allow to understand the persistence of the SA. In this chapter, after an embryological remind concerning hyostapedial, carotid and ophthalmic systems, the anatomical variations including the SA will be explained [27–29].
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
The stapedial artery (SA) is an embryological artery that allows the development of orbital and dural arteries as well as maxillary branches. Its complex embryological development explains numerous anatomical variations of the middle meningeal artery (MMA) and orbital arteries. A few anatomists dissected a human middle ear that bore a persistent SA describing the origin and course of this artery [1,2,3,4,5,6,7]. ENT surgeons reported cases discovered during middle ear surgeries explaining the technical difficulties of the surgery in the presence of a persistent SA [8,9,10,11,12,13,14,15,16,17,18,19,20]. More recently, neuroradiologists paid particular attention to this anatomical variant [21,22,23,24,25,26]. These studies, associated with the comparative anatomy, allow to understand the persistence of the SA. In this chapter, after an embryological refresh concerning hyostapedial, carotid and ophthalmic systems, the anatomical variations including the SA will be explained [27,28,29].
History
The first cadaveric case of persistent SA was described by Hyrtl in 1836, who called attention to an artery running across the obturator of the stapes in a human cadaver, with some similarities with a vessel found in hibernating animals [30]. In the first half of the twentieth century, the phenomenal publication of Dorcas Padget, based on the dissections of 22 human embryos of the Carnegie collection, gave lot of information about the embryological development of the cranio-facial arteries and in particular of the hyostapedial system [31]. In the same period, Altmann (1947) furnished a comprehensive explanation of the development of the aortic arches and of the carotid system variants [32]. However, only in the 1960s we find the first angiographic demonstration of MMA variations and in particular, the persistence of a SA [33, 34]. At the end of the 1970s, P. Lasjaunias was able to give a comprehensive explanation of all variations implicating the SA. Its more famous articles were published between 1975 and 1990 and are summarized in its textbook [23,24,25, 35,36,37,38,39]. Diamond in the 1990s also published some articles based on the comparative anatomy principally with the great apes that increased our understanding of the stapedial system [5, 27, 28]. In this last three decades, few case reports of interesting anatomical variations implicating the SA were published [17, 40,41,42,43,44,45,46,47].
Comparative Anatomy
The studies of comparative anatomy were fundamental in the understanding of the embryological development of the SA. At the beginning of the twentieth century, Tandler, studying the SA development in rats, explained the SA regression and the role of the external carotid artery (ECA) in the formation of the MMA [31]. Rats differed from human embryos, because the supraorbital branch of the SA is primarily dependent on the first aortic arch (mandibular branch). Fuchs (1905), in the rabbit, also showed that the first two first aortic arches participate in the formation of the ECA as in human embryos [20]. More recently, Diamond (1991) and Falk (1993) compared the development of the SA between human and great apes’ embryos and highlighted some analogies [28, 29].
Embryology
The development and regression of the SA is strictly related to some embryonic systems. In this part of the chapter, we will summarize the main steps of the internal carotid artery (ICA) and ophthalmic artery (OA) development. We will also present a detailed description of the hyostapedial system, in order to understand the different possible adult variations related to this artery.
The Carotid System
Since the detailed description of the ICA embryological development has been widely treated in the chapter “Embryological Development of the Internal Carotid Artery,” we will focus only on the understanding of the “aberrant flow of the ICA.” It consists in an intratympanic course of the ICA and could be associated with a persistent SA and consequent variation in origin of the MMA. The principal steps of ICA development are resumed in Fig. 1.

Embryological segments of the internal carotid artery. The illustrations (a–c) show consecutive stages of ICA embryological development. The first stages of development (a) are characterized by the presence of three aortic arches that link the ventral and dorsal aorta (VA, DA). The VA regresses together with the ventral part of the aortic arches. The dorsal remnants of the aortic arches persist as embryonic arteries. These embryonic arteries divide the ICA into seven embryological segments: (1) The cervical segment: it derives from the remnant of the third aortic arch (III AA). (2) The ascending intrapetrous segment: it is the remnant of the dorsal aorta (DA) between the second (II AA) and third aortic arches. The division point between segments 2 and 3 is at the point of origin of the Hyoid artery (HA), that is the dorsal remnant of the second aortic arch. (3) The horizontal intrapetrous segment: it is the remnant of the DA between the first (I AA) and second aortic arches. The division point is at the point of origin of the mandibular artery (MA), that corresponds to the dorsal remnant of the first aortic arch. (4) The intracavernous ascending segment: it originates from the DA between the first aortic arch and the primitive maxillary artery (PMA), that connects the DA of the two sides (DAl: dorsal aorta left; DAr: dorsal aorta right). At the junction between segments 4 and 5 also the trigeminal artery (TA) takes its origin. This latter represents a primitive connection between the cavernous ICA and the basilar artery (BA). (5) The horizontal intracavernous segment: it derives from the DA between the PMA and the primitive dorsal ophthalmic artery (PDOA). (6) The clinoid segment: corresponds to the DA between the PDOA and the primitive ventral ophthalmic artery (PVOA). (7) The terminal segment: the terminal ICA between (PVOA) and the primitive ICA bifurcation into the future anterior cerebral artery (ACA) and future posterior communicating artery (PComA). The figure shows also the hypoglossal artery (HypA) and the proatlantal artery (PA), which have their origin proximal to the third aortic arch and will contribute to the formation of external carotid artery (ECA) branches
In the first stage of Padget (4–5 mm embryos), the two first aortic arches initiate their natural regression allowing the ICA to be individualized. The embryological segments of the ICA are derived from the third aortic arch and from the dorsal aorta cranial to the third aortic arch [31, 32, 39]. The dorsal aorta also regresses at the same time between the third and fourth aortic arches. Embryologically, the ICA has been divided by Lasjaunias et al. in seven different segments [48]. The first one corresponds to the third aortic arch from the origin of the ventral pharyngeal artery (future ECA) to the junction between the third aortic arch and the dorsal aorta. The second segment is the dorsal aorta between the third and second aortic arches. The third segment is the dorsal aorta between the second and first aortic arches. The fourth segment is the dorsal aorta between the first aortic arch and the origin of the trigeminal artery (TA) (and the primitive maxillary artery (PMA)). The fifth segment is the dorsal aorta between the origin of the TA (and the primitive maxillary artery) and the origin of the primitive dorsal ophthalmic artery (PDOA, future inferolateral trunk (ILT)). The sixth segment is between the origin of the PDOA and the origin of the primitive ventral ophthalmic artery (PVOA). The seventh segment is between the PVOA and the primitive carotid bifurcation [23, 24, 31, 39].
It is important to note that the carotid bulb has not the same embryological origin than the other segments of the ICA. It originates from the pharyngo-occipital system that easily explains variations in origin of the ascending pharyngeal and occipital arteries. Agenesis or abnormal regression of one or more segments of the ICA explains an intratympanic course of the ICA (by pharyngo-carotid anastomosis) and also the different type of “reperfusion” in case of ICA agenesis [12, 32]. The intratympanic course of the ICA also named as “aberrant flow of the ICA” is the consequence of the abnormal regression of the first and second segments with anastomosis between the inferior tympanic artery (from the ascending pharyngeal artery) and the caroticotympanic artery (from the carotid artery) that infuses distally the ICA. The pseudo-ICA has consequently an intratympanic course without passing through the stapes [12, 39, 49].
The Hyostapedial System
“Hyostapedial artery” is the term used to describe the complete embryological development of the second aortic arch. The SA, that develops from the hyoid artery and takes its name after passing through the crus of the stapes, is an embryonic artery present between the stages III and VI of Padget (9–24 mm). This is an important embryological system from which numerous dural, orbital and facial arteries develop. The steps of SA development are summarized in Table 1.
The hyoid artery is the dorsal remnant of the second aortic arch which regresses early in the embryological development (4–5 mm, stage I of Padget) [2, 12]. After this arch regression, approximately at the 5–6 mm stage (stage II of Padget), the hyoid artery presents a rapid lateral elongation between the beginning of the 7th and 19th weeks of gestation, when it gives an anastomosis to the mandibular artery (remnant of the first aortic arch) [31]. During the stage III of Padget (7–12 mm embryos) the hyoid artery is prominent and grows cranially, passing, as stapedial artery, through the crus of stapes. In contrast, the mandibular artery is short and difficult to identify. During this stage the initial formation of the two main branches of the SA (supraorbital and maxillomandibular divisions) is visible, even if they are only completely established in the stage IV of Padget (12–14 mm). At the 16–18 mm stage (stage V of Padget), the SA continues its elongation to the gasserian region passing into the future tympanic cavity and particularly through the crus of the stapes reaching its maximal development [23, 31, 32]. The supraorbital artery, which follows the ophthalmic root of the trigeminal nerve, allows the development of orbital branches (supraorbital, lacrimal, ethmoids and frontal arteries) and also of the MMA [39]. The maxillomandibular artery gets out of the cranial cavity through the foramen spinosum and gives extracranially its two branches: infraorbital and mandibular arteries (future infraorbital and inferior alveolar arteries). After this maximal development of the SA, two annexations and two regressions occur to give the adult configuration of the MMA. Intracranially, the orbital branches are annexed by the primitive OA and the trans-sphenoidal segment of the supraorbital branch regresses leaving an anastomotic artery between the anterior branch of the MMA and the lacrimal artery (the sphenoidal artery) that penetrates the orbit through the superior orbital fissure (SOF). Extracranially, the ventral pharyngeal artery annexes the maxillomandibular artery of the SA, forming the proximal stem of the MMA and becoming the ECA. Consequently, by flow reversal into the SA, its tympanic portion regresses and leaves as remnants the caroticotympanic artery (from the ICA) and the superior tympanic artery (from the petrous branch of the MMA). These annexations and regressions concerning the SA happen during the stage VI of Padget (20–24 mm embryos) [31, 39].
Ophthalmic Artery
The embryogenesis of the primitive OA is related in some phases to the hyostapedial systems. These phases are resumed and related to the concomitant SA steps of development in Table 2.
The formation of the OA depends on two different arteries during 4–18 mm stages, the PDOA and the PVOA. The PDOA develops from the cavernous segment of the primitive internal carotid artery and the PVOA from the anterior division of the primitive ICA. The PDOA penetrates the orbit through the superior orbital fissure and the PVOA through the optic canal.
Then, two major anastomoses between these two arteries are formed. The first one is an intraorbital plexiform anastomosis supplied by the two arteries around the optic nerve (future second segment of the OA). The second anastomosis is intradural between the PVOA and the primitive ICA to form the definitive supraclinoidal origin of the OA. The proximal portion of the PVOA (between its origin on the ACA and its anastomosis with the ICA) then regresses to give the adult stem of the OA.
In a following step, the proximal part of the PDOA regresses and its remnant becomes the ILT of the primitive ICA.
Concomitantly, in 7 and 24 mm embryos (stages III–VI), the SA arises from the primitive hyoid arch of the petrous ICA. As described above, its supraorbital artery enters the orbit to give orbital branches. It divides into two different branches: the ethmoido-nasal and the lacrimal arteries. The ethmoido-nasal artery anastomoses with the primitive OA at the arterial ring around the optic nerve described before. Then, the trans-sphenoidal part of the supraorbital artery regresses and its orbital branches are annexed by the primitive OA to give the adult conformation.
The Trigeminal Artery
The trigeminal artery (TA) represents one of the carotid-basilar anastomoses, that appears in the 4–5 mm embryos (stage I of Padget) and disappears in the 12 mm embryos (stage III of Padget). It originates from the basilar artery (BA), between the superior and the anteroinferior cerebellar arteries, passes medial to the gasserian ganglion and follows the trigeminal nerve to the primitive ICA, at the level of the junction between the fourth and fifth segments of the ICA. The TA has a common origin on the ICA with the primitive maxillary artery. The carotid remnant of these two primitive arteries will be the future meningo-hypophyseal trunk, from which the lateral clival, marginal tentorial and inferior hypophyseal arteries will originate [31, 32]. On the other hand, the basilar remnant of the TA could be considered responsible for MMA possible origin from the BA in adult, thanks to its anastomosis in the gasserian region with a persistent stapedial artery. The details of this variant are explained in the chapter “Anatomy, Embryology and Variations of the Middle Meningeal Artery.”
Formation of the Inferolateral Trunk
As supported by the theory of P. Lasjaunias concerning the embryology of the ophthalmic artery, the ILT is the carotid remnant of the PDOA. The PDOA develops from the cavernous segment of the primitive ICA and penetrates the orbit through the SOF. In the 40 mm embryos, the proximal part of the PDOA regresses and its remnant becomes the ILT. At the adult configuration, the ILT is composed of four branches: (1) superior branch that supplies the roof of the cavernous sinus; (2) anteromedial branch which passes into the SOF; (3) anterolateral branch that runs into the foramen rotundum and (4) posterior branch passing medial to the gasserian ganglion. These branches present a lot of anastomoses in the cavernous region that correspond to remnants of primitive trigeminal, ophthalmic, stapedial and maxillary arteries.
Persistence of the Stapedial Artery
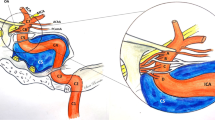
Changes in the events previously described can determine different adult configurations of the arteries related to the hyostapedial system, like the persistency of the SA. The incidence of this variant is estimated to 0.48% after a series of more than 1000 temporal bone dissections [6]. The possible variants related to the persistency of the SA are illustrated in Fig. 2, including the rarer association of SA persistency with the so called pseudo-petrous ICA.

Persistent stapedial artery and intratympanic flow of the ICA. (a) Normal regression process of the hyostapedial system. After the stapedial artery proximal regression, its proximal remnant originates from the ICA as carotico-tympanic artery (CTA). Its distal part is annexed by the ventral pharingeal artery, future external carotid artery, to give the internal maxillary artery (IMA) and its first and major branch, the middle meningeal artery (MMA). The MMA side of the stapedial artery persists as petrosal branch (PB), that gives birth to the superior tympanic artery (STA). The anastomotic branch to the hyostapedial system from the ascending pharyngeal artery (APhA) persists as inferior tympanic artery (ITA). The CTA, STA and ITA participate to the adult vascularization of the middle ear. (b) Persistent stapedial artery. In case of complete SA persistency, this artery originates from the petrous ICA, and gives intracranially the MMA and the IMA exits the middle fosse skull base through the foramen spinosum as extracranial branch. This variant is due to the lack of annexation of the maxillo-mandibular branch by the ventral pharyngeal artery. (c) Pseudo-petrous or aberrant intratympanic ICA. This variant is due to the agenesis of the cervical ICA (third aortic arch). The cervical ICA agenesis is bypassed through a hypertrophic inferior tympanic artery that courses from the APhA, through the inferior tympanic canal and reaches the CTA. The intratympanic ICA flow is called “aberrant” since the artery does not course into its usual canal, but into an enlarged inferior tympanic canal. (d) Pseudo-petrous ICA + SA persistency. This variant is due to two phenomena: the agenesis of the cervical ICA, and stapedial artery persistency. In this case the ITA does not reach the CTA but the persistent SA (pSA)
Complete Persistence of the Stapedial Artery
The complete persistence of the SA is a very rare variant, only two cases were published in a context of ICA aneurysm or PHACE syndrome [25, 39]. In these cases, the SA could be seen as in the embryo taking its origin from the petrous ICA, passing through the middle ear and giving its two branches: one intracranial that corresponds to the MMA and the other extracranial leaving the cranial cavity through the foramen spinosum. Consequently, the foramen spinosum is enlarged, the cochlear promontory is eroded and the IMA arises from the SA instead of the ECA. Such an anatomical variant could easily be explained by the embryology and particularly by the absence of annexation of the maxillomandibular branch (of the SA) by the ventral pharyngeal artery. Consequently, in absence of reversion of the arterial flow into the SA, its proximal (intratympanic) stem could not regress [39].
Partial Persistence of the Stapedial Artery
The partial persistence of the SA is more frequent and in this case, only the intracranial branch of the SA keeps its origin from the petrous ICA [1, 7, 21,22,23,24, 26, 32, 50, 51]. The foramen spinosum is absent (an orifice does not exist without its contents) or reduced in size and the MMA arises from the SA instead of the IMA. This variant is explained by the regression of the proximal part of the maxillomandibular artery instead of the proximal part of the SA [39]. A rare case of bilateral partial persistent SA is illustrated in Fig. 3. We must note that it is surprising that the complete persistence of the SA is even much rarer than its partial persistence. This could be the illustration of a lack of embryological knowledge of the SA or a misunderstanding, particularly in the annexation of the maxillomandibular branch by the ventral pharyngeal artery.

DSA with bilateral partial persistent Stapedial Artery in patient with moyamoya disease. (a) and (b) Show respectively right and left ICA injection with the MMA (red arrow) arising from the petrous ICA. This phenomenon is possible because of bilateral stapedial artery persistency. In (b) the ICA is narrowed and stopped at the level of the ophthalmic artery due to moyamoya disease. (c) Shows a 3D reconstruction of the right ICA from which the MMA originates (red arrow). (d) Shows the left ECA injection without the MMA enhancement, because of its absence
Persistence of the SA Associated with Aberrant Flow of the ICA (Pseudo-Petrous ICA)
In rare cases, the persistence of the SA is associated with an intratympanic course of the ICA (also known as “aberrant flow of the ICA”) [12, 21, 23, 24, 52, 53]. This association of vascular variants concerning both carotid and hyostapedial systems was described anatomically and angiographically [32, 39]. In these cases, the ICA enters the skull base through an enlarged inferior tympanic canal (narrowing of the vessel on digital subtraction angiography (DSA)), passes into the tympanic cavity to bend anteriorly and reaches its normal carotid canal. The MMA arises from the ICA in its tympanic segment and passes through the stapes to have the same course described in the previous paragraph. The exocranial orifice of the carotid canal is therefore absent in these cases [23, 24]. The intratympanic course of the ICA is explained by the agenesis of the two first segments of the primitive ICA. The cervical segment is in fact the ascending pharyngeal artery with a hypertrophied inferior tympanic artery that maintains its anastomosis with the caroticotympanic artery (branch of the ICA) into the tympanic cavity. The correct term of this aberrant flow of the ICA is, in reality, tympano-caroticotympanic variant [39]. It bypasses the absence of the cervical ICA and provides a collateral circulation to the remainder of the ICA. Therefore, the cervical and intratympanic segments of this artery do not derivate from the carotid system but from the pharyngo-occipital and hyostapedial systems (pseudo-ICA) [23, 24, 39]. The agenesis of the first two segments of the ICA could be partial and the ICA appears duplicated [52].
Pharyngo-Tympano-Stapedial Artery
This very rare variant was first described by Lasjaunias (1977) in its original publication [23]. The same case served as illustration in the textbook Surgical Angiography and only one similar case was published by Baltsavias et al. (2012) [39, 42]. The MMA arises from the cervical portion of the ICA, ascends along the cervical ICA, enters the tympanic cavity through the inferior tympanic canal and follows the usual course of the SA. The two cases described were presented as “partial” persistence of the SA with only the MMA arising from the SA and the absence of the foramen spinosum. In this variant, an annexation of the SA by the inferior tympanic artery (branch of the ascending pharyngeal artery) with regression of the proximal part of the SA explains this vascular configuration. Therefore, the SA arises from the cervical instead of the petrous segment of the ICA.
MMA Origin of the OA
The first description of orbital branches arising from the MMA was published in 1872 by Curnow [54]. He already noted in its case dissection that all orbital branches were supplied by the MMA except the central retinal artery which kept its origin from the supracavernous ICA. Twenty years after this first description, Musgrove (1893) presented another case of MMA origin of the OA where also the central retinal artery was supplied by the MMA and the ICA did not give any orbital branch [55]. After other isolated cases found during cerebral or orbital dissections, Hayreh (1962) reported 6 cases among its 170 orbital dissections whom the MMA participated completely (2 cases) or partially (4 cases) in the orbital supply [1, 27, 56,57,58]. Then, Moret (1977) and Lasjaunias (1977) described with precision the contribution of the MMA to the orbital vascularization and gave us a more comprehensive explanation of this anatomical variation [23, 36].
The incidence of OA that arises from the MMA and penetrates the orbit through the SOF is estimated between 1.4% and 2.5% [56]. In this anatomical variation, orbital arteries are supplied by the anterior division of the MMA passing through the SOF or through the spheno-orbital foramen. The central retinal artery generally keeps its vascular supply from the supracavernous ICA but in few cases, also the central retinal artery is supplied by the MMA without ICA participation [59]. A case of complete supply of the orbital arteries by the MMA is illustrated in Fig. 4.

MMA origin of the OA. The figure shows a case of complete OA origin from the MMA. The selective injection of the MMA in antero-posterior projection (a) shows that the sphenoidal artery (SphA) arises from the MMA after its bifurcation into the anterior (red arrow) and posterior division (blue arrow). It courses along the inner surface of the temporal and sphenoidal bone and enters the orbit through the SOF to give rise to the OA. (b) Shows an oblique ICA injection highlighting the absence of the OA
As explained before, during the embryogenesis, the SA gives the supraorbital branch which allows the development of the lacrimal artery and anastomoses with the OA around the optic nerve (participation of the peri-optic circle). The proximal part of the supraorbital branch then normally regresses and the lacrimal artery is annexed by the OA [39]. The persistence of MMA branches (or, in extremis, MMA origin of the OA) could be explained by two different phenomena: the absence of anastomosis between the supraorbital branch and the OA, with consequent persistence of a dual supply of the orbital arteries, or the persistence of the proximal stem of the supraorbital branch of the SA with regression of the primitive OA (complete MMA origin of the OA). Although the supraorbital branch gives off its bifurcation (lacrimal artery laterally and ethmoido-nasal medially) usually inside the orbit, in 30% of cases it can divide outside the orbit in the middle cranial fossa. In such cases, the ethmoido-nasal artery passes through the SOF, but the lacrimal artery penetrates the orbit through its own canal: the spheno-orbital foramen (canal of Hyrtl, lacrimal foramen, sinus canal). The medial branch, passing through the SOF, is named recurrent meningeal artery in its intra-orbital segment and sphenoidal artery in its intracranial segment [23, 31, 39, 56].
Conclusion
All the anatomical variations linked to the development of the SA, resumed in Table 3, could have a clinical impact. Since each vascular segment that regresses during the embryological life, usually persists as anastomosis between two arteries, the knowledge of the embryology and presence of these anastomoses is of paramount importance in case of external carotid branches embolization. Each residual anastomosis represents a potential undesired leak of liquid embolic agent, with possible clinical implications. The knowledge of the embryological variants described in this chapter, represents the basis to understand the different possible adult configurations of the MMA and their clinical implications, which will be described in the chapter “Anatomy, Embryology and Variations of the Middle Meningeal Artery.” The most common pathologies, whose treatment requires a detailed knowledge of MMA embryology and variants, are dural arterio-venous fistulas, meningeal tumors and chronic subdural hematomas.
References
Fisher AG. A case of complete absence of both internal carotid arteries, with a preliminary note on the developmental history of the Stapedial artery. J Anat Physiol. 1913;48(Pt 1):37–46.
Kelemen G. Arteria stapedia, in bilateral persistence. AMA Arch Otolaryngol. 1958;67(6):668–77.
Sando I, Baker B, Black FO, Hemenway WG. Persistence of stapedial artery in trisomy 13–15 syndrome. Arch Otolaryngol. 1972;96(5):441–7.
Marano GD, Horton JA, Gabriele OF. Persistent embryologic vascular loop of the internal carotid, middle meningeal, and ophthalmic arteries. Radiology. 1981;141(2):409–10. https://doi.org/10.1148/radiology.141.2.7291564.
Diamond MK. Unusual example of a persistent stapedial artery in a human. Anat Rec. 1987;218(3):345–54. https://doi.org/10.1002/ar.1092180316.
Moreano EH, Paparella MM, Zelterman D, Goycoolea MV. Prevalence of facial canal dehiscence and of persistent stapedial artery in the human middle ear: a report of 1000 temporal bones. Laryngoscope. 1994;104(3 Pt 1):309–20. https://doi.org/10.1288/00005537-199403000-00012.
Kawai K, Yoshinaga K, Koizumi M, Honma S, Tokiyoshi A, Kodama K. A middle meningeal artery which arises from the internal carotid artery in which the first branchial artery participates. Ann Anat. 2006;188(1):33–8. https://doi.org/10.1016/j.aanat.2005.08.013.
Baron SH. Persistent stapedial artery, necrosis of the incus, and other problems which have influenced the choice of technique in stapes replacement surgery in otosclerosis. Laryngoscope. 1963;73:769–82. https://doi.org/10.1288/00005537-196306000-00014.
House HP, Patterson ME. Persistent Stapedial artery: report of two cases. Trans Am Acad Ophthalmol Otolaryngol. 1964;68:644–6.
Maran AG. Persistent stapedial artery. J Laryngol Otol. 1965;79(11):971–5.
Davies DG. Persistent stapedial artery: a temporal bone report. J Laryngol Otol. 1967;81(6):649–60.
Steffen TN. Vascular anomalites of the middle ear. Laryngoscope. 1968;78(2):171–97. https://doi.org/10.1288/00005537-196802000-00001.
Marion M, Hinojosa R, Khan AA. Persistence of the stapedial artery: a histopathologic study. Otolaryngol Head Neck Surg. 1985;93(3):298–312. https://doi.org/10.1177/019459988509300304.
Boscia R, Knox RD, Adkins WY, Holgate RC. Persistent stapedial artery supplying a glomus tympanicum tumor. Arch Otolaryngol Head Neck Surg. 1990;116(7):852–4.
Pahor AL, Hussain SS. Persistent stapedial artery. J Laryngol Otol. 1992;106(3):254–7.
Govaerts PJ, Marquet TF, Cremers WR, Offeciers FE. Persistent stapedial artery: does it prevent successful surgery? Ann Otol Rhinol Laryngol. 1993;102(9):724–8. https://doi.org/10.1177/000348949310200914.
Wardrop P, Kerr AI, Moussa SA. Persistent stapedial artery preventing successful cochlear implantation: a case report. Ann Otol Rhinol Laryngol Suppl. 1995;166:443–5.
Jain R, Gandhi D, Gujar S, Mukherji SK. Case 67: persistent stapedial artery. Radiology. 2004;230(2):413–6. https://doi.org/10.1148/radiol.2302021108.
Yamamoto E, Hirono Y. Persistent stapedial artery associated with otosclerosis. ORL J Otorhinolaryngol Relat Spec. 1988;50(6):382–4. https://doi.org/10.1159/000276017.
Hogg ID, Stephens CB, Arnold GE. Theoretical anomalies of the stapedial artery. Ann Otol Rhinol Laryngol. 1972;81(6):860–70.
Guinto FC Jr, Garrabrant EC, Radcliffe WB. Radiology of the persistent stapedial artery. Radiology. 1972;105(2):365–9. https://doi.org/10.1148/105.2.365.
Teal JS, Rumbaugh CL, Bergeron RT, Segall HD. Congenital absence of the internal carotid artery associated with cerebral hemiatrophy, absence of the external carotid artery, and persistence of the stapedial artery. Am J Roentgenol Radium Ther Nucl Med. 1973;118(3):534–45.
Lasjaunias P, Moret J, Manelfe C, Theron J, Hasso T, Seeger J. Arterial anomalies at the base of the skull. Neuroradiology. 1977;13(5):267–72.
Lasjaunias P, Moret J. Normal and non-pathological variations in the angiographic aspects of the arteries of the middle ear. Neuroradiology. 1978;15(4):213–9.
Rodesch G, Choi IS, Lasjaunias P. Complete persistence of the hyoido-stapedial artery in man. Case report. Intra-petrous origin of the maxillary artery from ICA. Surg Radiol Anat. 1991;13(1):63–5.
Sheikh BY, Coates R, Siqueira EB. Stapedial artery supplying sphenoid wing meningioma: case report. Neuroradiology. 1993;35(7):537–8.
Diamond MK. Homologies of the meningeal-orbital arteries of humans: a reappraisal. J Anat. 1991;178:223–41.
Diamond MK. Homologies of the stapedial artery in humans, with a reconstruction of the primitive stapedial artery configuration of Euprimates. Am J Phys Anthropol. 1991;84(4):433–62. https://doi.org/10.1002/ajpa.1330840408.
Falk D. Meningeal arterial patterns in great apes: implications for hominid vascular evolution. Am J Phys Anthropol. 1993;92(1):81–97. https://doi.org/10.1002/ajpa.1330920107.
Hyrtl J. Über mehrere im Menschen vorkommende analogieen derjenigen arterie welche Otto bei mehreren Winterschläfern durch den Steigbügel verlaufend entdeckte. Neue beobachtungen aus dem gebieteder menschlichen und vergleichenden anatomie. Med Jahrb d Oesterreich Staates. 1836;10:457–66.
Padget DH. The development of cranial arteries in the human embryo. Contrib Embryol. 1948;212:205–62.
Altmann F. Anomalies of the internal carotid artery and its branches; their embryological and comparative anatomical significance; report of a new case of persistent stapedial artery in man. Laryngoscope. 1947;57(5):313–39.
Gabriele OF, Bell D. Ophthalmic origin of the middle meningeal artery. Radiology. 1967;89(5):841–4. https://doi.org/10.1148/89.5.841.
Salamon G, Guerinel G, Demard F. Radioanatomical study of the external carotid artery. Ann Radiol (Paris). 1968;11(3):199–215.
Seeger JF, Hemmer JF. Persistent basilar/middle meningeal artery anastomosis. Radiology. 1976;118(2):367–70. https://doi.org/10.1148/118.2.367.
Moret J, Lasjaunias P, Theron J, Merland JJ. The middle meningeal artery. Its contribution to the vascularisation of the orbit. J Neuroradiol. 1977;4(2):225–48.
Merland JJ, Theron J, Lasjaunias P, Moret J. Meningeal blood supply of the convexity. J Neuroradiol. 1977;4(2):129–74.
Moret J, Lasjaunias P, Vignaud J, Doyon D. The middle meningeal blood supply to the posterior fossa (author’s transl). Neuroradiology. 1978;16:306–7.
Lasjaunias P, Bereinstein A, Ter Brugge KG. Surgical neuroangiography. Berlin: Springer; 2001.
Maiuri F, Donzelli R, de Divitiis O, Fusco M, Briganti F. Anomalous meningeal branches of the ophthalmic artery feeding meningiomas of the brain convexity. Surg Radiol Anat. 1998;20(4):279–84.
Shah QA, Hurst RW. Anomalous origin of the middle meningeal artery from the basilar artery: a case report. J Neuroimaging. 2007;17(3):261–3. https://doi.org/10.1111/j.1552-6569.2007.00108.x.
Baltsavias G, Kumar R, Valavanis A. The pharyngo-tympano-stapedial variant of the middle meningeal artery. A case report. Interv Neuroradiol. 2012;18(3):255–8. https://doi.org/10.1177/159101991201800302.
Kumar S, Mishra NK. Middle meningeal artery arising from the basilar artery: report of a case and its probable embryological mechanism. J Neurointerv Surg. 2012;4(1):43–4. https://doi.org/10.1136/jnis.2010.004465.
Plas B, Bonneville F, Dupuy M, Sol JC, Chaynes P. Bilateral ophthalmic origin of the middle meningeal artery. Neurochirurgie. 2013;59(4–5):183–6. https://doi.org/10.1016/j.neuchi.2013.08.005.
Cvetko E, Bosnjak R. Unilateral absence of foramen spinosum with bilateral ophthalmic origin of the middle meningeal artery: case report and review of the literature. Folia Morphol (Warsz). 2014;73(1):87–91. https://doi.org/10.5603/FM.2014.0013.
Salem MM, Fusco MR, Dolati P, Reddy AS, Gross BA, Ogilvy CS, et al. Middle meningeal artery arising from the basilar artery. J Cerebrovasc Endovasc Neurosurg. 2014;16(4):364–7. https://doi.org/10.7461/jcen.2014.16.4.364.
Kimball D, Kimball H, Tubbs RS, Loukas M. Variant middle meningeal artery origin from the ophthalmic artery: a case report. Surg Radiol Anat. 2015;37(1):105–8. https://doi.org/10.1007/s00276-014-1272-z.
Lasjaunias P, Santoyo-Vazquez A. Segmental agenesis of the internal carotid artery: angiographic aspects with embryological discussion. Anat Clin. 1984;6(2):133–41. https://doi.org/10.1007/bf01773165.
Margolis MT, Newton TH. Collateral pathways between the cavernous portion of the internal carotid and external carotid arteries. Radiology. 1969;93(4):834–6. https://doi.org/10.1148/93.4.834.
McLennan JE, Rosenbaum AE, Haughton VM. Internal carotid origins of the middle meningeal artery. The ophthalmic-middle meningeal and stapedial-middle meningeal arteries. Neuroradiology. 1974;7(5):265–75.
Martins C, Yasuda A, Campero A, Ulm AJ, Tanriover N, Rhoton A Jr. Microsurgical anatomy of the dural arteries. Neurosurgery. 2005;56(2 Suppl):211–51; discussion 51.
Koenigsberg RA, Zito JL, Patel M, Swartz JD, Goldofsky E, Zahtz G. Fenestration of the internal carotid artery: a rare mass of the hypotympanum associated with persistence of the stapedial artery. AJNR Am J Neuroradiol. 1995;16(4 Suppl):908–10.
Silbergleit R, Quint DJ, Mehta BA, Patel SC, Metes JJ, Noujaim SE. The persistent stapedial artery. AJNR Am J Neuroradiol. 2000;21(3):572–7.
Curnow J. Two instances of irregular ophthalmic and middle meningeal arteries. J Anat Physiol. 1873;8(Pt 1):155–6.
Musgrove J. Origin of ophthalmic artery from middle meningeal. J Anat Physiol. 1893;27(Pt 2):279–81.
Hayreh SS, Dass R. The ophthalmic artery: I. origin and intra-cranial and intra-canalicular course. Br J Ophthalmol. 1962;46(2):65–98.
Chanmugan PK. Note on an unusual ophthalmic artery associated with other abnormalities. J Anat. 1936;70:580–2.
Bossi R, Pisani C. Collateral cerebral circulation through the ophthalmic artery and its efficiency in internal carotid occlusion. Br J Radiol. 1955;28(333):462–9. https://doi.org/10.1259/0007-1285-28-333-462.
Louw L. Different ophthalmic artery origins: embryology and clinical significance. Clin Anat. 2015;28(5):576–83. https://doi.org/10.1002/ca.22470.
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2023 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Bonasia, S., Bojanowski, M.W., Robert, T. (2023). Embryological Development of the Hyostapedial System. In: Robert, T., Bonasia, S., Bojanowski, M.W. (eds) Anatomy of Cranial Arteries, Embryology and Variants. Springer, Cham. https://doi.org/10.1007/978-3-031-32913-5_32
Download citation
DOI: https://doi.org/10.1007/978-3-031-32913-5_32
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-32912-8
Online ISBN: 978-3-031-32913-5
eBook Packages: MedicineMedicine (R0)