
Abstract
The purpose of this chapter is to present a model for cleft repair based upon principles of developmental anatomy. The final configuration of a cleft is the result of tissue deficiency or fusion failure during in embryogenesis leading to displacement and distortion of otherwise normal adjacent structures. It has long been my conviction that understanding why and how this process occurs would provide the key to surgical strategy based on unraveling a mechanism rather than imposing a geometric design. Developmental field reassignment (DFR) evolved as means to reassign developmental fields that are physically mismatched so that, with all fields restored into normal relationships, growth can proceed normally and relapse can be avoided. We shall first consider relevant developmental anatomy, the microform cleft, dissection technique, dental arch management, protocols, and their implications.
My goal with these illustrations is to show exactly how to carry out DFR, including variations depending upon the philosophical goals of the surgical team. We will start with the original DFR design to point out what aspects proved to be of true functional value and what aspects could be either be discarded or used as an option.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Rotation-advancement
- Subperiosteal plane
- Alveolar cleft bone graft
- Bone morphogenetic protein-2
- Mesenchymal stem cells
- Cleft rhinoplasty
Introductory Remarks
All plastic surgeons involved in the care of cleft-affected children and adults experience first hand both the rewards and the limitations of our craft. This work requires a certain mindset characterized by four qualities: an intense curiosity about cleft biology, a relentless pursuit of good technique, an unflinching assessment of results (both good and bad), and humility in the constant search for better ideas. Of all these characteristics, humility is perhaps the most important. It leads us to constantly search out and appreciate the work of other surgeons. Superior concepts or protocols should be embraced, not rejected.
In this Chap. 1 shall discuss how my thinking about developmental field reassignment (DFR) for the management of cleft lip and cleft lip nose has morphed over the years. The principle sources of change stem from long-term observations of the outcomes of surgical interventions (my own and those of others) and intellectual contributions of colleagues that made so much biologic sense to me that they demanded to be incorporated. I intend this as a narrative, with personal observations and conjectures. There will be some historical materials as well. Surgical evolution does not take place in a straight line; I have therefore included blind alleys and misconceptions as well, because, in their resolution, value information was gained. This I hope to pass on for your consideration.
We shall cover the following topics
-
Evolution of the DFR model
-
Embryologic strategy for cleft repair: problems and solutions
-
Developmental anatomy of the central lip–nose complex
-
Functional lip repair: the 5 As
-
DFR: toward a rational protocol
-
Technical details of DFR cleft lip–nose repair
-
Addendum: Sotereanos alveolar cleft procedure
Developmental Field Reassignment Evolution of a Concept
As we launch into this, our final discussion of cleft management, I would like to share with you a brief explanation of how DFR came into being. What now seems a rational system, based on neuroembryology, neurovascular fields, and biochemical signals started out as a more simplistic model which evolved in fits and starts, in several iterations.
The Process Concept
Developmental field reassignment surgery is a return to basics. The anatomy we see represents a rearrangement of the original building blocks of embryonic tissues. But, unlike the jig saw puzzle on the front of cleft craft, the anatomy is not in two dimensions, rather it is a four-dimensional problem. The event may be as early as early as stage 9, the assembly of the first arch, but its consequences are played out over time until by 8 weeks all the components of the pathology established. Fetal growth merely cements this anatomy into its final form.
As previously recounted, upon returning from Nicaragua, I could not escape the conviction that something was missing in our surgical management of clefts. Why was relapse so common? Why did it always take the same form? Why the need for secondary surgery? It was as if there were processes, unleashed by the cleft event that were not addressed by initial surgery, and that these would lead to an inevitable deterioration of the results (Fig. 19.1).

Cleft: a biologic problem in four dimensions. Three processes, a deficiency state leads to a division or maldistribution of tissue. This leads to a displacement of otherwise normal structures. Over time these structures undergo distortion. This case of bilateral cleft, incomplete, right more than left (Top left) demonstrates an acceptable initial result (Top right), albeit with asymmetry of the nose, the right ala being displaced downward (Top right). By late childhood/mixed dentition (Bottom left) the nose is flattening further. (Bottom right): In the late teens (lower right) orbicularis asymmetry is obvious. The nasal airway is compromised. He is a mouth breather. Nose creases over the ala representing nasalis tethering. We shall see him again the end of this photo essay. [Courtesy of Michael Carstens, MD]
It occurred to me that a process-oriented cleft repair would have to address four issues.
Deficiency of tissue at some specified site (I was not sure, but it seemed to be in the nasal floor). Somehow this was related to a division or cleft ranging in severity from microform to complete. The false insertion of muscles and the consequent imbalance [1] would lead to displacement, as in the alar base, columella, and premaxilla. Finally, over time, the sum of these processes would worsen, causing distortion of otherwise normal tissues. I set out to see how these processes could be undone so that relapse could be avoided (Figs. 19.2 and 19.3).

Pathologic anatomy of unilateral cleft. (Left): soft tissues; (Right): skeletal anatomy. Medial prolabium “acquires” displaced tissue from premaxilla
• Philtral prolabium
• Non-philtral prolabium
Medial crus displaced downward
• “Shoulder” of columella flat
Nasal tip distortion
• Foreshortening of nasal floor
Division
• External rotation of premaxilla
• Stem cells distributed asymmetrically in space
Lateral
Pre-turbinate deficiency site > displacement of lateral crus
• Nasal tip distortion
• Inward rotation of nostril sill
Deficient/absent frontal process > misinsertion of nasalis
• “band” in lateral wall
• Dilator becomes constrictor
Division of force vectors
[Courtesy of Michael Carstens, MD]

4 Ds of cleft repair (developmental model for cleft prior to injection studies). Deficiency > Division > Displacement > Distortion. Key: Blue = Frontonasal skin with r1 neural crest and StV1 ethmoids. Red = vestibular lining with r1 neural crest and StV1 ethmoids. Tan = r2 medial. This older drawing does not appreciate the invasion of first and second arch elements into the lateral nasal process (thereby changing its color). [Reprinted from Carstens MH. Functional matrix cleft repair: Principles and techniques. Clin Plast Surg 2004; 31: 159–189. With permission from Elsevier.]
While a resident and fellow at the University of Pittsburgh, I saw the ability of the wide subperiosteal approach designed by Sotereanos to close alveolar clefts of any dimension. At the same time, it gave an aesthetic correction of the midface, a centralization of the lip and nose which remained stable over time. The sliding sulcus procedure was merely an application of the concepts of Soteranos and Delaire to primary cleft repair. As initially reported, the lip dissection remained rotation-advancement. Although primary gingivoperiosteoplasty (GPP) using a gingival incision proved to be impractical the biologic lessons stuck with me (Figs. 19.4, 19.5, 19.6, 19.7 and 19.8).
-
Avoid a buccal sulcus incision to protect osteogenic periosteal cells from injury
-
Transfer those cells into correct position to make membranous bone where desired during the period of ensuing rapid facial growth
-
Freeing the alar base
-
Correction of the septum

Sotereanos muco-gingivoperiosteoplasty (GPP). Developmental field reassignment originated from experience in alveolar cleft bone grafting and in observations in multiple secondary cases, all showing the same pattern of relapse. Sliding sulcus flap (S) transfers two dental units of tissue per side. Bilateral flaps can close fistulas of almost any size. The S flaps are accompanied by a wide periosteal dissection (Delaire). This results in a tension-free centralization of the entire midface soft tissue envelope with normalized projection on the cleft side. In bilateral cases the effect is striking. What also occurs is a reassignment of bioactive stem cell populations from the periphery to the center. On the cleft side, left to their own devices, stem cells will correctly synthesize bone in the wrong position in space. In 1997 stem cells per se were not on my radar screen but my thinking underwent in shock treatment in 2000 when Martin Chin and I translated Boyne’s work with rhBMP2 in orthopedics to morphogen-driven craniofacial bone reconstruction, in situ osteogenesis (ISO). See Chap. 20. (Top left): Full-thickness incisions in oral side mucoperiosteum of cleft-side maxillary (pink) and non-cleft side premaxillary. (Bottom left): nasal side mucoperiosteal flaps (yellow on both sides) reflected backward into the cleft. (Top right): counter-incisions (transverse and vertical) made in maxillary mucoperiosteal advancement. (Bottom right): Maxillary flap translocated mesially two dental units. [Reprinted from Cohen M, Figueroa A, Aduss H. The role of gingiva mucoperiosteal flaps in the repair of alveolar clefts. Plast Reconstr Surg 1989; 83(5): 812–816. With permission from Wolters Kluwer Health, Inc.]

The sliding sulcus procedure. Initial attempt to address facial asymmetry, both as perceived, and with future growth, was based on subperiosteal centralization of the soft tissue envelope. This had the effect of reassigning the membranous bone-forming “factory” into correct position such that future osteogenesis would take place centrically and not in a displaced position. If the car factory belongs in Detroit, don’t make your Fords in Flatbush. [Reprinted from Carstens MH. Functional matrix cleft repair: Principles and techniques. Clin Plast Surg 2004; 31: 159–189. With permission from Elsevier.]

Subperiosteal dissection with sliding sulcus flaps. (Left): Original construct of sliding sulcus repair used a gingivoperiosteoplasty (GPP) with incision along the margin of the alveolus as in the sotereanos alveolar cleft procedure (shown at the end of this essay). Bone production was not adequate and did not prevent lateral segment from collapsing but forward growth was stimulated. (Right): The same mobilization can be obtained without the gingival incision, reserving the S flaps for later use to close the alveolar cleft at age 4 (ideally) or later. [Reprinted from Carstens MH. Functional matrix cleft repair: Principles and techniques. Clin Plast Surg 2004; 31: 159–189. With permission from Elsevier.]

Original case of sliding sulcus subperiosteal repair with gingivoperiosteoplasty. Midline symmetry is striking and position of the left alar base is maintained at 3 years. Unfortunately, would be many years before I recognized the importance of nasalis reassignment restoration of the airway. It also correcting the alar base, which is here asymmetrical. [Reprinted from Carstens MH. The sliding sulcus procedure: simultaneous repair of unilateral clefts of the lip and primary palate—a new technique. J Craniofac Surg 1999; 10(5): 415–429. With permission from Wolters Kluwer Health, Inc.]

An original subperiosteal repair with gingivomucoperiosteoplasty. The photos are very old but what is so striking in the incredible degree of centralization and the complete forward re-positioning of the left ala. [Reprinted from Carstens MH. The sliding sulcus procedure: simultaneous repair of unilateral clefts of the lip and primary palate—a new technique. J Craniofac Surg 1999; 10(5): 415–429. With permission from Wolters Kluwer Health, Inc.]
The Functional Matrix Concept: Enter Neuroembryology
In 1999–2001 five events came together to jolt my thinking into a higher level. From 1997 to 1999 I prepared a series of papers regarding applications of process theory and subperiosteal repair for unilateral, bilateral, and cleft nose deformities. This process required extensive review of the literature, including turning all three volumes of cleft craft into an underlined and annotated shambles. So many papers seemed to be variations on design with little embryology to back them up. But I was impressed by the functional matrix concept of Moss and Opitz, the idea, born out of orthodontics and genetics, that tissues were organized into blocks with individual behaviors. I was not aware at the time of Taylor’s work with angiosomes. To understand this better I devoured standard embryology texts but found them utterly lacking regarding craniofacial development. In particular, I wanted to understand better the blood supply to the central face, nose, columella, and lip. Somehow there must be functional matrices involved in the pathology of clefts.
About this time, I had the good fortune to meet and work with fetal pathologist, Dr. Geoffrey Machin at Kaiser Hospital in Oakland. Geoff and I procured fetal specimens, some with normal facies, some with variants of holoprosencephaly and a right unilateral cleft lip and alveolus with an intact secondary palate. We injected them with results published previously [2] and discussed in Chap. 18 (Figs. 19.9 and 19.10). In so doing we discovered the anatomic boundaries of what I thought (falsely) to be the internal carotid/ophthalmic supply to the midline. This first developmental field map was of neurovascular origin; although it lacked a developmental rationale it was immediately practical, as is suggested a different way to dissect the lip–nose complex, initially termed Functional Matrix Repair. The functional matrix concept gave much better facial symmetry and nasal projection but did not give a better airway (Figs. 19.11 and 19.12).

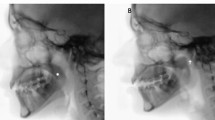
Blood supply to the prolabium: a change in perception. Injection study showing dye from ethmoid circulation descending the columella and entering the prolabium, which is shown here in hemisection to reveal the vessels. [Courtesy of Michael Carstens, MD]

Field separation: AP. Fetal cadaver nasal dorsum showing paired StV1 anterior ethmoid arteries descending over the nasal tip for the columella 0. Midbrain neural crest: nasal skin (hp2 neural fold epithelium), vestibular skin (hp2 placode epithelium). Hindbrain neural crest: vomer, premaxilla (r2). Cleft prolabium = philtral prolabium + non-philtral prolabium. [Courtesy of Michael Carstens, MD]

Contemporary surgical designs violate embryonic fields. Incisions (yellow) in both triangular (Tennison-Randall, Fisher) and rotation-advancement transect the embryonic supply to the philtrum. They do not give direct access to the nose. Key: light blue = PNC skin and StV1 ethmoid mesenchyme; red = placodal vestibular skin and StV1 ethmoid; blue-gray = hindbrain mucosa and r2 medial nasopalatine axis; dark blue = hindbrain mucosa and r2 lateral nasopalatine axis [3]. [Reprinted from Carstens MH. Functional matrix cleft repair: Principles and techniques. Clin Plast Surg 2004; 31: 159–189. With permission from Elsevier.]

Primary subperiosteal repair with gingivoperiosteoplasty. Symmetry is maintained at 2 years. Good growth of the ipsilateral maxilla which was recessed pre-op. The left nostril is slightly smaller (no nasalis repair). Note normal eruption of lateral incisor into the cleft site. There is discoloration of the enamel. [Courtesy of Michael Carstens, MD]
The discovery of the anatomic basis of developmental fields I attribute directly to Dr. Machin. In the process of our dissections Geoff told me about a new theory that he found intriguing: the existence of homeobox genes and how that related to the body axis. At Machin’s prompting, I discovered the 1989 work on rhombomeres and cranial nerves by Lumsden and Keynes [4]. This had instant application to the Tessier system classification system of craniofacial clefts. Neuroanatomy had always been one of my obsessions. When I first learned about the Tessier system in 1987, it was apparent these rare clefts had some relationship to the sensory distribution of the trigeminal nerve (Figs. 19.13 and 19.14). What’s more, rhombomeric compartments seemed to give a neuromeric explanation for the pharyngeal arches which was directly relevant to understanding the development of the face and the cleft condition.

Tessier cleft classification system. From the get-go I realized there had to be a neuroanatomic explanation for the cleft zones because they were concordant with the trigeminal system. This perception in 1987 would eventually morph into neurovascular fields based on neuromeres. Left: [Courtesy of Michael Carstens, MD]. Right: [Reprinted from Carstens MH. Developmental field reassignment cleft surgery: reassessment and refinements. In: Bennun R, Harfin J, Sandor G (ed). Cleft Lip and Palate Management: A Comprehensive Atlas. Hoboken, NJ: John Wiley & Sons; 2016:83–111. With permission from John Wiley & Sons.]

Tessier cleft zones. Skull bones on the left are color coded for the origin of the neural crest mesenchyme: prosomeric (blue), mesomeric (white), rhombomeric (tan). On the right, individual cleft zones are depicted. The exact biologic relationship between orbital and maxillary cleft zones could be due to similarities in the homeotic code (or a different gene code) specifying their respective neurovascular axes. The following fields are all in register with rhombomere r2 and supplied by StV2 branches of the stapedial from the sphenopalatine fossa: fluorescent = medial nasopalatine axis, tan/flesh = lateral nasopalatine axis, light green = descending palatine axis (greater and lesser), green = medial infraorbital axis, blue = lateral infraorbital axis, gold = superior alveolar axis and purple = zygomaticofacial axis, and pink = zygomaticotemporal axis. Fields supplied by the non-stapedial external carotid system are colored orange. Fields supplied by StV1 branches are light blue for hp2 frontonasal skin and red for hp2 placodal skin. [Reprinted with permission from Sari, E. Tessier Number 30 Facial Cleft: A Rare Maxillofacial Anomaly. Turkish Journal of Plastic Surgery, 2018; 26(1): 12–19.]
Very murky, however, was the basis for development and vascularization of the upper face. Why the reliance of a supposedly internal carotid source of supply for tissues unrelated to the forebrain? Or, more profoundly, were they actually related? Perhaps it was not that “the face predicts the brain” but rather that “the brain predicts the face.” In the meantime, I heard that John Rubenstein’s group at UCSF was doing work with hox genes. So it was, that one foggy morning, I traipsed over to his lab on Parnassus hill. There I discovered the prosomeric system. Luis Puelles and Rubenstein had successfully linked the development of the entire embryonic neuraxis to the homeotic system [5]. I knew that a functional matrix cleft repair could be designed to make use of a neuromeric tissue map.
Functional matrix cleft repair abandoned rotation-advancement forever. The identity and anatomy of the non-philtral prolabium (NPP) was clear from injection, consistent with r2 mesenchyme and supported by a known neuroangiosome, the medial nasopalatine artery. Functional matrix dissection applied equally well to both unilateral and bilateral clefts, eliminating the cognitive dissonance shared by all surgeons doing two very different procedures for the very same pathology, something that never made sense to me. The rationale and design of the functional matrix repair were published in 2000 and updated in 2004 with better understanding of the nasal tissues.
But in 2000 along came Flor, a little girl born with a severe left-sided lateral facial cleft involving structures on both sides of the midline. I sat by the bedside and made a drawing of what I saw. Suddenly, an entire map of the face appeared with upper deck structures from r2 and r4 matching up with lower deck structures of r3 and r5 neural crest. This fit the Dlx system I had learned about from Michael DePew at Rubenstein’s lab. It was obvious that all pharyngeal arches had to have the same mapping system to create compartments that would become developmental fields. We shall meet Flor again in our final chapter (Chap. 20) as she posed a reconstructive challenge that lead to the first application of recombinant human bone morphogenetic protein-2 in craniofacial surgery: it was the birth of in situ osteogenesis.
Developmental Field Reassignment: The Impact of Bruce Carlson and Dorcas Padget
Let’s fast forward to 2006. The functional matrix was working nicely. I was working with Martin Chin on application of rhBMP2 to the alveolar cleft and closing it up at the primary surgery. [As noted before, I have now come to the conclusion that secondary repair at age 4 (a Talmant concept), again, ideally with rhBMP2, is the best management for the dental arch.] Blood supply issues between alveolar extension palatoplasty (AEP) (Chap. 17) and the lip made a lot of sense. By this time with texts of molecular embryology by Bruce Carlson and of developmental biology Scott Gilbert incorporated into neuromeric theory things were falling into place. But I found myself lost at sea when it came to the vascular system. The arterial system was the door to understanding the developmental map of the face. But where was the key?
References are a good thing. Hinrichsen’s invaluable SEM studies of facial development (which I recommend to all those interested) were just such a footnote. So too, as I looked for clues to the vascular system, I found Carlson’s text a reference research by a woman artist working at the Carnegie Institution on the arterial supply to the head in staged embryos. Her fascinating story, recounted elsewhere, was an inspiration and her drawing superlative. In this book I have attempted to bring them alive with color coding and simplified concepts. Nevertheless, Padget showed me the unifying concept of blood supply as multiple iterations through development with the stapedial system as the crowning innovation explaining those structures of the face that are never considered as part of the first arch yet form the interface between maxilla and brain.
Once the neurovascular map was complete, it seemed appropriate to rename the conceptual basis for cleft repair as DFR. The DFR concept applies to many craniofacial problems, both as a better means to understand their pathogenesis and promoting more rational surgical strategies. In the future others will use field mapping to explore old problems in a new way. As for the remainder of this chapter, we shall see how DFR technique was further modified, in particular, in service to a functional aspect of clefts that has previously defined my best efforts and those of many others: the nasal airway (Fig. 19.15).

Developmental field map: UCL and BCL. (Left): Prolabium consists of two pairs of fields, StV1 (yellow) and StV2 (pink). Normal embryogenesis assigns StV2 as medial nasal floor above the central and lateral incisor fields, PMxA and PMx B. In the cleft state tissues from r1 remain abnormally fused with r1. This results in an island of medial nasopalatine non-philtral prolabium (pink) located on one of both sides of the true anterior ethmoid philtrum. (Right): Bilateral clefts have symmetrical fields. Key: yellow = StV1 anterior ethmoid; pink = StV2 medial nasopalatine; magenta = StV2 lateral nasopalatine; orange = StV2 infraorbital + ECA facial Gray vermilion flaps = ECA. Note: DFR flaps. Diagram shows NPP elevated along with LCC, but these can be separated (as per Matthews). Lateral wall mucoperiosteum is elevated from what was formerly PMxF is now Mx F. Subvestibular mobilization with elevator continues under the periosteum inside the nostril until reaching nasal bone. Mucosal flaps are dropped as “diapers” without committing them until the end of the case. M can be sutured to the lateral lip for coverage. It can also be based proximally and rotated into the sulcus release site under the ala which gave access to the bone. M can provide coverage to the lip or turned up into the vestibule. [Courtesy of Michael Carstens, MD]
And herein, at risk of being a bit informal, I’d like to include some personal details as to how these innovations have emerged. No craniofacial surgeon works in a vacuum. The contributions of long-term work by David Matthews, Jean Claude Talmant, Jean-Pierre Lumineau, and Luis Monasterio are essential components in the evolution of the developmental field model. What is remarkable is that each of us, coming from different experiences, has arrived at a very similar set of priorities and techniques. My purpose here is to bring these perspectives together into a single unitary philosophy based on the developmental field model of cleft formation. As Victor Veau put it so well “the surgery of clefts is merely experimental embryology.” This chapter intends to prove him right.
Toward an Embryologic Strategy for the Surgical Management of Clefts
The organization of this chapter is built around five sets of issues that I have found most problematic. We shall consider each of these in turn.
Medial Wall Dissection
Certain aspects of the DFR design produced results that were not ideal or did not make sense. (1) In some cases, I observed flattening of the cupid’s bow at the intersection of the white roll and the cleft-side philtral column. (2) Since 2003 I have been advancing the cleft-side medial crus into the nasal tip using an anterolateral columellar incision. Although this design worked well (with excellent scars) I had the nagging suspicion that a simpler design would accomplish the same goals. (3) I realized that the intranasal extension of the prolabial incision was an embryologic challenge because it would have to conform to the neurovascular field map of the medial nasal wall. What were the precise boundaries between skin of forebrain neural crest origin and skin originating from the hindbrain neural crest?
Lateral Wall Dissection
Nasal airway expansion, a top priority in DFR surgery, was difficult to maintain, despite the near-perfect fit of the NPP flap into the lateral wall releasing incision. Why should the cleft-side nasal airway arrive at its particular shape? What cause or causes could explain the functional limitations of breathing on the cleft side? There seemed to be a missing piece to the puzzle. What was it?
Dental Arch Management
Coming to clarity regarding the surgical-orthodontic sequence with respect to the alveolar arch was a very frustrating problem for me. The long-standing debate regarding presurgical orthopedics has been characterized by loudly stated opinions and a near-total lack of developmental biology. Understanding the cleft maxilla and its reconstruction requires the juxtaposition of new input from developmental field biology, with a surgical and orthodontic protocol in functional agreement with basic science. Above all, a biologic protocol should be backed up by long-term results.
Microform Cleft
It has long been observed that a small number of patients have a nasal deformity characteristic of cleft lip but a minimal to absent affectation of the lip itself. The literature on this subject is limited but very consistent. All such patients have asymmetries of the nasal floor/alar base; some degree of septal deflection is always present. The microform cleft is truly the “form fruste” in the problem. It demonstrates most elegantly that the source of the pathology lies within the piriform fossa, not the lip. In addition, the microform cleft demonstrates how differences in the fluid mechanics of fetal ventilation cause deformation of the septum. Of late, I have been able to find more satisfactory answers to these questions.
The Airway in Facial Clefts
Breathing begins in utero. As we shall see, the biologic problems of facial clefts begin with the airway itself, as the deficiency state of the premaxillary field, no matter how slight, causes deformation of the piriform fossa, the nasalis muscle complex, and the vestibular lining of the lateral nasal wall. All other manifestations of cleft in the nose, as seen in the nasal tip, proceed from this triad. What’s more, if the palate is also cleft the nasal airway is compromised by two sites of deformation involving the septum. The achievement of correct nasal function is the Sahara Desert of cleft surgery, a harsh and uncompromising terrain, poorly understood and clinically neglected. No matter how much attention we place on prolabial designs and soft tissue flaps, at the end of the day, the majority of our patients are left with life-long breathing dysfunction on the cleft side. As we shall see, it turns out that basic features of the embryologic features of the cleft nose involving the septum, medial footplate, and the lateral crus are hugely significant. They point the way to understanding the pathology and correcting it completely. Understanding the pathology of the cleft airway and addressing it as an integral part of the initial repair are a major focus of DFR.
Developmental Anatomy of the Midline Lip–Nose Complex
Understanding the development of the premaxilla and its soft tissue coverage, the prolabium is fundamental for cleft lip surgery. Before we take on the issues raised previously, let’s review how the midline structures of the nasal envelope premaxillary platform, the prolabium, and the nose are organized.
Nasal Skin
Let’s return to Chap. 4. Recall that frontonasal skin flows forward and downward in development, forming a bilaminar envelope. Its outer surface is epidermis formed from the anterior prosencephalic folds of hypothalamic prosomeres hp1 and hp2. Its dermis originates from posterior prosencephalic neural crest from prosomeres p1–p3 (probably the latter). The inner lining develops from nasal placodes, in register with hp2. These sink into the mass on either side. In so doing they create two tunnels, the future nasal cavities. The unique vestibular epithelium of the nasal airways is derived from placodal ectoderm, the medial half of which is dedicated to the pheromonal accessory olfactory system and GnRH neurons of nervus terminalis, while the lateral half forms olfactory system. Growing forward into this mass we find extensions of r1 neural crest which are interposed in between the frontonasal layers like cheese in a sandwich. These are responsible for making nasal cartilages.
The nasal chambers wind up as U-shaped structures supplied by posterior and anterior internal nasal and septal branches of the posterior and anterior ethmoid arteries. Its innervation is V1. The nasal floor is constructed from r2 neural crest from the first arch, which is innervated by V2 and supplied by corresponding Stapedial artery related to cranial nerve V2 (StV2) branches from the pterygopalatine fossa. Tessier cleft zones 13–12 are represented within the nasal chamber.
The external nasal skin paired zones of 13–12 along the dorsum and zone 11 along the side walls. These are supplied, respectively, by external nasal branches of anterior ethmoids and by the dorsal nasal artery from the infratrochlear stem.
Columella
Although manipulated and or incised in most cleft repairs the columella has suffered from anatomic neglect. Over the years a vast literature has accrued regarding the components of the nasal tip and their support structures [6] but with little emphasis on the design of the columella. Previously, this author [2] demonstrated the presence of paired anterior ethmoid arteries that transmitted dye into the philtrum but not beyond the philtral columns (Figs. 19.9 and 19.10). As is expected, these vessels are surrounded by r1 neural crest fat filling the space between the medial crura all the way down to the footplates. The combination of fat, medial crura, and skin abutting the septum is the membranous septum. U-shaped incisions allow the upward mobilization of the lateral crus which remains perfused independently from the central columellar. The reason for this is the abundant blood supply reaching the membranous septum from vessels of the internal nasal chamber and rim. Furthermore, the entire philtrum-columella-membranous septum can be readily elevated in a plane anterior to the septum thus elevating the nasal tip, as seen in reports of bilateral cleft report from Trott (which we will discuss later on) (Fig. 19.16).

Trott–Mohan BCL design proved the vascular independence of columella and philtral prolabium. They elevated the entire PP and columella to gain exposure to the nasal tip. This is an extension of the Rethi and Harashina procedures. In general enough elasticity is present in DFR that elevating PP is not necessary. A specific indication for this however is in the insufficient prolabium. In this case PP becomes the columella and the Cupid’s bow is constructed with a graft. This can be later replaced with a composite skin-cartilage graft from the cymba of the ear the curvature of which will mimic the depth of Cupid’s bow. [Reprinted from Trott JA, Mohan N. A preliminary report on one-stage open tip rhinoplasty at the time of lip repair in bilateral cleft lip and palate: the alor Setar experience. Br J Plast Surg 1993; 46;215–222. With permission from Elsevier.]
Blood supply to the skin over the medial crura is quite independent of the columella. The lateral columellar incision described in the technical section an incision below the footplate and the parallel incision in the membranous septum creates a lateral columellar chondrocutaneous (LCC) flap that can be repositioned upward into the nasal tip. It is supplied by a network of StV1 vessels of the dorsum combine by ECA collaterals ascending around the rim of the nostril. This robust blood supply prompted Song to report the use of the columella flap rotated downward into the prolabium in primary unilateral repair. The design transects the columellar arteries to philtrum from above, a sort of Millard-in-reverse. But the technique is worth knowing because it demonstrates the abundant blood supply to the lateral columella (Fig. 19.17).

Lessons of the lateral columellar flap. Song’s variation on the original Reichert procedure (described by Honigmann) violates columellar fields but does illustrate alternative blood supply to the NPP. Skin flap extracted from lateral columella is a random extension of NPP, an idea that originated from Reichert. Both arteries to the philtrum are cut. Blood supply is reverse from the labials. NPP is separated from its vascular axis, becoming random based on philtral prolabium, the blood supply to which is now reduced by 50%. Compare this design to DFR. Pink, medial nasopalatine, magenta, lateral nasopalatine, lemon, anterior ethmoid, orange, ECA facial. [Adapted from Song R, Liu C, Zhao Y. A new principle for unilateral complete deft lip repair: The lateral columellar flap method. Plast Reconstr Surg 1998; 102:1848. With permission from Wolters Kluwer Health, Inc.]
Congenital absence of the columella is a rare occurrence but quite significant. It can occur as part of holoprosencephaly or as an isolated field defect. This condition was first reported by Jacobs [7]. It comes in two forms, total columella loss without and with caudal septum involvement. All cases report absent medial crura. A familial tendency has been documented by both Lewin and Mavili [8, 9] (Figs. 19.18 and 19.19). These cases reveal preservation of two mounds of tissue just overlying the footplates of the medial crura: this is referred to as the so-called “shoulder” of the columella. This tissue originates from r2 and is supplied by a terminal branch of medial nasopalatine artery. The philtrum is preserved. No case of unilateral columellar loss has been reported. This is because the anterior ethmoid arteries are paired into the philtrum; loss of one would be compensated by the other.

Congenital absence of columella 1. Note absence of medial crura but otherwise normal nasal tip with normal projection. Medial crura are not necessary for tip support. Orbicularis is normally inserted into both r1 columella and into the r2 fields below the footplates. In this case the normal sweep upward into columella is lost but lip “drape” remains. Nostril sills are hiding in the nasal floor. They normally blend into the r2 “shoulder” of columella. Columella represents paired independent developmental fields. Medial crus is programmed by columellar skin. Familial tendency (seen in twins). Proves vascular independence from the septum. [Reprinted from Mavalli ME, Akyúrek M. Congenital isolated absence of the nasal columella: Reconstruction with an internal nasal vestibular skin flap and bilateral labial mucosa flaps. Plast Reconstr Surg 2000; 106(2):393–399. With permission from Wolters Kluwer Health, Inc.]

Congenital absence of the columella. (a) Loss of columellar tissue in the center but with residual normal “flow” of columellar tissue into the “shoulders” at the base. (b) a more severe defect in which both midline and basal tissues are absent. Left: [Reprinted from Lewin ML. Congenital absence of columella. Cleft Palate Journal 1985; 25(1):58–63. With permission from SAGE Publications.] Right: [Reprinted from Ayhan M, Sevin A, Aytug Z, Gorgu M, Erdogan B, Reconstruction of congenital and acquired columella defects: Clinical review of 38 patients. J Craniofac Surg 2007; 18(6):1500–1503. With permission from Wolters Kluwer Health, Inc.]
The medial crura are lost in columellar aplasia. This makes an important point: Nasal tip cartilage, although seemingly a continuous structure, consists of distinct developmental subunits. Medial crus belongs to zone 13 and lateral crus to zone 12. They are connected by intermediate crus. For this reason, the number 13 cleft presents as medial alar notch and the number 12 cleft as a defect in the lateral crus.
The philtrum remains intact in columellar aplasia. This may occur because the PNC mesenchyme has reached the level of the lip and is vascularized by collateral circulation from the surrounding lateral lip elements. In the meantime, vascular failure eliminates the columella. I cannot find an example of congenital columellar aplasia in the presence of cleft lip. For example, case should show an abnormal or absent philtrum.
Prolabium
The prolabium refers to the non-mucosal soft tissue coverage of the premaxillae. In normal development it consists of a caudal extension of columella which includes frontonasal skin, anterior ethmoid arteries (usually paired) about 2–4 mm apart, and r1 neural crest fatty-fibrous tissue. This midline structure is known as the philtrum. Its innervation is V1 (Fig. 19.15).
Prolabial anatomy is “more than meets the eye” because the prolabium is a composite of tissues that originate from different locations in the embryo. Under normal conditions the original prolabium fuses with the lateral lip elements. Its lateral embryonic components become tucked inside the nose as the “shoulder” of the columella that gives soft tissue coverage of the lateral incisor. We tend to forget that they exist. Thus, the best way to study the embryology of the prolabium is in its “native” state: the complete bilateral cleft. We will then discuss how the prolabium in unilateral cleft patients contains extra tissues belonging to the first and second pharyngeal arches.
Prior to fusion with the lateral lip elements, the original prolabium looks just like that of the bilateral cleft. It has central PNC elements supplied by the Stapedial artery related to cranial nerve V1 (StV1) anterior ethmoids and lateral elements (the “shoulders”) supplied by StV2 facial. Note that in a bilateral cleft the entire prolabial complex can be elevated and the split into three component parts. In conventional techniques, this elevation proceeds up to the anterior nasal spine because of vascular concerns. In DFR, as a lesson from the Trott and Mohan design, the entire columella and membranous septum (philtral prolabium) can be elevated upward, leaving the columellar shoulders (NPP) in place. This is accomplished by means of a simple incision straight back underneath the alar footplates.
-
Philtral prolabium is PNC
-
Non-philtral prolabium is RNC
-
The prolabium in the embryonic state acquires muscle by fusion with lateral lip elements
-
Bilateral cleft lip repeats the embryonic state and is devoid of muscle
Lateral Lip Elements
Recall that in development first arch and second arch fuse to form a sandwich. This is clearly demonstrated by the structural layers of the lips. Consider the upper lip as a saddlebag suspended from the maxillary complex. Both inner and outer surfaces are r2 ectoderm: externally it forms skin and internally it forms mucosa. Muscles of mastication are dedicated to suspending the jaws and they do not extend into the lip. Thus, there is no layer of DIF fascia in the lips. Into this r2 “envelope” projects second arch myoblasts enclosed within SMAS (SIF) fascia. Deep orbicularis (DOO) is programmed by the mucosa, while superficial orbicularis (SOO) is programmed by the skin. DOO curls upward at the vermilion border where it comes into edge-to-edge contact with SOO. This transition is marked by the white roll (Fig. 19.20).

Lateral lip element contains three muscles. DOO follows the wet mucosa around in the form of the letter “J” with marginal fibers MDOO ascending beneath the dry mucosa. Boundary be between dry mucosa and skin is the white roll. SOO stops short of the white roll. Pit ends proximally with nasalis fibers at the level of the alar base and nostril sill. Nasalis (yellow) in cleft state is located subcutaneously from the nostril sill into the vestibule as far in an inferior turbinate. [Reprinted from Park C, Ha B. The Importance of Accurate Repair of the Orbicularis Oris Muscle in the Correction of Unilateral Cleft Lip. Plast Reconstr Surg 1995; 96(4): 780–8. With permission from Wolters Kluwer Health, Inc.]
All second arch muscles, both those of mastication and those of facial expression, arise from Sm6. The former are associated with deep investing fascia (DIF), while the latter make a primary insertion into the SMAS and then pursue three options (Figs. 19.21, 19.22, 19.23 and 19.24)
-
Facial muscles can remain within the SMAS, inserting into a contralateral counterpart from the other side, and thereby form a sphincter as in orbicularis oculi and orbicularis oris. Frontalis has a secondary insertion into the muscle-free zone of central galea aponeruotica which is stationary, acting like a tendon that permits frontalis to lift the forehead.
-
SMAS muscles can drop downward to insert into bone as in zygomaticus major and minor, nasalis, and depressor septi nasi (DSN). Note that the SMAS over the vertex of skull is devoid of muscle but acts like a tendon, into which insert both frontalis and occipitalis.
-
SMAS muscles can be directed upward to insert into skin, as in corrugator superciliaris.

Superficial musculoaponeurotic system upper lip. Lip and nose anatomy both demonstrate SMAS which conforms to the five criteria of Tessier (see text). Left: [Reprinted from Pensler JM, Ward JW, Parry SW. The superficial musculoaponeurotic system in the upper lip: an anatomic study in cadavers. Plast Reconstr Surg 1985; 75(4):488–492. With permission from Wolters Kluwer Health, Inc.] Right: [Reprinted from Letourneau A, Daniel RK. The superficial musculoaponeurotic system of the nose. Plast Reconstr Surg 1988; 82(1): 48–57. With permission from Wolters Kluwer Health, Inc.]

Nasal SMAS histology. Key: Mallory’s triple stain: 1, hair follicle; 2, sebaceous gland; 3, superficial fatty layer; 4, fibrous septae; 5, anterior ethmoid artery, terminal branch; 6, fibromuscular layer (i.e., procerus muscle); 7. arteriole; 8, epidermis; 9, dermis; 10 SMAS; 11, deep fatty layer; 12, perichondrium. [Reprinted from Letourneau A, Daniel RK. The superficial musculoaponeurotic system of the nose. Plast Reconstr Surg 1988; 82(1): 48–57. With permission from Wolters Kluwer Health, Inc.]

Superficial musculoaponeurotic system (SMAS) of the nose. The SMAS contains the Sm6 muscles of the nose. These have a primary insertion into the fascia and a secondary insertion into soft tissue structures of the nasal envelope. SMAS transmits forces in differential ways due to its flexibility. (Left): Superficial layer of SMAS. The nasal SMAS before its division (black arrow); note its insertion into the internal nasal valve, which appears yellowish (red arrow). The medial superficial layer covers the lower lateral cartilage (blue arrow) and inserts on to the skin of the alar margin. (Right): Deep aspect of nasal superficial musculoaponeurotic system (SMAS). SMAS insertions have been separated at the level of the internal nasal valve (black arrow) and the margin of the nostrils (red arrow). [Reprinted from Saba Y, Amodeo CA, Hammou J-C, Polselli R. An anatomical study of the nasal superficial musculoaponeurotic system: surgical applications in rhinoplasty. Arch Facial Plast Surg 2008; 10(2):115. With permission from Mary Ann Liebert, Inc. Publishers.]

Superficial musculoaponeurotic system (SMAS) of the nose. (Left): Basal view of the nasal SMAS. This layer is penetrated by vascular elements lying immediately beneath the dermis (black arrow). (Right): Superficial medial layer of the nasal SMAS running into the columella. Note the columellar arteries (black arrow) and the relationship between this layer and the fibers of the depressor septi nasi muscle (red arrow). [Reprinted from Saba Y, Amodeo CA, Hammou J-C, Polselli R. An anatomical study of the nasal superficial musculoaponeurotic system: surgical applications in rhinoplasty. Arch Facial Plast Surg 2008; 10(2):115. With permission from Mary Ann Liebert, Inc. Publishers.]
Lower lip development precedes that of the upper lip. Sm6 buccinator from mandible inserts into the modiolus first followed by that from the maxilla. This criss-cross of fibers is important to recreate in cases of lateral clefting in which the vest-over-pants commissure must be reconstructed.
Under normal conditions the lateral lip elements fuse to the PNC mesenchyme overlying premaxilla, bringing three sets of muscles into position, each of which has a distinct fate.
-
SOO stops at the sidewall of the prolabium. It is incapable of penetrating frontonasal mesenchyme. The interface between the mesenchyme beneath philtral skin and SOO results in the formation of a philtral column. Reports in the literature of the philtral columns arising by criss-crossing interdigitation are incorrect. This concept, propagated by Latham, resulted as an artifact of the methodology used. These fibers are not seen in the dissection of unilateral cleft. Once one has pared the cleft side, one sees skin, a fibrous mesenchyme devoid of muscle, and distinct layer of DOO.
-
DOO, on the other hand, follows the r2 mucosa as it covers over the premaxilla, making a sulcus. DOO flows in a plane deep to sub-philtral mesenchyme, achieving continuity with itself across the midline. Note that the branches of facial artery run in the plane between DOO and SOO.
-
DSN is a small muscle associated with DOO, that moves into the midline at anterior nasal spine [10]. The fibers diverge to insert into the preseptal ligament and into the alar bases. Cadaver dissections of DSN show the fibers descending from SMAS into the periosteum below the ANS [11, 12] (Fig. 19.25). This small muscle is important because it can pull down the nasal tip or, on smiling, give the appearance of a transverse crease, the “tight lip.” Clinical approaches to DSN are reviewed by Sinno et al. [13].
-
I am not aware of any reports documenting the presence of DSN in unilateral clefts, but it must surely be so. Barbosa documents an asymmetrical insertion but I am not aware of the circumstances.
-

Depressor septi nasi has primary insertion into fossa above the central incisor. These fibers are often muddled together with those of nasalis in which the triangular muscle complex bears the obscurantist name myrtiformis. [Reprinted from Barbosa MV, Nahas FX, Fereria LM. Anatomy of the depressor septi nasi. J Plast Surg Hand Surg. 2013 Apr;47(2):102–5. With permission from Taylor & Francis]
Premaxilla and Vomer
The neuroembryologic model of cleft remains the same and has been well described elsewhere [14,15,16] and pari passu in this book. In broad brush strokes, the spectrum of unilateral cleft lip alone or in combination with cleft palate results from defects along the axis of the medial nasopalatine neuroangiosome (MNP), the most medial of all the branches emanating from the sphenopalatine fossa. This neurovascular axis supplies two r2 bone fields: the premaxilla and the vomer. Both bones are paired and for the sake of simplicity, I am going to refer to them in the singular (Figs. 19.26 and 19.27).

Anatomy of premaxilla. (Left): Testut’s original rendition of the premaxilla. Upper figure shows right PMx in sagittal section. Lower figure shows left PMx in axial section. Key: gray (PMx) = medial nasopalatine; blue (Mx) = lateral nasopalatine and medial branch from infraorbital to frontal (ascending) process; APm = PMxF, APl + MxF. (Right): Coronal view of piriform fossa: bone fields (left) and angiosomes (right). Note how in the fetal state frontal process of premaxilla (violet/light gray) is in parallel with frontal process of maxilla (blue). Later, MxF overlaps PMxF. Key: gray, PMx and voner/V (medial nasopalatine), dark blue, Mx (infraorbital); lighter blue, inferior turbinate (lateral nasopalatine); red, middle turbinate (ethmoid); aqua, nasal bone (ethmoid and infratrochlear; magenta, lacrimal (infratrochlear). Key (French) Above; premaxiliaire, canal incisive, fossette, incisive, processus naso-frontalis, silion lachrymal, eperon nasal. Below: alviole de l’incisive laterale, alviole de la canine, alveole de dents de remplacement, paquet vaselo-nerveus dans canal incisive, procesus palatine median, procesus palatine laterale, suinure de Albrecht. Left: [Reprinted from Testut L. Traite d’anatomie humaine: anatomie descriptive, histologie, developpement [French]. Paris, France: Gustave Doin, 1899.] Right: [Courtesy of Michael Carstens, MD]

Growth of premaxillary fields. Observe that frontal process grows upward out of the PMxb (here labeled i2). The two frontal processes, MxF and PMxF, are initially separate but fuse. [Reprinted from Barteczo K, Jacob M. A re-evaluation of the premaxillary bone in humans. Anat Embryol (Berlin) 2004; 207(6):417–437. With permission from Springer Nature]
Each premaxilla consists of three distinct zones (from oldest to newest): central incisor, lateral incisor, and a frontal process stretching upward from the lateral incisor all the way to the frontal bone. When pathology strikes the premaxilla, the frontal process is the first to be affected (causing the scooping out of the piriform rim). If the pathology is more extensive, the lateral incisor zone takes the hit. Very rarely, the entire field may be wiped out [17].
Note that the number of incisors in basal mammals is three (in dinosaurs, it was four) (Fig. 19.28). For this reason, it is entirely normal for the medial branch of anterior superior alveolar artery to supply, via a separate branch to the frontal process of maxilla, a third incisor, with its main vessel supplying the canine.

Premaxilla in evolution. (a) Premammal tetrapods had 4 peg-like teeth, as in Tyrannosaurus Rex. Mammals had specialization into incisors. Placental mammals have basal formula of 11 teeth per quadrant with three incisors: I3, C1, P4, M3. (b) Key: i1, central incisor (PMxA); i2, lateral incisor (PMxB); 3, frontal process of premaxilla (PMxF); 4, frontal process of maxilla (MxF); 5, canine eminence (lateral head of nasalia); 6, lateral incisor eminence (medial head of nasalis). (a) [Reprinted from Barteczo K, Jacob M. A re-evaluation of the premaxillary bone in humans. Anat Embryol (Berlin) 2004; 207(6):417–437. With permission from Springer Nature]. (b) [Courtesy Michael Carstens, MD]
Vomerine bones are triangular in shape. Because the vomer sits under the septum, its vertical height anteriorly is very small but, as one proceeds posteriorly, the height increases. Development of the vomer is (1) anterior-to-posterior and (2) dorsal-to-ventral. When pathology strikes the vomer, the posterior height is affected first. The deficient sector of vomer will fail to reach the plane of the palatal shelves.
Defects of the medial nasopalatine neurovascular axis can affect the premaxilla, the vomer, or both, as we shall see below.
The vascular anatomy of medial nasopalatine artery was discussed in Chaps. 14 and 15 but let’s recap the important point. The axis descends along the septum with side branches distributed to mesenchyme such that deficiencies hits the posterior parts of the field first. At incisive foramen the artery flows laterally with the distal zones affected first. These facts explain the spatial sequence of cleft lip and of midline cleft palate involving the vomer.
Premaxillary deficiency always causes a contour deformity of the piriform fossa (the cleft lip nose). Depending upon its severity, isolated cleft lip or cleft lip plus alveolar defect can occur. Vomerine deficiency always affects the hard palate. When associated with premaxillary deficiency, the combination of cleft lip and cleft palate is observed.
The construction of the hard palate involves multiple neuroangiosomes. The intranasal anterior ethmoid and posterior ethmoid supply the perpendicular plate and septum. The medial nasopalatine supplies the vomer and premaxilla. The greater palatine supplies the oral surface of the secondary hard palate: the palatine bone fields P1 and P2 with what we term the palatine bone P3 supplied by the lesser palatine axis. The lateral nasopalatine supplies the nasal surface of the secondary hard palate and the inferior turbinate. Thus, the spectrum of cleft palate is more complex than that of cleft lip alone. Embryologic classification of cleft palate is a subject unto itself and was discussed in Chap. 14. In this chapter, we shall place our emphasis strictly upon the medial nasopalatine axis: the premaxilla and prolabium.
The connection between underlying bone field pathology and soft tissues is as follows. Whenever membranous bones are synthesized, BMP-4 is released. This protein diffuses upward through overlying soft tissues until it reaches the cleft margin. En route, it fulfills 3 functions (Fig. 19.29).
-
BMP4 signals the attachment of nasalis muscle over the lateral incisor and canine.
-
If the signal if absent, nasalis inserts into nostril sill and piriform fossa.
-
-
BMP4 is required for orbicularis muscle development in late embryogenesis (stages 18–23). It holds the myoblasts in a proliferative state until the proper number is achieved and the system can be pass into terminal differentiation.
-
If BMP4 is reduced, muscle fibers in the distal 5 mm of the cleft margin commit prematurely while, in the embryonic state, forming fewer fibers in a disorganized array.
-
-
BMP4 acts on the epithelium where it blocks sonic hedgehog (SHH), a stabilizer of the epithelium. This block leads to epithelial breakdown and mesenchyme fusion between lateral lip and prolabium.
-
Insufficient [BMP4] leads to fusion failure. This mechanism is quantitative and directional. Defects in [BMP-4] affect the extent of downward diffusion. Thus, a minor reduction creates a cleft of the vermillion. The greater the reduction in [BMP-4], the higher the soft tissue cleft ascends [18,19,20].
-

Mesenchymal deficiency = morphogen deficiency. Signaling interactions in the developmental palatal shelves. BMP bone morphogenetic protein, FGF fibroblast growth factor, FGFr fibroblast growth factor receptor, Shh sonic hedgehog. [Reprinted from Carlson BM. Human Embryology and Developmental Biology, sixth edition. St. Louis, MO: Elsevier; 2019. With permission from Elsevier.]
Neuroangiosomes are the functional basis of embryology and of facial clefts. Before we proceed onward with mapping out the prolabium and premaxilla let us get one concept straight. Sensory nerves induce arteries. As the face develops, various families of arteries arise, reorganize, and interact. How this takes place is a fascinating story covered elsewhere.
As we have seen, in the middle stages of development, just at the time of emergence of the cranial nerves, a new stapedial arterial system appears. Its stem is the dorsal remnant of the defunct second aortic arch artery. It traverses the middle ear, where it divides, one part entering the skull and the other part exiting into the face. All branches of the stapedial system, both intracranial and extracranial, are programmed by sensory branches of cranial nerves [21,22,23].
The consequences of this neuroembryology were intuitively grasped by Paul Tessier when he developed his classification system of rare craniofacial clefts. Orbital clefts (zones 10–13) represent individual “knock-outs” of branches from the V1-induced supraorbital stapedial. Maxillary clefts (zones 1–8) represent individual “knock-outs” of branches from the V2 and V3-induced infraorbital stapedial. Clefts in zone 9 are considered the most rare because of its dual blood supply from StV1 lacrimal and StV2 recurrent meningeal. Recall that trigonocephaly may arise in this zone so it is not so obscure as originally thought. Zone 14 does not exist: it merely represents the failure of normal tissue involution (apoptosis) required to approximate the facial midline. Such patients have normal brains but large ethmoid complexes and widened interorbital dimensions [24, 25] (Fig. 19.11).
Having waded through the deep waters of vascular development, we are now in a position to “map out” craniofacial developmental fields using the tools of neuroembryology. Remember, arteries represent inductions from sensory nerves. Thus, prolabium in the bilateral cleft can be “mapped” into four distinct embryonic zones.
-
The philtral prolabium In the center, paired extranasal anterior ethmoid nerves and arteries from the V1 stapedial system run down the columella about 4 mm apart. The anterior ethmoid fields make up the philtrum, that is, the Cupid’s bow. The width of the philtrum = the width of the columella (as defined by the footplates of the inferior crura).
-
The non-philtral prolabium Laterally, additional tissues flank the philtrum. These are supplied by the MNPs, from the V2 stapedial system [26, 27].
The skin and subcutaneous tissues of the philtral prolabium (PP) are unique: ectoderm and mesoderm are not present. Recall that the mature embryonic forebrain is divided into five developmental zones: diencephalon develops from prosomeres, p1–p3, and secondary prosencephalon develops from two hypothalamic prosomeres, hp1 and hp2. Orbitofrontonasal skin has its epidermis from the non-neural ectoderm (not neural crest) of hp1 and hp2. Its dermis arises from p1 to p3 neural crest. Nasal placed contributes a unique form of lining. Take a look inside your nose: the color difference between epithelial nasal skin and placodal vestibular lining is obvious. The boundary is marked by nasal vibrissae.
-
All the remaining facial skin consists of r2–r3 ectoderm and neural crest dermis.
-
One does not encounter true mesodermal dermis until dermatome level C2.
Embryologic Cleft Surgery: Core
Principles of Medial Dissection: Prolabium, Nasal Tip, and Medial Nasal Wall (Figs. 19.16, 19.17, 19.20, 19.30, 19.31, 19.32, 19.33, 19.34, 19.35, 19.36, 19.37, 19.38, 19.39 and 19.40)
Under normal conditions, unification between the premaxilla and the maxilla involves two sets of structures. The frontal process of the premaxilla fuses with its its counterpart, the frontal process of the maxilla. The lateral incisor zone of the premaxilla fuses with the canine zone of maxilla. This unites the MNP with the lateral nasopalatine neuroangiosome. In this way, the soft tissues covering the premaxilla and those of the nostril sill become internalized within the floor of the nose.

David Matthews innovations
improved prolabial incision
• Marking the PP flap wider based on the footplates allows for back-cut directly above point 3. This allows downward displacement of 1–2 mm which is filled with a half z-plasty flap from the lateral lip. Enables a final “cut-as-you-go” strategy for DFR
Non-invasive elevation of medial crus
• Medial crus can be elevated using a “stealth” incision under the alar footplate separates NPP from LCC, making them two independent flaps
NPP–LCC elevated in continuity with vomer flap (V). Septum (S), gold color, accessed from vomer, yellow. Circle denote pedicle to NPP at level of nasopalatine duct. M flap can pared from the NPP or kept with it
[Courtesy of Michael Carstens, MD]

DFR markings. Numbering is based on the original numerical sequence popularized by Millard. The width of the “true” philtral prolabium (P) is the width of the columella at the level of the alar footplates at the tips of the medial crura. Let us call this distance x. Point 2 is the normal/non-cleft PMx philtral column at the white roll. The new philtral column on the cleft side, point 3, is 2-x. Point 1 in the Millard system, the visual “center” of the cupid’s bow, is therefore irrelevant. Points 4 and 10 are the centers of the alar bases on the non-cleft and cleft sides, respectively, as defined by the light reflex. Point 5, the Millard back-cut, is irrelevant. Points 6 and 7 are the commissures. Point 8 is the tentative location of the cleft-side philtral column on the lateral lip element at the white roll. This can be adjusted. Measuring distances 2–6 and 7–8 are rough guides to equality but not terribly useful. The height of the Cupid’s bow is measured from the alar footplates, with point 13 on the non-cleft side and point 12 on the cleft side. Distance 13–2 is the true height of the lip and will equal 12–3 with the addition of the Matthew’s triangle (discussed below). 13–2 will equal 8–9. Point 9 can be marked as the highest point on the skin margin of the lateral lip element. The alar base on the cleft-side is rotated inward and this translates the nostril sill internally. The nostril sill is a triangle defined on the non-cleft side by 4–14, with point 14 being the terminus of the sill. This is usually 3–4 mm. You can take the compass and measure across the sill from point 4 into the nose and find the other leg of the triangle. In similar fashion, the nostril sill on the cleft side can be marked out from point 10 based on the measurements on the normal side. The tip of nostril sill flap, point 11, is inserted at the base of point 12 to re-establish the normal width of the nostril floor. Note: The NPP field (green) is powered by a designated branch of medial nasopalatine located just at the septopremaxillary junction, i.e., at incisive foramen. It is below the footplate of the medial crus. [Reprinted from Carstens MH. Developmental field reassignment cleft surgery: reassessment and refinements. In: Bennun R, Harfin J, Sandor G (ed). Cleft Lip and Palate Management: A Comprehensive Atlas. Hoboken, NJ: John Wiley & Sons; 2016:83–111. With permission from John Wiley & Sons.]

Medial dissection of NPP flap: independent flap vs. composite with LCC. Markings of the both versions of DFR are show here. Here the incision separating the non-prolabium (NP) from the philtral prolabium (PP) sweeps upward along the lateral margin of the columella: open-closed rhinoplasty, a modification of Trott a small counter-incision in the membranous septum helps lift up the medial crus like a “boot strap.” This will leave a 3–4 mm gap. One can replace it by backing up the “shoulder” of the NPP into the deficit at the same time as it is repositioned into the nasal floor. This may require some stretching around the pedicle to gain mobility. Another alternative is to simply rotate NPP on its existing axis and fill the gap with a composite punch graft from the ear. The lower paring incision of the NP flap is carried over the alveolus. If the alveolar cleft is complete, it can reach to the vomer to elevate an optional vomer flap and thus achieve closure of the nasal floor. A lateral nasal incision elevates tissues in front of the inferior turbinate. With the Talmant dissection of the nasalis complex, this lateral design is not useful. There may be situations in which the access gained by the lateral columellar incision justifies its use. The resulting scar behaves well and is inconspicuous. Matthew’s modification brings the incision underneath the alar footplate. It can stop at point 1. Placing curved scissors through the incision beneath the footplate places the surgeon immediately beneath the medial crus. The dissection extends readily up to the tip, where it becomes superficial to the intermediate crus and to the lateral crus. Option 2 follows the alveolus backwards to the vomer permitting elevation of a vomer flap to close the floor of the nose. Complete closure of the nasal floor over an alveolar cleft is critical to avoid an iatrogenic fistula and for successful grafting at age 4. In the Talmant protocol, soft palate closure is achieved at the first surgery but nothing is done with the hard palate. Certain surgical situations, where recall is uncertain, may require closure on the nasal side because it will never be more readily accessible. The extension of the NPP flap incision can be readily carried backward on the vomer to accomplish this goal, leaving completion of the hard palate with mucoperiosteal flap mobilization to a later stage in the sequence. Note that the lateral columellar incision lifts up the entire medial crus and re-sets it into position vis-a-vis the normal side. This maneuver proved highly effective with exceedingly good scar but was superseded by Matthew’s innovation. [Reprinted from Carstens MH. Developmental field reassignment cleft surgery: reassessment and refinements. In: Bennun R, Harfin J, Sandor G (ed). Cleft Lip and Palate Management: A Comprehensive Atlas. Hoboken, NJ: John Wiley & Sons; 2016:83–111. With permission from John Wiley & Sons.]

Access to the nose via anterior columellar incision. Harashina showed a variation of the 1919 Rethi incision which opens the lateral columella in front of the medial crura and includes part of the central columella, transecting it, as in an open-rhinoplasty. Had he proceeded further down into the lip and philtrum he would have isolated the entire StV1 field. Trott and Mohan extended this idea. Perfusion of remaining lateral columella is profuse vessels of the rim and septum. (a) Transcolumellar incision with extension along the rim of the non-cleft side and, on the cleft side, along the caudal border of the alar cartilage. (b) Skin flaps elevated and fibro-fatty tissue elevated away from the nasal tip. (c) Suture fixation of the lower lateral cartilage and cleft-side alar cartilage is advanced. (d) Narrowing of tip with interdomal sutures. (e) Fibro-fatty tissue replaced back over tip. (f) Closure. [Reprinted from Harashina T. Open reverse U incision technique for secondary correction of unilateral cleft lip deformity. Br J Plast Surg 1990; 86:872–881. With permission from Elsevier.]

Trott and Mohan came very close to the open-closed rhinoplasty. This design for UCL demonstrates typical tension with BCL where developmental fields are obvious in their bilateral design the NPP was staring them in the face. If they had only carried over these observations to the UCL repair they might have been able to use a full NPP. Unfortunately the dogma of the Millard “c flap” proved to be a stumbling block to innovation. Note my superimposed DFR incision marks (orange) [28,29,30]. [Reprinted from Song R, Liu C, Zhao Y. A new principle for unilateral complete cleft lip repair, the lateral columellar flap method. Plast Reconstr Surg 1998; 102(6):1848–1852. With permission from Wolters Kluwer Health, Inc.]

Composite NPP–LCC flap and “open-closed” rhinoplasty. Left: NPP–LCC is a small flap with a long name: non-philtral prolabium-lateral columellar chondrocutaneous flap. Right: NPP flap (green) is in continuity with LCC. Vascular pedicle is outlined by black tape. Footplate of the medial crus is in the raw tissue just above the skin of the flap. Flap includes subcutaneous scar which can be trimmed. Note that the NPP flap has been dissected using a lateral columellar incision. The blue tape marks the pedicle. NPP flap is a skin flap. Note here the mucosal extension to the NPP flap. Note: In the cleft state, the prolabium is wider than normal. This is because non-philtral tissue that should be assigned to the premaxilla becomes included. I have therefore considered the cleft “prolabium” to be a composite structure consisting of a “true” philtral component (supplied by the terminal branches of the StV1 anterior ethmoid axis) and a “false” non-philtral component (supplied by the StV2 medial nasopalatine axis). Thus, the incision that runs upward from the lip at point 3 separates the P philtral prolabium, from the NPP non-philtral prolabium. Note: The true position of the NPP is to provide soft tissue coverage over the lateral incisor, that is, the introitus of the nasal floor. The vessels supplying the NPP flap come from the vomer–premaxillary junction. These can be readily visualized, but blunt dissection in the subperiosteal plane is protective. Note here (right) that the NPP flap has been dissected using a lateral columellar incision. The blue tape marks the pedicle. NPP flap is a skin flap. Note here the mucosal extension to the NPP flap. Left: [Reprinted from Carstens MH. Developmental field reassignment cleft surgery: reassessment and refinements. In: Bennun R, Harfin J, Sandor G (ed). Cleft Lip and Palate Management: A Comprehensive Atlas. Hoboken, NJ: John Wiley & Sons; 2016:83–111. With permission from John Wiley & Sons.] Right: [Reprinted from Carstens MH. Functional matrix cleft repair: Principles and techniques. Clin Plast Surg 2004; 31: 159–189. With permission from Elsevier.]

Creation of composite flap NPP–LCC. Original design of NPP–LCC had two incisions: (1) anterior columella into the nose, and (2) membranous septum 50%. Results was a U-shaped “bucket handle” flap. Here we see the separation of non-philtral prolabium (NPP) from philtral prolabium (PP). Note how incision flares around columella until just before the footplate. It then ascends up the side of lateral columella in front of the medial crus and into the nose as far as intermediate crus. To elevate the flap, I used to make a counter-incision in the membranous septum the medial crus. Mobilization around the nasopalatine foramen is blunt to avoid injury to the pedicle. The medial crus is elevated into the nasal tip, bringing NPP with it. At this point, I don’t think the counter-incision is necessary, just good mobilization. [Courtesy of Michael Carstens, MD]

Vissiaronov flap. In cleft surgeries that leave behind the NPP, subcutaneous tissues (buried) are still capable of supporting a full-thickness skin flap harvested from the scar but widening below to include blood supply (now random). This was the basis of the flap described (in Russian) by Vissiaronov. We are indebted to Millard for preserving his work. Lateral columella chondrocutaneous flap (LCC) is elevated as a “bucket handle” with two parallel incisions connected beneath the footplate. But the surprising finding with the NPP scar flap (recall that some NPP tissue may have been discarded at the previous surgery is much more than the skin. Tracing subcutaneous tissue (including the scar) will lead you right up to the pedicle just beneath the footplate. When LCC advances, the “shoulder” of NPP can be “backed up” into the defect, with the remainder of the flap rotated into the nasal floor and trimmed to fit the defect (as there is always an excess of tissue). [Courtesy of Michael Carstens, MD]

Closure of the nasal floor. NPP–LCC elevated in continuity with vomer flap (V). Septum (S), gold color, accessed from vomer, yellow. Circle denote pedicle to NPP at level of nasopalatine duct. M flap can pared from the NPP or kept with it.
• Advantage: open-closed rhinoplasty
• Advantage: no trans-columellar scar as in Harashina; lateral columella scar well tolerated
• Advantage: counter incision in membranous septum behind the medial crus is not required
• Advantage: direct access for mobilization of medial nasal vestibular lining (Talmant 1 maneuver)
• Disadvantage: additional closure time (15 min)
[Courtesy of Michael Carstens, MD]

NPP flap alternative incision design (Matthews). Lateral columella incision and membranous septum incisions are not done. NPP extends proximally in curvilinear fashion just below footplate of medial crus. It puts the access incision to columella above the NPP flap. From here, once again, the NPP flap can be kept continuity with vomer… preserving the pedicle… and used to close the nasal floor above the alveolus by suturing NPP-V to the mucoperiosteum elevated from nasal side of the secondary palate
• Advantage: Talmant 1 elevation of medial nasal vestibular lining can be done
• Disadvantage: open-closed rhinoplasty not available
Key: blue, columella with anterior ethmoid; pink, septum with anterior/posterior septals from ethmoid; orange, medial nasopalatine angiosome, lemon mixture of greater palatine (if continuity with one maxilla) with medial NP
[Courtesy of Michael Carstens, MD]

Lateral columellar incision vs. infra-footplate incision. Subperiosteal dissection of NPP from premaxilla takes you safely right to the pedicle (circle)
• Lateral columella incision: very easy to elevate medial cruse, direct access to septum
• Infra-footplate incision accesses the septum. Elevation of medial crus is blind
NPP-vomer flap = makes use of entire medial sphenopalatine neuroangiosome
• Access the septum from below
• Elevation of medial crus less facile
[Courtesy of Michael Carstens, MD]
Under normal conditions of lip fusion, the two layers of orbicularis oris do not “migrate,” instead they passively accompany the epithelium with which they are associated. Mucosa of the lateral lip fuses with its tr2 counterpart lying beneath prolabium and covering premaxilla. Like a train following a track, DOO migrates medially beneath philtrum. SOO follows different “rules.” It develops in association with r2 skin of the upper lateral lip element (medial infraorbital neuroangiosome). This skin fuses with the frontonasal skin of the philtrum (external anterior ethmoid neuroangiosome). These structures are biologically incompatible. SOO will not penetrate prosomeric mesenchyme. SOO-containing skin is inherently thicker than prolabial skin. The philtral column results from this discrepancy.
The central theme of medial dissection is the separation of embryonic fields with preservation of angiosomes. The philtrum as seen in the unilateral complete cleft philtrum is the ideal model. Bilateral cases are simply a variation on this theme (Figs. 19.15, 19.16, 19.34 and 19.35).
-
Non-cleft side consists of the lateral lip element, a normal philtral column, the r1 columella and its r2 “shoulder” which lies in the nasal floor, and the PP. Note that SOO stops at Cupid’s bow and DOO continues all the way to the cleft side.
-
Cleft side consists of a skin envelope derived from r2: the “shoulder” of columella (which is not truly part of columella), and the NPP.
The non-philtral prolabium develops from hindbrain neural crest. It represents a distal soft tissue extension of the medial sphenopalatine artery, originating as the terminal branch of the internal maxillary system in the sphenopalatine fossa. Under normal conditions, NP represents the skin tissue coverage of the medial the nasal floor. Tucked inside the nostril sill, it is difficult to appreciate. The reason NPP is displaced into the nasal floor is because the lateral lip element is inserted between PP and NPP.
-
In cleft lip patients, NPP is externalized and readily seen, being a lateral “add-on” to the true philtrum. The abnormal fusion is due to failure of the lateral lip element to reach the midline. Because NPP remains connected to the nasal floor, it draws the cleft side philtrum upward, giving the impression of “shortness.”
-
Surgical separation of non-philtrum from the Cupid’s bow makes use of this embryonic fusion plane. NPP is reassigned into the nasal floor and medial nasal wall nasal floor. It joins with tissues from the alar to recreate the missing nostril floor anterior to that of the mucoperiosteum. NPP reassignment releases the medial crus and expands the airway. Simultaneously, the ipsilateral philtrum is dropped into position. Modifications of the DFR incisions based on the contributions of Matthews and Talmant make the surgical design embryologically accurate and (not surprisingly) give significant functional improvement.
Medial Dissection: Modifications
Ten years ago, I met David Matthews at the ASPS in Orland, Florida. Over the years we have developed a close professional cooperation. We found that, from a technical standpoint, our dissections were very similar. Several years ago, Dr. Matthews began to incorporate the NPP flap into the nasal floor. However, when we compared long-term outcomes my cases demonstrated a flattening of the Cupid’s bow at the junction of the white roll and the cleft-side philtral column, whereas my colleague’s patients maintained eversion all along the Cupid’s bow. Clearly, there was something amiss with the design of my straight-line incision as it approached the caudal margin of the Cupid’s bow (Figs. 19.30, 19.31 and 19.32).
-
Matthews #1: The answer was the preservation of the uppermost fibers of the deep orbicularis oris by means of a small back-cut incision that drops the muscle into proper position with respect to that of the non-cleft side. Technical details of this modification will be described below.
-
Matthews #2: I also observed that the proximal limb of Matthews’ NP incision could be nicely carried into the nasal floor just beneath the footplate. The trick was to incise just the skin, leaving the subcutaneous pedicle powered by intact MSP artery branches. The resulting NP flap rotates perfectly into the lateral nasal incision. Furthermore, the incision permits access to the internal surface of the medial crus. One simply inserts the curved scissors inward, locating the internal surface of the medial crus, and elevates. From the very moment I saw his approach it was obvious to me that the misplaced medial crus could be corrected perfectly well without an external incision. Simple is always better.
The embryologic implication of Matthews’ incision is that the true width of the PP was not (as I had previously thought) merely the width of the columella alone. Instead, PP equals the distance between the alar footplates. This adds 2–3 mm to the width of the Cupid’s bow along the white roll. As we shall see, the terminal point of Matthews’ back-cut is aligned with the old definition for PP.
Medial Dissection: Embryologic Lessons, Access to the Nose
A side benefit of Matthews’ critique of the DFR incision design (not the concept) was that it forced me to rethink the embryologic boundaries between the columella, the nasal floor, and the lip. Landmarks were needed to define the junction between frontonasal skin (of forebrain origin) and facial skin (of hindbrain origin). To illustrate this point, we must discuss the embryologic origins of body skin, facial skin, and frontonasal skin. All three arise in a radically different manner. Here is a quick and dirty summary. Armed with this information, we can understand how the developmental fields of the lip and nose are laid out.
Body skin arises from two tissue sources. Epidermis comes from ectoderm, whereas dermis is mesoderm. During gastrulation (the creation of a three-layer embryo) the mesoderm becomes segmented into somites. Each somite is supplied by a sensory nerve, the nucleus of which lies within the corresponding developmental unit of the CNS: the neuromere. These tissues are in genetic register with each other. T3-innervated dermis is genetically connected with the third thoracic neuromere. Each “swatch” of mesoderm is “assigned” to a zone of overlying ectoderm; together they share a common genetic definition. This skin unit is what textbooks refer to as a “dermatome” (sic). Precisely where does body skin begin? The alert reader may ask, “Why is there no C1 dermatome?” All somites contain dermatomes, but those of the first four occipital somites and the first cervical somite are unstable and degenerate. Thus, the body skin (dermatome-derived dermis) does not appear until the second cervical somite.
Facial skin arises from two different sources as well. The epidermis comes from ectoderm but the dermis arises from neural crest. Once again, neuroanatomy comes to our rescue. Facial skin is innervated by V2 and V3, whereas frontonasal skin is innervated by V1. Since the V2 nucleus resides within the second rhombomere, it makes sense that the neural crest dermis of maxillary division skin arises from the neural fold just above r2. These tissues share a common genetic “signature.” Mandibular dermis arises from the r3 neural crest.
Frontonasal skin is utterly different: it arises from a single source. Here, specialized non-neural ectoderm of the neural folds overlying the secondary prosencephalon (hp1 and hyp2 brain) gives rise to all frontonasal epidermis except vestibular lining of the nose. This specialized epithelium contains neurons and arises from the nasal placode which lives within the anterior forebrain neural fold. The source of dermis for frontonasal skin if neural crest from prosomeres p1–p3 encoding the diencephalon.
For readers new to the subject of neuromeric mapping, the above description probably seems abstract. Let’s convert this to anatomic terms understandable to all surgeons, that is, to neuroangiosomes. Vestibular skin (hp2 placode) and fronto-orbital-nasal skin (hp1–hp2) are innervated by V1. Arterial supply to both these regions is from branches of the V1 stapedial branches of ophthalmic axis. Vestibular “skin” consists of septal mucoperichondrium and lateral nasal wall mucoperiosteum. These tissues are supplied by the posterior and anterior ethmoid arteries, both of which send out nasal branches to their respective targets. External nasal envelope skin is supplied by terminal branches of the anterior ethmoid. These exist from beneath the nasal bones and run downward to supply the distal (non-vestibular) internal nasal skin and the columella. The skin lying immediately beneath the alar footplates belongs to a separate developmental field, the r2 NPP, supplied by the medial nasopalatine axis.
Cartilages of the nose develop when neural crest mesenchyme comes in contact with, and is “instructed” by an epithelial program. The size and shape of the upper lateral (triangular) cartilages are vestibular lining of zone 12, whereas the contours of the lower lateral (alar) cartilages are fixed by interaction with zone 12 skin. The medial crura are programmed from nasal skin of zone 13.
-
The footplates of the medial crura are the landmark for where skin ends and where r2 upper lip skin begins.
Freeing the Medial Crus: Lateral Columellar Incision
The whole idea of the DFR incision is field separation. In the original iteration of DFR I took the prolabial separation incision right up the side of the columella and combined two unrelated neuroangiosomes: r1 frontonasal + r2 medial nasopalatine. The flap was large: NPP skin (medial nasopalatine) in continuity with the entire ipsilateral columella (anterior ethmoid). It merited an equally awkward term: NPP–LCC (lateral columella chondrocutaneous) flap. This design did not achieve embryologic field separation, but it seemed to work. I rationalized this compromise by convincing myself that the downward displaced, to be repositioned correctly, the cleft-side medial crus needed a surgical release (Figs. 19.35, 19.36, 19.37 and 19.38).
Freeing the Medial Crus: the Infra-Footplate Incision
This concept proved to be unnecessary. Correct separation of PNC and r2 RNC skin takes place just below the footplates. This permits elevation of the media crus from below just as effectively as the lateral columella incision with shorter time of closure. The incision, just through the skin, permits gentle spreading of areolar tissue. It automatically puts the scissors inside the columella; by hugging the medial crus one can advance right the way up into the nasal tip with virtually no bleeding. Finally, the infra-footplate incision puts one safe above the medial nasopalatine pedicle permitting safe dissection of the NPP flap (Figs. 19.39 and 19.40).
Principles of Lateral Dissection: Lip–Nose Muscles, Nasal Dorsum, and Lateral Nasal Wall (Figs. 19.41, 19.42, 19.43, 19.44, 19.45, 19.46, 19.47, 19.48, 19.49, 19.50, 19.51, 19.52, 19.53, 19.54, 19.55, 19.56, 19.57, 19.58, 19.59, 19.60, 19.61, 19.62, 19.63, 19.64, 19.65, 19.66 and 19.67)
Wide Subperiosteal Release
This is one of the first principles I learned that comes directly from work of Delaire and Soterianos [31]. What I did not understand 20 years ago was that this was really a case of functional stem cell transfer. We shall recount its history and literature.

Jean-Claude Talmant: cleft surgery must restore airway. His brother demonstrated movements of the nasal cartilages with fetal breathing. The nasal chamber expanded and contracts with the flow of amniotic fluid. Distortions of airway diameter affect development of nasal envelope and, in particular, the shape of the septum. [Courtesy Dr. Jean Claude Talmant]

Inferior turbinate flap (after Noordhoff). Very useful “patch” for lateral nasal wall. I did not recognize extent of mobilization until the Talmant 2 maneuver. Simple excision, spreading, and turbinate flap did not give reliable advance of the alar cartilage. At the present time, with the extensive mobilization, the deficit created sometimes not reliably covered with NPP. Important not to tighten the airway. Thus, turbinate flap becomes relevant again. [Courtesy of Michael Carstens, MD]

Consequences of conventional surgery: two problems. Traditional approaches to cleft rhinoplasty include lateral approaches to the lateral crus via the ala, via vestibular incisions. These can be combined with medial approaches, such as Rethi incision. Here we see two parallel incisions, pre-cartilage and intercartilage to “free” the alar cartilage to reposition in into the tip. Recall that the cartilage is “programmed” by the vestibular lining. Thus, freeing the vestibular lining for overlying soft tissues (Talmant) accomplishes lasting alar cartilage stability. Failure to do so results in
• Tip definition compromised
• Relapse of the lateral nasal wall
• Improper nasal valve function
The nasalis complex (red) is not reassigned to its normal insertion site (dotted lines) but transferred to the midline where it acts as a constrictor of the airway, as times seen as paradoxical motion of the ala with forced inspiration
• Constriction of the airway
• Vestibular “web”
[Courtesy of Michael Carstens, MD]

Alar anatomy: cleft surgery must restore airway. Soft triangle defined by gap between compressor and pars alaris. Nostril sill is that segment that connects ala with “shoulder” of columella. It corresponds to Simonart’s band between LNP and MNP. 1a, nasalis pars alaris (dilator) inserts above lateral incisor; 1b, nasalis pars transversus inserts into canine eminence; 3, anomalous nasi (dilator) is part of nasalis; 4, anterior dilator nasi is functionally part of pars alaris; 5, compressor nasi; 6, levator labii et alaque nasi); 7, oblique fibers of Delaire from SOO are superficial to insertions of nasalis and; 8, depressor septi nasi (no numbered) is deep so SOO, inserts over central incisor, and is shown inserting into the columella. Note pars transversus of nasalis is often labeled as a compressor but, acting as a chain attached to accessory cartilage and thence to the canine, is really a dilator. When surgically mal-inserted into the midline, it acts as a constrictor and a powerful constraint on the lateral crus. [Reprinted from Oneal RM, Beil J. Surgical anatomy of the nose. Clin Plast Surg 2010; 37(2): 191–211. With permission from Elsevier.]

Myrtiformis. Key, (left): A alar margin, C canine, D inner fibers of myrtiformis (act as a depressor), E outer fibers of myrtiformis (resemble myrtle, its berries or leaves). Myrtiformis act as both (1) a depressor/stabilizer of the columella and ala and (2) via SMAS as nasalis, (both pars alaris and pars dorsalis) as a dilator of the airway. Key, (right): A alar margin, N nasalis, L labial portion of levator labii et alaque nasi; O orbicularis, M inner fibers of myrtiformis (oblique fibers of Delaire from SOO?) [Reprinted from Figallo EE, Acosta JA. Nose muscular dynamics: the tip trigonum. Plast Reconstr Surg 2001;108(5):118–1126. With permission from Wolters Kluwer Health, Inc.]

Vestibular web is due to nasalis. Left: The mal-inserted nasalis exerts a “bowstring” effect. Right: Insertion into piriform fossa of deep head, pars transversus, is shown by the cotton tip applicator. Note: If you look under the skin of the nostril sill, just where it flows into the lateral lip, you will see fibers of pars alaris. They are the same thickness as SOO (0.5 mm) and are 1–1.5 cm wide. After dissecting the superficial plane of pars alaris, one dives beneath the muscle (while staying on top of pars dorsalis). This plane takes you directly to bone. Slip the elevator inside the piriform fossa and elevated pars dorsalis. It can be “delivered” from the fossa. The two heads are sutured as one unit above the canine. With release of nasalis, the “web” goes away. [Courtesy of Michael Carstens, MD]

Lateral dissection 1. Lateral dissection opens the lip and separates: (1) the skin from the superficial orbicularis—about 5 mm; (2) the superficial orbicularis (SOO) from the deep orbicularis (DOO); and (3) the nasalis from both SOO and DOO. NB: actual separation of the nasalis is most easily accomplished after the muscle has been completely dissected out because it can be tensed away from the lateral lip element. Recall that the nasalis has two heads, both of which become displaced in the cleft condition. The superficial head should insert over the lateral incisor but instead becomes stranded beneath the nostril sill. The deep head should insert over the canine but instead it falls inside the piriform rim and attaches low on the wall. Dissection of the nasalis proceeds in two planes. First, the skin of the nostril sill is elevated off the muscle. Dissection is then extended up into the nose undermining the vestibular mucosa off the muscle, proceeding in front of the inferior turbinate, and extending all the way up to the edge of the lateral crus. Next, while grasping the muscle, one proceeds deep to it, separating it from the orbicularis and going directly to the rim of the piriform fossa. One then proceeds inward and, hugging the inner wall of the piriform rim, one elevates the deep head off the floor of the piriform. A McKenty or a Molt #9 elevator can prove useful as one “fishes out” the muscle from the piriform fish tank. Note here that the superficial plane of the nasalis (pink arrow) is dissected by proceeding just beneath the skin of the nostril sill and thence upward beneath the vestibular lining until one achieves the edge of the lateral crus. The deep plane of the nasalis (blue arrow) is achieved by following the mucosa down to the piriform fossa where one proceeds in the subperiosteal plane along both its internal aspect and upward to the nasal bones. [Reprinted from Carstens MH. Developmental field reassignment cleft surgery: reassessment and refinements. In: Bennun R, Harfin J, Sandor G (ed). Cleft Lip and Palate Management: A Comprehensive Atlas. Hoboken, NJ: John Wiley & Sons; 2016:83–111. With permission from John Wiley & Sons.]

Lateral dissection 2. Field separation, sagittal first step. Height of the lateral lip (8–9) = cleft-side philtral column (3–12) = non-cleft-side philtral column (2–13). The lateral lip incision ascends from point 8 to point 9 which is tucked just inside the nostril rim. Alar base, point 10 and tips of nostril sill flap, point 11 are clearly seen. From 9 the incision then darts lateral behind and in parallel to the nostril sill. It then ascends for about a cm just in front of the inferior turbinate. The triangular nostril sill flap, the apex of which is point 11, is of the same dimensions as that of the non-cleft nostril floor (or slightly greater for overcorrection)
• Superficial orbicularis (SOO) and deep orbicularis (DOO) both in green are separated in the fatty vascular plane (magenta). The two heads of nasalis (yellow and tan) are released from orbicularis by
• Dis-insert pars alaris (yellow) from vestibular lining
• Pars alaris is intermingled with the upper fibers of orbicularis for about 1 cm
• Just transect OO away from ala
• Dis-insert pars dorsalis/transversus (orange) from internal surface of piriform fossa
• Note: pars transversus is positioned behind pars alaris. To get at it you must first reflect up the more superficial muscle
[Courtesy of Michael Carstens, MD]

Lateral dissection 3. Field separation, sagittal, second step
• Dis-insert pars alaris (yellow) from skin of nostril sill and extend up under vestibular lining past the inferior turbinate
• Dis-insert pars transversus (orange) from internal surface of piriform fossa, above half-way
Key: The skin/mucosa envelope of lateral lip element are shaded. DOO is thicker than SOO. Muscles separated by fat plane with axial vessel running the length of the flap. PP are in gray. SOO ends about 5 mm below the alar crease. Nostril sill flap (point 11) is separated from lateral lip margin (point 10)
Pars alaris (yellow) required two planes of dissection. In the superficial plane it is separated from upper margin of SOO and followed up hard under the skin and vestibular lining. The deep plan goes between SOO and DOO down to bone. Holding the muscle with a forcep one follows it right the way around the outside of piriform, hugging the bone. One will encounter the previous subperiosteal plane
Pars transversus is elevated from the inner surface of piriform. It can readily be grasped and extracted. The two heads are gathered together and secured to the sulcus at the canine with a 4–0 vicryl or PDS mattress suture
[Courtesy of Michael Carstens, MD]

Nasalis complete cleft: dissection and reposition
• primary cases, NPP in continuity with vomer flap
• secondary cases NPP developed separately including scar
[Courtesy of Michael Carstens, MD]

Nasalis, incomplete cleft. DFR ± Talmant technique: In this case of primary CP only, nasal floor scooped up from depressed bone with optional alveolar augmentation using ICBG, Osteocel®, rhBMP2. (Left): Lip incision shown curving around footplate and descending to just above white roll where Matthews back-cut is planned. If height adjustment is needed It will to receive a half z-plasty at the conclusion of the procedure. Lateral incision from 8 to 9 is seen turning lateral behind the nostril sill. (Right): Forcep (left) tractions well defined nasalis. Undermining of skin for 5 mm demonstrates SOO, the fee border of which is hanging down. NPP is suspended from its pedicle. [Courtesy of Michael Carstens, MD]

Subperiosteal dissection external to the piriform fossa (Delaire). Left: Wide subperiosteal dissection of the maxilla and nasal bone (black arrows). Dissection is carried out immediately after the superficial plane dissection of nasal skin via the rim incisions. Right: The next step, deep plane dissection, takes one directly down to the bone of the piriform rim. From that point the maxilla is widely freed up, sometimes if more mobility is needed, with a counter-incision, going up the buttress. Subsequently the elevator is brought along the external surface of the maxillary frontal process, all the way to the nasal bone. It is then brought downward and forward, passing bluntly into the nasal tip, as described by Tessier. Note that the plane passes below pars dorsalis of the SMAS (orange). [Courtesy of Michael Carstens, MD]

Subperiosteal dissection internal to the piriform fossa (Talmant). Left: Talmant dissection inside the piriform is bidirectional (orange arrows). It frees the entire internal surface of frontal process of maxilla. It sweeps upward to the nasal bones and then pulls downward and forward, stretching the lining. This causes (1) passive repositioning of the cartilages and (2) expansion of the airway. Right: Vestibular lining (pink, StV2 and white, StV1) is completely mobilized with Talmant 1 freeing the medial wall and Talmant 2 freeing the lateral wall. [Courtesy of Michael Carstens, MD]

Lateral nasal wall subperiosteal dissection: lengthening the nasal lining as elevator descends it dislodges “scroll” of upper lateral cartilage from inferior margin of nasal bone. Orange nasalis pars transversus yellow nasalis pars alaris. [Courtesy of Michael Carstens, MD]

Lateral nasal wall dissection: variations (Trott and Talmant). (Left): Trott and Mohan design for mobilization of the lateral nasal mucosa (indicated in gray stipple). Talmant elevation is in yellow. (Right): I could never get the lateral nasal wall right until I became aware of Talmant’s findings. Formerly, I had tried lateral crus elevation using a V–Y incision, broad subperiosteal freeing, and so on, all to no avail. Nasalis dissection is key but another maneuver worked out by Talmant is very helpful. Using a long curved elevator, one proceeds inside the piriform rim and frees it all the way to the nasal bones. Then you sweep downward—always in the subperiosteal plane—until you reach the nasal mucosa underlying the alar cartilage. Now, you do exactly the same maneuver outside the piriform fossa—again in the subperiosteal plane—until you reach the nasal bones and then proceed downward bluntly separating the SMAS muscle layer from the underlying upper lateral (triangular) cartilage and thence to the lower lateral (alar) cartilage. The alar cartilage will be nicely mobilized. You will also fall into the dissection plane you previously created by the medial dissection with scissors up the columella. Recall Tessier’s concept of blunt tissue dissection following embryonic planes. These maneuvers are simple and quick but a nicely sharpened elevator is a must. Left: [Courtesy of Michael Carstens, MD]. Right: [Reprinted from Carstens MH. Developmental field reassignment cleft surgery: reassessment and refinements. In: Bennun R, Harfin J, Sandor G (ed). Cleft Lip and Palate Management: A Comprehensive Atlas. Hoboken, NJ: John Wiley & Sons; 2016:83–111. With permission from John Wiley & Sons.]

Open-closed rhinoplasty: elevation of the nasal tip
Step 1: mobilization of the medial and lateral crura
Step 2: Elevation of the medial crus by 5–0 PDS sutures to the contralateral side. Alar cartilage repositioning
Step 3: Achieving definition of the nasal tip
• Intercrural suture at the inflection between the medial and intermediate crura
• Interdomal suture to align the dome on the cleft side with the non-cleft side
• Suspension sutures placed to septum or upper lateral cartilage, depending on the in situ geometry. NPP is now ready to be placed into the nasal floor. If it has been raised with anterior vomer flap the closure is more extensive
Note: In the case of complete cleft palate, the width of the cleft may be considerable. The use at primary lip–nose repair of a concomitant soft palate closure will result in an inevitable narrowing of the secondary palate defect, making vomer flap closure easier. At definitive palatoplasty, access to the nasal mucoperiosteal flap when the lip is closed remains quite simple. A small incision in the vestibular wall in front of inferior turbinate will allow one to slip in a narrow periosteal elevator. Proceed straight backward, hugging the bone. Now sweep medially until one reaches the edge.
Closure of the nasal floor is essential to block oronasal fistula
[Courtesy of Michael Carstens, MD]

Open-closed rhinoplasty: nasal tip suture fixation. Medial crus elevation and stabilization to the septum (which has been mobilized)
• Medial crura joined (red sutures) and fixed to septum (green sutures)
• Oblique head SOO attached to base of columella (arrow) > fixation of septum
Nasal tip control: interdomal suture to narrow the angle of domal divergence (2), intercrural fixation to elevate cleft-side tip defining point (3), and suspension of lateral crus from the contralateral upper lateral cartilage or septum (4)
Restoration of the nasal floor
[Courtesy of Michael Carstens, MD]

NPP flap augments nasal floor; can be combined with vomer flap. NPP flap augments nasal floor; can be combined with vomer flap. Optional nasal floor closure if NPP combined with vomer flap. NPP-V flap (pink) has two parts NPP flap is rotated 90° and “fits into” the vomer flap
• Liberate mucoperiosteum on nasal side of the secondary palate using anterior approach in front of vomer
• Unification of neuroangiosomes = medial SP (NPP and vomer) + lateral SP (inf turbinate)
• MSP (pink), LSP (gold), greater palatine (yellow), infraorbital (dark orange), anterior ethmoid (light orange)
Left: Dorsal view of the lip–nose complex showing un-elevated NPP flap in situ
Right: Oral view showing anatomic continuity between NPP and the anterior vomer flap
[Courtesy of Michael Carstens, MD]

Nasal floor closure: how to harvest the lateral mucoperiosteal flap. This brings me to a final detail regarding the lateral wall that is important if one wants to achieve a hermetic closure of the nasal mucoperiosteum. As we all know, elevating the lining away from the nasal/dorsal aspect of the palatal shelf can be challenging. Two maneuvers make this easier and safer. First, after making the incision along the edge of the palatal shelf, a small dental amalgam packer can be used to elevate off all fibers of periosteum. The head of this instrument is angulated and delicate. It can nicely lift up the edge without tearing it. Second, the incision to elevate the triangular nostril sill flap places one directly in front of the inferior turbinate—in the subperiosteal plane. Using a periosteal elevator or the amalgam packer, one proceeds straight backward beneath the turbinate, elevating the mucoperiosteum away from the vertical wall of bone. Then, at the horizontal palatal shelf, one simply proceeds in the same plane, elevating lateral to medial until one gains the cut edge of the palatal shelf. Hard palate mucoperiosteum
• Nasal side + inferior turbinate = medial sphenopalatine neuroangiosome (yellow)
• Oral side = greater palatine branch of descending palatine (lemon)
[Courtesy of Michael Carstens, MD]

Nasal floor closure: anterior approach to the nasal layer of palatal mucoperiosteum. Anterior sub-turbinate approach elevates mucoperiosteum from nasal side of P1 (). NPP flap with extension to the vomer (pink) reaches easily. Note the L-shaped closure as seen in Fig. 19.58b. Maxillary lamina of alveolus and the face of maxilla (orange) are supplied by infraorbital. Dedicated branch from medial IO toe zone 4 supplies frontal process of maxilla (not seen here). [Courtesy of Michael Carstens, MD]

NPP inset into lateral wall. (Left): NPP flap is lifted up on its pedicle. It is 3 mm wide. Left nostril sill has not been rotated outward. (Right): nostril flap out-rotated, leaving tissue deficit behind it. NPP flap (forcep) fits into the slot, permitting nostril sill to be exteriorized. [Courtesy of Michael Carstens, MD]

NPP inset and nasalis. Another case shows NPP flap fitting into the release of nostril sill. (Left): Nasalis seen below nostril sill. It is clearly separated from SOO. PDS or Vicryl suture about to be placed through sulcus at the level of the canine, being passed in mattress fashion through the bulk of the nasalis and then returned to the oral cavity, where it is tied at the base of the nose at the end (literally) of the case. (Right): roomy nostril and good nostril sill seen at closure around the Talmant stent. Note cleft-side nostril overcorrected. [Courtesy of Michael Carstens, MD]

Final closure of the nasal floor +/− back wall of alveolar cleft. On the left NPP (pink) is shown achieving closure behind the release incision of the nostril sill, medial branch of infraorbital (orange). This is frequently not possible, necessitating mobilization of LNP nasal flap (gold) seen on right from the opposite side. If one wants to close back wall of the alveolar cleft another option is to include the anterior sector of the vomer flap as a mucoperiosteal extension of NPP. After advancement of the vestibular lining, with proper mobilization (using Talmant technique) no residual raw surface remains (as seen on right). If a raw surface is present it can be patched using a turbinate flap or a skin graft (full thickness or composite) from the back of the ear. [Courtesy of Michael Carstens, MD]

Muscle closure: DOO and SOO. Left: NPP flap placed into the floor of the nose to close the upper margin of the “box.” Right: DOO (brown) is sutured first to the non-cleft side at its highest point just below anterior nasal spine. This suspends the lip. SOO is sutured to the mesenchyme of the prolabium. Recall that in unilateral clefts contralateral SOO does not extend past the philtral column. Sm6 muscle cannot penetrate r1 neural crest mesenchyme. [Courtesy of Michael Carstens, MD]

Orbicularis repair: what not to do. Muscle repair is nuanced as per the individual surgeon. I think the key factor is anchoring DOO to its counterpart right under the nasal spine. This serves to set the height of the lateral lip flap. SOO is repaired to the subdermal tissue high in the flap. The upper corner (fibers of Delaire) can be sutured to the base of the columella to see if an improvement in the “aesthetic drape” of the lip is obtained. Remember that SOO is superficial to nasalis. This case was performed before I learned about nasalis re-insertion. I include it here because it shows a very common MISTAKE of lip repair. (Left): Nasal stents not yet placed. The gingival flaps are being turned inward from the cleft margin. NPP flap is sutured into place in the nasal floor. Just under the skin of the lateral lip element you can see muscle. The uppermost 1–1.5 cm is nasalis (here left in situ with orbicularis. The subcutaneous plane for freeing nasalis from the nostril sill is obvious. (Right): Here you can see the fibers under the nostril see sutured to the columellar base. Do not do this. Suture nasalis to just above the canine with a mattress that catches the periosteum and the sulcus. The knot is in the mouth. Do not tighten this or you will depress the alar base. Just gently approximate it and let nature do the rest. This is the final step in the DFR procedure. [Courtesy of Michael Carstens, MD]

Oblique fibers of Delaire. This final suture can help the aesthetic “drape” of the lip. It is placed as a mattress into the upper margin of SOO through in incision to free nostril sill about 1–1.5 cm back from the leading edge of the muscle. It can be mattressed into the base of the columella. 5–0 PDS glides nicely through the tissue permitting you to see if it accomplishes an improved suspension. If not, simply remove it. [Courtesy of Michael Carstens, MD]

Talmant nasal stent. Use 0.5 mm silicon sheeting and appropriate glue. Let dry overnight. These are also commercially available. Note: Cases 1–6 are primary DFR repairs using followed for 5 years. Case 7 is a primary repair using rotation-advancement and revised with DFR with results shown at 1-year post-revision. Case 0 began this chapter and is followed demonstrating DFR correction of multiple secondary problems at 1-year post-revision. [Courtesy of Michael Carstens, MD]
Separating the Orbicularis Layers in the Lateral Lip Element
DOO and SOO insert differently. DOO needs to extend further forward to the midline to joint its opposite just under anterior nasal spine. SOO, on the other hand, has a different vector and is sutured to the mesenchyme of the PP. Paring of the mucosal border exposes these two layers and the artery that runs downward between them to the vermilion border. Naturally, the vascular axis is surrounded by adipose tissue which can be gently spread in the upper half of the lateral lip to gain mobility between the layers. The vertical incision running upward along the anterior border of the alveolar cleft provides access to the subperiosteal plane for wide mobilization. A counter-incision through the mucoperiosteum running up the buttress can be used as needed to convert the entire lip–cheek unit into a giant bipedicle flap.
Management of the Lateral Nasal Wall: Trial and Error
The Vestibular “tightness”: A Case of Misunderstood Mismatch.
In normal patients, the lateral nasal wall just in front of the inferior turbinate contains the soft tissues and bone of the premaxillary frontal process. In virtually all cleft patients this zone appears contracted. A prominent vertical fold or web is seen running downward from the distal margin of the lateral crus and terminating at the palatal shelf. My initial concept of the cleft nasal deformity was that the vestibular lining was somehow deficient. This was very much influenced by the work of Markus, Delaire, and Precious (more on this later). The deficiency site could be readily “patched” with an inferior turbinate flap, rotated 180° forward. This became part of what I considered the “process-oriented” cleft rhinoplasty [32]. Alternatively, I tried filling in the deficiency site with composite skin-cartilage graft from the ear. The cartilage was intended to prevent contraction and relapse. Despite stenting, I was disappointed in the results. As it turned out, the model was wrong and I had not yet stumbled on the developmental field concept.
Upon discovery of the PMx subfields I thought I has found a better solution. Absence of the PMx subfield seemed to explain the entrapment of the lateral crus. After working out the neurovascular anatomy of the prolabium in 2000, it seemed logical that this problem could be addressed by wide subperiosteal release, mucosal release incision, and addition of the NPP flap into the defect. As an adjunct, the nasal airway was supported by postoperative stenting for up to 3 months.
By 2010 I was convinced that this idea was insufficient. Although the initial release seemed to create a nearly perfect airway, in many patients the expansion was unstable. The nostril sill was better but airflow in these patients continued to be reduced compared to the normal side. Furthermore, on physical examination, I started to note a dynamic component to the cleft airway. Watching unilateral patients breathe from below and from above demonstrated an asymmetry of alar movement. Despite the addition of adequate soft tissues, forceful inspiration would be accompanied by an inward contraction. It was time to return to the drawing board: Gray’s Anatomy and the work of Jean Claude Talmant (Fig. 19.41).
Reassignment of the Nasalis Muscle and Vestibular Lining
I was fortunate to meet Dr. Talmant in 2005 as invited faculty for the Indian Society for Cleft Lip, Palate and Craniofacial Anomalies in Guwahati, Assam. His emphasis on the nasal airway as a key to maxillary growth made biologic sense and his results were impressive. His concepts are well summarized in “Evolution of the functional repair concept for cleft lip and palate patients,” published in IJPS 2006; 39(2):197–209. This article should be required reading for all cleft surgeons. This being said, I must confess that my initial review of Talmant’s work was too superficial. I underestimated the physiologic significance of the nasalis muscle and I missed its embryologic relationship to the frontal process of the premaxilla [33].
Four years later, Dr. Talmant’s work came to my attention once again via David Matthews. After visiting Talmant in Nantes he brought back three key ideas: (1) reassignment of the nasalis muscle to its correct position; (2) subperiosteal release and the internal nasal lining; and (3) effective techniques to correctly reassign the nasal lining using packing and a much better technique for airway stenting. These three maneuvers made instant biologic sense. Cleft surgery without them is inconceivable.
These maneuvers describe below are based on three embryologic principles
-
First, the concept of functional muscle repair, as per Delaire–Markus, is of great value but contains a fundamental embryologic misconception about the nasalis muscle. By assuming that it inserts into the midline at or near the anterior nasal spine, and by failing dissect it out as a separate structure, all standard lip repairs, be the supraperiosteal or subperiosteal, anchor nasalis into the midline, thereby condemning it to function as a constrictor, instead of as a dilator.
-
Virtually 100% of complete unilateral and bilateral cleft repairs having the nasalis centralized will have paradoxical alar movement during respiration and impaired nasal breathing.
-
-
Second, the surface area of vestibular lining is normal. It is simply mismatched with respect to the overlying skin envelope. The layers can be separated bluntly with an elevator and the internal lining rotated upward toward the midline. The distal anchorage point is release as well during the process of bone dissection into the alveolar cleft and piriform fossa.
-
Third, low-grade stenting in the postoperative period is very effective and, depending on the technology used, quite well tolerated.
Talmant #1
Like all muscles innervated by the facial nerve, the nasalis muscle originates from the paraxial mesoderm of somitomere 6. It flows forward within the substance of the SMAS along the trajectory of the buccal branch of VII. Upon reaching its destination it inserts at two distinct locations. Under normal conditions, the muscle has two proximal sites of attachment, both of which are into the mucoperiosteum of the canine fossa. A lateral “head” lies over the root of the canine while the medial “head” is located above the lateral incisor. These proximal attachments require the correction position of the embryologic fields making up the piriform rim. First to develop is the frontal process of the maxilla, arising from just above the canine. Second to develop is the frontal process of the premaxilla arising from just above the lateral incisor. The proximal heads of nasalis form their insertions in the same sequence. The SMAS then migrates upward, tracking along the vestibular lining, carrying the remaining nasalis myoblasts upward into the nose. At the lateral crus, the cartilage-vestibular attachment forces the SMAS over the dorsal surface of the alar cartilage. Here it forms a distal insertion into the perichondrium of the lateral crus.
In cleft patients the frontal process of the premaxilla is reduced or missing. The proximal muscle mass is pathologically displaced into the piriform fossa. It is attached to the mucoperiosteum immediately in front of the inferior turbinate, the exact location of the missing frontal process of the premaxilla. Distal nasalis anatomy remains unaffected. At operation, the muscle is readily encountered, using the same type of releasing incision as for DFR. The proximal muscle mass is substantial, one simply has to look for it. It is detached from two points: (1) from the mucoperiosteal lining, using sharp dissection; and (2) from the internal surface of the piriform margin using the subperiosteal plane. The distal muscle mass may be optionally released from the lateral crus using the subperichondrial plane. Reassignment of the displaced nasalis is accomplished by placing a mattress suture into the distal muscle mass and then suturing it to the alveolar mucoperiosteum just above the canine. This maneuver instantly corrects the pathologic vector of muscle contraction; it now acts functionally to open the airway, not to constrict it.
Talmant #2
Redistribution of the nasal lining is simple and elegant. It involves four maneuvers: (1) elevation of the septal mucoperichondrium all the way backward and upward to the nasal bone; (2) subperiosteal dissection from the internal piriform margin all the way backward and upward to the nasal bone; (3) subperiosteal dissection of the maxilla, external piriform margin and nasal bones; and (4) optional subperichondrial dissection via a rim dissection. Note that (3) and (4) involve the sub-SMAS plane.
Nasalis detachment requires entry into the piriform fossa. Using a sweeping motion, the mucoperiosteal sleeve of the lateral nasal wall is elevated off the bone completely, all the way up to the nasal bone. Here, the lateral dissection becomes continuous with the medial dissection of the septal mucoperichondrium. The result of these maneuvers is the complete liberation of the nasal lining; the lateral wall rotates internally. Embryologic anchorage of the lateral crus to the underlying vestibular skin (its “program”) ensures that it will be dragged into a correct position. At this juncture, the tip defining point is assessed. If additional medicalization is needed, the lateral crus can be freed up from its overlying nasalis attachment, using the subperichondrial plane. This requires a rim incision.
Talmant #3
Once separation of the nasal lining from the SMAS is achieved, how can one effectively reposition the lower lateral and upper lateral cartilages? A roll of thin silicone sheeting is placed into each nostril and secured with transfixion sutures. During this maneuver, the now mobilized lining can be physically manipulated and the position of the alar cartilage observed. The silicone stents are then packed with vaseline gauze. This expands the nasal cavity and effectively pushes the upper lateral cartilage into a new relationship with the nasal bone and with the septum.
Lateral Wall Dissection: Embryologic Implications
The shape of the piriform fossa dramatically affects the appearance of the nasal soft tissues. Recall that this structure is bilaminar. It is composed, externally, by the frontal process of the maxilla and, internally, by the frontal process of the premaxilla. The alar base is pinioned to the external rim. The vestibular lining contains the “program” of the lateral cartilages, determining their size, shape, and position. In cleft patients, a deficiency or absence of the premaxillary frontal process causes the rim to be posteriorly recessed and inferiorly scooped out. The canine fossa is ablated. This causes pathologic displacement of the distal nasalis. The medial head is transferred from above lateral incisor to within the piriform fossa. The lateral head shifts from above the canine to the skin of the nostril sill. A lateral vestibular “web” is the result. The internal nasal skin envelope contracts into the piriform fossa “sink.” This further flattens the lateral nasal.
Talmant’s first contribution is the recognition of the physiologic role of the nasalis to maintain a patent airway. By demonstrating over two decades the consistent displacement of the nasalis muscle inside the piriform fossa, his work explains observable anatomy long misunderstood, that is, the nasal “web.” Moreover, it confirms the role of the frontal process in constructing the normal nasal vault.
Talmant’s second contribution is supported by two important embryologic points. (1) Soft tissues produce membranous bone. Biosynthesis of the internal piriform fossa depends upon stem cells within an internal lining of adequate dimensions. When the lining is contracted (as in clefts), over time bone deposition and resorption of the piriform fossa cannot take place normally. Freeing up the internal lining reassigns stem cells into a correct functional position. Thus, growth over time creates a normal nasal cavity. The end result of Talmant’s mucoperiosteal dissection is a redistribution of the osteosynthetic envelope into a developmentally correct position. (2) The frontal process of the premaxilla is the missing Lego® piece of the piriform fossa. Its absence results from an absolute lack of premaxillary soft tissue immediately in front of the inferior turbinate. This deficit causes the “scooped out” appearance of the piriform fossa in cleft patients. The soft tissues of the NPP represent the “housing” of the missing lateral incisor and frontal process fields. The NPP flap (based on the medial nasopalatine artery) can be separated from the PP and transferred into the floor of the nose and the lateral nasal wall. It fits into the releasing incision required for nasalis dissection.
-
By replacing the missing frontal process field, NPP reassignment accomplishes two important goals: (1) it potentiates Talmant’s lining release and (2) it increases the surface area at the external valve.
Nasal breathing plays a vital role in shaping the nasal air passage. It determines, in no small measure, the physical dimensions of the nasopharynx and, by extension, of the maxilla itself. This is vital for speech because the functional position of the soft palate depends upon the bony platform to which it is attached.
Cleft patients have impaired nasal breathing for the following reasons
-
Deficit of the soft tissues responsible for synthesis of the frontal process of premaxilla reduces the surface area of the vestibular lining, dragging the lateral crus downward.
-
In the absence of the frontal process, the proximal attachments of nasalis are malpositioned. Muscle contraction restricts the airway rather than opening it.
-
Contraction of the vestibular lining means that ongoing osteosynthesis of the piriform fossa cannot be normal. Its physical dimensions become abnormal.
-
The lower and upper lateral crura provide support for the vestibular lining. In particular, correction of the upper lateral cartilage has been largely ignored in cleft surgery.
The sum of these four factors is increased turbulence at the external valve and reduced airflow. Talmant’s concepts provide an embryologically sound answer to these issues.
Microform Cleft: Premaxillary Deficiency and Poiseuille’s law 𝚫P = 8uLQ/𝛑r4
The underlying problem in the microform or “minimal” cleft is no different than in more explicit manifestations of cleft. The stigmata arise from a mesenchymal deficiency in the premaxilla. The frontal process zone deficit causes a warping of the piriform fossa. A deficit in the lateral incisor zone causes the nasal floor to be “scooped out.” To the degree that the bone volume is reduced, the strength of the BMP-4 signal will also be diminished. SHH, the protein within lip tissue that stabilizes the epithelium, will be less inhibited [34].
In sum, the spectrum of minimal clefting is absolutely quantitative, with the degree of soft tissue involvement proportional to the reduction in bone stock. Premaxillary deficits always affect the insertion of the nasalis muscle with consequent depression of the alar cartilage. Nasalis misinsertion, when significant, manifests itself as a “band” in the lateral nasal wall. That these factors should also cause a deviation of the anterior nasal septum is not intuitively obvious.
Talmant deserves credit for drawing our attention to the effect of hydraulic forces resulting from fetal ventilation of amniotic fluid upon the shape of the nasal fossae. As previously mentioned, sagittal real-time ultrasound studies demonstrate how the alar cartilages and septum respond to the influx and efflux of fluid through the nares. In the case of cleft lip associated with cleft palate, Talmant postulates a difference in pressure between the two sides, the non-cleft nostril having higher pressures than those within the labiomaxillary cleft. It is easy to conceive how the anterior septum could be warped.
The “fly in the ointment” of this argument is the minimal cleft. Here, the septal deformity is exactly the same yet the nasal floor remains intact. The answer, once again, lies in the reduction in bone volume within the frontal process and lateral incisor fields of the premaxilla. Poiseuille’s law predicts that minor increases in radius will significantly reduce the pressure drop (ΔP) in a linear tube of length L. Let us consider the nostrils as two tubes of equal length in parallel sharing a common wall (the septum). Fluid viscosity (u) and fluid flow velocity (Q) upon entry into the tubes stays constant. Although the piriform fossae are not circular, the premaxillary deficit increases the overall perimeter on the cleft side. This translates into a non-traditional “radius” that exceeds that of the non-cleft piriform fossa. Unequal pressures within the nostrils result in the difference being maximal at the level of the external nasal valve. Growth of the nasomaxillary complex and of the septum takes place in the fetal period. Unequal intranasal pressures during this time account for warping of the septum as seen in the microform cleft and in all other variants.
Finally, displacement of the lower and upper lateral cartilages in the microform cleft demonstrates the importance of epithelial–mesenchymal programming, a concept we have invoked previously. Nasalis in the microform is minimally displaced but the piriform fossa deformity remains. The finite deficiency of nasal skin from the defective premaxilla creates an insufficiency state within the lateral nasal wall. The vestibular lining, displaced downward, synthesizes a perfectly normal cartilage in the wrong place. A similar effect can be observed, on occasion, in microform clefts in which one notes an asymmetry of the nasal bones, with slight flattening on the side of the cleft. The effect can only be explained by an abnormally positioned epithelial program.
The surgical consequence of repairing the microform cleft using a Talmant intranasal subperiosteal dissection is that the biologically active vestibular “bone factory” will be repositioned into its normal state. As membranous bone synthesis continues over time one can expect progressive improvement in the shape and volume of the piriform fossa.
Dental Arch Management
The impact of cleft surgery on maxillary growth after surgery has been intensively studied. Blame in such cases is usually attributed to surgical intervention versus maxillary “hypoplasia.” Nonetheless, multiple studies of unoperated CL(P) patients demonstrate two patterns of fundamental scientific importance. (1) The cleft-side maxilla has an abnormal piriform fossa; this can exert distal effects on the overall shape of the anterior maxillary wall. (2) The cleft-side alveolar process has normal growth potential. We have already discussed the first phenomenon so we shall concentrate on the second. What is at the root of all this confusion?
Prior to the era of molecular embryology and stem cell biology, the mechanism of membranous bone growth was poorly appreciated. Textbooks would describe the maxilla as developing in membrane from a single ossification center. This model, based upon nineteenth-century descriptive embryology, is completely out of date.
We now know that osteoblasts arise from stem cells residing in the cambium layer of periosteum. The relationship between membranous bone and soft tissue is like butter and bread: the bread synthesizes the butter. More precisely, membranous bone develops when a unique population of neural crest cells organized as a neuroangiosome comes in contact with an epithelial surface (mucosa, skin, meninges) from which it receives a “program” determining the size and shape of the bone product.
Some membranous bones are unilaminar. The nasal bone is synthesized from forebrain neural crest (sixth prosomere). Its “program” is nasal mucoperiosteum supplied by the V1 anterior ethmoid neuroangiosome. Others membranous bones are bilaminar, with two sources of programming. Such bones are characterized by a separation plane occupied by sinuses or bone marrow. The prepalatine bone (Sic, the horizontal plate of the maxilla) is synthesized from hindbrain neural crest (second rhombomere). It has two “programs.” The upper (nasal) layer is the V2 lateral nasopalatine neuroangiosome. The lower (oral) layer is the V2 greater palatine neuroangiosome (GPA). Sinuses in the palatine bone fields of the maxilla (P1 and P2) are a well-described anatomic variant. Membranous bones of the calvarium, such as the parietal bone, arise from dual sources of mesenchyme (dermis and dura); these are programmed by skin and neuroepithelium. The interspace of the parietal bone contains marrow.
Let us now consider the membranous bones of the dental arch. The purpose of the alveolus is to house the dental apparatus. Two factors determine the size and shape of the arch: the number of dental units present and the effective biosynthesis of its bony walls. Both the upper and the lower dental arches are constructed in exactly the same way. They are “sandwiches.” In the center is a dental field arising from neural crest cartilage. It has its own intrinsic neurovascular supply. The lingual wall is composed of four distinct bone fields: premaxilla, P1–P3. The buccolingual wall is composed of an apparently single bone, the maxilla proper, divided into three developmental zones supplied by branches of the superior alveolar arterial system. Stem cells of neural crest origin within these various neuroangiosomes lay down membranous bone on either side of the dental anlage, much like armor plating.
The following table lists the neurovascular territories of the three layers (see Table 8.1). Here are the main take-home points: (1) The alveolar bones of both jaws are complex structures composed of multiple neuroangiosomes. (2) Alveolar osteogenesis is utterly dependent upon the integrity of these fields. (3) Malformations of the dental arch arise from abnormalities in the number or size of teeth, deficits in the alveolar housing, or both.
The nasomaxillary complex is a series of neural crest bones, each of which is innervated by a specific branch of V2. Using a VEGF mechanism each nerve induces an accompanying artery from the stapedial system (the internal maxillary). All these neuroangiosomes have a common point of origin: the pterygopalatine fossa. The maxilla per se has a partial role in the alveolus: it is responsible for the dental field and the external lamina. The prepalatine and palatine bones form the internal lamina. Although the premaxilla is an autonomous bone, under normal conditions, its external lamina receives mesenchyme of the maxilla.
The mandible is equally complex. In its primitive tetrapod form it consisted of multiple bones. The original dental lamina arose from Meckel’s cartilage and consisted of a dentary bone plus two to three coronoid bones. The body of the mandible supporting the teeth had two splenial bones. The primitive ramus had four bones: pre-articular and articular were lingual. The fates of these bones are listed. The mammalian mandible bears muscle insertions corresponding to the genetic “idea” of these ancient bone fields.
Response of the Dental Arch in Clefts to Surgery
Clefts are experiments of nature. They present us with clues as to the mechanisms behind craniofacial embryology. Nowhere is this more apparent than in the completely divided palate. The cleft-side maxilla is dynamic; it responds to surgical intervention in two primary ways: retroposition (growth restriction) and collapse (cross-bite). We shall deal with these problems in sequence.
Retroposition
At birth, the cleft-side maxilla appears smaller and/or retropositioned when compared with its normal counterpart. This situation likely results from disruption of force vectors in utero. Forward, centric growth of the ethmoid, vomerine, and premaxillary fields is disconnected from the cleft-side maxilla (in bilateral clefts, from both maxillae). Nonetheless, multiple studies of unoperated CL(P) patients demonstrate two patterns of fundamental scientific importance. (1) The cleft-side maxilla has an abnormal piriform fossa; this can exert distal effects on the overall shape of the anterior maxillary wall. (2) The cleft-side alveolar process has normal growth potential. We have already discussed the first phenomenon so we shall concentrate on the second. Why does the dental arch in cleft patients present as it does? Why does it self-correct over time (in the absence of surgery)? How does surgical intervention affect this process?
Recall that the primary determinant of alveolar size is the number of teeth. Assuming that the cleft-side alveolus has all six dental units, one can expect that it will eventually achieve normal proportions. Dental development involves the interaction between the epithelial program and the underlying mesenchyme. Although this process is delayed on the cleft-side, it eventually takes place. Assuming a normal mucoperiosteal envelope, the alveolar walls of the cleft maxilla will respond to accommodate the dentition. For this reason, patients with unoperated complete cleft palate can present with a dental arch of normal dimensions.
Restriction in maxillary growth after surgery has been intensively studied. Blame in such cases is usually attributed to surgical intervention versus hypoplasia intrinsic to the maxilla. Studies by Bardach (in the supraperiosteal plane) showed that increasing degrees of dissection were accompanied by increased growth reduction. At the same time, long-term work by Tessier and Delaire demonstrates that extensive subperiosteal dissection does not result in growth. What is at the root of all this confusion?
As previously discussed, biosynthesis of membranous bones is wholly dependent upon the vascular integrity of the neuroangiosomes from which they are constructed. These contain stem cells, located in the cambium layer of the periosteum. Any surgical intervention that invades or falsely subdivides a neuroangiosome will potentially compromise the blood supply to the stem cell population and negatively affect osteogenesis.
Traditional techniques of lip repair have mobilized the lateral lip element using the readily accessible supraperiosteal plane. As a consequence, cleft lip surgery disrupts blood supply to the stem cell rich cambium layer. Restriction in growth of the maxillary wall (Tessier zones 4, 5, and 6) can lead to malposition of the alveolus per se. Blood supply to the buccolabial mucoperiosteum can also be reduced. Bardach’s findings in beagles, using the supraperiosteal plane, were originally interpreted as a caution against wide dissection [35,36,37]. In point of fact, these papers demonstrate just the opposite: the use of the supraperiosteal plane violates the principles of embryology by devascularizing developmental fields. Because membranous bone is just a by-product of its soft tissue envelope, preservation of the resident stem cell population in the cambium layer of the periosteum requires the use of the subperiosteal plane. Applications of this concept to cleft surgery were described in detail with a detailed literature review by this author [31, 38, 39].
In cleft palate repair, oral mucoperiosteal flaps are commonly elevated using an incision placed at the junction between the lingual alveolar wall and the horizontal palatal shelf. This maneuver disrupts blood vessels directly laterally from the greater palatine axis to the lingual mucoperiosteum of the alveolus. As a consequence, blood supply to the lingual mucoperiosteum becomes random, based upon collateral flow from the opposing buccolabial mucoperiosteum and upon the underlying alveolar bone.
Just as the whole is greater than the sum of its parts, the most negative effects on alveolar development are observed in patients having combined surgical interventions for cleft lip and cleft palate surgery. The resulting process, sequential vascular isolation, is the primary cause of maxillary growth arrest after cleft surgery [32].
Collapse: A Reversible Phenomenon
Unoperated patients with complete clefts of the lip and palate demonstrate a gap corresponding to the absence of the two premaxillary fields (lateral incisor and frontal process). In the growing child, lip repair re-establishes force vectors across the alveolar cleft that are unopposed by bone: collapse ensues. This situation is dynamic; it is easily reversed by orthodontic expansion. The Achilles’ heel of this approach to cleft management is access to orthodontic care, a resource not readily available in many countries with a high-volume cleft population.
In theory, primary alveolar stabilization would prevent collapse and reduce or simplify subsequent orthodontic management. Inclusion of soft palate repair with the primary lip intervention ameliorates, but does not prevent, arch collapse. Long-term splinting and delayed repair of the hard palate lead to good size and position of the arch but poor speech. Over the years, various attempts have been made to address this problem using soft tissue procedures with or without some form of grafting. Early work suffered from soft tissue considerations stemming from use of the supraperiosteal plane. Observation of maxillary growth restriction led to a consensus that primary grafting should be abandoned in favor of secondary grafting at ages 6–9 (eruption of the canine).
This dogma was successfully challenged by Kernahan and Rosenstein at the University of Illinois [40, 41]. Using extremely limited dissection and rib grafts, stabilization was achieved. Long-term growth studies (up to 30 years) in these patients demonstrated arch development without maxillary restriction. Of note, the grafts were placed as struts high up over the alveolar cleft, leaving the actual walls of the cleft site largely untouched. Rosenstein’s final report is of great scientific importance for several reasons. First, it shows that primary arch stabilization is achievable. Second, and more importantly, the relationship between grafting and maxillary growth restriction reported in previous studies is an artifact of the surgical technique resulting from entry into the supraperiosteal plane.
More recently, GPP using local flaps at the alveolar cleft site has been reported with the hope that autologous bone would be generated at the cleft site (an idea dating back to Skoog) [42]. This approach is limited by the dimensions of the cleft site (wide clefts are not amenable). The dissection is technically demanding: flaps are taken directly over alveolar bone that is extremely delicate. The amount of bone generated by this technique has been disappointing. Elevation of the flaps may compromise the stem cell population within them. Furthermore, scar formation at the site may have consequences for dentition, such as tipping or rotation. Such problems are certainly amenable to orthodontic correction, but the limited benefits of GPP and its technical nuances do not recommend it.
Contributions of Jean Delaire
Delaire’s work demonstrates the surgical importance of the subperiosteal plane in cleft surgery [1, 43]. Elevation and medicalization of such flaps from the face of the maxilla represents the transfer of bioactive stem cell-bearing mucoperiosteum into a correct midline position. This concept is of fundamental importance for DFR cleft repair because it corrects the displacement of soft tissues that is present on both sides of the cleft. Such displacement is asymmetrical in the unilateral cleft and symmetrical in the bilateral cleft.
As previously mentioned, an unintended side effect of the Delaire’s dissection is that it draws the malpositioned nasalis into the midline along with the rest of the lateral lip musculature. This changes the function of the nasalis and constricts the airway. Talmant, having inherited many of Delaire’s patients, recognized this problem and worked out how the nasalis could be repositioned to its natural attachment at the canine fossa. This simple dissection frees both heads of the muscle and passes them below the orbicularis complex. Technical details are provided in the addendum.
Using the Delaire dissection plus alveolar cleft closure as described by Demas and Sotereanos, in 1996 this author described use of an extended alveolar flap that can span a distance of two tooth units [38]. This size of this flap permitted abundant tissue to be mobilized backwards from the alveolar margins for a secure soft tissue closure in larger clefts. The sliding sulcus technique was therefore more versatile than the GPP. It worked well for patients 2 years of age and up. In younger patients, the sliding sulcus had the same technical drawbacks of soft tissue manipulation at the alveolar cleft site. In 2004–2006 the problem of inadequate stem cell response seen in the above techniques was addressed by placing rhBMP2 in the alveolar cleft site. CT scans demonstrated bone formation that filled nearly the entire vertical dimensions of the cleft site. At the end of 10 years of work and observation, I came to the conclusion that gaining alveolar cleft closure at the primary surgery was doable but not worth the effort. A simpler approach was required. As we shall see, the search for this better solution led eventually to the Talmant protocol.
The bottom line is this: Only nature can make a dental arch ... and only the orthodontist can set it right. In the un-grafted complete cleft palate the dental arch is unstable; all forms of lip repair cause alveolar collapse. Fortunately, such perturbation is only temporary. DFR transfers stem cell populations into correct, centric positions such that they produce bone where desired. DFR is designed to assist the orthodontist in all other aspects of cleft palate development: airway, soft palate positioning, and achieving complete closure of the anterior hard palate.
Functional Lip and Palate Repair: The Five As
Let’s put the dental arch into the overall scheme of the cleft problem. Patients with complete clefts of the lip and palate require a prioritized, functional approach characterized by the “five As”: airway, articulation, anterior fistula avoidance, arch, and aesthetics. What is the rationale behind this order? There is no question that intelligible speech is the most important social benefit for these patients, so why prioritize the airway before articulation?
Airway
The anatomic and physiologic relationships between the hard and soft palate can be described by the pinball machine model. The soft palate is like the “flipper” in a pinball machine (a moveable lever that prevents the ball from escaping and keeps it in play). The hard palate is the platform to which the flipper is attached. The function of the soft palate flipper is to reach backward and upward to make contact with the pharynx; thereby closing off the nasal airway during phonation. The ability of the soft palate to accomplish this goal depends on: (1) normal motor innervation from cranial nerves V3 and IX; (2) normal muscle mesenchymal mass from somitomeres 4 (first arch) and 7 (third arch); and (3) normal length of the bony platform to which the soft palate musculature is attached. In other words, one must be able to control the flipper. It must be of normal length and not be floppy. Above all, the flipper mechanism must be positioned on the platform in such a way that it closes off the escape of the ball.
The size and shape of the hard palate bones (prepalatine and palatine) determine the positioning of the soft palate. When these bones are deficient, the soft palate musculature cannot make contact with the pharynx. In some patients, however, an apparently normal hard and soft palate complex is inadequate, causing velopharyngeal incompetence. What is the problem here? The secondary hard palate is not an isolated entity. It is part of a larger bone complex, the nasal chamber. The prepalatine and palatine bones are in anatomic contact with the frontal process of the maxilla, the inferior turbinate, and the sphenoid. Thus, the overall dimensions of the nasal airway determine the spatial relationship between the soft palate and the posterior pharynx and consequently its function.
Surgical implications of the above are as follows: First, the vestibular lining of the cleft nose is malpositioned due to deficiency/absence of the premaxillary frontal process. If left in this state, the ongoing process of osteogenesis during the first years of life will produce a permanent deformity in the bony dimensions of nasal chamber. Although the primary determinants of soft palate position are the prepalatine and palatine bones, these in turn are affected by the spatial dimensions of the surrounding bones, that is, the nasal airway. Correction of the airway by reassignment of the internal lining field (and therefore the cartilages) and of the nasalis is required to achieve optimal hard palate dimensions.
When should nasal airway correction be undertaken? Talmant believes that if nasal breathing patterns are not established in the first 6 months of life, oral breathing patterns will develop. Mouth breathing has negative effects that are difficult to reverse. These include head positioning, body posture, and maxillary shape. This logic is indisputable but it is unclear to me how rigid one should be on this issue. Children can present late, with wide clefts or with complications. The good news is that airway correction can be easily done secondarily, either concomitant with lip revision or as an isolated procedure. It seems thus reasonable to conclude that nasal breathing should be accomplished in all children by age 2.
A caveat to the reader here: technical details regarding intraoperative nasal packing and postoperative nasal stenting are of the utmost importance for airway correction. These accomplish the following: (1) a subtle, but very precise, control of alar cartilage position; (2) upper lateral cartilage repositioning; and (3) adequate diameter of the external nasal valve. These maneuvers will be described in the technical section that follows ... stay tuned.
Articulation
Timing of palate closure is yet another area of cleft management that is rife with controversy. As we have previously seen, palate clefts can be classified by embryologic mechanism, depending on which components are affected. Cleft palate arises from faulty mesenchyme involving bones (neural crest from rhombomeres r1, r2, r3), muscles (paraxial mesoderm from somitomeres 4 and/or 7), or a combination of both. In the absence of such classification, studies relating surgical technique to speech are inherently unreliable; apples cannot be compared with oranges.
Nonetheless, some general concepts of cleft palate repair make sense. First, soft palate closure mechanically positions the tongue, helping prevent the acquisition of motor patterns that are difficult to correct later. This can be done as early as 6 months, as in the Talmant protocol (see below). Soft palate repair establishes normal force vectors across the posterior midline. These balance against force vectors created by lip repair or lip adhesion. Any form of early lip closure exerts force vectors across the alveolar cleft leading to arch collapse and cross-bite. Thus, lip repair and soft palate repair are a rational combination at 6 months.
A quick review of the embryology of the soft palate musculature is required. Five sets of muscles are involved. First pharyngeal arch muscles arise from paraxial mesoderm of the fourth somitomere. The sole Sm4 palatal muscle is tensor veli palatini.
Third pharyngeal arch muscles arise from the seventh somitomere. These are levator veli palatini, palatoglossus, palatopharyngeus, uvulus, and superior constrictor.
These muscles can readily be tested using a Peña® muscle stimulator (Integra Life Sciences, Plainsboro, New Jersey). Normal function is graded as follows: normal = 2, hypotonic = 1, and no response = 0. Differences frequently exist between the left and right sides. Such information is valuable for classification of the palate cleft and for diagnosis of speech problems, if these should arise.
Soft palate muscles exist in two layers. Nasal mucosa provides the “program” for the tensor and levator. Oral mucosa is the “program” for the palatopharyngeus and palatoglossus. Uvulus is programmed by both mucosal surfaces. The upper pharyngeal mucosa contains the “program” for the superior constrictor.
Only the muscles of the nasal layer (tensor and levator) are responsible for palatal elevation/elongation. Only these muscles are directly attached to the palatine bone. These are the one that need to be set back. The dissection plane between the nasal and oral layers is readily defined. Extensive three-layer intervelar veloplasty is not embryologically rational and causes unnecessary scarring. Soft palate repair is best viewed as a “first-pass” event. If the functional result is good, nothing more is required. If VPI is present, and the elevation pattern is V-shaped, secondary lengthening with a double-opposing z-plasty will readily resolve the issue.
Anterior Fistula Avoidance
The GPA supplies two bone fields: the horizontal shelf of the palatine bone and the oral lamina of the prepalatine bone. It runs all the way forward to the junction of the canine and lateral incisor. There it makes an anastomosis with the MNP. In traditional cleft palate surgery, hard palate incisions located at the junction of the palatal shelf and lingual alveolar wall constitute an embryologically incorrect subdivision of the greater palatine fields. Not only does this incision interrupt blood supply to the alveolar mucoperiosteum; it also creates short flaps. Troublesome anterior fistulae result from this design.
In the unoperated state, the length of the GPA field is designed to reach the premaxilla. Achieving a water-tight closure is simply a matter of using the entire neuroangiosome, using a pericoronal incision (or one just lingual to the teeth). The term AEP refers to the additional tissue gained by this flap. The anatomy and developmental biology of this procedure have been previously described. The AEP procedure is also useful in secondary cases. When enough time has elapsed after the primary palate repair, collateral flow is re-established between the palatal shelf and alveolar mucoperiosteum. One can therefore use the AEP incision design to re-elevate the entire mucoperiosteum of the hard palate and lingual alveolus as a single flap. In unilateral clefts, the AEP flap from the non-cleft side always projects beyond the alveolar cleft, while that from the cleft side reaches the back wall of the alveolar cleft. These flaps permit closure of anterior fistulae in virtually all situations.
Arch
Surgeons and orthodontists get along best when goals of each specialty are respected
-
Early lip closure (at 6 months) will cause the alveolar cleft to narrow down and may create a cross-bite of the primary dentition, but this is readily correctible.
-
Orthodontist to surgeon: Alveolar soft tissues over un-erupted teeth are delicate ... leave them alone.
-
-
Achieving a perfect closure of the entire nasal floor is a major goal of the primary surgical sequence. The most technically difficult site for this is the anterior floor, just over the alveolar cleft, exactly where the collapse has taken place. When nasal palatine mucoperiosteum is joined with that of the vomer, the floor will always be short in the anterior–posterior dimension. The NPP flap corrects this deficit: it is dissected out of the lip, transferred into the nasal floor, and sutured to the vomer-maxilla closure.
-
Surgeon to orthodontist: The anterior nasal floor is tight and tricky ... can you expand the alveolar cleft prior to hard palate repair and then keep it open until I graft it? What is required here is a means of palatal expansion that can be maintained over time. This also favors proper positioning of the lateral nasal wall. Such a device cannot be fixed; it must be capable of adapting to changes in the arch. As detailed below, the modified quad helix as designed by Lumineau fulfills these requirements and produces, often by age 4, a dental arch ready for bone grafting.
-
Developmental Field Reassignment
At the beginning of the chapter, we set out to define a set of biologic goals for cleft repair. At this point, the protocol that comes closest to achieving them is that published by the Nantes group. Their results have been gained over 20 plus years. Credit should be given to Jean Delaire, from whose work Talmant’s concepts have evolved. In any case, the outcomes of the airway and dental arch using this protocol not only are the best I have seen but also have a sound basis in developmental biology (Figs. 19.68, 19.69, 19.70, 19.71, 19.72, 19.73, 19.74, 19.75, 19.76, 19.77, 19.78, 19.79, 19.80, 19.81, 19.82, 19.83, 19.84, 19.85, 19.86, 19.87 and 19.88 reference DFR cases done with 5-year follow-up.)

Case 1 5-year result: left UCL, incomplete with complete cleft of primary and secondary palate. Patient has microphthalmia and ptosis. Nasal projection maintained. No nasalis was done so nasal tip is asymmetric and nostril axis is oblique versus normal vertical. Top/bottom left pre-op; top/bottom right 5 years post-op. [Courtesy of Michael Carstens, MD]

Case 1 Alveolar extension palatoplasty with rhBMP2. Top left: surgical defect. Top right: flaps elevated. Note their length; they exceed the boundaries of the alveolar cleft. Bottom left/right: At 5 years arch is preserved and lateral incisor has erupted. [Courtesy of Michael Carstens, MD]

Case 1 3-D CT scan AP and palatal views pre-op versus 5 years showing complete filling of the primary cleft with good height of bone maintained at 5 years. [Courtesy of Michael Carstens, MD]

Case 1 Dental models taken at age 5. Occlusion is Class 1. Note the left lateral incisor is in small degree of cross-bite. [Courtesy of Michael Carstens, MD]

Case 2 Primary BCL/P, complete on right and incomplete on the left. Palate cleft was complete, unilateral on the right. AP and submental vertex views show that airway symmetry at 5 years is maintained. [Courtesy of Michael Carstens, MD]

Case 2 Subperiosteal release reassigns soft tissues without tension despite projection of premaxilla. Upper panel show projection premaxilla. Lower panel at 5 years shows symmetrical facial growth. Note equality of nasal creases. Good nasal tip projection. [Courtesy of Michael Carstens, MD]

Case 2 Comparison of 3D CT scans pre-op versus 5 years showing that graft with rhBMP2 has stabilized the arch but anteriorly the cleft is only about 50% filled. Will need more graft for implant placement if desired. [Courtesy of Michael Carstens, MD]

Case 2 Skull models made from the 3-D CT scan: pre-op and 5 years. Courtesy of Medical Modeling, Inc. Vertex submental views (upper left) and submental vertex views (lower right) show symmetrical midface growth at 5 years. Lower left: Apical dental film shows collar of bone around the erupted canine in the cleft. [Courtesy of Michael Carstens, MD]

Case 3 BCL incomplete with bilateral CP, incomplete on right, and complete on the left. Upper panel demonstrates a small philtrum. Lower panel shows 5-year aesthetics with good nasal tip projection. Despite the initial size of the philtrum DFR achieved good repositioning of the columella without need for NAM. [Courtesy of Michael Carstens, MD]

Case 3 Nasal tip anatomy and projection. Upper panel compared with lower panel shows good aesthetics at 5 years. DFR achieves good repositioning of the columella. [Courtesy of Michael Carstens, MD]

Case 3 At 5 years, unification of arch with rhBMP2 at palatoplasty has resulted in stability, class 1 and a nicely rounded, projection maxilla. [Courtesy of Michael Carstens, MD]

Case 4 Primary BCL with long-term growth of midface, stability of nasal projection, and protection of the airway. Note at 5 years (upper right and left) the nostrils are widely patent. [Courtesy of Michael Carstens, MD]

Case 4 5-year growth of the midface with stem cell reassignment has brought the premaxillae and maxillae into a normal relationship. Lower panel demonstrates ability of DFR to bring soft tissue envelope forward without compromising nasal projection. [Courtesy of Michael Carstens, MD]

Case 5 Personal comparison of rotation-advancement and DFR: same patient, same surgeon, different concepts, better biology. Complete UCL/P (wide) with primary repair using rotation-advancement presents with typical secondary deformities. Shown here is the on-table pre-op appearance at primary repair for which I was the attending surgeon. On-table results were nice but she represented at age 9 with the complete panoply of secondary deformities. [Courtesy of Michael Carstens, MD]

Case 5 Shown here (left) at 9 years post-rotation-advancement the patient has a lip notch, muscle asymmetry, nasal deformity and mouth breathing. Right panels show after secondary repair with DFR (Talmant) restoration of symmetry and a functional airway. [Courtesy of Michael Carstens, MD]

Case 6 Lateral views showing midface and nasal profiles. Note on the left panels aesthetic improvement of projection in what is apparently the “normal” right side. [Courtesy of Michael Carstens, MD]

Case 7 Secondary BCL with nasal airway restoration. Upper panel shows typical compression of the nose and compromise of the nasal airways. Lower panel demonstrates functional and aesthetic improvements after DFR. [Courtesy of Michael Carstens, MD]

Case 7 Bilateral secondary repair: lateral views show nasal projection and forward drape of the upper lip. Lower panel demonstrates increased tip projection and angulation of the alar plane. [Courtesy of Michael Carstens, MD]

Case 0 From the beginning, we return to this story of relapse, now treated with DFR and rhBMP graft. Original pathology was complete UCL/P on the right with incomplete/microform on the left. At 1-year post-op from DFR cleft lip–nose revision (lower right) a striking correction of the “recessed” maxillary soft tissue envelope has been achieved. [Courtesy of Michael Carstens, MD]

Case 0 Lower panel shows fullness of the midface and normalization of upper–lower lip aesthetics and significant changes in alar angulation, nasal projection, and normalization of the nose–lip relationship. [Courtesy of Michael Carstens, MD]

Case 0 CT demonstrates ability of rhBMP2 to achieve a complete fill of this large alveolar defect, extending back to incisive foramen. Orthodontics closed down the arc giving him good aesthetics and bite. [Courtesy of Michael Carstens, MD]
Talmant–Lumineau Protocol
-
Six months: soft palate repair plus full lip/nose repair. Wide bilateral subperiosteal dissection is carried out, both extrapiriform and intrapiriform. In some countries, OR and anesthesia conditions may require that the soft palate repair be carried out separately.
-
Step 2. As soon a primary dentition permits, a modified quad helix is placed. The cleft site is expanded over a period of approximately 1 month and then maintained. Unlike traditional quad-helix designs, it is not fixed to the molars. Instead, it is secured with three wire ligatures. The expander has four “eyelets.”
-
These are 360° loops placed posteriorly at the level of the second molars (E) and anteriorly at the level of the canines. The expander is attached to each molar, with the “pull” of each wire loop placed in opposite directions.
-
A third wire is passed through the posterior “eyelet” and over the second wire. The purpose of this wire is to prevent backward slippage of the expander. The third wire can be tightened to keep the device in place. In this way, the quad helix remains in situ but can be advanced forward to accommodate dental arch growth. Expansion is maintained until alveolar bone grafting is completed.
-
-
Two years: hard palate repair.
-
Four years: alveolar bone grafting. The timing for the procedure follows the extent of distraction: the maxillary intercanine distance must exceed the mandibular intercanine distance by 6 mm.
-
4.5 years later: remove quad helix.
Comments
The Talmant lip repair accomplishes the following goals. (1) It establishes an adequate nasal airway in both the short and the long term. (2) Subperiosteal elevation of the internal nasal lining elevates nasal cartilages into correct position, providing immediate support for the external nasal valve. (3) Reassignment of nasalis into an anatomic insertion changes the force vectors of breathing, both immediately and in the long term. (4) Lining release reassigns biosynthetic mucoperiosteum into a correct relationship with the developing bony cavity.
Talmant’s lip/nose repair does not touch the nasal mucosa of the hard palate; furthermore, it does not make use of a vomer flap. The reader should be aware that some schools of thought (particularly in Europe) maintain that vomer flap closure to the nasal mucoperiosteum is to be avoided. Certainly, repair of the anterior nasal floor is not required to accomplish the four goals listed above. However, the decision to not unite the nasal floor completely, once and for all, may be technically disadvantageous.
Concomitant with the lip repair, soft palate repair helps with early tongue posturing, narrows the posterior palate cleft, and offsets the anterior forces created by the cleft lip repair. Talmant wants the hard palate cleft to narrow down in anticipation of palatoplasty at 2 years. He also knows that Lumineau’s quad helix will compensate for collapse.
Hard palate repair at 18 months has no particular drawbacks. As we shall see (below) the eruption of dentition and the quality of alveolar bone make AEP flaps (when needed) simple to design and execute.
At age 4, the mucoperiosteum lining the (now fully expanded) cleft is easy to elevate. The alveolar bone is strong as well, protecting unerupted permanent dentition. Intervention at this age is rational because it does not pose a risk to the stem cell layer nor to the tooth buds.
DFR-Modified Talmant–Lumineau Protocol
-
Six months: DFR including complete nasal repair + soft palate with buccinator interposition palatoplasty.
-
Soft palate repair can be performed separately as soon as conditions permit.
-
-
At eruption of dentition: Lumineau adjustable quad helix.
-
18–24 months: hard palate repair.
-
If lip adhesion was done, revise with DFR. Steps 4 and 5 as per Talmant–Lumineau.
-
-
Four years: alveolar bone grafting. The timing for the procedure follows the extent of distraction: the maxillary intercanine distance must exceed the mandibular intercanine distance by 6 mm.
-
4.5 years later: remove quad helix.
Comments
Doing a full lip repair in isolation from the hard palate presents theoretical problems directly over the alveolar cleft site. (1) Prolabial tissue is brought into the nasal floor and remains isolated from hard palate mucoperiosteum. The NPP flap, to be most effective, brings with it periosteum from the lateral “shoulder” of the premaxilla. NPP is sutured under direct vision of the nasal palate closure, periosteum to periosteum. This type of continuity may be difficult to achieve at a second surgery. (2) These authors do not subscribe to a “no touch” approach to the vomer. From a developmental standpoint, the nasal side of the floor of the nose is supplied by two neuroangiosomes: medial nasopalatine from the vomer and the lateral nasopalatine coming from the lateral nasal wall below the inferior turbinate. The oral side of the nasal floor is supplied by the greater palatine. Thus, in normal embryologic conditions, each vomer is united to the maxillary crest of the ipsilateral P1–P2 shelf.
Achieving definitive nasal repair at 2 years versus 6 months represents an obvious trade-off. Do the requirements of cerebral “programming” demand nasal surgery at 6 months? Management of nasal stents is undoubtedly simpler in younger patients. Lip–nose surgery performed as a secondary procedure offers technical simplicity (the structures are larger and less delicate, have secure tissue planes, and a readily accessible continuity between the mucoperiosteum of the premaxilla, vomer, and septum). The length of the AEP flaps is such that the entire floor of the alveolar cleft will be covered. No oronasal fistula is produced. The technical design of doing a simultaneous lip and hard palate closure closes five of the six sides of the alveolar “box”: roof, floor, sides, and back wall. Thus, soft tissue closure of the alveolar cleft is possible at this stage. The relative merits of this are unknown.
Embryologic Basis of Occlusion and the Lateral Facial Cleft
The maxilla and mandible develop within the confines of the first pharyngeal arch. In its initial state, PA1 hangs downward from the axis of the embryo like a saddlebag. Pharyngeal arches are almost exclusively composed of neural crest. Running through the core of each arch is a mesodermal structure, known as an aortic arch. Let us explore how the first aortic arch develops.
Recall that the primitive embryonic vascular system consists of paired dorsal aortae running the entire length of the embryo. In the most anterior region of the stage-8 embryo (anterior to the brain) the dorsal aortae form a U-shaped loop. Here, the dorsal aortae take a different name, primitive heart tubes; these fuse to form the embryonic heart. The most distal (the most anterior) segment of the primitive heart is the atria, being connected to the vitelline veins. Stage 9 is characterized by three major events: embryonic folding, neural crest migration, and the appearance of the first pharyngeal arch. Brain growth is the driving force behind folding. The heart turns 180°, brining it ventral to the embryonic face. The functional components of the heart also undergo a 180° reversal. As a result, the atria now are positioned posterior to the future ventricular outflow tract. The two vessels connecting the cardiac outflow tract with the dorsal aortae become surrounded by the neural crest mesenchyme migrating downward from rhombomeres 1–3 of the hindbrain. The connecting arteries acquire a new name: the first aortic arch arteries.
From an evolutionary and embryologic standpoint these bones are not single entities; maxillary bone fields arise from neural crest of the second rhombomere, whereas those of the mandible arise from neural crest of the third rhombomere. In its initial state, the first arch hangs down like a saddlebag. Its rostral half is all r2, while the caudal half is made from r3. Thus, maxillary and mandibular bone fields sit directly across from one another along the “neuromeric fault line.” As we shall see, this situation explains their eventual occlusal relationship.
Oral mucosa is an ectodermal tissue all the way back to the buccopharyngeal membrane (represented in the fetal state by Waldeyer’s ring). Beyond that point, pharyngeal mucosa is endodermal in origin. Oral mucosa contains the “program” for each tooth. Neural crest mesenchyme simply responds, producing the appropriate dental units. For this reason, ectodermal dysplasias can result in malformed or absent teeth. The premaxilla and the dentary bone (being formed from neural crest) merely “present” potential tooth germs to the epithelium.
In any case, all five pharyngeal arches are divided into quadrants by a set of distal-less genes (Dlx) genes: Dlx-1, Dlx-2, and so on. Recall that the first and second pharyngeal arches fuse together very early, probably by the time the third pharyngeal arch makes its appearance (Carnegie stage 11). It can be postulated that a nested set of genes related to, or controlled by, the Dlx system exists along the midline of each arch. This would ensure that a potential zone of apoptosis exists in the first arch running along the longitudinal midline from distal to proximal. Epithelium invading the apoptotic zone contains “mirror image” genes specifying opposing maxillary and mandibular dentition.
Under normal conditions, the apoptotic event is confined to the deep-lying bone fields of the first arch, not the skin. However, if the signals responsible for this division are also located in the second arch and/or in the overlying skin, a lateral facial cleft will be present. Such a cleft extends progressively proximal from the oral commissure to the external auditory canal, as successive Dlx fields are involved. Thus, the cleft described as number 7 has nothing whatsoever to do with Tessier cleft zone 7, that is, the jugal bone and overlying malar soft tissues.
Technical Details of Developmental Field Reassignment Surgery
Children with CL(P) affecting the primary hard palate only (intact arch) are brought to the OR at 6 months of age for definitive repair [44].
When CL(P) affects both the primary palate and secondary hard palate, that is, when the cleft is complete (through the secondary palate) and narrow, DFR/Talmant lip–nose repair is done at 6 months.
In some cases, cleft lip accompanied by a wide complete palate cleft may benefit from a passive splint or from a Dynacleft® device prior to surgery. Lip adhesion should not be considered a “failure.” This procedure has a definite role; the resulting scar is not an issue because it will be relocated within the nose along with the non-philtral flap.
Embryologic Definition of the Philtral Prolabium
As previously discussed, the lip incision in DFR cleft surgery divides the prolabium into two distinct embryologic components: a “true” PP and a “false” NPP.
In normal patients, the central lip element (Cupid’s bow) and the PP are synonymous. The true PP is the terminal extension of columella and nasofrontal skin, all of which (epidermis, dermis, and subdermal tissue) originate from p5 forebrain neural crest. The philtrum consists of two paired fields; both are supplied by a V1 sensory nerve and a terminal branch of the anterior ethmoid artery. The two AEA vessels run about 2–4 mm apart. The developmental field of the prolabium is therefore in continuity with the entire columella, including the lateral walls all the way to the pink p6 septal mucoperiosteum. Contained within the philtral–columellar fields are the medial crura and footplates of the lower lateral cartilages. In the Matthews model, the philtral columns ascend from the white roll to the footplates. In this interaction, the width of the PP is no longer considered just that of the columella; it is defined by the transverse distance between the footplates.
In cleft lip patients the prolabium contains additional tissue, the NPP. This tissue is in continuity with the mucoperiosteum of the underlying premaxilla and vomer. The mesenchyme of the non-philtrum originates from r2 hindbrain neural crest. NP, premaxilla, and vomer share a common neurovascular axis: the medial sphenopalatine (nasopalatine) artery. Note that the mucoperiosteum of vomer and septum appears to be continuous, but these are actually embryologically separate fields. The soft tissue walls within which the septum is synthesized are derived from p6 forebrain neural crest. They are therefore innervated by branches of V1 and supplied by anterior and posterior nasal branches from the anterior ethmoid axis.
In normal patients, NP is never seen. It lies within the nasal floor as the skin cover for two premaxillary sub-fields: the lateral incisor field and the frontal process field. In the cleft situation, however, the neural crest bone elements of lateral alveolus and frontal process are gone. NPP becomes “shipwrecked,” a lonely mass of mesenchyme is cast ashore alongside the pre-existing PP. In bilateral clefts the prolabium will therefore have four separate fields: two PP fields in the center and one NPP field on either side.
In sum, the normal prolabium consists of a tissue complex with four layers, the skin and the underlying non-muscle-bearing mesenchyme. Both arise from forebrain neural crest (prosomere 5). These are supplied by the terminal branches of the anterior ethmoid artery. All remaining layers arise from the hindbrain. An intervening layer of fat conveys branches of the facial artery to the fascia and muscle of the DOO oris. The fascia is neural crest from rhombomere 5, while the muscle arises from somitomere 6. The facial nerve supplying the DOO has its nucleus in r5 as well. The mucosa is a neural crest structure arising from rhombomere 2. It is supplied by branches from the medial infraorbital artery.
Medial Dissection: the Non-Philtrum Flap, Septum, the Medial Nasal Fossa, and Nasal Tip
The DFR incision is designed to separate out tissues that are embryologically distinct. Recall that the purposes of the non-philtral flap are (1) to add length to the nasal floor, that is replace the missing “housing” of the lateral incisor; (2) to replace tissue deficit resulting from release of the lateral nasal wall from the piriform fossa, that is, reconstruct the missing frontal process of the premaxilla; (3) to release the alar footplate and permit advancement of the lower lateral cartilage into the nasal tip; and (4) to create direct access to the septum from below.
In harvesting the NP flap, knowing the width of the PP is critical. Previously, this author had described the transverse dimensions of the true philtrum as being equal to those of columella proper. Matthews’ definition above is broader and better. The width of the true philtrum is equal to the transverse distance between the alar footplates. This has two implications: (1) the resulting Cupid’s bow is embryologically more accurate; and (2) surgical dissection using these landmarks has technical advantages. Both these points will be discussed later (Figs. 19.30, 19.31 and 19.32).
The prolabial incision we are describing has implications for lip height and lip aesthetics. Using the former (narrower) definition of philtral width (in which 2–3 equals columellar width) only lip height is almost uniformly equal. That is, the height of the philtral column on the non-cleft side (13–2) equals the distance on the cleft side (12–3) equals the height of the lateral lip element (8–9). The aesthetic problem with this incision resides at the white roll at the cleft-side philtral column. The normal upper lip has natural pucker (a slight eversion) all along the white roll. This aesthetically important feature is caused by the presence of marginal fibers of the DOO. Recall that anatomic territory of DOO is biologically “programmed” oral mucosa. Thus, the distal margin of DOO follows the vermillion, curving upward in the form of a fishhook. It terminates at the mucosa–skin interface, that is, at the white roll. In a non-cleft situation, these terminal fibers extend right across the philtral column. By lending bulk to the vermillion just below the while-roll, the terminal fibers are responsible for eversion.
Matthews has pointed out that that when a DFR incision is brought straight down based upon columellar width alone, eversion at the white roll is obliterated. How can this be avoided and, at the same time, preserve lip height? Three simple steps will do the trick. (1) The definition of Cupid’s bow width is expanded, marking it out using the transverse dimensions of the footplates and normal mucosal landmarks. (2) The DOO fibers should be conserved all across the philtrum. (3) A measured back-cut above the “ridge” of the DOO will preserve lip height. Over time this incision becomes a straight line and conserves the aesthetically important eversion of the lip all the way across the repair.
In the Matthews modification of DFR, the PP is mapped out as follows. Point 1 (the center of PP) is located directly above the frenulum. Point 2 is the non-cleft philtral column. Point 3 (the cleft-side philtral column) is marked out with distance 1–2 equal to 1–3. Remember that PP and columella share a common source of mesenchyme (forebrain neural crest) and a common neurovascular supply (V1 and the terminal branches of anterior ethmoid arteries). The width of these fields is the transverse distance between the footplates. In a unilateral cleft the non-cleft footplate is point 13 and the cleft-side footplate is point 12. Matthews’ prolabial markings create a philtrum of the same width. Thus, 2–3 = 13–12. These medial markings will match those of the lateral lip element 8–9.
The eversion (pucker) of the lip is marked all the way across the philtrum. Because the DOO follows the mucosa, this will be observed to curve slightly upward. With our new definition, cleft-side lip height will fall short by 3–4 mm. One compensates for this by lengthening PP using a back-cut made above the natural roll of the muscle along the margin of the Cupid’s bow. The back-cut will dart inward about 2 mm and then back outward by the same amount at a 45° angle, where it joints the original curve of the DFR incision. This gives two benefits: (1) parity of lip height and (2) a natural fullness of muscle at the base of the new philtral column.
The proximal component of the prolabium incision ascends toward the columella. It circles around the base of the columella, passing beneath the footplate of the medial crus. The incision terminates at the transition between the columellar skin and the septal mucosa. Initially, the depth of this incision is skin only (Figs. 19.39 and 19.40).
The depth of the prolabial incision is important. Initially, it is entirely cutaneous. This permits elevation of the philtral skin from the underlying non-muscle-bearing mesenchyme for about 5 mm. Once the initial incision and undermining are accomplished, the prolabial incision is deepened from the white roll upward to the base of the columella (the underlying mucosa is spared). At the columellar base the incision remains cutaneous only. This is to preserve the underlying mesenchyme within which is contained the pedicle from the medial nasopalatine artery. This tissue is left intact. The cutaneous incision beneath the alar footplate permits entry into the columella with curved scissors. The tips of the scissors follow the medial aspect of the medial crus all the way into the nasal tip (Fig. 19.39).
A paring incision is now done from the white roll up into the nose. Again, at the base of the columella, care is taken to make this incision cutaneous only. The non-philtrum flap is now elevated with a generous subcutaneous base. The flap is lifted off the premaxilla in the subperiosteal plane up to, but not beyond, the junction of the premaxilla with the vomer. This ensures mobility of the flap without compromise of the pedicle. In sum, the non-philtrum flap is a skin island supported by subcutaneous tissue supplied by the nasopalatine artery.
The disposition of the medial mucosa is at the discretion of the surgeon. The M flap can be dropped downward like a baby’s diaper. It can be included with the non-philtrum flap for greater epithelial width. Finally, it can be inferiorly based and rotated. I prefer option 1 because it leaves option 3 open. The width of the non-philtrum flap is not strictly determined by the skin paddle. Subcutaneous tissue will readily epithelialize.
Septal mobilization is carried out next. This can be done by extending the incision like a hockey stick upward at the anterior septal border. A dental amalgam packer is very useful because its curved tip is flat and cross-hatched. It can rasp through the mucoperichondrium to the correct plane with great delicacy. The septum is completely dislocated.
Herein we encounter part 1 of Talmant’s nasal fossa reconstruction: the dissection is extended upward to the nasal bone and the vestibular lining is freed in the subperiosteal plane from beneath the nasal bone. This frees up the upper lateral cartilage from beneath the nasal bone. Recall that the abnormal anatomy of the nasalis predisposes to a flattening of the nasal bone and entrapment of the upper lateral cartilage. This maneuver also lengthens the dimensions of the nasal fossa [45].
We have now completed the medial dissection. It is now time to ensure that the goals of our nasal dissection are accomplished. The nasalis complex needs to be freed in the subperiosteal plane from the underlying upper and lower lateral cartilages. A small infracartilaginous incision provides access to the nasal dorsum. It is a good idea to visualize the alar cartilage (once again, the amalgam packer proves helpful). A McComb dissection is carried out. One should drop downward to encounter the bone along the piriform fossa. Spatial limitations of the nasal incision limit what one can accomplish. We must await the lateral dissection to ensure that our subperiosteal nasal dissection is complete.
Lateral Dissection: Muscle Separation, Nasalis Transposition, and the Lateral Nasal Fossa
Paring incision proceeds from point 8 all the way into the nasal cavity. Recall that distances 8–9 = 13–2 = 12–3. Also recall that the width of the keratinized mucosa at point 8 should equal that at point 3. With the lateral lip element stretched, one immediately undermines the skin from the SOO oris for a distance of about half a fingertip. This separation is continued upward to the nasal skin, involving both the lower alar base and the nostril sill. The superficial and DOO muscles are separated by a fat pad analogous to that found in the prolabium. SOO and DOO are separated proximally all the way to the termination of the deep layer (Figs. 19.20, 19.47, 19.48 and 19.49).
Nasalis and SOO are still confluent. Our next task is to separate them and gain control of the nasalis. This is most easily done using the concept of two planes: subcutaneous and subperiosteal. Recall that the medial head of the nasalis is inserted into the vestibular lining, while the lateral head of nasalis resides in the piriform fossa, extending halfway up its vertical extent. We shall start with the medial head. Extension of the skin under-mining into the nasal cavity now continues in a plane superficial to the medial head of the nasalis. The dissection should extend in front of the inferior turbinate and all the way up to the lower lateral cartilage. One now proceeds in the subperiosteal plane beginning at the entrance into the piriform fossa, that is, in the triangular zone corresponding to the nostril sill. One is now beneath the medial head of nasalis. The muscle is grasped and sectioned away from the bone. With the lateral lip once again on tension, the SOO is divided away from medial nasalis at the level of the alar base. Tension placed on the nasalis at this point will transmit to the lateral nasal wall but will not be free.
The lateral rim of the piriform fossa is swept clean upward to the halfway point. One has now gained access to the lateral margin of the fossa. An elevator is passed upward along the lateral wall of the nose until it achieves the nasal dorsum. At this point one should switch to an elevator that is sharp and strong, with a broad and curved tip, such as a McKenty. A customized Molt elevator works well. The elevator is swept downward underneath the SMAS (including the nasalis) all the way to the nasal tip. The result of this McComb dissection should be a complete liberation of the dorsal nasal skin and SMAS from the underlying upper and lower lateral cartilages (Fig. 19.52).
Recall that all unilateral cleft noses have an overall deviation of the soft tissue envelope on the non-cleft side away from the midline. This is an opportune moment to correct this problem and to centralize the entire midface envelope. Wide subperiosteal dissection is carried out over the face of the maxilla on both sides. Care should be taken to free up the non-cleft alar base as well. Because the mucoperiosteal envelope contains the stem cells required for future membranous bone synthesis, one has now “centralized the biosynthetic envelope” such that the external dimensions of piriform fossa (and the maxillae in general) will continue to auto-correct over time.
We now turn our attention to inside the piriform fossa to complete the dissection of the lateral head of nasalis. Using a curved elevator the nasalis muscle fibers lying within piriform are literally “scooped out” of the fossa. At this point the two heads are evaluated. They may appear distinct or as a single mass. The functionality of the muscle dissection is now tested. Traction is placed on the nasalis; motion along the lower lateral cartilages will be seen. The range is between 5 and 10 mm. If restriction is encountered, one can release the muscle further by first re-entering the piriform fossa to take down any residual attachments. One can then proceed along the external aspect of the muscle about halfway up, spreading it away from the overlying skin. Excessive external dissection will encounter blood supply to the muscle from the facial arcade.
Repositioning of the nasalis is done by anchoring the muscle to the mucoperiosteum of the canine fossa and the sulcus using a mattress suture of 4–0 vicryl. The suture is passed from the buccal sulcus upward behind the orbicularis. It loops through the muscle and is returned to the mouth where the suture is placed on a clamp. It will be approximated as the final maneuver of the surgery (Figs. 19.51 and 19.52).
Having cleaned out the piriform fossa we are now in position to complete part 2 of Talmant’s nasal fossa reconstruction. A curved elevator is passed anterior to the inferior turbinate and is then directed backward and upward until the nasal bone is reached. Using this combination of subperichondrial and subperiosteal dissection, a complete freeing of the mucoperiosteal lining of the nasal fossa is achieved. Once again, the biologic advantage of this maneuver is to reposition the stem cell envelope correctly such that osteosynthesis within the piriform fossa proceeds in a normal manner from 6 months onwards. The goal is normalization of the internal dimensions of the piriform fossa over the course of time.
Closure and Nasal Splinting
At this juncture, the dissection is complete. All cleft surgeons have their preferred sequence of steps to achieve closure. I will simply make note of a few maneuvers that I have found helpful over time. The closure sequence begins inside the nose. Access incisions to the septum and the nasal tip are addressed first. The non-philtrum flap is sutured to the release incision in the lateral nasal wall. If a vomer flap has been raised, the NP flap is sutured on its back side to the vomer flap. Otherwise, the nostril sill flap created by the releasing incision previously described is brought along the front side of the NP flap and sutured with 5–0 vicryl sutures taking care to maximize the “fit” between the two flaps. A double hook is then placed in the nose and gently lifted. The posterior margin of the NP flap is sutured to the footplate of the alar cartilage. In this way one strives to re-establish the elusive “shoulder” of the columella (Figs. 19.61, 19.62, 19.63 and 19.64).
Lateral lip height is first established by suturing the extreme tip of the DOO to its counterpart at the anterior nasal spine. Alignment of the white roll is carried out using 4–0 or 5–0 vicryl in the DOO where it curls upward toward the white roll. A confirmatory suture of 5–0 monocryl is placed at the dermal–epidermal junction of the white roll. An additional vicryl suture is placed at the wet–dry mucosal junction. With the lip on traction, the remainder of the mucosa and DOO is closed as a single layer. Final decisions regarding the M and L flaps are taken at this time. The use of the L flap as a “diaper” to cover the raw area of the medial lip vermillion often makes sense.
The extreme tip of SOO (sometimes referred to as the oblique head of the SOO) is attached by a mattress suture through the columella just below the footplate of the lateral crus on the non-cleft side. The suture is tightened to check the position but not tied. The SOO is then sutured to the non-muscle-bearing mesenchyme of the prolabium with three to four vicryl sutures. This helps achieve some bulkiness beneath the new philtral column. Several sutures of 6–0 monocryl (or nylon, if nothing else is available) carefully placed at the dermal–epidermal junction seal up the skin. A running 6–0 vicryl suture completes the closure. Ophthalmic 6–0 is particularly good because of its dissolution characteristics.
Suspension of the nasal cartilages is not a part of the Talmant technique. Nonetheless, I think that tip projection is improved by suturing the medial crura together in their new position with two sutures of 5–0 vicryl/monocryl. The unified crura can then be elevated and sutured to the septum with 4–0 vicryl/monocryl. Luis Monasterio has a very ingenious tip graft which is shaped like the number 1 with the upper limb pointing backwards. The number 1 graft is placed through a lateral columellar incision exactly the way I used to do if for the non-philtrum flap. The incision falls just anterior to the medial crus. The graft is harvested from the septum. Closure of the incision with 6–0 chromic is virtually unnoticeable. For simplicity and versatility, the number 1 graft is my first choice for secondary rhinoplasty or in adult cleft cases. The graft is fixed in place using the same suture sequence: medial crural unification followed by fixation to the septum.
Silicon stents cut from 0.5 mm sheeting are then placed (Fig. 19.67). The initial two stents are only temporary. They are curled up like snails inside each nasal cavity. An additional sheet of silicon is placed over the dorsum and the tubes are secured with two sutures of 4–0 prolene, beginning with the non-cleft side. When one places these sutures on the cleft side, a very important phenomenon takes place. On the second pass from the nasal cavity back to the dorsum, the tip of the needle is engaged with the silicon sheeting. Moving the sheeting further repositions the alar cartilage. The needle is then thrust upward through the sheeting, fixing the cartilage in its final position.
Vaseline gauze or Xeroform® gauze is then used to pack the nose. This further expands the nasal airway. During the packing phase, antibiotics are given. At 1 week, both the packing and the temporary silicone stents are removed. Nasal stenting is then continued for 4 months using the design as illustrated. Once again 0.5 mm silicon sheeting is used. In countries where this is not available, intravenous solution bags can be cut up instead.
ADDENDUM: Sotereanos Muco-Gingivoperiosteoplasty (see Fig. 19.4)
Alveolar cleft closure (with DFR modification) is extremely useful as part of secondary cleft repair or as a stand-alone procedure. The following comments are intended to simplify the steps.
Non-DFR flaps are shown in the original illustration with nasal floor closure using nasal mucosa flaps from below as a blind procedure. Under DFR protocol the nasal floor is addressed at primary lip repair (6 months) with the back wall and floor created by AEP at 18 months. If the nasal floor has not been closed, prior repair was non-DFR. Secondary lip repair is done with nasalis dissection and water-tight closure.
Step 1 Upper Left
Mucoperiosteal flap (r2, pink) GPP flaps (sliding sulcus flap) is mobilized via pericoronal incision with a back-cut up the buttress. The S flap is supplied by medial and lateral branches of StV2 infraorbital and by overlying ECA facial artery. The dotted lines indicate the level of a counter-incision that will be made along the undersurface of the flap. Mucoperiosteum of premaxilla (orange) has also a mixed blood supply from StV1 anterior ethmoid, from StV2 medial nasopalatine, and from StV2 medial branch of infraorbital. Periosteal flap on maxillary side (yellow) is supplied by greater palatine. Periosteal flap on the premaxillary side (white) has a variable blood supply. If the cleft is unilateral, it receives StV2 greater palatine + StV2 medial nasopalatine and back flow from StV1 anterior ethmoid. If the cleft is bilateral, it is perfused by the latter two arteries only. Note the cut shown between lateral incisor and canine on the non-cleft side. In DFR this is usually unnecessary for two reasons: (1) the non-cleft flap can be mobilized in a similar manner. (2) A similar periosteal releasing incision. Bilateral S flaps can cover a total of up to four dental units without tension. Dissection time is fast and bloodless, under 5 min per flap.
Step 2 Upper Right
Backwall flaps are sutured, maxillary (yellow) to premaxillary (white). The counter-incision along the undersurface of the S flap is indicated by a white line with red dots. The incision is placed at the level of the buccal sulcus. Although the S flap is mobilized over the entire face of the maxilla, the counter incision frees the alveolar component to translate with ease, with no tethering from above.
Step 3
Mesial translation of maxillary S flap (pink) and premaxillary S flap (orange). Dead space for the flaps is transferred to the first and second molar.
Step 4
Final closure with bone graft/rhBMP2 in place. Incision at premaxillary–maxillary junction on the non-cleft side is not necessary.
A closing quote, re-stated: “Cleft surgery is applied embryology.” (Victor Veau) Better outcomes for the treatment of facial anomalies will inevitably be achieved when empiricism yields to an understanding of the normal mechanisms of development (Figs. 19.89 and 19.90).

Pioneers in molecular embryology. (Top left) Luis Puelles, Universidad de Murcia, Spain. (Top right) John Rubenstein, University of California San Francisco. (Bottom left) Neural crest mapping: Nicole LeDouarin. (Bottom right) Mesodermal mapping: Drew Noden, Cornell University. [Top left: Courtesy of Luis Puelles, MD]. [Top right: Courtesy of John Rubenstein, University of California, San Francisco]. [Bottom left: Reprinted from Flickr. Retrieved from https://www.flickr.com/photos/ministere-enseignementsup-recherche/11046611646/in/album-72157638039342903/. With permission from Creative Commons License 2.0: https://creativecommons.org/licenses/by-sa/2.0/.] [Bottom right: Courtesy of Drew M. Noden PhD]

Developmental biology: engine of change for craniofacial surgery. Swiss anatomist Wilhelm His invented the microtome for histologic sections and became the father of descriptive embryology. To his right is an original drawing by His from his embryo collection. Scanning electron microscopy enabled Hinrichsen to carry out his monumental study of facial development. And now, with gene tracers and dynamic imaging, the development of the face can be understood as never before. Top left: [Reprinted from Wikimedia. Retrieved from https://commons.wikimedia.org/wiki/File:Wilhelm_His.jpeg.]. Top right: [Wilhelm His drawing, circa 1889]. Bottom left: [Courtesy of Prof. Kathleen K. Sulik, University of North Carolina.] Bottom right: [Reprinted from Gilbert SF, Barresi MJK (ed). Developmental Biology, 11th ed. Sinauer Associates; 2016. With permission from Oxford Publishing Limited.]
References
Delaire J, Precious S, Gordeef A. The advantage of widesuperiosteal exposure in primary surgical correction of labial maxil-lary clefts. Scand J Plast Reconstr Surg. 1993;22:710.
Carstens MH. Development of the facial midline. J Craniofac Surg. 2002;13(1):129–87.
Tse R. Unilateral cleft lip: principles and practice of surgical management. Semin Plast Surg. 2012;26(4):145–55.
Lumsden A, Krumlauf R. Patterning the vertebrate neuraxis. Science. 1996;274:1109–15.
Rubenstein JLR, Puelles L. Homeobox gene expression during development of the vertebrate brain. Curr Top Dev Biol. 1994;29:1–64.
Rohrich RJ, Adams WP, Ahmad J, Gunter JP. Dallas rhinoplasty: nasal surgery by the masters. Stuttgart: Quality Medical/Thieme; 2014.
Jacobs KF. Angeborene aplasia von nasensteg und knorpeligem septum. Laryngol Rhinol Otol (Stutgartt). 1984;f63:344.
Lewin ML. Congenital absence of the nasal columella. Cleft Palate J. 1988;25(1):58–63.
Mavili ME, Akyürek M. Congenital isolated absence of the nasal columella: reconstruction with an internal nasal vestibular skin flap and bilateral labial mucosal flaps. Plast Reconstr Surg. 2000;106(2):393–9.
De Souza Pinto EB, Da Rocha RP, Filho WQ, et al. Anatomy of the median part of the septum depressor muscle in aesthetic surgery. Aesthet Plast Surg. 1998;22:111.
Jardini Barbosa MV, Nahas FX, et al. Anatomy of the depressor septi nasi muscle: the basis for correction of deformities of the lip-nose junction. J Plast Surg Hand Surg. 2013;47(2):102–5.
Rohrich RJ, Huynh B, Muzaffar AR, Adams WP Jr, Robinson JB. Importance of the depressor septi nasi muscle in rhinoplasty: anatomic study and clinical application. Plast Reconstr Surg. 2000;103(1):376–83.
Sinno S, Chang JB, Saadeh PB, Lee MR. Anatomy and surgical treatment of the depressor septi nasi muscle: a systematic review. Plast Reconstr Surg. 2015;135(5):838e–48e.
Carstens MH. Neural tube programming and the pathogenesis of craniofacial clefts, part I: the neuromeric organization of the head and neck. Handb Clin Neurol. 2008b;87:247–76.
Carstens MH. Pathologic antatomy of the soft palate, part 1: embryology, the hard tissue platform, and evolution. J Cleft Lip, Palate, Craniofac Anom. 2017a;4(1):37–64.
Carstens MH. Pathologic anatomy of the soft palate, part 2: the soft tissue lever arm, pathology, and surgical correction. J Cleft Lip Pal Craniofac Anom. 2017b;4(2):83–108.
Bartezko K, Jacob M. A re-evaluation of the premaxillary bone in humans. Anat Embryol. 2004;207:417–37.
He F, Hu X, Xiong W, et al. Directed BMP4 expression in neural crest cells generates a genetic model for the rare human bony syngnathia birth defect. Dev Biol. 2014;39(2):170–81.
Zhang Y, Zhang Z, Zhao X, et al. A new function of BMP4: dual role for BMP4 in regulation of sonic hedgehog expression in the mouse tooth germ. Development. 2000;127:1431–43.
Zhang Z, Song Y, Zhao X, et al. Rescue of cleft palate in Msx-1 deficient mice by transgenci Bmp-4 reveals a network of BMP and SHH signaling in the regulation of mammalian palatogenesis. Development. 2002;129:4135–46.
Diamond MK. Homologies of the meningeal artery of humans, a reappraisal. J Anat. 1991a;178:223–41.
Diamond MK. Homologies of the stapedial artery in humans, with a reconstruction of the primitive stapedial artery configuration in euprimates. Am J Phys Anthropol. 1991b;84:433–62.
Georgiou C, Cassell MD. The foramen meningo-orbitale and its relationship to the development of the ophthalmic artery. J Anat. 1992;180:119–25.
Tessier P. Anatomical classification of facial, craniofacial, and latero-facial clefts. J Maxillofac Surg. 1976;4:70–92.
Tessier P, Hervouet F, Lekieffre M, et al. Plastic surgery of the orbit and eyelids. Paris: Masson (distributed by Year Book Medical Publishers); 1977. Translated by S. Anthony Wolfe
Ewings EL, Carstens MH. Neuroembryology and functional anatomy of craniofacial clefts. Indian J Plast Surg. 2009;42:S19–34.
Moss ML, Vilman H, Das Gupta G, Sklak R. Craniofacial growth in space-time. In: Carlson DS, editor. Craniofacial biology. Ann Arbor: University of Michigan; 1981.
Honigmann K. Experiences with the Reichert procedure in closure of unilateral cleft lips. Plast Reconstr Surg. 1980;65(2):164–8.
May H. The Rethi incision in rhinoplasty. Plast Reconstr Surg. 1951;8(2):123–31.
Reichert H. Die Formung des Philtrumsbeim primären Verschluss einseitiger Lippen-und Lippen-Kiefer-Spalten. Fortschr Kiefer Gesichtschir. 1973;16:72.
Carstens MH. Correction of the unilateral cleft lip nasal deformity using the sliding sulcus procedure. J Craniofac Surg. 1999a;10:346–64.
Carstens MH. Sequential cleft management with the sliding sulcus technique and alveolar extension palatoplasty. J Craniofac Surg. 1999b;10:503–18.
Talmant J-C. Evolution of the functional repair concept for cleft lip and palate patients. Indian J Plast Surg. 2006;39(2):197–209.
Carstens MH. The spectrum of minimal clefting: process-oriented cleft management in the presence of an intact alveolus. J Craniofac Surg. 2000b;11:270–94.
Bardach J, Kelly KM, Salyer KE. Comparative study of facial growth following lip and palate repair performed in sequence and simultaneously. Plast Reconstr Surg. 1993;91(6):1008–16.
Bardach J, Kelly KM, Salyer KE. Effects of lip repair with and without soft tissue undermining and delayed palate repair on maxillary growth: an experimental study in beagles. Plast Reconstr Surg. 1994a;94(2):343–51.
Bardach J, Kelly KM, Salyer KE. Relationship between sequence of lip and palate repair and maxillary growth: an experimental study in beagles. Plast Reconstr Surg. 1994b;93(2):269–78.
Carstens MH. The sliding sulcus procedure: simultaneous repair of unilateral clefts of the lip and primary palate – a new technique. J Craniofac Surg. 1999c;10:415–34.
Carstens MH. Correction of the bilateral cleft using the sliding sulcus technique. J Craniofac Surg. 2000a;11:137–67.
Rosenstein SW. Early bone grafting of alveolar cleft deformities. J Oral Maxillofac Surg. 2003;61(9):1078–81.
Rosenstein SW, Grassechi M, Dado DV. A long-term retrospective outcome assessment of facial growth, secondary surgical need, and maxillary lateral incisor status in a surgical-orthodontic protocol for complete clefts. Plast Reconstr Surg. 2003;111(1):1–13.
Skoog T. The use of periosteum and Surgicel® for bone restoration in congenital clefts of the maxilla. Scand J Plast Reconstr Surg. 1967;1(2):113–30.
Precious D, Delaire J. Surgical considerations in patients with cleft deformities. In: Bell WH, editor. Modern practice in orthognathic and reconstructive surgery. Philadelphia: Saunders; 1992. p. 390–425.
Carstens MH. Chapter 15: developmental field reassignment in unilateral cleft lip: reconstruction of the premaxilla. In: Losee JE, Kirschner RE, editors. Comprehensive cleft care. Lippincott; 2008a. Note: the description given in this paper departs from the Losee chapter but it contains valuable material and references.
Figallo EE, Acosta JA. Nose muscular dynamics: the tip trigonum. Plast Reconstr Surg. 2001;108:1118–26.
Bardach J, Morris HL. Multidisciplinary management of cleft lip and palate. Philadelphia: W.B Saunders; 1990.
Chancholle AR, Saboye J, Tournier JJ. Prise en charge des fentes labio-palatines par l’équipe du docteur Chancholle à Toulouse’. Ann Chir Plast Esthét. 2002;47:92–105.
Semb G, Shaw WC. Facial growth related to surgical methods. In: Berkowitz S, editor. Chap. 15: Cleft lip and palate. Diagnosis and management. 3rd ed. Berlin: Springer; 2013. p. 325–43.
Carstens MH. Chap. 1: Mechanism of cleft palate: developmental field analysis. In: Bennun RD, et al., editors. Cleft lip and palate management. A comprehensive atlas. New York: Wiley Blackwell; 2016. p. 3–21.
Talmant JC. Réflexions sur l’éthiopathogénie des fentes labio-maxillo-palatines et l’évolution de leurs traitements’. Ann Chir Plast Esthét. 1995;40:639–56.
Bartolo D, et al. Les liquides actifs, du chaos à l’ordre. La Recherche. 2022;570:92–7.
Mooney MP, Siegel MI. Premaxillary development in normal and cleft lip and palate human fetuses using three-dimensional computer reconstruction. Cleft Palate Craniofac J. 1991;28:49–54.
von Bertalanffy L. General theory of systems. Dunod; 1973.
Tokoro M. Une nouvelle méthode pour de nouveaux enjeux. Pour Sci Suppl Innov. 2015;447:64–6.
Zwirm H. Les Systèmes complexes. Odile Jacob; 2006.
Oblak P. The importance of nasal airways in cleft patients. In: Bardach J, Morris HL, editors. Chap. 82: Multidisciplinary management of cleft lip and palate. W.B. Saunders; 1990. p. 682–8.
Hairfield WM, Warren D. Dimensions of the cleft nasal airway in adult: a comparison with subjects without cleft. Cleft Palate J. 1989;26:9–13.
Millard DR, Berkowitz S, Latham RA, Wolfe SA. A discussion of presurgical orthodontics in patients with clefts. Cleft Palate J. 1988;25:403–12.
Berkowitz S. Cleft lip and palate. Diagnosis and management. 3rd ed. Berlin: Springer; 2013.
Majourau-Bouriez A, et al. Maintenir ou rouvrir l’espace correspondant à l’incisive absente ou dysmorphique au niveau de la fente alvéolaire en vue de sa réhabilitation. Prise en charge interdisciplinaire. In: Grollemund B, Etienne O, editors. Part II: Chapt 1. Fentes labio-palatines: approches thérapeutiques chirurgicales, orthodontiques et prothétiques. Editions CDP; 2022.
Malek R. Cleft lip and palate. Lesions, pathophysiology and primary treatment. Martin Dunitz; 2001.
Fudalej P, Hortiz-Dzierzbicka M, Dudkiewicz Z, Semb G. Dental arch relationship in children with complete unilateral cleft lip and palate following Warsaw (one stage repair) and Oslo protocols. Cleft Palate Craniofac J. 2009;46:648–53.
Goldwyn RM. Consecutive patients. Plast Reconstr Surg. 1990;86:962.
Talmant J-C, Talmant J-C, Rousteau G, Lumineau JP. Fentes labiales et palatines. Traitement primaire. Encyclopédie médico-chirurgicale. Techniques chirurgicales. Chir Plast Reconstr Esthét. 2018;13(4):1–21.
Herzog G. Spécificité du traitement orthodontique des fentes labio-palatines. Cas cliniques. In: Grollemund B, Etienne O, editors. Part II: Chap 1. Fentes labio-palatines: approches thérapeutiques chirurgicales, orthodontiques et prothétiques. Editions CDP; 2022.
Doucet J-C, et al. Speech outcome of early palatal repair with or without intravelar veloplasty in children with complete unilateral cleft lip and palate. Br J Oral Maxillofac Surg. 2013;51:845–50.
Touzet-Roumazeille S, et al. Osseous and dental outcomes of primary gingivoperiosteoplasty with iliac bone graft: a radiological evaluation. J Oral Maxillofac Surg. 2015;43:950–5.
Dissaux C, et al. Evaluation of 5-year-old children with complete cleft lip and palate: multicenter study. Part 1: Lip and nose aesthetic results. J Oral Maxillofac Surg. 2015;43:2085–92.
Dissaux C, et al. Evaluation of 5-year-old children with complete cleft lip and palate: multicenter study. Part 2: functional results. J Oral Maxillofac Surg. 2016;44:94–103.
Dissaux C, et al. Evaluation of success of alveolar cleft bone graft performed at 5 years versus 10 years of age. J Oral Maxillofac Surg. 2016;44:21–6.
Doucet JC, et al. Facial growth of patients with complete unilateral cleft lip and palate treated with alveolar bone grafting at 6 years. Cleft Palate Craniofac J. 2018;56(5):1–9. https://doi.org/10.1177/105566561872791.
Moreau A, et al. Speech outcome after intravelar veloplasty. J Stomatol Oral Maxillofac Surg. 2021;122:147–50.
Dissaux C, et al. Aesthetic and psychosocial impact of dentofacial appearance after primary rhinoplasty for cleft lip and palate. J Cranio Maxillofac Surg. 2021;49:914–22.
Nicol M, de Boutray M, Captier G, Bigorre M. Primary cheilorhinoplasty using the Talmant protocol in unilateral complete cleft lip: functional and aesthetic results on nasal correction and comparison with the Tennison–Malek protocol. Int J Oral Maxillofac Surg. 2022;51(11):1445–53. https://doi.org/10.1016/jijom.2022.04.004.
Quenel L, et al. Impact of orthodontic gap closure versus prosthetic replacement of missing maxillary lateral incisor on dental arch relationship and symmetry in 212 patients with cleft palate. Retrospective study. Plast Reconstr Surg. 2022;150:613e–24e. https://doi.org/10.1097/PRS0000000000009477.
Ruppert E, Wagner D. Prise en charge pluridisciplinaire autour des fentes labio-palatines, autres que l’odontologie et la chirurgie: Le syndrome d’apnées hypopnées obstructives du sommeil. In: Grollemund B, Etienne O, editors. Fentes labio-palatines: approches thérapeutiques chirurgicales, orthodontiques et prothétiques. Editions CDP; 2022.
Further Reading
Ayhan M, Sevin A, Aytüg Z, Gorgu M, Erdogan B. Reconstruction of congenital and acquired columellar defects: clinical review of 38 patients. J Craniofac Surg. 2007;18(6):1503.
Bennun R, Harfin J, Sandor G, editors. Cleft lip and plate management: a comprehensive atlas. New York: Wiley; 2015.
Bilkay U, Tokat C, Ozek C, Erdem O, Cagdas A. Reconstruction of congential absent columella. J Craniofac Surg. 2004;15(1):60–3.
Carstens MH. Functional matrix cleft repair: principles and techniques. Clin Plast Surg. 2004;31:159–89.
Carstens MH. Neural tube programming and the pathogenesis of craniofacial clefts, part 2: mesenchyme, pharyngeal arches, developmental fields, and the assembly of the human face. Handb Clin Neurol. 2008c;87:277–339.
Carstens MH. Developmental field reassignment cleft surgery: reassessment and refinements. In: Bennun R, Harfin J, Sandor G, editors. Cleft lip and palate management: a comprehensive atlas. New York: Wiley; 2015.
Demas PD, Sotereanos GC. Closure of alveolar clefts with corticocancellous block graft and bone marrow. J Oral Maxillofac Surg. 1988;46(8):682–7.
Etchevers HC, Couly G, Le Douarin NM. Morphogenesis of the branchial vascular sector. Trends Cardiovasc Med. 2002;12(7):299–304.
Fisher DM. Unilateral cleft lip repair: an anatomical subunit approximation technique. Plast Reconstr Surg. 2005;116:61–71.
Fisher DM, Sommerlad BC. Cleft lip, cleft palate, and velopharyngeal insufficiency. Plast Reconstr Surg. 2011;128(4):342e–60e.
Flores-Sarnat L, Sarnat HB, Hahn JS, et al. Axes and gradients of the neural tube form a genetic classification of nervous system malformations. Handb Clin Neurol. 2007;83:3–11.
Harashina T. Open revers-U incision technique for secondary correction of unilateral cleft lip deformity. Br J Plast Surg. 1990;86:872–81.
Honigmann K. Experiences with the Reichert procedure in closure of unilateral cleft lips. Plast Reconstr Surg. 1980;65(2):164–8.
Le Douarin NM, Kalchiem C. The neural crest. 2nd ed. Cambridge: Cambridge University Press; 1999.
Lee MR, Malafa M, Roostaeian J, Rohrich RJ. Soft tissue composition of the columella and potential relevance to rhinoplasty. Plast Reconstr Surb. 2014;134(4):621–5.
Letourneau A, Daniel RK. The superficial musculoaponeurotic system of the nose. Plast Reconstr Surg. 1988;82:48–57.
May H. The Rethi ncision in rhinoplasty. Plast Reconstr Surg. 1951;8(2):123–31.
Mohler LR. Unilateral cleft lip repair. Plast Reconstr Surg. 1987;80:511–7.
Oneal RM, Beil J. Surgical anatomy of the nose. Clin Plast Surg. 2010;37(2):191–211.
Oneal RM, Izenberg PH, Schlesinger J. Surgical anatomy of the nose. In: Daniel RK, editor. Aesthetic plastic surgery rhinoplasty. Boston: Little, Brown and Company; 1993. p. 3–37.
Onizuka T. A new method for the primary repair of unilateral cleft lip. Ann Plast Surg. 1980;4:516–22.
Pensler JM, Ward JW, Parry SW. The superficial musculoaponeurotic system in the upper lip: an anatomic study in cadavers. Plast Reconstr Surg. 1985;75(4):488–92.
Ribatti D, Nico B, Crivellato E. The role of pericytes in angiogenesis. Int J Dev Biol. 2011;55:261–8.
Reichert K. Die Formung des Philtrums beim pimären Verschluss einseitiger Lipp-und Lipp-Liefer-Spalten. Forts Kiefer Gesichtschi. 1973;16:72.
Saba Y, Amodeo CA, Hammou J-C, Poiselli R. An anatomic study of the nasal superficial musculoaponeurotic system: surgical applications in rhinoplasty. Arch Facial Plast Surg. 2008;10(2):115.
Salyer K, Bardach J. Atlas of cleft and craniofacial surgery. Philadelphia: William & Wilkins Lippencott; 1999.
Sari E. Tessier number 30 facial cleft: a rare maxillofacial anomaly. Turk J Plast Surg. 2018;26(1):12.
Song R, Liu C, Zhao Y. A new principle for unilateral complete deft lip repair: the lateral columellar flap method. Plast Reconstr Surg. 1998;102:1848–52.
Talmant JC, Talmant JC. Cleft rhinoplasty, from primary to secondary surgery. Ann Chir Plast Esthet. 2014;59(6):555–84. https://doi.org/10.1016/j.anplas.2014.08.004.
Talmant JC, Talmant JC, Lumineau JP. Secondary treatment of cleft lip and palate. Ann Chir Plast Esthet. 2016;61(5):360–70. https://doi.org/10.1016/j.anplas.2016.06.012.
Talmant JC, Talmant JC, Lumineau JP. Primary treatment of cleft lip and palate. Its fundamental principles. Ann Chir Plast Esthet. 2016;61(5):348–59. https://doi.org/10.1016/j.anplas.2016.06.007.
Testut L, Latarjet A, Latarjet M. Tratado de Anatomía Humana. Bacelona: Salvat Editores S.A; 1988.
Thomas C. Primary rhinoplasty by open approach with repair of the bilateral complete cleft lip. J Craniofac Surg. 2009;20(Suppl 2):1715–8.
Thomas C. Primary rhinoplasty by open approach with repair of the unilateral complete cleft lip. J Craniofac Surg. 2009;20(Suppl 2):1711–4.
Trott JA, Mohan N. A preliminary report on open tip rhinoplasty at the time of lip repair in bilateral cleft lip and palate: the Alor Setar experience. Br J Plast Surg. 1993;46(3):215–22.
Trott JA, Mohan N. A preliminary report on open tip rhinoplasty at the time of lip repair in unilateral cleft lip and palate: the Alor Setar experience. Br J Plast Surg. 1993;46(5):363–70.
Acknowledgements
The author wishes to credit the following individuals for their intellectual and technical contributions to developmental field reassignment cleft surgery: Paul Tessier, M.D. (deceased)—for his pioneering work in facial cleft classification; Jean Delaire, M.D., D.M.D.—for his demonstration of wide subperiosteal dissection in cleft surgery; David Matthews, M.D.—for his refinements in the design of the non-philtral prolabial flap; Jean Claude Talmant, M.D.—for his discovery of the nasalis muscle and the importance of correct nasal airway restoration; Jean-Pierre Lumineau, M.D., D.M.D.—for his unique technical advances in the management of the alveolar cleft; and Luis Monasterio, M.D.—for demonstrating the importance of the developmental field approach to cleft palate repair using the alveolar extension palatoplasty to treat or prevent fistulae and maintain dental development.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Commentary: Jean-Claude Talmant
Commentary: Jean-Claude Talmant
Writing the afterword of this multidisciplinary work on the treatment of cleft lip and palate is a delicate mission and a great responsibility. With good reason, our patients always expect more from us. May this contribution remind us that anatomy is nothing but the support of function and that we must understand it and respect it to order to convert their hopes into progress.
If I had to translate my thought in a few lines, these aphorisms would be my interpreters:
“Diagnose before you treat.” Harold Gillies
“Repair must be applied embryology.” Victor Veau
“When planning the restoration, the first consideration should be function.” Harold Gillies
History has left us with a confusing situation in which, with consistency, most cleft teams, regardless of country, treat lip/palate clefts according to their own protocol, the one they were taught, one which is reassuring and comfortable. They feel legitimate since their patients have the expected acceptable result. As Janus Bardach [46], with rare lucidity, suggested in the preface to his book, even the most respected of protocols are not evaluated over the long term for the simple reason that their less-than-optimal results deter them from doing so. It is as if we have resigned ourselves to a certain mediocrity, to the point that our primary goal has become to help patients accept their handicap by attenuating their deformity and preparing their families and society to welcome them well [47]. This renunciation traps many therapists who spend their lives following others, complacent in their routine. For them, difficulty is not an opportunity. They know. This knowledge has been passed on to them. You must not challenge it. But then, how to do better?
Should One Prioritize the Judicious Choice of Technique or to the Magic of Its Execution?
Twenty years of daily management of a generation of patients are barely enough for the surgeon to evaluate his protocol. It will take yet another generation to usefully compare that protocol with others and to identify if progress has (or has not) been made. Understanding is a matter of patience. Senior surgeons finally have the experience and the time to reflect on the consistency of their choices… if only they take the risk to share their regrets. Their doubts are born of their imprecise perceptions about cleft biology itself, and of their sense that a hierarchy of techniques actually exists; and that these techniques can somehow be put into play in an attempt to restore functional anatomy without compromising growth. Surgery is, in any case, iatrogenic. However, there exist studies that maintain that some surgeons are less iatrogenic than others [48]. These assessments are not innocent. Sneakily, they add to the confusion, attributing results to talent. Whatever the dexterity or the visual intelligence of a surgeon might be, what really underlies a successful outcome is the reasoned choice of a protocol, and of those procedures required for its execution.
Our understanding of facial growth mechanisms has not seen any major advances in recent decades. The quality of dialog required to achieve progress in the field of cleft management continues to be hindered by an uneven level of culture and knowledge of basic anatomy, combined with a lack of curiosity and a resistance to open-mindedness. Certainly, technical concepts differ from one school to another, but the extent of our misunderstanding of basic science is appalling. In 2011, a renowned chairman, commenting on my presentation on the treatment of bilateral cleft lip and palate, threw out this comment to the audience: “But, Jean-Claude, why do you think that nasal breathing might play a role in facial growth?” This question left me speechless for a moment… So we are still there!
What is the Nature of the Cleft?
Craniofacial embryology, focused on molecular biochemistry, has clarified many questions but has remained without translation into surgical practice. Nevertheless, Michael Carstens, seeking the similarities between the vascular supply of the maxilla during the last stages of embryology and the clinical classification of facial clefts established by Paul Tessier, demonstrated their perfect correspondence [49]. Thus, the very essence of the labio-maxillary-palatal clefts finds its genesis at the level in a specific development field, that of the premaxilla. A dysfunction in the forward advance of the median naso-palatine vascular pedicle which supplies this field, on or before the sixth week of uterine life, changes the course of things. The earlier this occurs, the more it affects the entire bone structure, where, according to an immutable cephalo-caudal and medio-lateral spatio-temporal chronology, there exist 3 subfields: the central incisor subfield is the most primitive, followed by that of the lateral incisor, and, finally, that of its distal extension, the frontal apophysis of the premaxilla. This latter structure doubles the thickness of the outline of the piriform orifice as it underlies the ascending apophysis of the maxilla. A deficiency state of the apophysis is present in all clefts, even in their most subtle form fruste. Most often, the entire embryological field of the lateral incisor is affected with a wide variety of expressions of the deficit of its mesenchyme, especially as an additional vascular supply, coming from the lateral naso-palatine artery and from a branch of the greater palatine artery, joins the canine sector of the lateral maxillary segment. This model enables us to better understand the frequency and the considerable variety of anomalies of the lateral incisor, from its duplication in a good number of labio-alveolar clefts, to an isolated precanine, passing through its pure and simple agenesis in more than 50% of cases of complete clefts.
This direct link between cleft lip and palate and agenesis of the lateral incisor is perfectly consistent with the chronology of embryology and the clinical experience of clefts. It is the illustration of Kundera’s aphorism: "It is at the moment of its genesis that the essence of a phenomenon is revealed.” If the volume of the mesenchyme is sufficient, the joining of the frontonasal, external nostril, and maxillary buds forms the epithelial wall whose disappearance conditions their fusion, ensuring the continuity of the facial envelope, the buccal orifice, and the tip of the nose. The diffusion of BMP-4 from the mesenchyme is necessary to inhibit the action of Sonic hedgehog (Shh) which stabilizes the epithelial wall. In the event of a cleft, the BMP-4 deficiency linked to that of the mesenchyme leaves the epithelial wall the opportunity to oppose the fusion of the buds.
Influence of a Missing Embryonic Territory on Facial Muscular Organization
In a complete cleft, the distribution of the muscular network of the facial envelope and the beginning of nasal ventilation take place posterior in time to the development of the lateral incisor field. This territory is a strategic site, the keystone, both of the narrowest section of the respiratory tract, the nasal valve, and of the intercanine distance governing the centric and symmetrical mastication useful for facial growth. It should be emphasized that this is agenesis of the lateral incisor in the context of a cleft in the maxillary structure, on its two sides (nasal and oral) and not agenesis within an arch not split with diminished consequences. The alveolo-dental and maxillary impact of clefts must therefore be approached in a much more active and efficient way. In a nutshell, early orthopedic action is needed in the cleft space to restore the width useful for canine function.
Self-organization of Muscles in Space and Time
The image of the cutaway with its radiant arrangement of the facial skin muscles centered on the orbicular muscles is in no way different from the diagram of the forces acting within the facial envelope. A rupture in this system necessarily modifies this distribution. From the 7th week of uterine life, the premuscular cells amassed in the second pharyngeal arch invade the facial envelope to join the midline, eight weeks later, thereby constructing the nasalis muscle, the cupid’s arch, and the philtrum of the lip. As soon as the premyoblasts become contractile, around the 9th week, they polarize and orient themselves where they are, according to the axis of the forces which reign within the envelope. They then come together in bundles like iron filings in an electric field [50]. If the skin muscle bundles have, by definition, a maxillary or zygomatic bone insertion initiated by the power of attraction exerted by BMP-4, their orientation responds to a simple, self-organizing rule of physics, without genetic sophistication. This concept is largely demonstrated by advances in the new physics of soft matter and active matter. Beginning in the years 2000–2010, studies in this novel field have explored the mechanisms of mobility for both intra- or extra-cellular living structures as well as their orientation [51].
With the disappearance of the embryological field of the lateral incisor, the ruptured lateral facial envelope loses its support on the midline. The new distribution of forces within it changes the muscular organization. Held back by the buccinator, the envelope finds a new anterior anchorage on the end of the small maxillary fragment. Nasalis and myrtiformis deprived of their common extensions in the nostril sill merge into a single muscular body, the lower head of which is the myrtiformis. Powerful, because very stressed by its new function, the muscle slides on the lateral crus of the alar cartilage and constrains it to a caudal rotation. Their superposition raises a vertical webbing in the nostril vestibule, the correction of which requires complete separation between myrtiformis and alar cartilage by extensive dissection, which is best performed in primary surgery. Orthopedics can only act on the width of the cleft. It has no effect on the abnormal relationship between cartilage and muscle.
Influence of the Missing Element on Fetal Ventilation
Fetal ventilation, by mobilizing the incompressible amniotic fluid 50 times/minute in the nasal fossae, has a considerable morphogenic role from the 11th week onward. Opening the nasal cavity into the oral cavity causes the expiratory pressure to drop on that side, so that the relative hyperpressure widens the opposite nasal cavity and pushes the septum and vomer back into the cleft nostril with dislocation of the septal cartilage out of its vomer groove. On the cleft side, all the structures retract, and the weak cyclic stimulation explains the quantifiable growth defect at the level of the premaxilla from the 14th week, whereas the cleft appeared between the 6th and 7th weeks and the ventilation starts at the 11th week [52]. This chronologic correlation reinforces the credibility of the hypothesis which recognizes the primordial role of fetal ventilation. The pressure states induced to fetal ventilation also explain why, in the unilateral cleft, the median vomero-septal axis is disrupted, whereas, in the bilateral cleft, the axis remains aligned. Only surgery can correct these nasal cartilaginous deformities; they are inaccessible to orthopedics. Their magnitude is such that the vomer in some total unilateral clefts can actually horizontalize, thus seeming to be integrated into the palatal vault. Exploiting this deformity by closing the hard palate with the vomerian flap does not reconstruct a normal anatomy and is not without harmful consequences, as we will see.
The Concept of Facial Envelope
In utero, the combined actions of ventilation and the facial envelope shape and transform the entire nasal capsule and upper lip into a sophisticated anatomy called upon to manage future air ventilation. The facial envelope is all the soft tissues of the face; its physical characteristics exceed the sum of those of its constituents and its mechanics become so different that it is necessary to master this concept to analyze the dysfunctions and better treat them. As for the morphogenesis of the nasal and perinasal structures, it is the work of ventilation. Nothing escapes it, from the prow of the philtrum to the concha of the turbinates, passing through the individualization of the tip of the nose, the telescoping of the plica nasi, the conformation of the nasal valve and its takeover by the commissural pillars of the facial envelope. With this in mind, one can decipher, after birth, both the role of the facial envelope in the rest posture for the control of ventilation (its only vital function), and that of swallowing, which is subordinate to it (ventilation) although synchronized.
We can read influence of the facial envelope on the more or less brachyfacial or dolichofacial evolution of each of us according to our anatomical particularities and its performance. We no longer wonder if nasal ventilation influences facial growth... We can see it! The bases of essential knowledge, concerning the facial envelope, are gathered in the report of the 76th scientific meeting of the SFODF on: “The vertical dimension” by J. Talmant et al, published in “L’Orthodontie Française,” 2003; 74, 2: pp 137–313.
Respect the Coherence of the Theory of Open Systems
The trampling of our knowledge and the confusion of protocols show that Descartes’ theory is no longer sufficient to understand the complex interactions linking ventilation and facial growth. It is necessary to appeal to the systems theory of von Bertalanffy [53]. The ventilatory system which constantly adjusts the balance between nasal and oral ventilation on demand is a particularly complex open system: It constantly exchanges and interacts with the environment [54, 55]. It is itself composed of subsystems which interact and evolve over time and with external factors, such as the change of fluid before and after birth, or the change of dentition. The ventilatory system, because it is the only one to be vital for life, is the great organizer of all the subsystems which interact with it, such as those of mastication and phonation. It is therefore not coherent for the practitioner, nor healthy for the patient, to align the teeth by extraction or close spaces in the dental arch resulting from missing teeth. This represents an optimization of the masticatory subsystem at the expense of the system as a whole, with ventilation being primordial.
Optimal ventilation is nasal, at rest and in sleep. It alone ensures the oral posture (associating lingual posture and posture of the facial envelope in charge of labial occlusion and regulation of the nasal valve) that is necessary for normal and balanced facial growth [56]. Hairfield and Warren have clearly shown that the ventilation of almost 75% of children operated on for a complete uni- or bilateral cleft is more oral than nasal. Overall on inspiration, the smallest section of the nasal passages of an adult which corresponds to that of the nasal valve is reduced by 40% on inspiration in the event of uni- or bilateral total cleft [57]. By the Hagen–Poiseuille equation, a reduction by half of the diameter of the nasal valve reduces its flow 16 times; thus we can measure the functional influence of the closure of the missing territory of the lateral incisor.
Precocious Closure of the Lateral Incisor Space: Its Consequences
After birth, the widths of the nasal valve and the piriform orifice remain essential. They are governed by the dimensions of premaxilla, and therefore by its dental contents. If the lateral incisor is missing in a total cleft, preserving its space is essential.
The Latham and Millard Protocol
We can verify the consequences of an early closure of this space sought by the supporters of preoperative active orthopedics. As soon as the two edges come into contact, gingivoperiosteoplasty leads to bone fusion, the base of which is supposed to facilitate future labial-nasal repair. Millard and Latham had developed this protocol in 1977, for the difficult treatment of bilateral clefts [58]. After the initial enthusiasm, they became disillusioned, while refraining from publishing their results. Berkowitz took charge of this, emphasizing the very harmful impact of this protocol on facial growth [59]. At that time, the retired Millard had incidentally dedicated his photo to Tony Wolfe, his former student and partner, writing on the silver print: “Tony, plastic surgery is not as easy as it seems”, thus testifying to his awareness of this unfortunate choice.
The Latham appliance took effect in 3–4 weeks, the two premaxillae separating to pass on each side of the vomer in a telescoping movement. After stopping the compression, the premaxilla was expected to resume its growth, but the vomero-premaxillary suture had been abused too much. The brutality of this form of dental orthopedics has been called into question. Nonetheless this protocol has a lesson to teach. Closure of the space was a tacit recognition of a previously existing field defect. Once fusion occurred, the deficit could no longer be reproduced naturally. Maintaining the space for the defect and then reconstructing it at the proper time are logical alternatives.
Nasoalveolar Molding by Grayson and Cutting (NAM)
Grayson has developed a "soft" version of this closure of the cleft, in 3–4 months, without forcing the vomero-premaxillary suture, with a Hotz plate on which the addition of nasal extenders simultaneously ensures the expansion of the columella and the projection of the wing dome. This nasoalveolar molding (NAM) remains quite in vogue. Like their predecessors, Grayson and Cutting were tight-lipped about their long-term results. After careful assessment, Berkowitz denounced the frequency of dental crowding and the major problems of maxillary growth; this led Cutting to announce in 2011, during a congress in Perth, in which I participated, that he had abandoned this technique in unilateral cleft. In this book, the chapter by Aurélie Majourau, begins with the case of a young patient, born in New York and treated according to the NAM protocol [60]. Figure 19.91 details the evolution, revealing the terrible repercussions of the first bone fusion of the cleft. Closing and sealing this space in order to then try to reopen it with frenzied techniques are illogical and do little for the patient other than to obligate him to undergo unnecessary and heavy procedures. Obviously, these practitioners have given no credence to the key role of the lateral incisor field. It is also true that nasal ventilation is not their cup of tea.

Revision of a left cleft operated according to Malek. Canine and central incisor are in contact. The space of the missing lateral incisor was closed early with the one-plane vomer flap. The septum touches the lateral nasal wall. The vomer lies on the nasal floor; between the two the inferior meatus is excluded from any ventilation. The nasal fossa, amputated by the width of the lateral incisor, is obstructed
The Case of the Vomer Flap
Due to its simplicity and the opportunity, it offers to treat the entire cleft at one time, the most universal technique for closing the hard palate is the one-plane vomer flap. The face of the flap left raw heals by secondary epithelialization, known for the persistence of its intense retractile power. The vomerian flap is an integral part of the Malek protocol, which has been widely used in France and in French-speaking countries in recent decades [61]. We have repeatedly observed its drawbacks by surgically exploring its labial, nasal, and alveolar sequelae. Early scar retraction closes the space of the lateral incisor, damage increased in case of agenesis. It touches the vault of the palate, an eminently functional structure which has all its growth to do. This retractile tissue installed early in the cleft is a nuisance that is not limited to the arch and the anterior palate but hits the nasal valve, the piriform orifice, and the entire nasal cavity hard. The inferior meatus, between the vomer and floor, is excluded from any ventilation while the residual space is obstructed by the septal deformity (Fig. 19.91).
Malek preferred the vomerine flap to large maxillary and palatal subperiosteal dissections, which he accused of being the cause of growth defects. Many European publications of the last decade compare protocols which, in fact, all use the vomerian flap, simple and universal. Can we be surprised by their conclusions which show that these protocols are equal and that none really stands out? [62]. The harmfulness of the vomerian flap is the most probable cause of the leveling of the results whose mediocrity, accepted with resignation, is wrongly attributed to the cleft.
It is clear that 50 years of statistics have not been enough to provide useful information. We probably didn't ask the right questions! Rare are the publications showing with the most perfect photographic rigor, the results at the end of growth (20 years of follow-up) of all the first consecutive cases operated according to a precise protocol of a unilateral and bilateral complete cleft lip and palate. The first ten cases of a protocol are binding objectives, indeed difficult to fulfill by the surgeon as well as the patient [63], but so rich in teaching! I know of only two such attempts. The first is that of Harold McComb of Perth, who became a very dear friend 30 years ago, who encouraged me, given my liberal status, to build my credibility in this way. At the end of 2018, I thus published the first 5 consecutive cases of total unilateral cleft lip and palate treated with my protocol since 1997, and also did so for bilateral clefts in the “Encyclopédie médico-chirurgicale” thanks to digital possibilities [64].
Abandoning subperiosteal dissections during labial-narinal repair limits the projection of the soft tissues in the naso-labial angle. This contributes to the "cleft look," a defect that Georges Herzog, in his chapter on the closure of the lateral incisor space, regrets in his patients after Lefort I osteotomy [65]. He is not responsible for it. He undergoes the situation created by the surgery. His chapter is very informative, with a good iconography of the palatal vault and the evolution of occlusion as is rarely shown. Only the incidences that allow the nasal sequelae to be seen are lacking. The protocol implemented in these observations is that of Malek.
Most of my students came from this protocol and adopted mine because I showed my results, and they wanted to improve theirs or those of their master. Comparative studies by them in a variety of fields do justify their choices [66,67,68,69,70,71,72,73,74,75]. The Lausanne Protocol has a Lefort I osteotomy rate close to 60% cases in complete unilateral clefts. This situates its harmfulness on growth. The 3 cases of complete unilateral clefts presented by Georges Herzog reveal the consequences of the choices made during primary surgeries that we will analyze below.
Preoperative Passive Orthopedics
It is clear that the palatal orthosis does not have the expected effects on the protection of the palatal sutures, the conformation of the arch and maxillary growth. This confirms the work of teams who have compared series of patients treated with or without passive orthopedics. They could not demonstrate any lasting advantage in any area.
The Vomerian Flap: Its Flaws
The major problem is the vomer flap itself. We are not going to insist, again, on the constant nasal obstruction of this protocol. In the first case, the vomer itself is largely integrated into the palatal vault. The resulting rigidity does not facilitate recentering of the midpoint. In another context, orthopedics in deciduous dentition on a normalized palate could have solved this problem and avoided this adventure of a Lefort I osteotomy at 9 years old, when the maxillary arch had good proportions. This resulted in the extractions of 14 and 24, a second osteotomy, the persistence of a fistula and a labial “cleft look.”
The vomer mucosa does not have the qualities required to replace the palatal fibromucosa. Observations 1 and 3 show recurrent bucco-nasal fistulas, very wide in the 3rd case. These fistulas disappeared in our two-stage palatal closure protocol. Putting the fibromucosa of the banks in its place according to Victor Veau’s principle solves this problem definitively from the primary stage. The vomer flap does not facilitate the restoration of the alveolar cleft: it contracts it. In the absence of bone graft, the quality of the periodontium at the edge of the cleft is mediocre, especially for the lateral if it is present. In our protocol, any tooth, even dysmorphic, is respected until the end of the treatment—with very rare exceptions—as a precious bone capital to be preserved in total clefts. Here, it is the dental capital which is adapted to the narrowness of the arch contracted by the vomer flap. The missing part of the embryological field of the lateral incisor that can be assessed with precision by reestablishing the maxillary intercanine distance in relation to its mandibular reference is quite simply obliterated—both compacted in the patient and erased in the mind of the therapist. Should we really continue onward this path?
The Logic: Open the Space of the Lateral Incisor
This situation can be reversed, subject to action. A labial-nasal, septal, and alveolar revision is possible at any age (the ideal is between 5 and 6 years old) and with the usual precautions (post-operative nostril conformation of 4 months). This secondary surgery is technically demanding but effective. Prepared by an anterior maxillary expansion for an iliac cancellous bone graft, it restores its effectiveness in orthodontics. Any procrastination is a waste of luck while the growth potential that is known to be good in the cleft becomes exhausted.
Aurélie Majourau’s sub-chapter on maintaining lateral incisor space provides many answers [60]. Read it and reread it. It is solid, didactic, ambitious, full of hope for today’s patients, addressing all the questions of a multidisciplinary practice where surgeons and orthodontists must play the same score with a repair that respects the structures, modeled on the embryology. Cleft orthodontics must be concerned with the need for efficient orthopedics, therefore early and active in the deciduous dentition and at the beginning of the mixed dentition. With the progress made, the prosthetic solution of the implant which requires an excellent periodontium and will stabilize the final result is the logical conclusion which awaits only a generalized implementation of the protocol and the procedures which are described, undoubtedly still perfectible, but whose broad outlines should be confirmed with time.
Nothing is easy in this pathology, but inaction is not defensible when oral ventilation can “make the bed” for obstructive sleep apnea and its consequences (See the good sub-chapter on OSAS by Elisabeth Ruppert and Delphine Wagner [76]). Orthognathic surgery is not an end in itself. It lacks the harmony of normal growth and it is not uncommon for it to fail in cleft lip and palate where recurrences are feared. It would be preferable for the quality of the primary surgery to preserve us from this scenario, but, if an orthognathic procedure becomes necessary, the primary repair surgery can create conditions for a favorable result: a reduced negative overjet, a single median scar on the palate, no fistula, concordant arches, good bone volume in the alveolar cleft, normal intercanine width with preservation of the lateral incisor space, and a functional nostril. Of course, a precanine lateral incisor of sufficient size, with a quality periodontium, must be preserved. After coronoplasty, this living tooth will be preferable to an implant.
Finally, there is a dysfunction whose considerable influence we all underestimate, especially in the clefts: the persistence of primary swallowing. This is the most probable cause of our current therapeutic limits. This subject must concern us and become a priority from the age of 5–6 years.
A Note from Dr. Carstens
Jean-Claude Talmant
The work of Jean-Claude Talmant will perhaps not be fully appreciated for another generation. He has single-handedly explored the relationship between fetal breathing mechanics facial anatomy, using facial clefts as his model. Pressures exerted normally within the nasopharynx and nasal cavity during development play an undeniable role in shaping the morphology of the lip-nose complex at birth. Dr. Talmant documented these effects fetal ultrasound. But he did not stop with merely proving an anatomic point; he carefully and methodically developed techniques to address these effects during primary surgical repair with a resulting restoration of not only breathing mechanics but of their cerebral control. Jean-Claude’s life work also demonstrates a principle that is very much in the tradition of French anatomy: the careful observation and recording of natural events over time. In this regard, his contributions are a surgical version of Marcel Proust’s revolution 7-volume masterpiece, À la Recherche du Temps Perdus (Remembrance of Things Past). Like Proust, he has lovingly captured the passage of time as reflected in the visage of each one of his patients. Although his case series is small, it has been recorded with the utmost precision and care. No one has done it better and his attention to detail remains a model for surgeons to emulate, both now and in the future.
Rights and permissions
Copyright information
© 2023 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

{kind=link}
Cite this chapter
Carstens, M.H. (2023). DFR Cheilorhinoplasty: The Role of Developmental Field Reassignment in the Management of Facial Asymmetry and the Airway in the Complete Cleft Deformity. In: Carstens, M.H. (eds) The Embryologic Basis of Craniofacial Structure. Springer, Cham. https://doi.org/10.1007/978-3-031-15636-6_19
Download citation
DOI: https://doi.org/10.1007/978-3-031-15636-6_19
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-15635-9
Online ISBN: 978-3-031-15636-6
eBook Packages: MedicineMedicine (R0)