
Abstract
The importance of the cerebellum in basic as well as higher order domains of affect processing in the brain has been vividly elaborated and specified by the contributions collected in this book. Indeed, according to increasingly precise research findings in functional neuroimaging and functional neurophysiology, individually delineable areas of the cerebellum play a role in virtually all process levels of the responsible networks of emotion perception, attribution, and experience via a variety of reciprocal connections to the limbic system and distinct areas of the parietal, temporal, and prefrontal cortex. The works in this book identify alternative perspectives in neuroscience research that offer new directions in future investigations. Important aspects will be to pin down the precise cerebellar processes in multiple sensory integration and allocation in cognitive and affective evaluation, and also cognitive-affective as well as motor behavioral responses. In this context, imaging and electrophysiological techniques will highlight the spatial and temporal, and thus the topographic and topological, specificities of the cerebellar areas to the respective networks. In the final chapter, questions and suggestions for future neuroscientific investigations are identified, from whose developments several fields of neurological and psychological disciplines could benefit in order to open up therapeutic avenues for people with cerebellar disorders.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
We believe that the present compilation of basic and advanced principles of the cerebellum in emotive functions provides a timely and representative overview, fostering the fascinating development in neuroscience to unravel the neural mechanisms of human emotions. More specifically, appreciating the detailed outlining of the chapters in this book, the field is moving on to establish a more comprehensive understanding of the involvement of the cerebellum in many different avenues of emotion in the human brain. Overall, the emotional cerebellum appears both simple and complex at the same time. As the contributions in this book demonstrate, the cerebellum impacts a broad range of brain functions to basic as well as high-order affective processing domains, supporting the complex behavioral outcomes at the motor, cognitive, and emotion levels. With the growing evidence of neuroimaging, electrophysiological, and clinical study protocols, this book complements the ongoing research disentangling the underlying mechanisms and pathways of cerebellar actions in the widespread, even large-scaled neural affective and cognitive networks of the brain in different aspects of emotion.
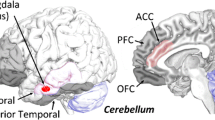
Emotion is a complex construct and still an open area of research, including its precise frame of definition (Lindquist and Barrett 2012; Scarantino and de Sousa 2021). This debate should be kept in mind in particular in appreciating the hitherto findings, and of course the intention, to describe the specific role of the cerebellum in emotion. Moreover, capturing the specific cerebellar computations of emotion-related information is complicated by the fact that in most studies the involvement of the cerebellum in emotion processing is tied to the condition of correlative rather than constitutive findings, which often lead to a report besides the central scope of intended investigation of the cerebral activations. According to task-based fMRI studies and meta-analytic approaches including step-wise heuristic models (Ebneabbasi et al. 2021), five large-scale brain networks have been identified that govern basic emotion processing and emotion regulation. (1) The visual and (2) auditory cortical regions for registering sensory stimuli, whereby (3) the salient information is processed in the insula, anterior cingulate cortex (ACC), and various subcortical regions (e.g., hypothalamus), (4) the self-referential importance of a stimulus appraised in the medial prefrontal and posterior cingulate cortex (PCC), and (5) the emotional responses by the amygdala and fusiform gyri (Riedel et al. 2018). Following the growing literature of functional neuroimaging in emotion, the responsible neural networks of emotion processing in bottom-up levels and emotion regulation in top-down levels emerge as interrelated (Morawetz et al. 2017), in particular highlighting the strong interaction between the amygdala and PFC for guiding a scaled regulatory behavior (Doré et al. 2017). The amygdala is generally assumed to be involved in the encoding of motivationally relevant stimuli, therefore generating the expressive and affective characteristics of the emotional response, whereas the PFC (particularly the DLPFC and VLPFC) is associated with the modulation of appropriate emotional response (Dixon et al. 2017). Especially, the VLPFC with its specific role in evaluating the ongoing emotion response is expected to project the regulation urgency to DLPFC for emotion regulation initiation (Etkin et al. 2015). Thus, the functional connectivity between these cerebral structures, in particular between the amygdala and the prefrontal cortex, is presumed to be crucial in emotion perception and behavior and psychopathology (Ebneabbasi et al. 2021). As articulated in the chapter of Thomasson and Peron, the neuroscience of emotion has long been focusing on the cortico-limbic networks and less so on other potentially relevant regions including the basal ganglia and cerebellum. In fact, the close anatomical connections between the basal ganglia and cerebellum in conjunction with their reciprocal connections of the cerebellum provide a neuroanatomic blueprint for temporally structured event representations in the stage transition from the detection of salient (emotional) information to motivational tendencies and motor responses. According to previous data on emotion processing in the brain, early and late stages can be distinguished, which essentially characterize mechanisms of bottom-up and top-down control processes (Sabatinelli et al. 2013). The vermis and hemispheric areas of the cerebellum were identified as contributing to the functional stages identified for the cerebral process levels. Not only for the early steps of perception and recognition of emotional cues, but also for the late integrative stages of emotional evaluation (Adamaszek et al. 2013; Styliadis et al. 2015). Subsequently, cerebellar signal processing is important for the target processes within cerebral pathways for not only optimal emotional, but also cognitive and motor expressive behavior, including in a social context. In this context, distinct areas of the cerebellum are assigned a core mode of associative learning in emotion processing, which supports excitatory and inhibitory neural activities within task-dependent networks of motor, cognitive, and affective behavior (Adamaszek et al. 2017). More specifically, the cerebellar role in these different domains of emotion perception, recognition, and evaluation deems to be anchored within its fine adjusted connections to associative cortical brain regions, in particular the prefrontal and parietal areas, by driving the neural information along internal models as already built, or going to be build, within the responsible cerebral-cerebellar loops as part of the task-dependent large-scaled neural networks (Ito 2008). This assumption is in line with the proposal of the cerebellar role in interfacing and therefore supporting the predictive property of complex patterns of neural information along specific task-related brain networks. The intracerebellar distribution to these task-dependent network processing is nevertheless an exciting request in current research, in particular considering the meta-analysis data of evidence from neuroimaging about the cerebellar contributions to cognition, showing that there are both distinct and overlapping cerebellar regions involved across these tasks depending on task demands (Stoodley and Schmahmann 2009; Keren-Happuch et al. 2014). Neuroimaging analysis ruled out a map distribution for associative learning in the cerebellum with a medial-to-lateral cerebellar contribution along emotional, motor, and cognitive functions (Schmahmann et al. 2019), coupling distinct task-dependent cerebellar areas to nonmotor functions in parallel to the evolutionary development of the large nonmotor portions of the cerebellum, prefrontal, and association cortices, which is consistent with a phylogenetic expansion in size of the cerebellum parallel to the growth of the frontal cortex (D’Angelo and Casali 2013; Marek et al. 2018), and as a result representing a behavioural variable of brain evolution (Barton 2012). These observations might be of interest in particular for the cerebellar activities to various emotion domains, which should consider also the cognitive and motor aspects in specific behavior lines.
Numerous intriguing issues about the topographical organization of the cerebellum in the generation and experience of emotion have been provided in the chapter of Habas. The challenge of gathering available neuroimaging literature about the cerebellum and emotion has resulted in a considerable amount of data which points toward distinct areas of the cerebellum in emotion processing. However, there are still outstanding conceptual and methodological issues in neuroimaging research about precise cerebellar mechanisms in emotion that need to be addressed. For example, the vast majority of brain neuroimaging studies in affective neuroscience remains focused on the cortico-limbic networks. In addition, the cerebellum is also technically tied to a low signal-to-noise ratio of blood oxygenation level dependent (BOLD) due to location and complex folding. Furthermore, fMRI of the cerebellum in task-dependent recordings faces different metabolic patterns without aerobic glycolysis, leading to weak or even absent cerebellar activations when assessed by blood oxygenation levels (Marek et al. 2018). Finally, even though the cerebellum demonstrates a reliable, individual-specific network organization, across individuals the cerebellum is significantly more variable than the cerebral cortex (Marek et al. 2018). These individual differences in variability appear largest in the lateral posterior lobes of the cerebellum, and future studies have to establish whether or not the observed signal variability subserves functional relevance and, if so, how this is coupled to the cortical association networks. Despite these challenges, the overview by Habas of the topographic mapping and task-based functional imaging of the cerebellar lobules provides fairly clear-cut insights into the distinct functional contributions of the vermis and hemispheres. In particular, the connections of the vermis to a limbic core network and main intrinsic networks constitute an “emotional” part of the cerebellum. According to this assumed involvement in a general emotion network (GEN), the cerebellum deems to be implicated in a complex functional topography of basic emotions, which follows a multi-dimensional aspect subserved by a context-specific and dynamic recruitment of functionally connected networks (Raz et al. 2016). The topographically subdivided cerebellar areas could therefore be regarded as active integrators and part of bridging circuits contributing to different task- or situation-dependent actions, reflected by specific and variable recruitment and synchronization of distinct networks in charge of processing multiple dimensions, including arousal and valence of emotions. This idea nicely fits with the growing insights of the cerebellum to linear and parallel signal processing in cognitive and emotion processing, expressing a high degree of interindividual and task-dependent differences in brain activation patterns (Marek et al. 2018).
Approaching the neurophysiological signatures of the cerebellar role in the emotion-dedicated cortical-subcortical networks, the chapter of Adamaszek and Kirkby provided an extensive discussion of available EEG research. The authors show that sensitive early and late potentials of the cortical-subcortical EEG activities in emotion tasks are of concern in delineating cerebellar inputs in guiding the top-down control of incoming affective information along bottom-up stages of primary and visual and auditory associative areas in encoding and forwarding emotion processing in responsible networks of the human brain (Schutter and van Honk 2009; Adamaszek et al. 2015; Styliadis et al. 2015; Grass et al. 2016; Houston et al. 2018). In fact, research of the neurophysiology of the cerebellum in emotion processing has shown a bias to negative basic emotions such as anger and fear, prompting special involvement in unconscious and conscious processing in affective regulation in particular for predicting suitable motor and cognitive as well as affective patterns of the desired behavioral outcome. This interpretation concurs with evolutionary accounts on the rapid activation of fight-flight responses when facing immanent threat. Moreover, recent research has fostered the multiple representations of cognitive and affective processing engaging focal areas within different cerebellar regions in parallel. More precisely, gradient analysis of relationships between functionally distinct cerebellar regions emphasizes the existence of at least three nonmotor representations in the cerebellum and a cerebellar functional specialization following a graded organization of goal-directed (i.e., task-specific) and goal-undirected (i.e., task-specific) processing (Guell et al. 2018; Schmahmann et al. 2019). Notwithstanding the debated concepts on emotions related to valence and arousal, or the discussion about the neuropsychological assumption of distinct universal emotions (Lindquist et al. 2016), the cerebellum could be assumed to interface among the different neural signal processing involved in bottom-up and top-down control at the millisecond level, leading to unified perceptions and actions to emotional triggers (Freeman and Ambady 2011; Dolcos et al. 2020). This interface-like function is based upon complex integration of timing and sequential steps of sensory information processing, which are tied to specific cerebellar-cerebral feedforward processing involving the parietal and prefrontal cortex with its subregions of the DLPFC and VLPFC, and also the OFC. These areas are not only of specific importance to emotion processing with their leading role in emotion conjugation and control, but also as being functionally coupled to subcortical areas such as the amygdala and the basal ganglia subserving for the appraisal of emotion (Pessoa 2013; Ebneabbasi et al. 2021). The importance of delineating the underlying temporal dynamics of cerebellar input to the segregated large-scaled networks in emotion processing along perception, recognition, and expression might be of interest to get a better understanding of the functional deficits caused by cerebellar lesions, resulting in timing and/or impediments in synergy of signal processing within the responsible networks. The fact that cerebellar damage can lead to impairments in predictive computations and goal-directed behavior (Cabaraux et al. 2020) may offer therapeutic opportunities by way of promoting neuroplasticity in the affected areas.
The chapter of Han about the cerebellar contributions to morality points to the now increasingly recognized relationship of emotional aspects between moral reasoning and behavior, which allows for an alternative concept to be developed that includes emotion in moral functioning (Bebeau 2002; Kristjánsson 2007; Han 2014). In particular, the connections between Crus I and Crus II and the ventromedial areas of the prefrontal cortex and precuneus in conjunction with the default mode network (DMN) and the central executive networks (CEN) are implicated in the emotive and cognitive aspects of morality (Habas et al. 2009; Keren-Happuch et al. 2014; Van Overwalle et al. 2015). Crus I and Crus II appear to operate as a hub for the networks regulating cognitive and affective aspects of moral functioning. In consideration of Han’s intriguing findings in his chapter, these two cerebellar areas within the right cerebellar hemisphere represent an exciting and potentially critical node in the neural circuitry underlying moral functioning. In concern to the role of the cerebellum in emotion in terms of its functional significance in human behavior, social cognition is one exciting subject of research (see a recent overview in Van Overwalle et al. 2020). The main focus of research has been on cognitive processes, revealing cerebellar activities to basic social skills like joint attention (Gordon et al. 2013), mentalizing and mirroring (Van Overwalle et al. 2014), and theory of mind (Mothersill and Donohoe 2016). Laricchiuta and colleagues discussed the possibility that the cerebellum could be an important hub associated with empathic abilities and social skills, impaired emotional identification and expression (e.g., alexithymia). This possibility is analyzed within the context of embodiment theory, which posits that cognitive and emotional processes are inherent parts of our biological constitution and depend on perceptual, somatosensory, motor, neuroendocrine, and autonomic nervous system activities (Critchley and Garfinkel 2017). Research has shown that the affective parts of empathy (e.g., feeling the emotion of the pain of others) are associated with activation of the posterior vermis, while the posterolateral regions, in particular Crus I and Crus II, show distinct activation when the cognitive aspects of empathy (e.g., understanding the emotion of another person) are involved. Additionally, D’Agata and Orsi in their chapter about the cerebellum in emotion recognition give a comprehensive overview in which they provided evidence for participation of the cerebellum in the underlying neural networks composed of the OFC, ACC, anterior insula, amygdala, lower ventral areas of the temporal lobes, fusiform gyrus, with their different activations along fast and slow neural strands (Adolphs 2009). Indeed, clinical and neurofunctional research has shown cerebellar involvement in the perception and evaluation of facial or bodily expressions (e.g., Schraa-Tam et al. 2012; Ferrucci et al. 2012), whereby the cerebellum takes part in the early and also late processing stages (Adamaszek et al., 2015). With the focus on the current aspects of cerebellar influences onto specific incoming sensory signals, the presumption of a specific role of the cerebellum in the perception and expression of music and art is congruent, with its demands of fine-graded resolution and its connections to forwarding to eligible brain regions, including the parietal, temporal, and prefrontal cortical parts in the further evaluation of artistic pieces and selection of suitable behavioral response. Similarly, the cerebellum has been proven to take part in several aspects of creativity in art, which involves a complex interplay between motor, cognitive, and emotional processes. In the chapter of Evers and Tölgyesi, the authors give a precise outlining of the cerebellar involvements not only in music perception and recognition, but also in production. They highlight specific network associations between certain parts of the cerebellum, in particular lobule VI and VII, the latter lobule with its Crus I and II, and the cerebral areas encompassing the frontoparietal and temporal cortex. In fact, the authors gathered a multitude of different components involved not only in music perception, including melody, harmony, rhythm, but also in the recognition of music chords, which emphasize the impact of memory and emotional processing to the temporal driven integration of sensory information during music perception and evaluation (Parsons 2001; Molinari et al. 2003; Paquette et al. 2017). Evers and Tölgyesi point out that from a topographical point of view, areas of the left hemisphere of the cerebellum deem to take a predominant role within music perception, which might be due to its specific role in low-pass filtered processes of prosodic and melodic properties, whereas the right hemisphere of the cerebellum might be more involved in segmental properties of music chords. A comparable topic related to the issue of cerebellar involvement in more complex functions was addressed in the chapter of the impact of the cerebellum in the experience of art, which follows the growing research field of neural underpinnings in aesthetics. According to the “aesthetic triad” of Chatterjee and Vartanian (2014), aesthetic experience represents the flexible interplay between the sensory-motor, emotion-evaluation, and meaning-knowledge levels along low- and high-level processes to aesthetic judgments in particular about visual and auditory stimuli from the occipital to frontal lobes (Boccia et al. 2016). Research on the role of the cerebellum in aesthetics has identified involvement of the posterior parts of the cerebellar hemispheres and sites within the vermis and paravermal regions. Thus, the cerebellum is suggested to be involved in the behavioral levels of creativity linking its unique cognitive and affective aspects (Petrosini et al. 2015), presumably by providing several internal models to solve the demands of motor, cognitive, and affective processing traces of the responsible networks in specific multimodal task processes (Ito 2008). Indeed, the reciprocal connections between the cerebellum and the lateral (DLPFC, IFG) and medial (vmPFC) areas of the PFC could, on the one hand, serve as an interface between intracerebellar convergence zones in order to progress to specific and novel outcomes within implicit and explicit processes of problem solving (Koziol and Lutz 2013) as a feature of creativity. On the other hand, the cerebellar contributions to art perception suggest similar modes of neural activities, as shown for other sensory processes with associative perception and recognition, such as emotional faces and vocalization. Therefore, activation of Crus I and II is frequently reported in specific fMRI tasks studying cerebral processing of emotion-laden visual and auditory art (e.g., Zeki et al. 2014), leading to the presumption that these cerebellar regions act as central hubs in diverse large-scaled neural networks. These cerebellar hubs arguably support aesthetic processing across different stimulus categories including not only paintings, music, and architecture, but also morality and mathematics (e.g., Kühn and Gallinat 2012; Vartanian and Kaufman 2013; Zeki et al. 2014). So art perception arguably involves the distribution of bottom-up and top-down processing stages that can be captured by early and late potential deflections in event-related potential protocols (Jacobsen 2013). The exact mechanisms of the responsible parts of the cerebellum serving as integrative modes need to be clarified in future studies.
One way of addressing the direct relationship between the cerebellum and the different emotion-related processing streams is with noninvasive brain stimulation techniques such as transcranial magnetic and direct current stimulation. These techniques can induce transient modulation of neural excitability of the human cerebellar cortex and associated (extracerebellar) regions. While cerebellar neurostimulation in the field of affective and clinical neuroscience is still in its initial phase, empirical evidence confirms direct involvement of the cerebellum in emotion and motivation. In addition, the chapter of Van Dun and colleagues deals with potential therapeutic options in affective disturbances in cerebellar disorders using transcranial magnetic and electric stimulation techniques. In consideration of the positive results described by Van Dun and colleagues in particular regarding cerebellar stimulation in the treatment of ataxia and emotion-related disorders, cerebellar stimulation protocols have been developed that may have neurorehabilitative potential. As Van Dun and colleagues outlined, the central aims of such neurorehabilitative approaches should be about targeting neural restoration of the responsible cerebellar circuitries. Moreover, specific psychological-oriented therapies, such as cognitive-behavioral therapy and/or goal management therapy to increase patient’s awareness of acquired disabilities, may serve as a kind of “external cerebellum” as an additional mode of therapy through the implementation of self-monitoring strategies (Ruffieux et al. 2017). In fact, a combined setting including such cognitive-behavioral approaches, psychiatric and occupational therapies, and individual adapted medical and noninvasive stimulation may be particularly beneficial to establish clinically relevant effects. However, the therapeutic efficacy of these combined and add-on treatment approaches warrants further research.
Following the chapter of Van Dun and colleagues about rehabilitative issues in the context of the cerebellum and emotion, Evers and Tölgyesi in their chapter reported some fascinating study observations of therapeutic implications of music targeting the neural reorganization within the cerebellum or the affected cerebello-cerebral connections, which has been proven in different clinical entities such as Alzheimer’s disease (King et al. 2019) or unresponsiveness wakefulness syndrome (Steinhoff et al. 2015). In sum, if plasticity of the brain following cerebellum lesions is a common characteristic (Bower and Parsons (2003), then the positive results of neurorehabilitative protocols for movement disabilities in cerebellar disorders may prompt comparable strategies for ameliorating deficits of perceiving and handling emotional cues (Ilg et al. 2014). According to the observation of cerebellar involvement in action perception coupling (APC) as an example of integration of motor and multisensory information (Christensen et al. 2014), a promising therapeutical approach might be a structured training of perceiving and evaluating visually presented scenes of daily situations with certain affective conditions or even practicing emotional facial expressions of basic emotions such as fear and anger.
Regarding the underlying mechanisms of the relevant cerebellar areas in guiding emotion processing, there is a highly uniform and stereotyped geometry at the microscopic level. The cerebellum can presumably be broken down to functional units, whereby several hundreds and thousands of multizonal microcomplexes serve as functional modules (D’Angelo and Casali 2013). Up to now, there is increasing evidence that the cerebellar areas work along corticonuclear microcomplexes subserving computational functions unique to the cerebellum. This overarching concept, known as the universal cerebellar transform (UCT), is considered a crucial feature of the cerebellum for the integration of internal representations with external stimuli and self-generated responses in an implicit and non-conscious manner (Schmahmann et al. 2019). Indeed, Baumann and Mattingley point out in their chapter that despite the value of a universal theory on cerebellar function, the unique contributions of the cerebellum (i.e., functional units) to specific functional domains are nonetheless important to consider. The authors show that the cerebellum partakes in several components of emotion processing, including physiological (bodily) responses underlying the “feeling” component of emotion, emotional expressions in social interactions, and cognitive appraisal processes linked to determine the significance of events and the subsequent regulation of emotions elicited by these events.
Similar to movement-related operations, the cerebellum coordinates and integrates cognitive and affective functions. Damage to the cerebellum can not only lead to ataxic motor symptoms (motor dysmetria), but also cause cognitive and emotional disturbances (dysmetria of thought) (Schmahmann 2021). Emotional dysmetria is now evolving. Within this framework, various hypotheses have been put forth that might form the basis of the UCT, such as error-based learning, error monitoring, forward control, prediction, timing, or sequencing (Argyropoulos et al. 2019). Notwithstanding the challenge to explain how a single brain structure whose neural circuitry is organized into uniform, repetitive pattern can play such an integral role in so many distinct functions and behaviors (Bower and Parsons 2003), the UCT model might be of relevance also to the cerebellar modes in emotion. This might be of interest for future research to further clarify the core computations and adaptability of the cerebellum in emotion and subsequent behavior. Sensitive clinical and neuropsychological assessments as well as fine-grained behavioral paradigms in functional neuroimaging targeting crucial cerebellar-cerebral pathways are warranted to address these questions on the specific intracerebellar computations of emotion and cognition (Argyropoulos et al. 2019; Schmahmann et al. 2019). As the clinical impact of cerebellar-related impairments in emotion perception and experience are know well recognized at the neuropsychiatric level (Schmahmann and Sherman 1998; Wolf et al. 2009; Schutter 2016; Zhou et al. 2017; Schutter 2020; Schmahmann 2021), psychopathological conditions can be conceptualized as either an emotional overshoot (hypermetria) or undershoot (hypometria) for the five neuropsychiatric domains of the cerebellum, that is (1) attentional and (2) emotional control, (3) social skills, (4) psychosis spectrum disorders, and (5) autism spectrum disorders (ASD) (Argyropoulos et al. 2019). Indeed, neuropsychological and experimental brain research has provided independent lines of evidence in support of cerebellar involvement in disorders of emotion. Medial cerebellar structures and their connections to the limbic system are involved in visceral aspects and generation of emotions, whereas the posterolateral cerebello-thalamo-cortical loops are implicated in emotion regulation and subjective sense of control. Disturbances within these cerebellar-centered circuits are proposed to underlie homeostatic dysregulation and emotion dysregulation that may perhaps even provide a transdiagnostic mechanism by which the cerebellum contributes to neuropsychiatric conditions. As the dysmetria of thought concept defines psychological issues from a cognitive-centered perspective, the term dysmetria of affect may be a more appropriate term when the psychological disturbances find their (subcortical) origins in the generation and experience of emotional states (Schutter 2020). As such, the significance of different cerebellar areas in various domains of emotion, as presented in this book, is in accordance with the notification of clinically relevant sequelae of cerebellar caused emotion processing (Schmahmann and Sherman 1998), substantiating the high interest for ongoing and outstanding research. Therefore, several issues in affective and clinical neuroscience to uncover the cerebellar principles and mechanisms of emotions and related disorders remain to be clarified in future research:
-
Topographical mapping and task-based functional imaging of the cerebellum have propagated the knowledge about intracerebellar areas not only in different categories of emotion such as valence and arousal, but also in combination to predominately cognitive processing modes. It would be interesting to clarify the specific topographical patterns of emotion processing within the responsible cerebellar regions in more detail, that is, by applying multivariate pattern analyses (Haxby et al. 2014). Moreover, it would be of interest to clarify the temporal features of each responsible cerebellar areas in more detail, capturing the task-dependent differences to each dynamic functional connectivity. In addition, intracerebellar connectivity between the medial and lateral regions, and the peculiarities of the regional overlaps, should be taken into account in future neuroimaging protocols. Finally, the importance of the intrinsic connectivity networks (ICN) subserving for maps of the cerebellum to various functional networks such as the default mode network (DMN), salience network (SN), and frontoparietal network (FPN) to certain associative processing in several domains such as creativity, imagination, or morality is warranted to be clarified.
-
Clinical and neuroimaging studies should disentangle the specific characteristics of the cerebellum, in particular its connections to limbic areas and the frontoparietal and temporal cortex. In this sense, future studies should shed a light onto the specific spatial segregation to evolutionary new regions of the cerebellum in terms of functional connectivity with limbic versus association cortices.
-
Does the cerebellum serve a more general role in emotion processing, or are there dedicated and functionally specialized regions? Moreover, how does the cerebellum contribute to the unconscious (subcortical) and conscious (cerebral cortical) processing streams?
-
Additional studies with specific moral task conditions as well as protocols with focal noninvasive stimulation of the posterolateral regions may obtain further detailed insights into the role of the cerebellum in morality.
-
Concerning the higher order associative processing of emotion, more research is needed to get a better understanding of how the cerebellum learns to support the discrimination of emotional valence in associative processing of multifaceted sensory perception and evaluation in music and visual art. So while the cerebellum has been found to be involved in art perception, explicating the stages deemed crucial in recognizing and evaluating the emotion features warrants further systematic investigation. Moreover, what are the functional contributions of Crus I and II to networks such as the default mode network (DMN) and central executive network (CEN) in the context of cognitive and affective processes in creativity and aesthetic perception? Finalizing these roads to specific avenues of the cerebellum to art disciplines, suitable music and art training protocols serving for therapeutic applications in the use of active as well as passive therapies to enhance the neural organization of afflicted neural cortical-subcortical networks need to be clarified.
-
Uncovering the precise role of the cerebellum in emotional prosody (i.e., the melodic and rhythmic features of speech that convey the emotional content) and the ability to recognize and describe one’s emotions (or lack thereof called alexithymia), as introduced in the chapter of Laricchiuta and colleagues, is an outstanding issue in language studies.
-
How do microcomplexes regulate signals related to emotion and how the cerebellar modules interfere to smooth motor and behavior? Specifically, what is the role of the climbing and mossy fibers in relationship with emotional regulation, and how these fibers interact with the monoaminergic afferent system? And will it be possible to extract patterns of discharges of the cerebellar cortex noninvasively?
-
Future studies are warranted to clarify individual parameters of transcranial cerebellar stimulation protocols, in particular the stimulation modes and target regions, which should be evaluated in more detail by neuroimaging approaches. In addition, neurorehabilitation protocols involving clinical physiatrist and occupational, psychological, and also individually tailored pharmacological treatment approaches should be evaluated in addressing affective disturbances following cerebellar disorders. Finally, the subdivision of the cerebellar syndrome into three subtypes (cerebellar motor syndrome, cerebellar vestibular syndrome, Schmahmann’s syndrome) can be used as a template to improve our handling of cerebellar disorders (Manto and Marien 2015).
References
Adamaszek M, Olbrich S, Kirkby KC, Woldag H, Heinrich A (2013) Event-related potentials indicating impaired emotional attention in cerebellar stroke: a case study. Neurosci Lett 548:206–211
Adamaszek M, Olbrich S, Kirkby KC, D’Agata F, Langner S, Steele CJ, Sehm B, Busse S, Kessler C, Hamm AO (2015) Neural correlates of disturbed emotional face recognition in cerebellar lesions. Brain Res 1613:1–12
Adamaszek M, D'Agata F, Ferrucci R, Habas C, Keulen S et al (2017) Consensus paper: cerebellum and emotion. Cerebellum 16(2):552–576
Adolphs R (2009) The social brain: neural basis of social knowledge. Ann Rev Psychol 60:693–716
Argyropoulos GPD, Van Dun K, Adamaszek M, Leggio M, Manto M et al (2019) The cerebellar cognitive affective/Schmahmann syndrome: a task force paper. Cerebellum 19(1):102–125
Barton RA (2012) Embodied cognitive evolution and the cerebellum. Phil Trans Soc B 367:2097–2107
Bebeau MJ (2002) The defining issues test and the four component model: contributions to professional education. J Moral Educ 31:271–295
Boccia M, Barbetti S, Picardi L, Guariglia C, Ferlazzo F et al (2016) Where does brain neural activation in aesthetic responses to visual art occur? Meta-analytic evidence from neuroimaging studies. Neurosci Biobehav Rev 60:65–71
Bower JM, Parsons LM (2003) Rethinking the “lesser brain”. Sci Am 289(2):50–57
Cabaraux P, Gandini J, Kakei S, Manto M, Mitoma H, Tanaka H (2020) Dysmetria and error predictions: the role of internal forward model. Int J Mol Sci 21:6900
Chatterjee A, Vartanian O (2014) Neuroaesthetics. Trends Cogn Sci 18(7):370–375
Christensen A, Giese MA, Sultan F, Mueller OM, Goericke SL et al (2014) An intact action-perception coupling depends on the integrity of the cerebellum. J Neurosci 34(19):6707–6716
Critchley HD, Garfinkel SN (2017) Interoception and emotion. Curr Opin Psychol 17:7–14
D’Angelo E, Casali S (2013) Seeking a unified framework for cerebellar function and dysfunction: from circuit operations to cognition. Front Neural Circuits 6(116):1–23
Dixon ML, Thiruchselvam R, Todd R, Christoff K (2017) Emotion and the prefrontal cortex: an integrative review. Psychol Bull 143(10):1033–1081
Dolcos F, Katsumi Y, Moore M, Berggren N, de Gelder B, Derakshan N, Hamm AO, Koster EHW, Ladouceur CD, Okon-Singer H, Pegna AJ, Richter T, Schweizer S, Van den Stock J, Ventura-Bort C, Weymar M, Dolcos S (2020) Neural correlates of emotion-attention interactions: from perception, learning, and memory to social cognition, individual differences, and training interventions. Neurosci Biobehav Rev 108:559–601
Doré BP, Weber J, Ochsner KN (2017) Neural predictors of decisions to cognitively control emotion. J Neurosci 37(10):2580–2588
Ebneabbasi A, Mahdipour M, Nejati V, Li M, Liebe T et al (2021) Emotion processing and regulation in major depressive disorder: a 7T resting-state fMRI study. Hum Brain Mapp 42:797–810
Etkin A, Büchel C, Gross JJ (2015) The neural bases of emotion regulation. Nat Rev Neurosci 16(11):693–700
Ferrucci R, Giannicola G, Rosa M, Fumagalli M, Boggio PS et al (2012) Cerebellum and processing of negative facial emotions: cerebellar transcranial DC stimulation specifically enhances the emotional recognition of facial anger and sadness. Cognition Emotion 26(5):786–799
Freeman JB, Ambady N (2011) A dynamic interactive theory of person construal. Psychol Rev 118(2):247–279
Gordon I, Eilbott JA, Feldman R, Pelphrey KA, Vander Wyk BC (2013) Social, reward, and attention brain networks are involved when online bids for joint attention are met with congruent versus incongruent responses. Soc Neurosci 8(6):544–554
Grass A, Bayer M, Schacht A (2016) Electrophysiological correlates of emotional content and volume level in spoken word processing. Front Hum Neurosci 10:326
Guell X, Gabrieli JDE, Schmahmann JD (2018) Triple representation of language, working memory, social and emotion processing in the cerebellum: convergent evidence from task and seed-based resting-state fMRI analyses in a single large cohort. NeuroImage 172:437–449
Habas C, Kamdar N, Nguyen D, Prater K, Beckmann CF, Menon V, Greicius MD (2009) Distinct cerebellar contributions to intrinsic connectivity networks. J Neurosci 29(26):8586–8594
Han H (2014) Analyzing theoretical frameworks of moral education through Lakatos’s philosophy of science. J Moral Educ 43:32–53
Haxby JV, Connolly AC, Guntupalli JS (2014) Decoding neural representational spaces using multivariate pattern analysis. Annu Rev Neurosci 37:435–456
Houston JR, Hughes ML, Lien MC, Martin BA, Loth F, Luciano MG, Vorster S, Allen PA (2018) An electrophysiological study of cognitive and emotion processing in type I chiari malformation. Cerebellum 17(4):404–418
Ilg W, Bastian AJ, Boesch S, Burciu RG, Clenik P et al (2014) Consensus paper: management of degenerative cerebellar disorders. Cerebellum 13(2):248–268
Ito M (2008) Control of mental activities by internal models in the cerebellum. Nat Rev Neurosci 9(4):304–313
Jacobsen T (2013) On the electrophysiology of aesthetic processing. Progr Brain Res 204:159–168
Keren-Happuch E, Chen S-HA, Ho M-HR, Desmons JE (2014) A meta-analysis of cerebellar contribution to higher cognition in PET and fMRI studies. Hum Brain Mapp 35(2):593–615
King M, Hernandez-Castillo CR, Poldrack RA et al (2019) Functional boundaries in the human cerebellum revealed by a multi-domain task battery. Nat Neurosci 22:1371–1378
Koziol LF, Lutz JT (2013) From movement to thought: the development of executive function. Appl Neuropsychol Child 2(2):104–115
Kristjánsson K (2007) Aristotle, emotions and education. Ashgate Publishing, Aldershot
Kühn S, Gallinat J (2012) The neural correlates of subjective pleasantness. NeuroImage 61(1):289–294
Lindquist KA, Barrett LF (2012) A functional architecture of the human brain: emerging insights from the science of emotion. Trends Cogn Sci 16(11):533–540
Lindquist KA, Satpute AB, Wager TD, Weber J, Barrett LF (2016) The brain basis of positive and negative affect: evidence from a meta-analysis of the human neuroimaging literature. Cereb Cortex 26(5):1910–1922
Manto M, Marien P (2015) Schmahmann's syndrome - identification of the third cornerstone of clinical ataxiology. Cerebellum Ataxias 2:2
Marek S, Siegel JS, Gordon EM, Raut RV, Gratton C et al (2018) Spatial and temporal organization of the individual human cerebellum. Neuron 100(4):977–993
Molinari M, Leggio MG, De Martin M, Cerasa A, Thaut M (2003) Neurobiology of rhythmic motor entrainment. Ann N Y Acad Sci 999:313–321
Morawetz C, Alexandrowicz RW, Heekeren HR (2017) Successful emotion regulation is predicted by amygdala activity and aspects of personality: a latent variable approach. Emotion 17(3):421–441
Mothersill O, Donohoe G (2016) Neural effects of social environmental stress - an activation likelihood estimation meta-analysis. Psychol Med 46(10):2015–2023
Paquette S, Fujii S, Li HC, Schlaug G (2017) The cerebellum's contribution to beat interval discrimination. NeuroImage 163:177–182
Parsons LM (2001) Exploring the functional neuroanatomy of music performance, perception, and comprehension. Ann N Y Acad Sci 930:211–231
Petrosini L, Cutuli D, De Bartolo P, Laricchiuta D (2015) The creative cerebellum: insight from animal and human studies. In: Animal creativity and innovation. Academic Press, San Diego, CA, pp 189–212
Pessoa L (2013) The cognitive-emotional brain – from interactions to integrations. Massachusetts Institute of Technology Press
Raz G, Touroutoglou A, Wilson-Mendenhall C, Gilam G, Lin T et al (2016) Functional connectivity dynamics during film viewing reveal common networks for different emotional experiences. Cogn Affect Behav Neurosci 16(4):709–723
Riedel MC, Yanes JA, Ray KL, Eickhoff SB, Fox PT et al (2018) Dissociable meta-analytic brain networks contribute to coordinated emotional processing. Hum Brain Mapp 39(6):2514–2531
Ruffieux N, Colombo F, Gentaz E, Annoni JM, Chouiter L (2017) Successful neuropsychological rehabilitation in a patient with cerebellar cognitive affective syndrome. Appl Neuropsychol Child 6:180–188
Sabatinelli D, Keil A, Frank DW, Lang PI (2013) Emotional perception: correspondence of early and late event-related potentials with cortical and subcortical functional MRI. Biol Psychol 92:513–519
Scarantino A, de Sousa R (2021) Emotion. In: Zalta EN (ed) The Stanford encyclopedia of philosophy (summer 2021). Metaphysics Research Lab, Stanford University, Stanford
Schmahmann JD (2021) Emotional disorders and the cerebellum: neurobiological substrates, neuropsychiatry, and therapeutic implications. Handb Clin Neurol 183:109–154
Schmahmann JD, Sherman JC (1998) The cerebellar cognitive affective syndrome. Brain 121(4):561–579
Schmahmann JD, Guell X, Stoodley CJ, Halko MA (2019) The theory and neuroscience of cerebellar cognition. Ann Rev Neurosci 42:337–364
Schraa-Tam CKL, Rietdijk WJR, Verbeke WJMI, Dietvorst RC, van den Berg WE et al (2012) fMRI activities in the emotional cerebellum: a preference for negative stimuli and goal-directed behavior. Cerebellum 11(1):233–245
Schutter DJLG, van Honk J (2009) The cerebellum in emotion regulation: a repetitive transcranial magentic stimulation study. Cerebellum 8:28–34
Schutter DJLG (2016) A cerebellar framework for predictive coding and homeostatic regulation in depressive disorder. Cerebellum 15(1):30–33
Schutter DJLG (2020) The cerebellum in emotions and psychopathology. Taylor & Francis, London
Steinhoff N, Heine AM, Vogl J, Weiss K, Aschraf A, Hajek P, Schnider P, Tucek G (2015) A pilot study into the effects of music therapy on different areas of the brain of individuals with unresponsive wakefulness syndrome. Front Neurosci 9:291
Stoodley CJ, Schmahmann JD (2009) Functional topography in the human cerebellum: a meta-analysis of neuroimaging studies. NeuroImage 44(2):489–501
Styliadis C, Ioannides AA, Bamidis PD, Papadelis C (2015) Distinct cerebellar lobules process arousal, valence and their interaction in parallel following a temporal hierarchy. NeuroImage 110:149–161
Van Overwalle F, Baetens K, Mariën P, Vandekerckhove M (2014) Social cognition and the cerebellum: a meta-analysis of over 350 fMRI studies. NeuroImage 86:554–572
Van Overwalle F, D'aes T, Mariën P (2015) Social cognition and the cerebellum: a meta-analytic connectivity analysis. Hum Brain Mapp 36(12):5137–5514
Van Overwalle F, Manto M, Cattaneo Z, Clausi S, Ferrari C (2020) Consensus paper: cerebellum and social cognition. Cerebellum 19(6):833–868
Vartanian O, Kaufman JC (2013) Psychological and neural responses to art embody viewer and artwork histories. Behav Brain Sci 36(2):161–162
Wolf U, Rapoport MJ, Schweizer TA (2009) Evaluating the affective component of the cerebellar cognitive affective syndrome. J Neuropsychiatry Clin Neurosci 21(3):245–253
Zeki S, Romaya JP, Benincasa DM, Atiyah MF (2014) The experience of mathematical beauty and its neural correlates. Frontiers Human Neurosci 8:68
Zhou M, Hu X, Lu L, Zhang L et al (2017) Intrinsic cerebral activity at resting state in adults with major depressive disorder: a meta-analysis. Prog Neuropsychopharm Biol Psychiatry 75:157–164
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2022 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Adamaszek, M., Manto, M., Schutter, D.J.L.G. (2022). Current and Future Perspectives of the Cerebellum in Affective Neuroscience. In: Adamaszek, M., Manto, M., Schutter, D.J.L.G. (eds) The Emotional Cerebellum . Advances in Experimental Medicine and Biology, vol 1378. Springer, Cham. https://doi.org/10.1007/978-3-030-99550-8_19
Download citation
DOI: https://doi.org/10.1007/978-3-030-99550-8_19
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-99549-2
Online ISBN: 978-3-030-99550-8
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)