
Abstract
Subphylum Cyanidiophytina is composed of a single class (Cyanidiophyceae) and three genera have been traditionally recognized Cyanidium and Galdieria (Cyanidiales) and Cyanidioschyzon (Cyanidioschyzogonales); a new genus (Cyanidiococcus) has been recently described in the order Cyanidioschyzogonales. All species are exclusive to inland habitats, occurring mostly in thermal waters, as thermoacidophiles, and also as mesophiles (aerophytic or endolithic). Subphylum Proteorhodophytina is represented in freshwaters by all four classes (Compsopogonophyceae, Porphyrideophyceae, Rhodellophyceae, and Stylonematophyceae). Class Compsopogonophyceae has three freshwater members in the single order Compsopogonales: genera Boldia (family Boldiaceae), Compsopogon and Pulvinaster (family Compsopogonaceae); all are monospecific. Class Stylonematophyceae is represented by five genera, four in the order Stylonematales, Chroodactylon (one species), Chroothece (four species), Kyliniella and Rhodospora (one species), and one in the order Rufusiales, the monospecific genus Rufusia. Class Porphyridiophyceae has freshwater members in the genera Flintiella (one species) and Porphyridium (three species). Class Rhodellophyceae has the monospecific genus Glaucosphaera.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Cyanidiophyceae
- Compsopogonophyceae
- Freshwater
- Porphyrideophyceae
- Rhodellophyceae
- Rhodophyta
- Stylonematophyceae
- Taxonomy
Subphylum: Cyanidiophytina; HS Yoon, KM Müller, RG Sheath, FD Ott, and D Bhattacharya, J Phycol 42:489
The characteristics of the subphylum are as follows (adapted from Nakayama and Yokoyama 2017): unicellular, coccoid; cells covered by a distinct wall or naked, with a single parietal blue-green chloroplast without a pyrenoid, with peripheral thylakoids; phycobilisomes with allophycocyanin, c-phycocyanin; with β-carotene and zeaxanthin; mitochondrion spherical or reticulate, with flat cristae. Asexual reproduction by binary fission or endospores; sexual reproduction unknown. Most species inhabit acidic and high temperature conditions in hot springs or acidic sulfur fumes; some are mesophilic (aerophytic or endolithic); some are facultative heterotrophs (e.g., Galdieria).
A multigene phylogeny showed the Cyanidiophyceae to represent the most ancient split within the Rhodophyta that was estimated to have occurred ca. 1.3 million years ago (Yoon et al. 2004). The class Cyanidiophyceae had been composed of a single order Cyanidiales, but Ott (2009) proposed a new order (Cyanidioschyzogonales) to accommodate the genus Cyanidioschyzon. Molecular data suggest potential undescribed lineages in the class (Ciniglia et al. 2004; Reeb and Bhattacharya 2010; Hsieh et al. 2015).
There are discrete morphological characteristics to distinguish the Cyanidiales genera, as well as ecophysiological, cytomorphological, and biochemical differences (Merola et al. 1981; Albertano et al. 2000; Reeb and Bhattacharya 2010; Liu et al. 2020). Cyanidioschyzon has ovoidal to clavate shape cells and reproduces by binary fission, whereas in Cyanidium and Galdieria cells are mostly spherical and differ in average cell size and number of endospores produced per sporangium (Reeb and Bhattacharya 2010). A fourth genus (Cyanidiococcus) was recently described based on genomic, morphological, and physiological traits (Liu et al. 2020). The four genera also differ in other cytomorphological characteristics: chloroplast and mitochondrion shape, presence/absence of vacuoles, and a rigid cell wall (Albertano et al. 2000; Reeb and Bhattacharya 2010; Liu et al. 2020).
Phylogenetic analyses support the division of the Cyanidiophyceae into four distinct lineages (Ciniglia et al. 2004; Yoon et al. 2004, 2006a; Reeb and Bhattacharya 2010; Fig. 2.1). The first is the Galdieria lineage (excluding G. maxima) that is divided into two clades: Galdieria A (G. sulphuraria, G. daedala, and G. partita) and Galdieria B (G. phlegrea). Galdieria A includes strains from worldwide locations and the order of branching within this clade follows a clear geographical pattern. Galdieria B contains taxa from dry and endolithic habitats exclusively from Italy. These two clades do not have distinguishing morphological features but differ with respect to ecophysiology (growth and photosynthetic rates). The second major lineage is represented by two clades: one containing mesophilic Cyanidium spp . and another with three minor clades with extremophilic representatives (C. caldarium , Cyanidioschyzon, and G. maxima). Only one lineage (the mesophilic Cyanidium spp.) has a defined geographic distribution, inhabiting nonacidic and nonthermal caves in Italy. The other two clades do not show a pattern consistent with the geographical distribution.

Phylogenetic tree of the main lineages of the class Cyanidiophyceae based on rbcL sequences showing the four major lineages (modified from Reeb and Bhattacharya 2010). The thick branches denote well-supported monophyletic groups
Based on genomic, morphological, and physiological traits, a new genus and species (Cyanidiococcus yangmingshanensis) was proposed by Liu et al. (2020) and should be applied to several strains that were misidentified as G. maxima. The taxonomic scheme with two orders and three families was proven to be consistent with the new data: the Cyanidiales with Cyanidiaceae (Cyanidium Geitler) and Galdieriaceae (Galdieria Merola) and the Cyanidioschyzogonales with Cyanidioschyzonaceae (Cyanidioschyzon De Luca, Taddei and L. Varano and Cyanidiococcus S-L Liu, Y-R Chiang, HS Yoon and H-Y Fu).
Key to the genera of Cyanidiophyceae
1a | Reproduction by binary fission, cells clavate, or ovoidal with one elongate chloroplast at one side of the cell | Cyanidioschyzon |
1b | Reproduction by endospores, cells mostly spherical with chloroplast occupying most of the cell | 2 |
2a | Cells with vacuoles and a multi-lobed chloroplast | Galdieria |
2b | Cells without vacuoles and spherical or variable in shape chloroplast | 3 |
3a | Chloroplasts spherical, endospores produced in groups of 4 | Cyanidium |
3b | Chloroplasts polymorphic, endospores produced in groups of 2–4 | Cyanidiococcus |
Order Cyanidiales TA Christensen in Böcher and M Lange, Botanik, Bind II, Systematisk botanik, Number 2, Alger (1962) emend Ott, Handbook of the taxonomic names of non-marine Rhodophycophyta:588 (2009)
-
Description: cells without vacuoles and chloroplasts parietal, not lobed; reproduction by two to many endospores. Two families have been established in the order: Cyanidiaceae and Galdieriaceae. Family Cyanidiaceae Geitler has only one genus and species that are described below. The genus has been widely accepted to have a single species.
-
Cyanidium caldarium Geitler, Arch Hydrobiol Suppl 14:624 (1934) emend A Merola, R Castaldo, P De Luca, A Gambardella, A Musacchio and R Taddei, Giorn Bot Ital 115:191 (1981) (Fig. 2.2a, f)
-
Type: NAP, P De Luca (Neotype); corresponds to strain 182 of the Algal Collection University Federico II (ACUF), Naples, Italy.
-
Type locality: Indonesia, Central Java, Mount Lawu, 7.627500° S, 111.194167° E (estimated).
-
Description: cells spherical or rounded-pyramidal, rarely united in a common mucilaginous matrix, containing one large parietal, plate-like chloroplast, 2–6 μm in diameter; endospores formed in groups of 4, tetrahedrally divided, 1–3 μm in diameter.
-
Remarks: Ott (2009) listed seven species of Cyanidium but most of them had been previously transferred to Galdieria (Albertano et al. 2000).
-
Representative sequences in GenBank: AY541298 (rbcL).
-
Habitat: the species is most often reported in thermal waters with temperatures of 32–56 °C and low pH (0.5–4.5) and less often in caves with lower temperatures (15–25 °C) and circumneutral pH 7.0–7.2 (Ciniglia et al. 2004; Pinto et al. 2010).
-
Distribution: Asia: Indonesia, Japan; Europe: Italy, Spain (Fig. 2.3).
-
Key references: Geitler (1934), Merola et al. (1981), Albertano et al. (2000).
Family Galdieriaceae Merola, Castaldo, De Luca, Gambardella, A Musacchio and Taddei, Giorn Bot Ital 115:193 (1981)
-
Description: cells with multi-lobed chloroplasts, more than one mitochondrion and large vacuoles. Although the genus Galdieria Merola has recently been treated as having more than one species, only one species has been widely accepted in the genus.
Galdieria sulphuraria (Galdieri) Merola, Giorn Bot Ital 115:193 (1981) (Fig. 2.2b, c, g)
-
Basionym: Pleurococcus sulphurarius Galdieri , Atti Reale Accad Sci Fis Mat Napoli, ser. 3 5:162 (1899).
-
Type: NAP, P De Luca, and R Taddei (Neotype); corresponds to strain 002 of the Algal Collection University Federico II (ACUF), Naples, Italy.
-
Type locality: Italy, Naples, Campi Flegrei, Pisciarelli Sulphatare, 40.828244° N, 14.135010° E.
-
Description: cells spherical, sometimes united in a common mucilaginous matrix, containing one parietal, multi-lobed chloroplast, 3–11 μm in diameter; each cell has one to several vacuoles; endospores formed in groups of 2–32.
-
Representative sequences in GenBank: AY541311 (rbcL).
-
Habitat: this alga has been reported on humic soils, pools, or streams at very low pH (0–4.5) and variable temperatures (18–55 °C) (Ciniglia et al. 2004; Pinto et al. 2010; Ciniglia et al. 2014).
-
Distribution: Asia: Indonesia; Europe: Iceland, Italy, Portugal (Azores), Spain; North America: Guatemala, Mexico, the USA (Fig. 2.3).
-
Key references: Merola et al. (1981), Albertano et al. (2000).
-
Remarks: although several species have been accepted in the genus, most are of uncertain taxonomic status and were transferred from the genus Cyanidium (Albertano et al. 2000), whereas some are recognized only from molecular data (e.g., G. phlegrea, Pinto et al. 2010). We followed Liu et al. (2020) who stated that G. maxima, G. partita, and G. daedala should be treated as invalid names since only cultures of these three species were designated as types. They should be regarded as practical but invalid descriptions. The genus warrants a full revision which is out of scope of this book. Thus, only the type species is treated here.
Order Cyanidioschyzonales FD Ott, Handbook of the taxonomic names of non-marine Rhodophycophyta:577 (2009)
-
Description: cells clavate, with nucleus, mitochondrion, and vacuoles positioned at one end; chloroplasts ovoidal or clavate; spores not formed and vegetative reproduction by the formation of two to four daughter cells.
-
Family Cyanidioschyzonaceae FD Ott, Handbook of the taxonomic names of non-marine Rhodophycophyta:578 (2009).
-
The family had initially only the genus Cyanidioschyzon but the recently described
-
Cyanidiococcus is shown to have a high similarity of genome structure and closely related phylogenetically justifying its classification in the family. Thus, the family description has to be expanded to include the characteristics of Cyanidiococcus. Three species of Cyanidioschyzon are known but only one was formally described (Ott 2009), and Cyanidiococcus is monospecific (Liu et al. 2020).
Cyanidiococcus yangmingshanensis S-L Liu, Y-R Chiang, HS Yoon, and H-Y Fu, J Phycol 56:1435 (2020) (Fig. 2.2d, h)
-
Type: HAST 144519, P-C Chen, 20.i.2015 (Holotype); HAST 144520–144522, TUNG THAL066-001-003 (Isotypes); corresponds to strain THAL066 of the Culture Collection at the Tunghai University, Taichung, Taiwan.
-
Type locality: Taiwan, Yangmingshan National Park, GengZiPing, 25.188333° N, 121.614722° E.
-
Description: cells sub-spherical or short rod-shaped, sometimes united in a common mucilaginous matrix, containing one parietal, polymorphic chloroplast, 1.2–3.8 μm in diameter; reproduction by 2–4 endospores.
-
Representative sequences in GenBank: MN431657 (chloroplast genome).
-
Habitat: the species can inhabit acidic geothermal microhabitats (pH 0.4–5) including pools, hot springs, streams, sulfur fumes, and endoliths.
-
Distribution: Asia: Japan, Taiwan (Fig. 2.3).
-
Key references: Liu et al. (2020).
Cyanidioschyzon merolae De Luca, Taddei and Varano, Webbia 33:43 (1978) (Fig. 2.2e, i)
-
Type: NAP , P De Luca (Holotype); corresponds to strain 199 of the Algal Collection University Federico II (ACUF), Naples, Italy.
-
Type locality: Italy, Naples, Campi Flegrei, 40.827000° N, 14.139000° E.
-
Description: cells clavate or ovoidal, with one elongate, polymorphic chloroplast, at one side of the cell, 3–4 μm in length, 1–2 μm in diameter; reproduction by binary fission that occurs longitudinally forming a V-shaped dividing cell.
-
Representative sequences in GenBank: AY541296 (rbcL).
-
Habitat: this species has been reported from thermal aquatic habitats in very acidic waters (pH 0–3) and usually high temperatures (≥32 °C and up to 55 °C) (Ciniglia et al. 2004; Pinto et al. 2010).
-
Distribution: Asia: Indonesia; Europe: Italy; North America: the USA (Fig. 2.3).
-
Key references: De Lucca et al. (1978), Albertano et al. (2000)

(a) Cyanidium caldarium : (a) spherical cells with endospores (arrows); (b, c) Galdieria sulphuraria : (b) spherical cells with vacuoles (arrows) and multi-lobed chloroplasts (arrowheads); (c) spherical cells with vacuole (arrow) and endospores (arrowhead); (d) Cyanidiococcus yangmingshanensis : spherical cells; (e) Cyanidioschyzon merolae: clavate or ovoidal cells; (f–i) schematic drawing of the main ultrastructural characteristics of the cell: ch chloroplast, m mitochondria, n nucleus, v vacuole; (f) Cyanidium caldarium ; (g) Galdieria sulphuraria; (h) Cyanidiococcus yangmingshanensis; (i) Cyanidioschyzon merolae. Scale bars: (a–e) = 5 μm; (f–i) = 0.5 μm (Image authors: Fig. (a) M Aboal; Fig. (b) K Mozaffari; Fig. (c) T Leya for Fraunhofer IZI-BB (Strain CCCryo 127-00); Fig. (d) H-Y Fu; Fig. (e) NBRP-Algae (National BioResource Project, Japan, Strain NIES-1332); Fig. (f, g, i) modified from Merola et al. (1981); Fig. (h) modified from Liu et al. (2020))

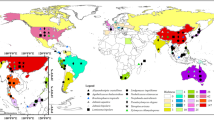
World map showing the distribution of Cyanidiococcus yangmingshanensis , Cyanidium caldarium , Cyanidioschyzon merolae, and Galdieria sulphuraria. Open symbols represent the type localities
Subphylum: Proteorhodophytina; SA Muñoz-Gómez, FG Mejía-Franco, K Durnin, M Colp, CJ Grisdale, JM Archibald, and CH Slamovits, Curr Biol 27:1680 (2017)
The characteristics of the subphylum are as follows (adapted from Muñoz-Gómez et al. 2017): red algae with unicellular, pseudofilamentous or filamentous thalli inhabiting mesophilic freshwater and marine environments. Members of this group do not develop macroscopic, pseudoparenchymatous, or parenchymatous seaweed-type thalli as those of the Eurhodophytina; variable plastid morphologies and organellar associations; plastid genomes with a large number of introns and a quadripartite organization in which the rRNA operon-containing inverted repeats are divergent relative to the small single copy region.
This group encompasses four classes (Compsopogonophyceae, Porphyridiophyceae, Rhodellophyceae, and Stylonematophyceae) with the largest plastid genomes sequenced (up to 1,127,474 bp). Within the subphylum two well-supported sister groups were detected, one with the classes Compsopogonophyceae and Porphyridiophyceae and another with the classes Stylonematophyceae and Rhodellophyceae. All four classes occur in freshwater.
Class: Compsopogonophyceae; GW Saunders and Hommersand, Am J Bot 91:1503 (2004)
Thalli filamentous , crustose, saccate or foliose, macroscopic or microscopic; sometimes with simple pit plugs (only plug core) between cells (Compsopogon and Rhodochaete); cells covered by cell wall; basically uninucleate. Chloroplasts variable in shape and number, blue-green, violet, or reddish, with peripheral thylakoid; phycobilisomes with allophycocyanin, phycocyanin, and phycoerythrin; with β-carotene, zeaxanthin, and antheraxanthin. Mitochondrion with flat cristae. Asexual reproduction by monospores generated by the oblique asymmetric division of vegetative cells. Sexual reproduction by oogamy (spermatia and carpogonia) and haplodiplontic life cycle reported in some species. Marine or freshwater. Description adapted from Nakayama and Yokoyama (2017). Freshwater members only in the order Compsopogonales.
Order Compsopogonales Skuja, Acta Horti Bot Univ Latv 11/12:34 (1939)
-
Thalli mostly macroscopic, filamentous, or saccate, with prostrate filamentous holdfast. Chloroplasts parietal, discoid or ribbon-shaped, lobed, without pyrenoid. Sexual reproduction unknown. Asexual reproduction by monosporangia. Mostly in freshwater streams or lakes, rarely in brackish water. Description adapted from Nakayama and Yokoyama (2017). Members of Compsopogonales are currently classified into two families: Boldiaceae and Compsopogonaceae.
Family Boldiaceae Herndon, Am J Bot 51:575 (1964)
-
Thalli monostromatic saccate to tubular, reddish, no pit plug.
-
Genus Boldia Herndon, Am J Bot 51:575 (1964).
-
The genus is monospecific and the characters are described below.
Boldia erythrosiphon Herndon, Am J Bot 51:576 (1964) (Fig. 2.4a–e)
-
Synonyms: Boldia angustata Deason and Nichols, J Phycol 6:40 (1970)
-
Type: US, WR Herndon, 05.vii. 1958, (Holotype, not found).
-
Type locality: USA: Virginia, Giles County, near Pembroke, Big Walker Creek, 37.315520° N, 80.643938° W (estimated).
-
Description: Thalli hollow , monostromatic sac attached to surfaces by a disc of cells, 1–20(–75) cm in height and 0.1–2 cm in diameter. Vegetative cells primarily circular, sometimes polygonal to rectangular in surface view, 5–20(–45) μm, depending on timing of cell division; chloroplasts discoidal or ribbon-shaped; larger, granular ovoidal cells scattered throughout the thallus, 25–40 μm in diameter; monosporangia derived from intercalary filaments, forming a network of cells, polygonal to ovoid, deeply pigmented, 5–9 μm in diameter; monospores germinate to form a disc of cells becoming a cushion-like mound that perennially produces the saccate thalli. No sexual reproduction known.
-
Representative sequences in GenBank: AF087121, AF087122 (rbcL).
-
Habitat: in the southern part of the distributional range, this alga often occurs on Pleuroceridae snails as well as attached to rock surfaces, but only epilithic in the northern part of the range. Stream conditions for this alga have been reported to be 6.1–8.5 pH, 3–71 cm s−1 current velocity, low to moderate specific conductance (18–290 μS cm−1) and a wide range in water temperature 12–25 °C (Sheath and Vis 2015 and references therein). It has often been noted to co-occur with other freshwater red algal taxa of the order Batrachospermales.
-
Distribution: North America: southeastern US and Canada in Ontario and Quebec (Fig. 2.5).
-
Key references: Herndon (1964), Howard and Parker (1980), Sheath and Hymes (1980), Sheath et al. (1989), Sheath and Vis (2015).
Family Compsopogonaceae F Schmitz, Natürl Pflanzenfam:318 (1896)
-
Branched filaments, blue-green to violet, sometimes with pit plugs. Two monospecific genera are currently recognized in the family (Compsopogon Montagne and Pulvinaster JA West, Zuccarello and JL Scott) and described below.
Compsopogon caeruleus (Balbis ex C Agardh) Montagne, Flore d’Algérie, Cryptogamie:154 (1846) (Fig. 2.6a–g)
-
Basionym: Conferva caerulea Balbis ex C Agardh, Systema Algarum:122 (1824).
-
Synonyms: listed by Necchi and Dip (1992), Vis et al. (1992), Rintoul et al. (1999), Sheath and Sherwood (2011), and Necchi et al. (2013) including all other species and infraspecific taxa described in the genera Compsopogon and Compsopogonopsis.
-
Type: LD, Bertero, 23.vii.1824, in Herbarium AGARDH Nr. 12400 (Holotype, not found); PC0047796, PC0047797 (Isotypes ).
-
Type locality: Puerto Rico, locality not specified.
-
Description: Thalli heterotrichous, attached by discoid holdfasts, uniseriate rhizoids or knot-like structures; basal system with 1–15 erect branches; spine-like branchlets occasionally produced, irregularly distributed in basal or middle portions. Main axes alternately, rarely unilaterally branched, at variable angles, 30–60(–90o), 100–2000(–3000) μm in diameter. Axial cells barrel-shaped or sub-spherical 45–450 μm in diameter, 50–400 μm in length; axial cells may break down with age leaving the thallus hollow or saccate-like. Cortex consisting of 1–4(–5) cell layers, 15–200(–380) μm in thick; cortication formed by vertical and regular or oblique and irregular divisions of axial cells forming or not rhizoidal filaments; outer cortical cells polygonal, tri- to hexangular or sub-circular in surface view, (10–)15–55(–75) μm in length, (5–)15–30(–40) μm in diameter. Uniseriate branches with disc-shaped or cubic cells, (5–)10–40(–55) μm in diameter, (4–)7–20(–30) μm in length; terminal cells dome-shaped or conical. Monosporangia variable in shape, elliptical, ovoidal, sub-circular, tri- to pentangular or fusiform in surface view, (10–)15–25(32.5) μm in length, (7.5–)10–20 μm in diameter. Microsporangia occur occasionally, arranged in sori, variable in shape, tri- to pentangular, ovoid or fusiform, 5–12.5 μm in length, 2.5–10 μm in diameter; microsporangial sori irregular in shape, circular, ovoid, square or polygonal in surface view, with (4–)8–35 microsporangia, (10–)40–100(–180) μm in diameter.
-
Representative sequences in GenBank: JX028177, JX028179, JX028186 (COI-5P); JX028155, JX028161, JX028167 (rbcL).
-
Habitat: this species has a wide tolerance of environmental variables ranging from clean to moderately polluted waters, slightly acidic to alkaline (pH 6.4–8.3), low ion concentrations up to brackish waters (18–1766 μS cm−1) and mid to high temperatures (most records ≥20 °C) (Necchi et al. 2013 and references therein). This wide ecological niche most likely contributes the ubiquitous distribution of Compsopogon in tropical–subtropical regions. It is a warm water species and the few records in temperate locations were at outflows of industries with warm water discharges.
-
Distribution: worldwide in tropical or sub-tropical regions , rarely in temperate areas (Fig. 2.5).
-
Key references: Necchi and Dip (1992), Vis et al. (1992), Rintoul et al. (1999), Eloranta et al. (2011), Necchi et al. (2013).
-
Remarks: Necchi et al. (2013) found little genetic variation based on a global sampling (25 specimens from locations in North America, South America, Europe, Asia, Australasia, and Oceania) for two markers (rbcL and COI-5P). They concluded from molecular and morphological evidence that there is only a single monospecific genus worldwide; the oldest specific epithet is C. caeruleus and all other species described in the genera Compsopogon and Compsopogonopsis to be treated as synonyms. This taxon is commonly referred to as “staghorn” algae in the aquarium trade.
Pulvinaster venetus JA West, Zuccarello and JL Scott, Phycologia 46:478 (2007b) (Fig. 2.6h, i)
-
Type: NSW 732559, J West, 14.vi.2005 (Holotype ).
-
Type locality: Vanuatu, Efate, Eton River, near a bridge 50 m from the river mouth, 17.733322° S, 168.550023° E.
-
Description: thallus consisting of a prostrate cushion up to 500 μm in diameter, of uniseriate branched filaments, formed by quadrate to ovate cells (9–14 μm in diameter), uninucleate, with a bluish-green parietal lobed or spiral plastid lacking a pyrenoid; older filaments in culture have a thick matrix and are bi- or multiseriate, with up to 25 μm in diameter; sometimes forming unattached branched filaments with elongate cells, 30–55 in length, 8–10 μm in diameter. Reproduction by monosporangia successively formed from lateral branch cells or by gelatinization of cell walls of a thallus sector; monospores spherical, 6–10 μm in diameter, showing a gliding movement. Golgi associated with endoplasmic reticulum; plastid with peripheral thylakoid; no pit connections between derivative cells; low molecular weight carbohydrate floridoside.
-
Representative sequences in GenBank: EF079964 (SSU rDNA); EF079965 (psbA).
-
Distribution: Pacific Islands: Vanuatu (Fig 2.5).
-
Habitat: Epiphytic on red macroalgae (Bostrychia and Caloglossa) in a shallow, slow-moving, freshwater stream (±2 psi).
-
Remarks: this taxon was invalidly published as Pulvinus venetus (West et al. 2007a) and later validated (West et al. 2007b) by referring to the Latin description and figures.

(a–e) Boldia erythrosiphon : (a) saccate translucent thalli (arrows) in clusters on rock surface; (b) numerous thalli (arrow) growing attached to one end of a snail; (c) collapsed tube-like thalli with dark red or light pink appearance; (d) thallus surface with enlarged clear cell (arrow) and isodiametric (arrowhead) cells having ribbon-shaped chloroplasts; (e) monosporangia (arrows) scattered in chain on the thallus surface. Scale bars: (a–c) = 1 cm; (d, e) 20 μm (Image author: Fig. (a, b) SA Krueger-Hadfield)

World map showing the distribution of Boldia erythrosiphon, Compsopogon caeruleus, and Pulvinaster venetus. Open symbols represent the type localities

(a–g) Compsopogon caeruleus : (a) general view with corticated (arrows) and uniseriate (arrowhead) filaments; (b) uniseriate filament with a young branch (arrow); (c) filament in early stage of cortication with vertical and regular divisions (arrows); (d) filament in early stage of cortication with oblique and irregular divisions forming rhizoidal cells (arrows); (e) corticated filament with axial cells (arrowheads) and one layer of cortical cells (arrows); (f) surface view of a corticated filament with monosporangia (arrowheads) and microsporangia (arrows); (g) surface view of a corticated filament with monosporangia (arrowheads) and cortical cells (arrows); (h, i) Pulvinaster venetus: (h) general view with uniseriate (arrows) and multiseriate filaments. (i) apices (arrows) of uniseriate filaments. Scale bars: (a) = 500 μm; (c, f–h) 50 μm; (b, d, e, i) = 25 μm (Image author: Fig. (h, i) JA West)
Class: Porphyridiophyceae; M Shameel, Pakistan J Mar Biol:240 (2001)
Thalli unicellular, sometimes united in a common mucilage. Cells usually exhibiting a gliding movement; uninucleate. Chloroplasts blue-green, green or reddish, without peripheral thylakoid, single, axial, stellate, without or with a naked pyrenoid; phycobilisomes with allophycocyanin, phycocyanin, and usually phycoerythrin; with β-carotene and zeaxanthin. Mitochondrion with tubular cristae. Asexual reproduction by binary fission. Sexual reproduction unknown. Marine, freshwater, or moist soils. Description adapted from Nakayama and Yokoyama (2017). Members of Porphyridiophyceae are currently classified in a single order (Porphyridiales Kylin) and family (Porphyridiaceae Kylin) with freshwater members in the genera Flintiella and Porphyridium.
Genus Flintiella FD Ott in Bourrelly, Algues D’Eau Douce 3:194 (1970)
-
The genus is monospecific and the characters are described below.
-
Flintiella sanguinaria Ott in Bourrelly, Algues D’Eau Douce 3:194 (1970) (Fig. 2.7a, b)
-
Type: FD Ott, xi.1964 (Holotype, not found).
-
Type locality: USA, Texas, Travis County, Austin, Barton Springs, 30.264546° N, 97.769241° W.
-
Description: unicellular, spherical, slightly ovoidal prior to division, surrounded by a non-laminated mucilaginous sheath, 6–20(–45) μm in diameter, with a central nucleus, a single parietal, irregularly and deeply lobed reddish chloroplast without pyrenoid.
-
Diagnostic characters: Flintiella can be differentiated from Porphyridium by the shape of chloroplast (parietal and lobed versus axial and stellate) and the absence of pyrenoid.
-
Representative sequences in GenBank: AY119774 (rbcL).
-
Distribution: North America: the USA (Fig 2.8).
-
Habitat: this alga was collected from the mouth of a spring where it appeared as a pink-colored growth on pebbles in rapidly flowing water with temperature (23–24 oC), pH (6.9), high hardness (301 mg L−1 CaCO3).
Genus Porphyridium Nägeli, Neue Denksch Allg Schweiz Gesellsch Gesammten Naturwiss 10:71 (1849)
-
Description: unicellular, but often grouped into irregular colonies with an ill-defined mucilaginous matrix, spherical, ellipsoidal, or ovoidal, with a stellate chloroplast and prominent central pyrenoid. Reproduction by cell division; sexual reproduction unknown.
-
Diagnostic characters: Porphyridium is characterized by having stellate chloroplasts with a central embedded pyrenoid. Diagnostic characters to recognize species are essentially based on color, reflecting distinct proportions of phycobiliproteins (allophycocyanin, phycocyanin, and phycoerythrin).
-
Habitat: species are freshwater , marine or occur on moist soils.
-
Distribution: most records are from Europe with some sparse and fragmentary reports for other continents (Fig. 2.8). According to Nakayama and Yokoyama (2017), the distribution of red microalgae is poorly documented and the reported distribution is just the region studied by experts in many cases.

(a, b) Flintiella sanguinaria : (a) group of cells; (b) detail of cells showing nucleus (arrow) and lobed chloroplast (arrowheads); (c-d) Porphyridium aerugineum: (c) group of cells; (d) detail of cells; (e, f) Porphyridium purpureum: (e) group of cells; (f) detail of cells showing pyrenoids (arrows) and stellate chloroplasts (arrowheads); (g, h) Porphyridium sordidum: (g) colony with cells embedded in a thin mucilage (arrow); (h) detail of a cell showing pyrenoid (arrow) and stellate chloroplast (arrowhead); (i, j) Glaucosphaera vacuolata: (i) group of cells surrounded by a mucilage (viewed with China ink); (j) detail of a cell showing nucleus (arrow) and vacuoles (arrowheads); (k) Chroodactylon ornatum: (k) pseudofilament with branches (arrows) and cells with pyrenoids (arrowheads). Scale bars: (i, k = 20 μm; (a, c, e, g) 10 μm; (b, d, f) = 5 μm; (h, j) 2.5 μm (Image author: Fig. (a, b, e–f) T Darienko (SAG Culture Collection of Algae, Goettingen University, Germany; © SAG); Fig. (c) T Friedl (SAG Culture Collection of Algae, Goettingen University, Germany; © SAG); Fig. (d) A Kryvenda (SAG Culture Collection of Algae, Goettingen University, Germany; © SAG); Fig. (g) CCAP (Culture Collection of Algae and Protozoa, Scottish Marine Institute, Scotland); Fig. (h) CCMP (Culture Collection of Algae, Bigelow Laboratory for Ocean Sciences, USA); Fig. (i, j) JA West; Fig. (k, l) C Carter)

World map showing the distribution of Flintiella sanguinaria , Porphyridium aerugineum , P. purpureum, P. sordidum, and Glaucosphaera vacuolata. Open symbols represent the type localities
Phylogenetic relationships among species: analyses of published rbcL sequences (Fig. 2.9) shows two single specimen branches and one clade within the genus Porphyridium representing three species: P. aerugineum, P. purpureum, and P. sordidum. The intraspecific variation for P. purpureum is low (0.2%).
Key to the species of Porphyridium
1a | Cells reddish | P. purpureum |
1a | Cells greenish-blue or greenish | 2 |
2a | Cells greenish | P. sordidum |
2b | Cells blue-greenish | P. aerugineum |

Maximum Likelihood tree based on rbcL sequences showing the phylogenetic relationships among the three species of Porphyridium. Support values are maximum likelihood bootstrap and Bayesian posterior probabilities
Porphyridium aerugineum Geitler, Öst Bot Z 72:84 (1923) (Fig. 2.7c, d)
-
Type: not found .
-
Type locality: Austria, Vienna, Vienna University Botanic Garden, 48.191671° N, 16.383968° E (estimated).
-
Description: unicells spherical or ellipsoidal, blue-green, 4.4–8.5(–10.6) μm in diameter.
-
Representative sequences in GenBank: AY119775, X17597 (rbcL ).
-
Distribution: Europe: Austria, Germany, Great Britain, Latvia; South America: Brazil (Fig 2.8).
-
Habitat: on moist soils or epiphytic on macrophytes in ponds and rivers.
-
Key references: Geitler (1923), Ott (1972, 1987), Pekárková et al. (1989), Eloranta et al. (2011).
Porphyridium purpureum (Bory) KM Drew and R Ross, Taxon 14:98 (1965) (Fig. 2.7e, f)
-
Basionym: Phytoconis purpurea Bory, Mémoire sur les genres Conferva et Byssus:55 (1797).
-
Type: PC0112173, Bory Saint-Vincent (Holotype).
-
Type locality: France, locality not specified.
-
Description: unicells, spherical, blood-red to brownish red, 6.9–12(–15) μm in diameter.
-
Representative sequences in GenBank: DQ308439, MN539012 (rbcL).
-
Distribution: Asia: China, India, Japan; Australasia: Australia, New Zealand; Europe: Croatia, France, Germany, Great Britain, Ireland, Hungary, Latvia, Romania, Slovakia, Slovenia, Spain, Sweden; North America: Canada, the USA; Pacific Islands: Micronesia; South America: Argentina, Brazil, Chile (Fig. 2.8).
-
Habitat: on moist terrestrial areas (soil, walls), particularly with calcareous components, brickwork in shaded areas or intermittently submerged in nutrient-rich rivers.
-
Key references: Ott (1972, 1987), Eloranta et al. (2011), Medina-Cabrera et al. (2020).
Porphyridium sordidum Geitler, Arch Protistenk 76:603 (1932) Fig. 2.7g, h
-
Type: not found.
-
Type locality: Austria, Pinggau, Steiermark, 47.445564° N, 16.066464° E (estimated).
-
Description: cells spherical or ovoidal, greenish (gray-green, olive-green, or yellow-green), 5.5–9.5(–11.5) μm in diameter.
-
Representative sequences in GenBank: DQ308440 (rbcL).
-
Distribution: Europe: Austria; North America: the USA (Connecticut, Iowa, Kansas, Massachusetts, Missouri, Nebraska) (Fig 2.8).
-
Habitat: on moist terrestrial areas (soil, walls), particularly with calcareous components, brickwork in shaded areas or intermittently submerged in nutrient-rich rivers.
-
Key references: Geitler (1932), Ott (1987), Eloranta et al. (2011), Medina-Cabrera et al. (2020).
Doubtful Species: A fourth species of Porphyridium (P. griseum Geitler ) has been recognized in some general treatments (Kumano 2002; Eloranta et al. 2011). However, there is no molecular or morphological evidence to support the recognition of this species. It is described as having grayish cells, but this could be only a variation within the color spectrum of other species, particularly P. sordidum that could have gray-green cells. Thus, we decided to place it provisionally as a doubtful species until further information is provided to define its taxonomic status.
Class Rhodellophyceae
Thallus unicellular, sometimes united in a common mucilage. Cells covered by an indistinct cell wall with copious mucilage; uninucleate. Chloroplasts blue-green to reddish, with peripheral thylakoid, axial, stellate to highly lobed, usually with pyrenoid; phycobilisomes with allophycocyanin, phycocyanin and usually phycoerythrin; with β-carotene and zeaxanthin. Mitochondrion with flat cristae. Asexual reproduction by binary fission; sexual reproduction unknown. Most marine, rarely in freshwater. Members of Rhodellophyceae are currently classified into three orders with the only freshwater member in Glaucosphaerales, characterized by a highly lobed peripheral blue-green chloroplast containing no phycoerythrin and without pyrenoid. Description adapted from Nakayama and Yokoyama (2017).
Genus Glaucosphaera A Korshikov, Arch Protistenk 70:222 (1930)
-
The genus is monospecific and the characters are described below.
-
Glaucosphaera vacuolata A Korshikov, Arch Protistenk 70:222 (1930). Fig. 2.7i, j
-
Type: not found.
-
Type locality: Ukraine, Kharkiv, 49.993500° N, 36.230383° E (estimated).
-
Description: cells spherical, 18–25 μm in diameter with a central nucleus, a single highly lobed, blue-green chloroplast, and numerous small vesicles visible throughout the cell; reproduction by binary fission.
-
Representative sequences in GenBank: DQ308427 (rbcL).
-
Distribution: Europe: Ukraine; Asia: Japan; North America: the USA (Fig. 2.8).
-
Habitat: this species is a minor component of phytoplankton in ponds; most reports refer to material isolated in culture.
Class: Stylonematophyceae; HS Yoon, KM Müller, Sheath, FD Ott, and D Bhattacharya, J Phycol 42:490 (2006b)
Thalli pseudofilamentous (uniseriate, multiseriate, or tubular), but some are unicellular, mostly microscopic. Cells covered by cell wall and copious mucilage; uninucleate. Chloroplasts blue-green to reddish, with peripheral thylakoid, stellate or discoid, sometimes with pyrenoid; phycobilisomes with allophycocyanin, phycocyanin, and usually phycoerythrin; with β-carotene and zeaxanthin. Mitochondrion with flat cristae. Asexual reproduction by fragmentation, archeospores, endospores (autospores), or binary fission; sexual reproduction unknown for most members, but spermatia and carpogonia reported in Kylinella. Most marine but some in freshwater. Description adapted from Nakayama and Yokoyama (2017). Members of Stylonematophyceae are currently classified into two orders: Rufusiales and Stylonematales.
Order Stylonematales KM Drew, Bot Tidsskr 22:73 (1956)
-
Thalli pseudofilamentous or unicellular; cells with an axial stellate chloroplast containing pyrenoid or parietal chloroplast without pyrenoid. Description adapted from Nakayama and Yokoyama (2017). There is only one family (Stylonemataceae Drew) with the same characters of the order. Four genera occur in freshwater.
Key to the genera of Stylonematales
1a | Cells with several parietal, discoid chloroplasts, without pyrenoid | 2 |
1b | Cells with one axial, stellate chloroplast, with one pyrenoid | 3 |
2a | Unicells, sometimes forming colonies | Rhodospora |
2b | Unbranched pseudofilaments | Kyliniella |
3a | Cells arranged in long pseudofilaments | Chroodactylon |
3b | Cells solitary, less often forming short pseudofilaments | Chroothece |
Genus Chroodactylon Hansgirg, Ber Deutsch Bot Ges 3:14 (1885)
-
Type species: Chroodactylon wolleanum Hansgirg, Ber Deutsch Bot Ges 3:14 (1885).
-
Description: pseudofilaments with false branching or unbranched, encircled in a broad, mucilaginous sheath with an irregular and uniseriate arrangement; cells sub-spherical to ellipsoidal, containing one blue-green, stellate, axial chloroplast, with a prominent pyrenoid. Reproduction by monosporangia and fragmentation. Sexual reproduction unknown.
-
Diagnostic characters: this genus is differentiated from others based on pseudofilaments with false branching and a large blue-green, stellate chloroplast with prominent pyrenoid; however, see discussion of Chroothece as this genus may also produce short pseudofilaments.
-
Habitat: primarily epiphytic on macroalgae in marine, brackish and freshwaters.
-
Distribution: this genus has been reported from marine and brackish coastal waters of North and South America, Asia, and Europe; it is widespread in alkaline freshwaters on numerous continents (Fig. 2.10).
-
Remarks: the number of species that should be recognized in this genus is unclear. There are three validly published and currently taxonomically accepted species (C. depressum, C. ornatum, and C. wolleanum); however, most modern freshwater treatments (e.g., Eloranta et al. 2011, Sheath and Vis, 2015) only recognize one species, C. ornatum and this scheme will be followed here. More research on the morphology and DNA sequence data is needed for this genus.
Chroodactylon ornatum (C Agardh) Basson, Bot Mar 22:67 (1979) (Fig. 2.7k)
-
Type: LD A1863, C Agardh (Holotype).
-
Type locality: Sweden, near Stockholm, Lake Mälaren, Bridge near Transberg, 59.446679° N, 17.466347° E (estimated).
-
Description: pseudofilaments with 0–6 false branches, 0.024–10 mm in length; cells sub-spherical to ellipsoidal, 7.1–16.6 μm in length, 5.8–11.6 μm in diameter, containing one blue-green, stellate, axial chloroplast, with a prominent pyrenoid.
-
Representative sequences in GenBank: DQ308429, EF660259, EF7660260 (rbcL).
-
Distribution: Africa: Madagascar; Asia: China, Japan, Philippines; Australasia: Australia; Europe: Austria, Croatia, Denmark, Hungary, Lithuania, Netherlands, Norway, Poland, Slovak Republic, Spain, Sweden, Switzerland, UK; North America: Canada, Mexico, USA; South America: Argentina (Fig 2.10).
-
Habitat: growing as an epiphyte on aquatic plants and macroalgae, especially Cladophora ; typically, in alkaline waters with pH 7.5–8.5 and conductivity 170–540 μS cm−1. This taxon has been reported in large streams, but more often in lakes; in North America, it occurs as a common epiphyte of Cladophora in the Great Lakes.
-
Key references: Vis and Sheath (1993), Eloranta et al. (2011)
-
Remarks: DNA sequence data available in GenBank are from North America, Philippines and Madagascar and differ by 1.1–1.8% suggesting that more research may be warranted to circumscribe the species and its geographic range.
Genus Chroothece Hansgirg, Öst Bot Zeit 34:353 (1884)
-
Type species: Chroothece richteriana Hansgirg in Wittrock and Nordstedt, Bot. Not. 1884:128 (1884).
-
Description: cells solitary or joined pole to pole, less often forming short pseudofilaments, immersed in a firm and stratified mucilaginous matrix, the basal pole extending into a lamellated stalk; cells ellipsoidal to cylindrical, containing one blue-green, stellate, axial chloroplast, with a prominent pyrenoid. Reproduction by cell division and by fragmentation. Sexual reproduction unknown.
-
Diagnostic characters : Chroothece is morphologically similar to Chroodactylon and they are typically separated by cell arrangement (Eloranta et al. 2011; Necchi 2016): unicells or small colonies in Chroothece and pseudofilaments in Chroodactylon. However, short pseudofilaments, branched or unbranched, are often observed in Chroothece species in culture and sometimes in the field (Aboal et al. 2018). Thus, there is overlap but the pseudofilaments are typically shorter in Chroothece in comparison to Chroodactylon and the two genera are clearly distinct based on rbcL sequences. More research is needed to determine if there are other morphological characters to separate these genera.
-
Habitat: species are mostly aerophytic within permanent seeps over different types of rocks or less often aquatic. Chroothece mobilis is reported as both aerophytic on limestones with permanent seepage and low irradiance (Aboal et al. 2018) or aquatic at pH 7.2–7.8, temperature 13° C and conductivity 340–570 μS cm−1 (Blinn and Prescott 1976). Chroothece lobata is found aquatic in calcareous and slightly saline water: temperature 11.4–28.8°C, pH 8.0–8.9, conductivity 281–3100 μS cm−1 and C. richteriana at temperature 11.9–22.3°C, conductivity 2640–3100 μS cm−1, oxygen 8.3–10.4 mg L−1 (Aboal et al. 2014, 2018). Chroothece thermalis occurs in a thermal spring (temperature 38° C) on detrital rock above water level (Aboal et al. 2018).
-
Distribution: one species (C. mobilis) is relatively widespread, whereas C. richteriana is mostly reported from Europe and two species (C. lobata and C. thermalis) are endemic, restricted to the type localities in Spain (Fig. 2.10).
-
Phylogenetic relationships among species: analyses of rbcL sequences from data reported by Aboal et al. (2018) supports recognition of Chroothece as a distinct genus forming a clearly separated clade from Chroodactylon (Fig. 2.11). Chroothece has two high-supported major clades: one with two sequences of C. lobata from Spain and another with sequences of three species (C. mobilis, C. richteriana, and C. thermalis) from distinct regions of the world.
Key to the species of Chroothece
1a | Mean cell diameter ≤10 μm | C. richteriana |
1b | Mean cell diameter ≥10 μm | 2 |
2a | Mean cell diameter ≤14 μm, colonies lobed | C. lobata |
2b | Mean cell diameter ≥14 μm, colonies forming mats | 3 |
3a | Aquatic in thermal spring (38° C) | C. thermalis |
3b | Aerophytic on cave wall or aquatic in cool waters (13° C) | C. mobilis |

World map showing the distribution of Chroodactylon ornatum, Chroothece lobata, C. mobilis, C. richteriana, C. thermalis, Kyliniella latvica, Rhodospora sordida, and Rufusia pilicola. Open symbols represent the type localities

Maximum Likelihood tree based on rbcL sequences showing the phylogenetic relationships among the four species of Chroothece and Chroodactylon ornatum. Support values are maximum likelihood bootstrap and Bayesian posterior probabilities
Chroothece lobata Aboal, BA Whitton, IS Chapuis, P Sánchez, and Necchi, Eur J Phycol 53:194 (2018) (Fig. 2.12a, b)
-
Type : MUB-ALGAE-2076, I Chapuis, P Sánchez and M Aboal, 14.ii.2014 (Holotype); GDA-A-6332 (Isotype).
-
Type locality: Spain, Murcia, Umbria, River Chícamo, 38.265316° N, 0.991772° W.
-
Description: colonies hemispherical or lobate, blue-green to yellow-orange; pseudofilaments of 3–4 cells frequent, occasionally branched; cells ellipsoidal, 11–31.1 μm in length, 7–14(–17.4) μm in diameter; mucilaginous sheath 1.2–2.2 μm thick, smooth.
-
Representative sequences in GenBank: KY962002, KY962005 (rbcL).
-
Distribution: Europe: Spain (Fig 2.10).
-
Key references: Aboal et al. (2018).
Chroothece mobilis Pascher and J Petrová, Arch Protistenk 74:490 (1933) (Fig. 2.12c, d)
-
Type : not found.
-
Type locality: Czech Republic, Bohemia, Franzensbad, 50.120059° N, 12.349681° E (estimated).
-
Description: Colonies forming a mat, blue-green to yellow-orange; pseudofilaments frequent, up to 5 cells, with short branches; cells ellipsoidal, (23.5-)28.5–38.5(–45) μm in length, 15.4–22.5(–27) μm in diameter; mucilaginous sheath 1.6–3–6.6 μm thick, smooth.
-
Representative sequences in GenBank: DQ308430, KY962004 (rbcL).
-
Distribution: Asia: Bangladesh; Australasia: Australia; Europe: Czech Republic, Germany, Great Britain, Spain; North America: the USA; Pacific Islands: Guam (Fig. 2.10).
-
Key references: Pascher and Petrová (1933), Blinn and Prescott (1976), Eloranta et al. (2011), Aboal et al. (2018).
Chroothece richteriana Hansgirg, Öst Bot Zeit 34:353 (1884) (Fig. 2.12e, f)
-
Type: not found.
-
Type locality: Czech Republic, Bohemia, Auzitz near Kralup, 50.241645° N, 14.310645° E (estimated).
-
Description: Colonies forming a mat or hemispheric, blue-green when young and brownish, yellowish, or orange with age; pseudofilaments not observed; cells ellipsoidal, 15–33 μm in length, 5–10 μm in diameter; mucilaginous sheath 2–4 μm thick, layered.
-
Representative sequences in GenBank: KY962003 (rbcL).
-
Distribution: Europe: Czech Republic, Germany, Great Britain, Russia, Spain; Asia: India (Fig. 2.10).
-
Key references: Eloranta et al. (2011), Aboal et al. (2014, 2018).
Chroothece thermalis IS Chapuis, P Sánchez, Aboal, and Necchi, Eur J Phycol 53:195 (2018) (Fig. 2.12g)
-
Type: GDA 6334, I Chapuis, 26.x.2012 (Holotype); MUB-ALGAE 5835 (Isotype).
-
Type locality : Spain: Granada, Santa Fé, 37.157639° N, 3.751861° E.
-
Description: colonies forming a mat, blue-green or yellowish; pseudofilaments not observed; cells cylindrical, (15–)20–29 μm in length, (12–)14–24 μm in diameter; mucilaginous sheath 2.5–4 μm thick, slightly undulated; stalks striated and wavy, transparent.
-
Representative sequences in GenBank: KY962006 (rbcL).
-
Distribution: Europe: Spain (Fig 2.10).
-
Key references: Aboal et al. (2018).
Genus Kyliniella Skuja, Acta Hort Bot Univ Latvi 1:4 (1926)
-
The genus is monospecific and the characters are described below.
-
Kyliniella latvica Skuja, Acta Hort Bot Univ Latvi 1:4 (1926) (Fig. 2.12h–j)
-
Type: S A36450, H Skuja, 20.viii.1926 (Holotype); MICH 660558 (Isotype).
-
Type locality: Latvia, Usma Parish, Lake Usma, 57.160699° N, 22.170287° E (estimated).
-
Description: pseudofilaments unbranched, arising from a discoid and pseudoparenchymatous base, 12–32 μm in diameter; cells discoid or cubic, contiguous, or separate, arranged within a broad mucilaginous sheath; each cell containing several parietal, reddish, discoid, or band-shaped chloroplasts, 4.9–17.5(–22) μm in length and 7.4–11(–19) μm in diameter, length/diameter 0.5–1.0; rhizoidal outgrowths occur at points of contact among cells, 17–25(–170) μm in length, 7–10–(12.5) μm in diameter. Asexual reproduction by small fragments (hormogonia) and monosporangia released from the sheath. Presumptive sexual reproduction with small colorless spermatia and large, pigmented carpogonia with tubular projections. Postfertilization structures unknown.
-
Representative sequences in GenBank: DQ30843 (rbcL).
-
Distribution: Europe: Austria, France, Germany, Latvia, Sweden; North America: northeastern United States (Rhode Island and New Hampshire) (Fig 2.10).
-
Habitat: this taxon has few records and is an epiphyte on macrophytes and macroalgae; in North America, it is a rare component in the littoral zone of soft water streams (Sheath and Vis 2015); in Europe, it is reported associated with macrophytes in hardwater streams with temperature 4.7–9.4° C, conductivity 515–785 μS cm-1, and pH 8.2–8.5 (García-Fernández et al. 2012).
-
Key references: Skuja (1926), Vis and Sheath (1993), Eloranta et al. (2011), García-Fernández et al. (2012).
Genus Rhodospora Geitler, Öst Bot Zeit 76:28 (1927)
-
The genus is monospecific and the characters are described below.
-
Rhodospora sordida Geitler, Öst Bot Zeit 76:28 (1927) (Fig. 2.12k, l)
-
Type: not found.
-
Type locality: Austria, Salzburg, Bad Gastein, 47.114089° N, 13.132661° E (estimated).
-
Description: unicells 4.5–18 μm in diameter, sometimes united into irregular colonies within a mucilaginous matrix which may be layered; each cell with several parietal, discoid, red-violet, olive-green or yellow chloroplasts; reproduction by cell division and formation of 4–32 autospores. Sexual reproduction unknown.
-
Representative sequences in GenBank: DQ308433 (rbcL ).
-
Distribution: Asia: Japan; Europe: Austria, Spain, Sweden; North America: the USA (Arkansas, Tennessee, Ohio) (Fig 2.10).
-
Habitat: this taxon has few records and is an epiphyte on macrophytes and macroalgae .
-
Key references: Geitler (1927), Johansen et al. (2005), Eloranta et al. (2011).
Order Rufusiales Zuccarello and JA West, J Phycol 44:392 (2008)
-
Thalli consisting of branched pseudofilaments; cells with several red to violet, parietal discoid to band-shaped chloroplasts without pyrenoid; asexual reproduction by fragmentation or by many endospores per cell; found on the hair of sloths. The order is monotypic with a single genus and species that are described below. The order has only one family (Rufusiaceae Zuccarello and West) with a single genus (Rufusia Wujek and Timpano).
Rufusia pilicola Wujek and Timpano, Brenesia 25/26:165 (1986) (Fig. 2.12m, n)
-
Type : figures 1–4, page 168 in Wujek and Timpano (1986, Holotype).
-
Type locality: Panama, Barro Colorado Island, 9.150722° N, 79.846437° W (estimated).
-
Description: vegetative cells barrel-shaped or short-cylindrical, 5.5–10 μm in diameter and 3.5–10 μm in length; mature reproductive cells 10–15 μm in diameter; endospores 1.5–2 μm in diameter.
-
Representative sequences in GenBank: DQ308435 (rbcL); DQ308455 (psaA).
-
Distribution: North America—Costa Rica, Panama (Fig 2.10).
-
Habitat: this alga grows within fissures and furrows of the hair of three-toed (Bradypus) and two-toed (Choloepus) sloths.
-
Key references: Wujek and Timpano (1986), Zuccarello et al. (2008).

(a, b) Chroothece lobata : (a) colony with stalks; (b) detail of cell showing stalk (arrow); (c, d) Chroothece mobilis: (c) colony with pseudofilament (arrow) and branch (arrowhead); (d) detail of cells with stellate chloroplasts and pyrenoids (arrows); (e, f) Chroothece richteriana: (e) general view of the mat in the field; (f) colony with stalks (arrows) and cells with pyrenoids (arrowheads); (g) Chroothece thermalis: (g) detail of cells with pyrenoids (arrows); (h–j) Kyliniella latvica: (h) pseudofilament with rhizoidal outgrowths (arrows); (i) pseudofilament with monosporangia (arrows); (j) pseudofilaments with cells showing band-shaped chloroplasts (arrows); (k, l) Rhodospora sordida: (k) colony with unicells (arrows) and cells with autospores (arrowheads); (l) cell with nucleus (arrow) and discoid chloroplasts (arrowheads); (m, n) Rufusia pilicola: (m) young filaments; (n) mature filaments. Scale bars: (c, f, n) = 20 μm; (a, d, g–k, m) 10 μm; (b) = 5 μm; (l) 2.5 μm (Image author: Fig. (a–c, g, j) M Aboal; Fig. (d, k) JA West; Fig. (e, f) C Carter; Fig. (l) J Johansen; Fig. (m, n) D Wujek)
References
Aboal M, Garcia-Fernández ME, Roldán M, Whitton BA (2014) Ecology, morphology and physiology of Chroothece richteriana (Rhodophyta, Stylonematophyceae) in the highly calcareous Rio Chicamo, south-east Spain. Eur J Phycol 49:83–96. https://doi.org/10.1080/09670262.2014.893018
Aboal M, Chapuis I, Paiano MO, Sánchez P, West JA, Whitton BA, Necchi O Jr (2018) Diversity of Chroothece (Rhodophyta, Stylonematales) including two new species. Eur J Phycol 53:189–197. https://doi.org/10.1080/09670262.2017.1402374
Albertano P, Ciniglia C, Pinto G, Pollio A (2000) The taxonomic position of Cyanidium, Cyanidioschyzon and Galdieria: an update. Hydrobiologia 433:137–143. https://doi.org/10.1023/A:1004031123806
Blinn DW, Prescott GW (1976) A North American distribution record for the rare Rhodophycean, Chroothece mobilis Pascher and Petrova. Am Midl Nat 96:207–210
Bory de St. Vincent JB (1797) Mémoire sur les genres Conferva et Byssus, du chevalier O. Linné. De l’Imprimerie de Louis Carazza, Bordeaux
Bourrelly P (1970) Les algues d’eau douce—Tome 3: les algues bleues et rouges, les Eugléniens, Peridiniens et Cryptomonadines. Editions N Boubée, Paris
Christensen T (1962) Botanik. Bind II. Systematisk botanik. Nr. 2. Alger. Kommission hos Munksgaard, København, pp 1–178
Ciniglia C, Yoon HS, Pollio A, Pinto G, Bhattacharya D (2004) Hidden biodiversity of the extremophilic Cyanidiales red algae. Mol Ecol 13:1827–1838. https://doi.org/10.1111/j.1365-294X.2004.02180.x
Ciniglia C, Yang EC, Pollio A, Pinto G, Iovinella M, Vitale L, Yoon HS (2014) Cyanidiophyceae in Iceland: plastid rbcL gene elucidates origin and dispersal of extremophilic Galdieria sulphuraria and G. maxima (Galdieriaceae, Rhodophyta). Phycologia 53:542–551. https://doi.org/10.2216/14-032.1
De Luca P, Taddei R, Varano L (1978) “Cyanidioschyzon merolae”: a new alga of thermal acidic environments. Webbia 33:37–44
Deason T, Nichols HW (1970) A new bangiophycidean alga from Alabama. J Phycol 6:39–43. https://doi.org/10.1111/j.1529-8817.1970.tb02354.x
Drew KM (1956) Reproduction in the Bangiophycidae. Bot Rev 22:553–611
Drew KM, Ross R (1965) Some generic names in the Bangiophycidae. Taxon 14:93–99
Eloranta P, Kwandrans J, Kusel-Fetzmann E (2011) Rhodophyta and Phaeophyceae. In: Büdel B, Gärtner G, Krientz L, Preisig HR, Schagerl (eds) Süsswasserflora von Mitteleuropa, vol 7. Spektrum Akad, Heidelberg, pp 1–155
Galdieri A (1899) Su di unálga che cresce intorno alle fumarole della Solfatera. Atti Reale Accad Sci Fis Mat Napoli 5:160–164. (Ser. 3)
García-Fernández ME, Chapuis IS, Aboal M (2012) Kyliniella latvica Skuja (Stylonemataceae, Stylonematophyceae), un rodófito indicador de buena calidad del agua. Limnetica 31:341–348. DOI https://doi.org/10.23818/limn.31.29
Geitler L (1923) Porphyridium aerugineum nov. sp. Öst Bot Z 72:84
Geitler L (1927) Rhodospora sordida nov. gen. et n. sp. eine neue Bangiaceae des Süsswassers. Öst Bot Z 76:25–28
Geitler L (1932) Porphyridium sordidum n. sp., eine neue Süsswasserbangiale. Arch Protistenk 76:595–604
Geitler L (1934) Diagnosen neuer Blaualgen von den Sunda-Inseln. Arch Hydrobiol Suppl 12:622–634
Hansgirg A (1884) Bemerkungen zur Systematik einiger Süsswasseralgen. Öst Bot Zeit 34:351–358
Hansgirg A (1885) Ein Beitrag zur Kenntniss von der Verbreitung der Chromatophoren und Zellkernen bei den Schizophyceen (Phycochromaceen). Ber Deutsch Bot Ges 3:14–22
Herndon WR (1964) Boldia: a new rhodophycean genus. Am J Bot 51:575–581. doi:https://doi.org/10.1002/j.1537-2197.1964.tb06673.x
Howard RV, Parker BC (1980) Revision of Boldia erythrosiphon Herndon (Rodophyta, Bangiales). Am J Bot 67:413–422. https://doi.org/10.1002/j.1537-2197.1980.tb07666.x
Hsieh CJ, Zhan SH, Lin Y, Tang SL, Liu SL (2015) Analysis of rbcL sequences reveals the global biodiversity, community structure, and biogeographical pattern of thermoacidophilic red algae (Cyanidiales). J Phycol 51:682–694. https://doi.org/10.1111/jpy.12310
Johansen JR, Fucikova K, Fitzpatrick MH, Lowe RL (2005) The red alga genus Rhodospora (Bangiophycidae, Rhodophyta): first report from North America. J Phycol 41:1281–1283. https://doi.org/10.1111/j.1529-8817.2005.00145.x
Korshikov AA (1930) Glaucosphaera vacuolata, a new member of the Glaucophyceae. Arch Protistenk 70:217–222
Kumano S (2002) Freshwater red algae of the world. Biopress, Bristol
Liu S-L, Chiang Y-R, Yoon HS, Fu H-Y (2020) Comparative genome analysis reveals Cyanidiococcus gen. nov., a new extremophilic red algal genus sister to Cyanidioschyzon (Cyanidioschyzonaceae, Rhodophyta). J Phycol 56:1428–1442. https://doi.org/10.1111/jpy.13056
Medina-Cabrera EV, Rühmanna B, Schmid J, Sieber V (2020) Characterization and comparison of Porphyridium sordidum and Porphyridium purpureum concerning growth characteristics and polysaccharide production. Algal Res 49:101931. https://doi.org/10.1016/j.algal.2020.101931
Merola A, Castaldo R, DeLuca P, Gambardella R, Musacchio A, Taddei R (1981) Revision of Cyanidium caldarium. Three species of acidophilic algae. Giorn Bot Ital 115:189–195. https://doi.org/10.1080/11263508109428026
Montagne C (1846) Flore d’Algérie. Ordo I. Phyceae Fries. In: Bory MM, Maisonneuve D (eds) Exploration scientifique de l’Algérie pendant les années 1840, 1841, 1842, Sciences physiques. Botanique. Cryptogamie, vol 1. Imprimerie Royale, Paris, pp 1–197
Muñoz-Gómez SA, Mejía-Franco FG, Durnin K, Colp M, Grisdale CJ, Archibald JM, Slamovitis CH (2017) The new red algal subphylum Proteorhodophytina comprises the largest and most divergent plastid genomes known. Curr Biol 27:1677–1684. https://doi.org/10.1016/j.cub.2017.04.054
Nägeli C (1849) Gattungen einzelliger Algen, physiologisch und systematisch bearbeitet. Neue Denkschr Allg Schweiz Gesellsch Gesammten Naturwiss 10:1–139
Nakayama T, Yokoyama A (2017) Classes Cyanidiophyceae, Compsopogonophyceae, Porphyridiophyceae, Rhodellophyceae and Stylonematophyceae. In: Kamiya M, Lindstrom SC, Nakayama T et al (eds) Syllabus of plant families; Adolf Engler’s Syllabus der Planzenfamilien—Part 2/2—Photoautotrophic eukaryotic algae—Rhodophyta. Borntraeger Sci Publ, Stuttgart, pp 19–30
Necchi O Jr (2016) Red algae (Rhodophyta) in rivers. In: Necchi O Jr (ed) River Algae. Springer, Cham, pp 65–91. https://doi.org/10.1007/978-3-319-31984-1_4
Necchi O Jr, Dip MR (1992) The family Compsopogonaceae (Rhodophyta) in Brazil. Algol Stud 66:105–118
Necchi O Jr, Garcia Fo AS, Salomaki ED, West JA, Aboal M, Vis ML (2013) Global sampling reveals low genetic diversity within Compsopogon (Compsopogonales, Rhodophyta). Eur J Phycol 48:152–162. https://doi.org/10.1080/09670262.2013.783626
Ott FD (1972) A review of the synonyms and the taxonomic positions of the algal genus Porphyridium Nägeli 1849. Nova Hedwigia 23:237–289
Ott FD (1976) Further observations on the freshwater alga Flintiella sanguinaria Ott in Bourrelly 1970 (Rhodophycophyta, Porphyridiales). Arch Protistenk 118:34–52
Ott FD (1987) A brief review of the species of Porphyridium with additional records for the rarely collected alga Porphyridium sordidum Geitler, 1932 (Rhodophycophyta, Porphyridiales). Arch Protistenk 134:35–41. https://doi.org/10.1016/S0003-9365(87)80024-6
Ott FD (2009) Handbook of the taxonomic names associated with the non-marine Rhodophycophyta. J Cramer, Berlin
Pascher A, Petrová J (1933) Über Porenapparate und Bewegung bei einer neuen Bangiale (Chroothece mobilis). Arch Protistenk 74:490–522
Pekárková B, Smarda J, Hindák F (1989) Cell morphology and growth characteristics of Porphyridium aerugineum (Rhodophyta). Plant Syst Evol 164:263–272. https://doi.org/10.1007/BF00940442
Pinto G, Ciniglia C, Cascone C, Pollio A (2010) Species composition of Cyanidiales assemblages in Pisciarelli (Campi Flegrei, Italy) and occurrence of Galdieria phlegrea sp. nov. In: Seckbach J (ed) Algae and Cyanobacteria in extreme environments. Springer, Dordrecht, pp 487–502. https://doi.org/10.1007/978-1-4020-6112-7
Reeb V, Bhattacharya D (2010) The thermo-acidophilic Cyanidiophyceae (Cyanidiales). In: Seckbach J, Chapman DJ (eds) Red algae in the genomic age. Springer, Dordrecht, pp 409–426. https://doi.org/10.1007/978-90-481-3795-4_22
Rintoul T, Sheath RG, Vis ML (1999) Systematics and biogeography of the Compsopogonales (Rhodophyta) with emphasis on freshwater genera in North America. Phycologia 38:517–527. https://doi.org/10.2216/i0031-8884-38-6-517.1
Saunders GW, Hommersand M (2004) Assessing red algal supraordinal diversity and taxonomy in the context of contemporary systematic data. Am J Bot 91:1494–1507. https://doi.org/10.3732/ajb.91.10.1494
Schmitz F (1896) Compsopogonaceae. In: Engler A, Prantl K (eds) Die natürlichen Pflanzenfamilien nebst ihren Gattungen und wichtigeren Arten insbesondere den Nutzpflanzen unter Mitwirkung zahlreicher hervorragender Fachgelehrten, Teil 1, Abteilung 2. von Wilhelm Engelmann, Leipzig, pp 318–320
Shameel M (2001) An approach to the classification of algae in the new millennium. Pak J Mar Biol 7:233–250
Sheath RG, Hymes BJ (1980) A preliminary investigation of the freshwater red algae in streams of southern Ontario, Canada. Can J Bot 58:1295–1318. https://doi.org/10.1139/b80-161
Sheath RG, Sherwood AR (2011) Rhodophyta (red algae). In: John DM, Whitton BA, Brook AJ (eds) The freshwater algal flora of the British Isles. An identification guide to freshwater and terrestrial algae, 2nd edn. Cambridge University Press, Cambridge, pp 159–180
Sheath RG, Vis ML (2015) Red algae. In: Wehr JD, Sheath RG, Kociolek JP (eds) Freshwater Algae of North America: ecology and classification, 2nd edn. Academic Press, San Diego, pp 237–264. https://doi.org/10.1016/B978-0-12-385876-4.00005-0
Sheath RG, Hamilton PB, Hambrook JA, Cole KM (1989) Stream macroalgae of the eastern boreal forest region of North America. Can J Bot 67:3553–3562. https://doi.org/10.1139/b89-437
Skuja H (1926) Eine neue Süsswasserbangiaceae Kyliniella n.g., n. sp. Acta Hort Bot Univ Latv 1:1–6
Skuja H (1939) Versuch einer systematischen Einteilung der Bangioideen order Protoflorideen. Acta Hort Bot Univ Latv 11(12):23–28
Vis ML, Sheath RG (1993) Distribution and systematics of Chroodactylon and Kyliniella (Porphyridiales, Rhodophyta) from North American streams. Jpn J Phycol 41:237–241
Vis ML, Sheath RG, Cole KM (1992) Systematics of the freshwater red algal family Compsopogonaceae in North America. Phycologia 31:564–575. https://doi.org/10.2216/i0031-8884-31-6-564.1
West JA, Zuccarello GC, Scott JL, West KA, Loiseaux de Goer S (2007a) Pulvinus veneticus gen. et sp. nov. (Compsopogonales, Rhodophyta) from Vanuatu. Phycologia 46:237–246. https://www.tandfonline.com/doi/abs/10.2216/06-66.1
West JA, Zuccarello GC, Scott JL, West KA, Loiseaux de Goer S (2007b) Correction to paper by West et al. Phycologia 46(3):237–246, Phycologia 46:478
Wilson SM, Pickett-Heaps JD, West JA (2006) Vesicle transport and the cytoskeleton in the unicellular red alga Glaucosphaera vacuolata. Phycol Res 54:15–20. https://doi.org/10.1111/j.1440-1835.2006.00404.x
Wittrock VB, Nordstedt CFO (1884) Algae aquae dulcis exsiccatae praecipue Scandinavicae quas adjectis algis marinis chlorophyllaceis et phycochromaceis. Bot Not 1884:121–128
Wujek DE, Timpano P (1986) Rufusia (Porphyridiales, Phragmonemataceae), a new red alga from sloth hair. Brenesia 25/26:163–168
Yoon HS, Hackett JD, Ciniglia C, Pinto G, Bhattacharya D (2004) A molecular timeline for the origin of photosynthetic eukaryotes. Mol Biol Evol 21:809–8818. doi:https://doi.org/10.1093/molbev/msh075
Yoon HS, Ciniglia C, Wu M, Comeron JM, Pinto G, Pollio A, Bhattacharya D (2006a) Establishment of endolithic populations of extremophilic Cyanidiales (Rhodophyta). BMC Evol Biol 6:78. https://doi.org/10.1186/1471-2148-6-78
Yoon HS, Müller KM, Sheath RG, Ott FD, Bhattacharya D (2006b) Defining the major lineages of red algae (Rhodophyta). J Phycol 42:482–492. https://doi.org/10.1111/j.1529-8817.2006.00210.x
Zuccarello GC, West JA, Kikuchi N (2008) Phylogenetic relationships within the Stylonematales (Stylonematophyceae, Rhodophyta): biogeographic patterns do not apply to Stylonema alsidii. J Phycol 44:384–393. https://doi.org/10.1111/j.1529-8817.2008.00467.x
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Copyright information
© 2021 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Necchi Jr, O., L Vis, M. (2021). Subphylum Cyanidiophytina, Class Cyanidiophyceae; Subphylum Proteorhodophytina, Classes Compsopogonophyceae, Porphyridiophyceae, Rhodellophyceae, and Stylonematophyceae. In: Freshwater Red Algae. Springer, Cham. https://doi.org/10.1007/978-3-030-83970-3_2
Download citation
DOI: https://doi.org/10.1007/978-3-030-83970-3_2
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-83969-7
Online ISBN: 978-3-030-83970-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)