
Abstract
Living organisms involved in wood biodeterioration range quite broadly across Eukarya, Bacteria, and Archaea domains with many wood degrading species in the latter two domains still likely undiscovered. This chapter focuses on some of the more prominent of these agents involved in the degradation of commercial wood products but also those that decay wood biomass in the environment. Because of the importance of select organisms relative to monetary costs and destructive capability, most of the chapter is focused on Eukaryotic organisms. Specific focus is on different species and types of fungi, insects, and marine organisms that degrade wood either structurally or, in some cases, just through changes in physical appearance making wood undesirable for esthetic reasons. Bacteria are also discussed briefly because, although abundant and very important in carbon cycling in the environment, bacteria typically cause less destruction to wood as a structural material, over a much longer time frame.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Fungi
- Decay
- Stain
- Brown rot
- White rot
- Soft rot
- Bacteria
- Archaea
- Termites
- Carpenter ants
- Beetles
- Shipworms
- Gribble
- Pholads
- Lignocellulose
1 Overview of Biodeterioration Agents Presented in This Chapter
Understanding of wood biodeterioration is not complete without knowledge of the underpinnings of how different organisms attack and deconstruct the substrate at the chemical and nanoscale; therefore, a portion of the content in this chapter is also devoted to the current state of art relative to what is known about degradative mechanisms (Fig. 4.1) [1]. It should be understood that in many cases, the exact mechanisms involved in the degradation of wood are still being refined by researchers. Therefore, an overview of general mechanisms is provided with insights on the direction or directions the field is taking at present to resolve unknowns from a scientific level. The information on mechanisms is important because it relates to our understanding of the basic biology of degradative organisms and their capacity to attack wood (see Chaps. 15; 16). However, it becomes even more important when one considers new ways to protect wood from deterioration, particularly with increasing restrictions on traditional biocides. New methods for the protection of wood, and for inhibiting the action of biological organisms in degrading wood, are required if nontoxic or low-toxicity protection systems that target specific mechanisms are to be developed or enhanced in the future.

Selected genera and species in three domains that are involved in the degradation of wood and woody biomass: Eukarya, Bacteria, and Archaea. The Eukaryotes with fungi, mollusks, arthropods, and even chordata organisms are considered in this chapter, together with some Bacterial systems. Because of limited information available on the Archaea, and the limited degradation that occurs by Archaea in structural environments, degradation of wood by the Archaea is not a focus. (From [1], licensed under CC BY 4.0)
2 Mechanisms of Biodeterioration
Mechanisms of biodeterioration are quite complex and depend on the specific organism(s) involved. In some cases, symbiotic action between two types of organisms is known to be required for successful degradation and metabolism of wood as a substrate and, for example, the action of some termites in attacking wood is not possible without the attendant bacteria, archaea, and protozoans in the termite gut. Biological degradation is more complex than simple chemical or biochemical action on the substrate since, in addition to having a system to break down lignocellulose into component molecules, those breakdown products must nourish the organism so that continued degradation of wood will occur. Further, breakdown products must also not be toxic to the organism or similarly, degradation will stop.
Mechanical action is an important mode for many higher wood degrading organisms in initially reducing the size of wood particles so that wood can be ingested. This is true, for example, with termites and some types of wood boring beetles [2], but also with several types of marine organisms, including some types of fish. These organisms must first reduce the size of wood to small particles before later action by chemical and enzymatic means to further digest wood to chemical components.
Further chapter sections will expand on specific known and presumed modes of action necessary for different organisms to break down wood and woody materials.
3 Fungi That Cause Wood Deterioration
For purposes of classification, the chapter considers fungi both from their phylogenetic perspective and practical damage-causation perspective. For example, Basidiomycota species are some of the most damaging fungi that cause what is known as either brown rot or white rot decay of wood. However, certain Ascomycota species also cause a decay of wood and therefore are classified as decay fungi causing “soft rot” of wood. This practical classification will be followed here.
Fungi that cause deterioration of wood are primarily filamentous in form, and the body of the fungus consists of fine (2–10 μM), hair-like, elongated cells that branch and connect end to end to form an interconnected network. Individual strands of the fungus are known as hyphae (singular hypha), whereas when may hyphae grow together in layers, they are called mycelia or a mycelial matt. Mycelia can also form unique structures such as sporophores (fruiting bodies) in sexually capable fungi. These fruiting bodies may take the form of cups or discs in some Ascomycota species, while in the Basidiomycota, mushrooms or bracket structures on the side of wood substrates are common. Common/colloquial names for the sporophores of Basidiomycota growing from wood include conks, punks, shelf fungi, and others.
Mold fungi, Stain fungi, Soft rots, Brown rots, and White rots are the groupings typically used for fungi that cause different types of degradation. Mold fungi are generally found in the Ascomycota. Traditionally many of the mold fungi were classified as Fungi Imperfecti or Deuteromycetes (not a formal taxa), and as time has progressed with new molecular techniques, many of these fungi have been able to be classified as either Ascomycota or Basidiomycota. As will be discussed later in the section on Molds, mold fungi do not penetrate wood surfaces more than one or two cells, and they do not metabolize structural wood components. Because they do not degrade cellulose, lignin, or hemicellulose, they do not cause appreciable mass loss as mold infection advances.
Stain fungi are able to penetrate into the sapwood of wood and sometimes living trees, and they feed on the sugars and starches primarily found in the parenchyma cells. They often initiate on the surface of wood and in that regard in early staining stages, they intergrade with and can be confused as being mold fungi. Although not common, some fungal species classified generally as stain fungi have also been found to attack the structural components of wood [3]. In general however, stain fungi cause only limited mass loss and cause only limited structural damage to wood.
Wood decay fungi including brown rots, white rots, and soft rot fungi generally have the capacity to attack both holocellulose and lignin components. The type of attack will vary as reviewed for the specific decay types throughout this chapter. However, as decay progresses, the fungi not only depolymerize all or portions of the lignocellulose, they metabolize the low molecular weight fractions of the substrate once adequate depolymerization has occurred to allow passage of the wood substrate molecules through the fungal cell wall and membrane. As depolymerization of holocellulose and lignin occurs, mechanical property loss occurs in the wood. In some cases, depolymerization is paced to match metabolism by the fungus, and in such cases, mass loss of the wood will parallel mechanical property loss. More often however, as in the initial stages of decay by brown rot fungi, rapid depolymerization of the wood cell wall occurs with little metabolism of the cell wall components by the fungus, so mechanical property loss can advance significantly before large mass loss has occurred. Concurrent with mass loss as decay progresses, shrinkage of wood occurs. In some decay types, the wood cell wall itself develops a porous structure so that bulk shrinkage may be less noticeable as mass loss increases. However, in brown rot of softwoods, it has been noted that the wood cell wall collapses upon itself [4] with rearrangement of modified lignin [5]. This ultimately results in wood shrinkage at the macroscale, and the cubical, friable appearance of brown rotted wood as this type of decay advances. Shrinkage is also observed in other types of decay but is often masked because of microporosity development within the cell walls, or when cells delaminate at the middle lamellae uniformly, as these two phenomena provide a more homogenous appearance of the wood without obvious ruptures/cracks at the macroscale.
Fungal hyphae are small enough to penetrate into the lumens of wood cells, and in stain and decay fungi, the hyphae also travel through and penetrate pit membranes such as bordered pits and ray parenchyma pits. The hyphae must ramify across the surface of the wood in the case of molds, or on the surface and throughout interior cells of wood in the case of stain and decay fungi, in order to obtain nutrients which are required for fungal survival. Extracellular enzymes, and often low molecular weight fungal metabolites, are secreted by the fungal hyphae to solubilize compounds ranging from simple oligosaccharides to polymerized lignin. The goal of most fungi is to obtain sugars or short-chain oligosaccharides that can be absorbed or actively transported into the fungal thallus. Degraded and modified fragments of lignin can also be metabolized by some fungi, and the hyphae of these types of fungi again are responsible for secretion of metabolites that can depolymerize lignin.
All fungi require moisture to grow in or on the surface of wood. Generally, the moisture content required for decay fungi to grow into wood and secrete metabolites into the wood cell wall must be above the fiber saturation point. This is because enzymes and other degradative components of the fungal secretome (low molecular weight metabolites involved in fungal decay processes) must be able to diffuse from the fungus through an extracellular fungal matrix (ECM) which surrounds all fungal hyphae, and to the cell wall. Liquid water must therefore be present in the lumens of the wood cells for this type of diffusion to occur at the surface of the fungal hyphae. The wood cell wall must also be at, or near, fiber saturation to allow low molecular weight fungal metabolites and ions to diffuse within the wall as a prerequisite for decay initiation. Mold fungi only require moisture to be present on the surface of wood, or other substrates, to grow. Many architectural manuals indicate that mold will grow on wood that is only at 20% MC. Technically, this is unlikely to occur because, as with all degradative fungi, some amount of liquid water is needed for extracellular enzyme secretion. Practically speaking however, the 20% MC figure is useful, particularly when the wood MC is in equilibrium with moisture in the air in an enclosed or poorly ventilated space. Depending on temperature, the relative humidity (RH) required to maintain 20% MC in wood is close to 90% RH. Over a broad temperature range from 3 °C to 32 °C, a decrease in temperature of only ~2 °C will allow the dew point to be reached, and condensation of liquid water at the surface of wood under such conditions can then occur. Although small amounts of surface water on wood would not allow decay in any form to advance, some molds and even surface stains could establish growth, as staining and mold can develop in a matter of a few days at moderate temperatures above approximately 10 °C.
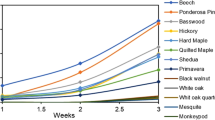
As a caveat to the discussion on moisture content limitations for fungal growth, very wet wood will also limit or stop fungal growth. Most decay fungi have a MC optimum for decay in the range between 50% and 100% MC (Fig. 4.2 [6]) and moisture content optima exist for other fungal species as well. For many soft rot fungi, moisture content optima for decay has been reported to be much higher, “near saturation”; however, aeration of wood samples undergoing soft rot attack has also been assessed to be critical for optimal soft rot decay [7]. This suggests that some soft rot fungi may be able to survive and thrive under conditions where they can pull dissolved oxygen from water. Relative to moisture optima for wood decay fungi in different density species, it has been observed that lower density woods will continue to decay at high moisture content levels that stop decay in high density species. This is because lower density wood species have proportionally greater cell lumen volumes compared to denser wood species. This is important because, like most Eukaryotes, wood decay fungi require oxygen for growth. Prior research has shown that some decay fungi are capable of decaying wood with oxygen levels as low as 1%; however, optimal levels were found at atmospheric oxygen levels (21%).

Decay optima vary with moisture content and also with the density of wood species. Many other factors, including fungal species, are important in determining rates of decay, but it is important to recognize that aggressiveness of decay can vary simply by altering moisture content; particularly in higher density wood species. (From [6])
3.1 Wood Decay Fungi: Basidiomycota Decay and Ascomycota Soft Rot Decay
Basidiomycota fungal genera that decay wood are generally classed informally as either white rot or brown rot fungi. Soft rot, Ascomycota, fungi are also included in this section because of the structural damage that they cause. These fungi in filamentous form grow through wood; first seeking sites within or on the surface of wood, where simple sugars and starches are available such as in the parenchyma of the wood or other lignocellulosic materials. The use of the simple substrates in initial colonization of woody substrates allows these fungi to build up mycelial biomass and move through the substrate without the need for expenditure of greater amounts of energy which is needed for the more complex tasks of deconstructing lignocellulose. With most species, when conditions are right and particularly when the simple sugars and starches are depleted, the cellular machinery for deconstruction of wood cell walls is started. With staining fungi, and white rot and brown rot, hemicelluloses are often then attacked, as these polysaccharides are the most accessible components of the wood cell wall, and they are generally more loosely packed at the molecular level. As decay progresses, the fungi then use different mechanisms for breaking down cellulose, lignin, and more tightly bound hemicellulose, as well as the more minor constituents of pectins, and extractives embedded in the cell wall. Inducible extracellular enzymes for deconstruction of cellulose and/or lignin are upregulated in many decay fungi when the more readily digested sugar, polysaccharide, and fatty acid substrates in wood are depleted. In these fungi, specific enzymes are secreted to depolymerize or hydrolyze specific chemical linkages in substrates. Glycoside hydrolase (GH) enzymes, for example, are enzymes that hydrolyze glycosidic bonds in either cellulose or hemicellulose. There are many different types of GHs and many families of this types of enzyme are now known [8]. Other enzymes that may be present in some fungal species will target specific linkages in lignin or hemicellulose, such as fungal esterases that have high affinity for acetylated carbohydrates such as would be found in some hemicelluloses. It has been suggested by some researchers that expansins or swollinens may also contribute to “loosening of the plant cell wall structure,” and upregulation of expansins during decay has been previously noted in some fungi [9]. However, the mode of action of expansins and swollinens remains largely unknown. Some fungal enzymes are also specific for deconstruction of specific types of pectin in the bordered pits or middle lamella, while other enzymes, particularly in white rot fungi, have broad specificity in degrading aromatic compounds.
White rot fungi decay wood primarily via the action of extracellular enzymatic systems. In some cases however, low molecular weight mediators or metal radical ions are known, or have been proposed, to work in conjunction with lignin-degrading enzymes in white rots to explain the types of degradation patterns observed at the nanoscale. Brown rot fungi decay wood use a low molecular weight catalytic system that can penetrate into the wood cell wall in early stages of attack by the fungus. The pattern of attack by this low molecular weight catalytic system is consistent with attack by hydroxyl radicals, and the most prominent theory on the low molecular weight degradation mechanism involved outlines a mechanism for hydroxyl radical attack [5, 10, 11] which is discussed in the section on nonenzymatic brown rot mechanisms in this chapter. Brown rot fungi discarded their machinery for production of most peroxidases (lignin degrading enzymes) and also many of their carbohydrate active enzymes (CAZymes) and oxidoreductases as they evolved a low molecular weight nonenzymatic system to bio-catalytically deconstruct cellulose and lignin. Although some fungal species are now considered to fall between brown rot and white rot fungi [12] relative to the mechanisms used to deconstruct wood, this chapter will maintain traditional definitions of the two for convenience and allow the reader to understand the differences at either end of the spectra.
Wood degrading Basidiomycota fungi evolved approximately 295 million years ago to produce Class II – peroxidase enzymes (PODs), enzymes which allowed these particular progenitors of current day white rot fungi to deconstruct lignin. Basidiomycota species prior to this possessed enzymes capable of depolymerizing and metabolizing cellulose, and therefore, they were able to decompose grasses and sedges. However, as tracheophytes evolved as the first plants to incorporate lignin into to their cell walls about 375 million years ago, the fungal species at that time were unable to deconstruct this new lignified plant cellular material. Lignification allowed plants to grow larger and taller. Researchers have suggested that the gap of approximately 90 million years between lignin production in plants and the ability of fungi to decay the lignin in those plants is at least in part responsible for the buildup of vegetation which resulted in current day coal seams in many locations on earth [13, 14].
It is important to note the different roles of extracellular enzymes and low molecular weight catalytic (LMWC) systems in the wood degradation process. There is a large body of literature that demonstrates that although extracellular fungal enzymes can erode the surfaces of plant cell walls at the molecular level, all enzymes (no matter the source) are too large to penetrate the intact structure of secondary plant cell walls for more than a nanometer at best. In plants such as corn and Arabidopsis, where secondary walls do not exist, and in particular when delignification procedures have been used [15], enzymatic erosion and penetration of one to a few elementary microfibril layers has been observed. However, in intact secondary cell walls of wood, enzymes do not readily penetrate. This has been demonstrated definitively in white rot fungi, brown rot fungi, and via the use of many different probes of sizes simulating the geometry of enzymes [16,17,18]. The importance of nonenzymatic mechanisms and the lack of penetration of enzymes into intact lignified cell walls is important although it is sometimes ignored by pathologists and microbial physiologists studying plant cell walls. Brown rot fungi and some types of white rot fungi employ LMWC systems to penetrate relatively deep into cell walls and even into the middle lamella to depolymerize and solubilize cell wall components and overcome the limitations imposed by the bulkiness and size of enzymes relative to the unmodified pore structure of the wood secondary cell wall [19]. In the soft rot Ascomycota, it has been suggested that low molecular weight mediator compounds are involved with some enzymatic attack, but this has not been well studied in this type of decay.
3.1.1 Brown Rot Fungi
Brown rot fungi comprise only about 6% of all known Basidiomycota species, yet they degrade approximately 80%, by mass, of wood in the northern hemisphere and other regions of the world [20]. Although brown rot fungi are present globally, in nature they tend to attack softwoods preferentially, and therefore, they have greater impact in the northern hemisphere (primarily boreal forests) of the world and in other regions where softwoods predominate. Research on the shifting brown rot/white rot paradigm highlights that some brown rots are mixed species or hardwood degrading fungi [21]. However, relative to the mass of wood degraded globally, brown rot degradation will remain the most prominent type of decay particularly in the northern hemisphere where coniferous species predominate. Brown rot wood decay fungi are perhaps the most destructive organisms of wood on earth, and for this reason, considerable space in this chapter will be devoted to their action.
Brown rot fungi are classified as such because of the color of the wood residue which remains after fungal degradation has progressed to advanced stages. The wood in this stage is typically brown in color and is classically described as being friable, crumbly, checked across the grain and sometimes having a cross-hatched appearance (Fig. 4.3). However, it is important to note that in early, or incipient, stages of decay by brown rot fungi, the wood often will appear visually unchanged from undecayed wood, other than appearing to be wet in some areas. Early/incipient decay stages are typically defined as wood where mass loss due to decay is at less than 10%. Because brown rot fungi have a unique LMWC mechanism for initiating decay in wood, the low molecular weight components can diffuse through the wood cell wall rapidly to depolymerize both lignin and holocellulose components. It is this depolymerization, particularly of the crystalline cellulose backbone of the elementary fibrils of wood, that dramatically reduces the mechanical properties of wood undergoing brown rot attack. In early work, Wilcox [22] reviewed literature demonstrating that 70% of both modulus of elasticity and modulus of rupture could be lost with degradation by brown rot fungi at a level of only 10% mass loss. Wood at this stage would not appear brown and crumbly, and this is the key reason why early stages of brown rot decay are considered dangerous, because it can be impossible to tell how much strength remains in a wood sample when it is unknown whether the wood has been attacked by brown rot fungi. For this reason, it is critically important to keep untreated wood protected from moisture when wood is used in either interior or exterior structural applications.

Brown rotted wood decay from the collection of Holzforschung München, Technical University of Munich, shown with the first author’s hand for scale. The interior wood is in an advanced stage of brown rot degradation. In advanced stages of decay, the wood is brown in color and often has a cross-hatched or cubical appearance
As reviewed above, in initial stages of brown rot degradation of wood, both holocellulose and lignin are rapidly depolymerized. The fact that lignin is extensively depolymerized has been recognized for some time, and more than 20 years ago it was recognized that hydroxyl radicals would rapidly depolymerize and then repolymerize lignin into a modified form [23, 24] as part of the LMWC mechanism employed in advance of enzymatic attack in brown rot fungi. Yelle et al. (2011) [25] confirmed that the depolymerization occurred and postulated further that repolymerization of the modified lignin would then also occur: “via radical coupling of the phenolic units that become enriched during attack on the aromatic rings by ·OH.” Because lignin is entangled with hemicellulose and encrusts the cellulose elementary fibrils in wood, depolymerization of lignin is an essential byproduct of hydroxyl radical attack on lignocellulose in brown rot degradation. (More on this topic is detailed under the nonenzymatic mechanisms of brown rot fungal degradation below.) Modification of the chemistry of the lignin phenolic units would not be possible without depolymerization. Further, the modified lignin residue that is described in older literature as “slightly modified” lignin is actually quite extensively modified in that aliphatic side-chain cleavage and demethoxylation have occurred, and then the majority of the modified aromatic subunits have repolymerized to produce the brown, crumbly polymerized mass that is observed in heavily brown rotted wood.
Genome sequencing of many brown rot decay fungi has occurred only within the past decade, and prior to that brown rot fungi were generally considered to be primitive species which had not evolved a complete suite of enzymes to deconstruct holocellulose or lignin. The reason why brown rot fungi evolved from the white rot progenitors and discarded many of their extracellular enzyme systems to evolve a LMWC mechanism is unknown. However, because of the more recent evolution of brown rots, only a small percentage (6%) of all Basidiomycota decay species currently known are brown rot species compared to the predominant white rot species. Despite evolving later, the brown rot fungi have exploited their environmental niche relative rapidly, and the vast majority of wood in the world is degraded primarily by brown rots. This suggests that an evolutionary advantage was gained by the brown rot fungi when shifting to a low molecular weight degradative system, with a smaller complement of extracellular CAZymes. The importance of this is discussed by Eastwood [14]: “The brown rot mechanism, where cellulose and hemicelluloses are decomposed leaving lignin modified but largely intact, has evolved from a white rot ancestry at least 5 times. This suggests that there is a strong selection pressure for ability to cast off the energetically expensive need to depolymerize lignin in conifer-dominated habitats.”
3.1.1.1 Nonenzymatic Mechanisms of Brown Rot Fungal Degradation
As noted previously, brown rot fungi lack the full complement of CAZymes and oxidoreductases necessary to deconstruct cellulose. Early researchers studying fungal attack of wood recognized the dilemma of having a class of fungi that aggressively decomposed wood, but did not have the biochemical machinery known at the time to be required for this degradation. Over 50 years ago, Cowling et al. [26, 27] noted that low molecular weight cellulases must be responsible, but 40 years later Cowling lamented calling the active component a very small cellulase rather than a low molecular weight catalytic compound of a more general type (Personal Communication). Highley and also Nicholas and their coworkers in the 1970s and 1980s noted that brown rot decay resembled wood that was treated with Fenton reagent (Fe2+ + H2O2 → Fe3+ + HO• + OH−); however, results using Fenton treatment of wood alone, or Fenton treatment with cellulolytic enzymes, were not able to mimic the action of brown rot. It is important therefore that brown rot degradation not be described by current researchers as employing simple “Fenton reactions” because the chemistries the fungi employ are more complex. Greater efficiencies are achieved, by both the brown rot fungi [28] and in industrial applications [29], when brown rot nonenzymatic chemistries are correctly interpreted.
Research in the 1990s explored the role of cellobiose dehydrogenase with the finding that this enzyme may play a role in iron reduction and subsequent generation of hydroxyl radicals; however, as a general mechanism for cell wall depolymerization in brown rots, this work has been discounted because of the limited number (one) of brown rot species found to produce cellobiose dehydrogenase. There are currently two viable working theories that describe how a low molecular weight catalytic (LMWC) system functions in brown rot fungi to solubilize wood cell wall components in advance of fungal enzymatic action occurring in the wood cell lumen:
-
1.
Glycopeptide theory: The laboratory of Tanaka and Enoki has posited that pyridine coenzymes provide electrons to low molecular weight glycopeptides which can then penetrate the wood cell wall to generate hydroxyl radicals [30]. Genomic analysis has found comparatively high levels of genes encoding for Fe3+-reducing glycopeptides in some brown rot species [12]. However, the mass of the encoded glycopeptides is larger than that of compounds reported to isolated from brown rot fungi with iron reducing activity [31], and it is unclear how the relatively large inferred mass of the glycopeptides would permit penetration into the wood cell wall. Further, like all enzymes, the proposed pyridine coenzymes would be unable to penetrate wood cell walls undergoing degradation to provide electrons to the glycopeptides at least until advanced stages of decay when cell wall porosity is increased. It also remains unclear whether these reductants required for the glycopeptide systems persist in extracellular environments. Given these caveats, it must be explored whether it is feasible for glycopeptides to diffuse into the wood cell wall to reduce iron and then diffuse back out to the coenzymes which would be in the environment surrounding the fungal hyphae. This would be necessary for the complete redox cycle which would permit hydroxyl radicals to be generated within the wood cell wall. Although problems exist in the current hypothesis relative to the function of glycopeptides and their ability to penetrate the wood cell wall, continued research is important to demonstrate what role expressed glycopeptides may play in brown rot degradation of wood, and whether perhaps there may be a role for glycopeptide activity, for example, at the lumen surface of the wood cell wall
-
2.
Chelator-mediated Fenton (CMF) theory: Work in the early 1990s through the current date by Jellison [32, 33], Goodell [24, 34,35,36,36], and others demonstrated that brown rot fungi produce low molecular weight compounds that function to reduce iron in a catalytic (repeated manner) and these compounds participate in oxidative reactions that depolymerize both cellulose and lignin. This hypothesis on the “chelator-mediated Fenton” (CMF) mechanism is now supported by other research groups [38,39,40,41] and has become an established mechanism [42] for nonenzymatic brown rot degradation in advance of enzymatic attack. Initial reports described the CMF mechanism as employing siderophores because related fungal metabolites were isolated using procedures adapted from those used to isolate siderophores from bacteria [35]. Because siderophore receptor sites were not identified on the fungal cell membranes and the catalytic function of siderophores that was observed in the compounds isolated from brown rot fungi has not been widely reported, the terminology of low molecular weight “chelators” has been used rather than “siderophore” in the brown rot literature. The CMF mechanism has been described elsewhere [1, 5, 10] but two types of chelators are involved, with oxalate first sequestering oxidized iron within the low pH environment of the fungal extracellular matrix (ECM) produced by the fungi within the wood cell lumen. As proposed in the CMF mechanism, the fungus regulates oxalate concentrations carefully [43] in the wood cell lumen and oxalate-bound iron diffuses into the cell at a rate limited by the fungus (particularly by the ECM of the fungus). Numerous groups have demonstrated the production of 2,5-dimethoxyquinone (2,5 DMHQ) and other “catecholate” or hydroxyquinone iron-reducing chelators by brown rot fungi including variegatic acid in some species. These compounds, as demonstrated in prior research on the CMF mechanism, will diffuse from the fungal hyphae into the wood cell wall [24]. In the higher pH environment of the wood cell wall (pH 5.5–6.0), and under conditions where iron-oxalate diffusion is limited by the fungal ECM, chelated iron is transferred from oxalate to the catecholate/hydroxyquinone chelators, and iron reduction then spontaneously occurs in this pH environment. Once this chemistry is initiated, a sustained iron-reduction reaction over multiple cycles will occur to generate a stream of hydroxyl radicals. Further, it has been demonstrated that, once initiated, fragmentation to expose lignin surfaces at the micro- and nano-scale caused as part of the decay process may perpetuate the CMF reaction [44]. If so, this would generate a moving radical front through the lignin backbone of the wood cell wall as previously proposed [10]. In all stages, adequate oxygenation of the cell wall must occur, which would be a prerequisite of fungal growth in any case
Relative to CMF chemistry, but also any mechanism which may be found to generate destructive oxygen radicals, the hydroxyl radical is the most potent oxidant known in biological systems, and it is therefore important that it be generated in a location where it cannot damage fungal hyphae. Continuing research in this area has advanced our understanding of how hydroxyl radicals are generated in the wood cell wall. As new research is undertaken and hypotheses developed, these must take into account either how the fungus is protected from hydroxyl radical action [45] or how hydroxyl radical generation is spatially controlled to react within the wood cell wall. CMF chemistry is more complex than neat Fenton reactions, and CMF chemistry has been demonstrated to be more efficient in use of peroxide in generating hydroxyl radicals [29]. It is therefore important that the nonenzymatic chemistries employed by brown rot fungi, not be described in the literature as a simple Fenton reaction.
Relative to other metabolites that may be involved in brown rot degradation of wood, current research suggests that early upregulation of a large group of low molecular weight compounds and enzymes occurs prior to upregulation of glycoside hydrolase enzymes [46]. Some of these metabolites are likely to be involved in LMWC action, with the staggered upregulation of CAZymes occurring later temporally. These findings are consistent with both glycopeptide and CMF working theories, and the staggered upregulation hypothesis also does not preclude mechanisms that allow spatial diffusion of radical-generating low molecular weight metabolites into the wood cell wall to prevent damage to fungal hyphae. Terpene and polyketide synthases have also been found to be upregulated in the brown rot fungi [12], and this may be important relative to any role these synthases play in the production of low molecular weight metabolites ranging from siderophores to fungal antibiotics. Some of these low molecular weight metabolites are structurally related to the low molecular weight compounds isolated from brown rot fungi that have been isolated and proposed for redox chemistries involved in nonenzymatic degradation schemes [24, 47, 48]. More research is needed in this area to explore nonenzymatic catalytic mechanisms in the brown rot fungi, but also in microorganisms in general.
3.1.1.2 Enzymatic Degradation in the Brown Rot Fungi
Riley et al. [12] provide a useful analysis of the CAZymes produced by the brown rot and white rot fungi and how some fungal species do not fit neatly within classical categories. As might be expected for fungi that do not metabolize lignin, lignin-degrading peroxidase enzymes have been lost in the brown rot fungi, although the genomes of some genera have been shown to encode for laccase. Brown rot fungi have a reduced number of enzymes acting on crystalline cellulose, with cellobiohydrolase enzymes absent or lacking a critical cellulose binding domain. Lytic polysaccharide monooxygenase (LPMO) enzymes are reduced in the brown rot fungi compared to white rot species. The Gloeophyllales and Boletales families have one and two cellobiose dehydrogenase enzymes, respectively, but other brown rot families and genera analyzed have lost these enzymes completely. Endoglucanases are considered to be one of the most important enzymes in brown rot fungi, yet they are reduced in number compared to the white rots. No clear difference has been observed between the brown rots and white rots and the number and types of hemicellulase and pectinase enzymes that are produced. Additional research must still confirm the hypotheses on the interaction between nonenzymatic and enzymatic brown rot mechanisms, but the with the reduction in CAZyme complement, including LPMO and endoglucanases in brown rots, the nonenzymatic systems take on a priority role in the brown rot fungi. More studies are required to understand whether interactions between enzymes and LMWC compounds are important in brown rot degradation and, for example, if the some of the low molecular weight metabolites produced by the fungi are important as electron donors for some enzymes like LPMOs. Further, understanding the importance of temporal staging in expression of low molecular weight metabolites and extracellular enzymes remains critical. Currently, it is unclear if staging of nonenzymatic and enzymatic systems is required to protect extracellular enzymes from nonenzymatic radical generating systems, or if generation of the nonenzymatic radicals within the wood cell wall (where enzymes have no accessibility) allows adequate protection of extracellular enzymes. Brown rot fungi may have additional mechanisms that have yet to be discovered, but certainly the use of a dual mechanism to protect critical enzymes is possible.
3.1.1.3 Dry Rot
“Dry rot” is a term that has been used to describe a particular brown rot decay type caused by the genus Serpula with some validity, as this genus has the ability to channel water from several meters away to the site of active decay. Therefore, although the wood still requires water to decay the wood, the requirement that the wood be wet before decay initiation does not hold with this type of brown rot fungus. Serpula lacrymans is common in Europe and Asia, whereas Serpula (Meruliporia) incrassata is the most common of the dry rot types in North America [49]. Serpula dry rots are true brown rot fungi, but their unique ability to channel water from sources at a distance from the wood being infected is permitted by the channeling of water through an elongated hyphal mass know as a rhizomorph structure. Multiple rhizomorphs are often present, and they have been reported to extend as much as 10 meters from the site of decay to reach sources of water. Interior decay of second story wood structures with fungal rhizomorphs extending to wet soil sites exterior to the structure has been observed by the first author.
The term “dry rot” is also sometimes used by laypersons as a misnomer, particularly in North America, to describe brown rotted wood caused by genera other than Serpula, when instead dried decayed wood is being observed. In these cases, the wood would have been decayed by a brown rot fungus in a wetted state, and it has the appearance of brown crumbly wood in the dry state. Typically, the wood would have dried out after the advanced decay stage was reached, stopping the decay process. The term may be used by laypersons (incorrectly) observing wood that has been wetted at some point in its history, and decay had initiated and progressed to an advanced state before water was removed and the wood dried out, but it is not an accepted use of the term dry rot by professionals in the field. As discussed generally for fungi that attack wood, the presence of water in amounts greater than the fiber saturation point (FSP) of the wood is necessary for all types of fungal degradation to occur.
3.1.2 White Rot Fungi
The term “white rot fungi” refers to a group of organisms which attack all of the major wood constituents, primarily via enzymatic action. Often white rotted wood, especially in late stages of decay, is characterized by residual wood material which appears bleached in appearance. In some common types of white rot decay, the wood will become soft and develop a “stringy” character where the softened wood fibers can be easily separated, allowing the wood to be peeled apart. In other types of white rot, pockets or zones of softened, deteriorated wood may appear. Like all wood degrading fungi, white rot fungi require oxygen, water, and a suitable wood substrate for growth. In general, the preferred substrate for white rot fungi is hardwood species or bamboo. Compared to brown rot attack, enhanced growth with corresponding greater mass loss will typically occur when hardwoods are inoculated with white rot fungi.
3.1.2.1 Characteristics of White Rot Wood Degradation
There are two distinct types of white rot from an appearance standpoint, and these are known by different common names. Wood that is white rotted in a uniform manner is variously known as “spongy white rot,” or “stringy white rot,” or because it is the most common type, just “white rot” (Fig. 4.4). The other appearance categorization of white rot is variously known as “pocket white rot” or “pecky white rot” and the wood appears to undergo advanced decay to produce isolated zones of heavily decayed wood, typically filled with white mycelium or bleached wood residue (Fig. 4.5). White rot fungi can also be categorized based on whether holocellulose or lignin is preferentially attacked within the wood cell wall. In “simultaneous white rot,” cellulose, hemicellulose, and lignin are all oxidized and metabolized at approximately the same rate. In “selective white rot,” the literature has suggested different scenarios and it is likely that in different white rot species, slight differences in substrate removal preference occur. Typically, in selective white rot, hemicellulose and lignin are attacked preferentially to cellulose, allowing the cellulose to remain relatively undegraded. However, in some reports, cellulose is also attacked, but it is removed at a slower rate than in simultaneous white rot. Selective white rot fungi have been proposed for their potential use in selective delignification of woody biomass in biotechnological applications including in future biorefineries [50, 51]. As noted in the introduction, in nature there is an intergradation of different decay types [12], and different strains of both simultaneous and selective white rot decays can be produced by the same fungal species [52, 53].

The classic stringy and bleached appearance of white rotted wood in a hardwood. This type of decay is often colloquially referred to as spongy white rot, or stringy white rot

Pocket white rot in a hardwood on the slopes of Mount Kilimanjaro with the 1st author’s hand for scale. (Note the presence of white rotted wood and fungal mycelium within the pockets)
White rotted wood has historically been used for a variety of applications ranging from insulated paneling in Russian refrigerator trucks/vehicles, to cattle feed. In the later application, the selective white rot fungi free the cellulose from the lignin fraction of the wood or fiber, improving the digestibility in ruminant animals. Biotechnological and biorefinery applications of white rot fungi have been explored for more than 30 years [54,55,56] with the initial focus being to free cellulose from lignin to enhance “biopulping” and paper production prospects. More recently, interest in biorefineries, and the production of cellulose-derived sugars for fermentation, with direct conversion to biofuels and platform chemical has been the target. White rot species including Ceriporiopsis subvermispora, Phlebia subserialis, Dichomitus squalens, Pycnoporus cinnarbarinus, and many others have now been tested for potential use in biorefinery applications [57]. Pretreatment by these organisms has been shown to reduce the energy required in refining wood chips in mechanical pulp production and helps improve certain strength and brightness properties of paper. White rot fungi such as Phanerochaete chrysosporium, Phanerochaete crassa, and Pleurotus pulmonarius have been studied as potential bio-bleaching agents for both Kraft pulp and sulfite pulp. Given the need for sustainable production of a wide range of products beyond pulp and paper however, more interest in the past 5 years has been on the conversion of biomass, including lignin, to produce platform chemicals for biopolymers. The use of lignin as an “economic pullthrough” product to provide additional high-value products over and above the holocellulose-derived products is of increasing interest in the field [58, 59]. This has opened up potential for the use of both white rot fungi and lignin residues from brown rots in these applications.
3.1.2.2 Enzymes and Mechanisms of White Rot Degradation
White rot fungi are characterized by their ability to produce a complete enzymatic system capable of degrading hemicellulose and cellulose and able to directly or indirectly oxidize and mineralize lignin [1, 60, 61]. Both endo-glucanases and exo-glucanases that can act synergistically on crystalline cellulose are produced. Enzymatic systems for breakdown of holocellulose include lytic polysaccharide monooxygenases (LPMOs), endo-1,4-b-glucanases, ß-glucosidases, cellobiohydrolases cellobiose dehydrogenases [62], as well as xylosidases, xylanases, acetyl xylan esterases, glucuronidases, and arabinofuranosidases; these later enzymes being necessary for complete depolymerization and oxidation of hemicellulose [63]. The nonenzymatic processes known in white rot fungi, particularly for hemicellulose depolymerization and selective white rot attack, are less well understood, but low molecular weight compounds moving into the wood cell wall in advance of, and in association with, enzyme action have been well documented by Daniel’s laboratory [4, 64] and are reviewed below for specific enzymes.
Relative to holocellulase-active enzymes, white rot fungi have a complete complement of CAZymes including the endo-acting enzymes that also are possessed by brown rot fungi. The white rot fungi also possess processive exo-acting cellulases, also known as cellobiohydrolases (CBH). The cellulase enzymes in white rot fungi are primarily in the glycoside hydrolase (GH) families which attack cellulose hydrolytically, but also include the LPMO enzymes in auxiliary enzyme (AA9) family which oxidatively cleaves cellulose. White rot fungi have many more enzymes that are active on crystalline cellulose, in particular enzymes in the GH6 and GH7 (CBH enzyme) families. In addition, they have many more cellulose binding module (CBM1) family enzymes. For most GH enzymes to be optimally functional, inclusion of a CBM as part of the enzyme is important. Seven known families of hemicellulase enzymes and 11 pectinase enzymes are encoded in the Basidiomycota, but as noted in the section on brown rot fungi, there are no notable differences between white rot and brown rot species relative to the enzyme families for hemicellulases and pectinases in the two types of decay fungi [12].
Because of the unique nature of the lignin degrading enzymes in white rot fungi, this section will cover primarily those enzyme and systems. These include lignin peroxidases, versatile peroxidases, manganese peroxidases, dye decolorizing peroxidases, and laccase. Individual white rot fungal species will possess one or more of these enzymes.
The degradation of lignin in the white rot fungi is biochemically complex. Lignin degrading enzymes and the biochemical mechanisms employed by these enzymes to oxidize lignin with low molecular weight compounds have been well described in a number of reviews [65, 66]. A recent review of Trametes showed the genome encoded for nine putative lignin peroxidases, seven putative short manganese peroxidases, and two putative versatile peroxidases; however, only one manganese peroxidase and one versatile peroxidase were consistently expressed under different growth conditions [67]. This suggests the importance in this common white rot fungus of these later two enzymes, but also indicates that the fungus does not rely on these enzymes alone, and it possesses several “back-up strategies” should conditions require them. Some enzymes have the capability of reacting directly with lignin as a substrate, while others use mediators or low molecular weight compounds involved in electron transfer. Some enzymes are also metal-dependent, requiring chelated or free transition metals to be in close proximity to the enzyme and substrate in ways not yet fully elucidated. Nonenzymatic systems in the white rot fungi do not penetrate as deeply into the wood cell wall as in the nonenzymatic mechanisms of brown rot fungi. Instead, the low molecular weight compounds in these systems appear to be more intimately associated with the enzymes and may be mediator-radicals, oxygen radicals, or chelate-radicals that are generated by action of the enzymes and/or transition metal cofactors. These low molecular weight compounds typically are described as penetrating the wood cell wall in advance of the enzymes [68, 69]. In some selective white rot fungal degradation, altered cell wall structure associated with these low molecular weight compounds has been observed using transmission electron microscopy (Fig. 4.6). The altered lignocellulose structure suggests that these low molecular weight compounds penetrate completely through wood cell walls including through the middle lamella regions in advance of enzymes [4, 68, 70] which runs contrary to other findings where close association of low molecular weight compounds and enzymes in the white rots is typically reported. Despite reports for more than 20 years of low molecular weight compound penetration of wood cell walls in advance of cellulose and lignin degrading enzymes in white rot fungi, research on these compounds is still quite limited as reviewed below for specific lignin degrading enzymatic systems. A better understanding of the nature of low molecular weight compounds, and which particular enzyme systems they are associated with, would enhance understanding of fungal mechanisms in white rot fungi, and it would also aid future research on utilization of fungal systems in biorefineries and in other bioprocessing efforts.

Selective white rot degradation showing altered wood structure in advance of lignin peroxidase enzymatic penetration in the wood indicating the action of low molecular weight compounds. This type of low-molecular weight radical in the degradation of wood is also associated with manganese peroxidase (MnP), versatile peroxidase, and laccase enzymes [71] in selective white rot degradation. Scale Bar = 1 μm. (Image courtesy of Dr. Geoffrey Daniel)
Lignin Peroxidases
Lignin peroxidase (LiP) is able to oxidize both phenolic and nonphenolic lignin groups; the later which comprises 70–90% of the wood lignin fraction. Lignin peroxidase will oxidize only the exposed components of the wood cell wall at the lumen surface and therefore it is proposed to oxidize smaller intermediates such as veratryl alcohol. The veratryl alcohol radical is smaller and would have the capacity to penetrate into the wood cell wall and may be involved in the gradual erosion of the wood cell from the S3 layer, outward in white rot degradation. The affinity of the radical for nonpolar substrates such as lignin could potentially provide a role for such a radical (assuming it is capable of diffusing from the enzyme) for the selective attack on lignin in selective white rot degradation. Lignin peroxidase’s ability to oxidize nonphenolic lignin has made it an attractive candidate for use in biopulping, dye decolorization, and biorefinery applications, as biorefinery development has grown. Publications discussing the use of LiP including those by Fernández-Fueyo et al. [72], Mäkelä et al. [73] and Pérez-Boada et al. [74] propose that veratryl alcohol may function as a cation radical that can oxidize lignin substrates indirectly, whereas versatile peroxidase (below) is able to oxidize Mn2+ directly allowing direct enzymatic action on substrates. Although they speculate that the cation radical is closely associated with the enzyme, it is possible that lignin monomers functioning as cation radicals could function as low molecular weight agents diffusing ahead of LiP.
Manganese Peroxidases
Manganese peroxidases (MnP) are Class II fungal peroxidases capable of oxidizing Mn2+ to Mn3+. Low redox potential phenolic compounds are able to be oxidized directly through this action. However, mediators such as unsaturated fatty acids or glutathione are required for the degradation of high redox potential nonphenolic compounds. It has been reported that carboxylic acids can also function as a mediator in the degradation of high redox potential substrates [75]. Some aliphatic acids such as malonate, lactate, and oxalate produced by white-rot fungi function as metal chelating agents and increase the oxidation rate of Mn2+ [76] to allow substrate attack. The chelated Mn3+ functions as a low-molecular weight, diffusible redox-mediator that can penetrate the wood cell wall to attack phenolic lignin and produce unstable radical species. As such, this is potentially one of the low molecular weight diffusible species observed by Daniel as cited above; however, diffusion of these radicals from the parent enzyme is typically observed as being quite limited compared, for example, to the diffusion of LMWC agents identified in brown rots. MnP has a lower redox potential than LiP and, under physiological conditions, can only oxidize phenolic lignin structures.
Versatile Peroxidases
Versatile peroxidase (VP) is considered to be a hybrid between LiP and manganese peroxidase [77], and it is an enzyme with broad substrate activity. VPs can directly oxidize nonphenolic substrates such as veratryl alcohol as well as phenolic substrates [72, 73]. VPs are more widely produced in white rot fungi than originally considered given that they were first cloned and sequenced only in the year 2000. They are now known to be produced in many fungi such as Phanaerochate chrysosoporium, Ceriporiopsis subvermispora, Pleurotus ostreatus, P. eryngii, and Bjerkandera sp. [78]. VP’s ability to oxidize phenolic and nonphenolic substrates occurs directly without the aid of a mediator.
Dye Decolorizing Peroxidases
The dye decolorizing peroxidases (DyP) were first reported in 1995 in a Geotrichium (Ascomycota) species, but have recently become more widely reported in white rot fungi. The DyP-type peroxidases have no homology to other known peroxidases. They are unable to oxidize Mn2+, and they are unique enough that they are classified in their own superfamily of heme peroxidases (EC 1.11.1.19). They are similar to VPs in that they can oxidize Reactive Black 5, phenols, and veratryl alcohol, but unlike VPs they also have the ability to oxidize recalcitrant anthraquinone dyes [79, 80].
Laccase
Unlike the peroxidases, the laccase enzyme employs a mechanism involving a 4-electron reduction of oxygen to degrade lignin. Laccase is produced by many microorganisms, including both wood-degrading and nonwood-degrading species, and may play multiple metabolic roles. Laccase alone is not capable of oxidizing nonphenolic lignin compounds but can oxidize “mediator” compounds to their radical forms, and in the presence of laccase, these compounds then catalyze the oxidation of nonphenolic lignin. Natural mediators have been reported including ortho-diphenols, para-diphenols, methoxy-substituted phenols, diamines, and benzenethiols. Since laccase is a robust enzyme, its potential for industrial use is high and many uses have been studied ranging from the grafting of polymers to lignin to the production of fiberboard products [81]. Several lignin-derived compounds [82] and also hydroxamate siderophores from fungi also have been reported as useful in mediating laccase-aided delignification processes [83]. Fungi are generally reported to produce hydroxamic acid siderophores, but research on wood degrading fungi has isolated many catecholate compounds that are “siderophore-like” in their structure. Given that the structure of lignin has more catecholate properties, it would be useful also to assess the catecholate/phenolate compounds similar to siderophores that are produced by wood decaying Basidiomycota species.
The role of low molecular weight compounds produced by fungi should be explored in future research, as understanding the potential activity of these compounds aiding in enzymatic mechanisms is still in its infancy. The role of enzyme mediators and electron donors for the ligninase enzymes as well as the CAZmes such as the Lytic polysaccharide monooxygenases still can be advanced to benefit biorefinery applications.
3.1.3 Soft Rot/Ascomycota Fungi
Soft rot fungal degradation is generally considered to be a surface attack of wood caused primarily by Ascomycota genera, although in some cases the species involved may still informally be classified as Fungi Imperfecti as they may not have yet been examined using molecular systematics. Surface wood hardness and density loss typically occur as fungal attack progresses. Soft rot can occur when the wood is very wet, or even in waterlogged conditions. However, it is now known that soft rot fungal attack can occur in both relatively wet and dry environments, and soft rot attack has also been observed several centimeters deep, particularly in buried wood in some field tests and in-service poles [84], in some cases when the wood is not at saturation levels. Although soft rot typically is observed as a surface phenomenon, researchers have pointed out that in wet environments, oxygen is excluded deep in the wood, and this would explain the lack of growth of soft rot fungi other than at the surface of wood [7]. Soft rot fungi seem to have a more selective requirement for oxygen, and therefore, decay fungi or bacteria that attack wood would potentially outcompete soft rot fungi in many environments. All wood undergoing any type of decay ultimately has reduced density, so to some degree the term soft rot is misnomer. Daniel and Nilsson [85] have also reported that soft rotted wood, particularly in CCA-treated poles, is quite hard at the surface. Upon drying, soft rotted wood typically becomes brown and develops surface checks across the grain as the wood shrinks. Although the decay may be superficial, the surface appearance may be similar to brown rot decay. In advanced stages of soft rot decay, the wood will fail in a brash or brittle manner when a surface sliver is lifted using a knife or sharpened probe – also very similar to brown rotted wood. The wood may assume a weathered appearance, similar to unpainted “barn board” [86]. Several common genera of soft rot include Chaetomium, Humicola, and Lecythophora.
The importance of soft rot decay in North America became more widely recognized in association with severe damage observed in utility poles treated using a preservative treatment known as the Cellon process [87, 88]. Cellon used liquified propane (LP) gas or other volatile solvents as the carrier for pentachlorophenol which was subsequently pressure-infused into the wood and the solvent carrier vaporized leaving a relatively clean wood surface. Chemical analysis indicated that there was adequate bulk retention of pentachlorophenol in the wood, but later analyses showed that the chemical often only passed into the cell lumens without extensive penetration into the wood cell walls. The treatment was also often variable with nonuniform distribution of the chemical resulting in a variable treatment that the fungi could circumvent. Caustic washing of the poles to reduce pentachlorophenol blooming also resulted in the surface leaching of pentachlorophenol contributing to additional treatment variability. Since soft rot fungi have some resistance to pentachlorophenol treatments, they were able to penetrate through the wood cell walls within the S2 layer and therefore attack and reduce the structural integrity of the wood. Over 800,000 utility poles were treated using the Cellon process [89] starting in the 1960s, and many were ultimately degraded in service by soft rot fungi. Prior to this experience, this type of fungus had been known primarily to attack wood in very wet locations such as the wooden slats of industrial cooling towers. Soft rot in North America is less commonly reported as a structurally degrading microorganism compared to brown rot, and it is more widely reported other environments/locations ranging from tropical to temperate and even polar environments [90].
Two different types of attack on the wood cell wall can be produced by soft rot fungi. Type I soft rot is characterized by cavity formation which typically occurs in the S2 layers of the wood cell wall, although these cavities may also be formed in some cases in the S1 layers, often in the same cell. Type II is a general erosion of the wood cell wall layers similar to that seen in white rot, starting from the S3-lumen interface and working outward. Often, particularly in hardwoods, both Type I and Type II attack can be produced by the same fungus in the same sample. Soft rot fungi are sensitive to high levels of lignin and preferentially grow in low-lignin hardwoods such as Populus or Betula. Lignin levels in the wood also impact the pattern of attack which can occur within the wood cell wall [85].
Soft rot fungi typically initiate Type I attack using fine microhyphae, to penetrate from the lumen and bore perpendicularly into the wood cell wall [4, 91, 92]. The fungal microhyphae re-orient and realign in the direction of the S2 cellulose microfibrils when they reach the S2 layer. T-branching of the hyphae also occurs once the hyphae penetrate the wood cell wall. Prior to the initiation of cavity formation, the microhyphae extend longitudinally into the S2 cell wall via development of what is known as a proboscis hypha. The process of enzymatic secretion and cavity formation then is used to produce a series of longitudinally oriented diamond-shaped, or elongated conical cavities in what is known as Type I attack (Fig. 4.7). Multiple hyphae often invade a wood cell wall and can completely riddle the wall in advanced stages of degradation. The conically pointed ends of the cavities are formed presumably because of the way that the extracellularly secreted enzymes interact with the crystalline structure of cellulose in the cell wall.

Transmission electron micrograph of a southern pine wood cross-section. The wood was used as a cooling tower slat and was being decayed in a Type I soft rot attack. The image shows portions of four pine tracheids with fungal hyphae decaying parts of the each of the cell walls (CW) of the tracheids. Cavity formation shows the fungal hyphae growing within the S2 layer of the wood cell walls and producing cavities (white arrows) surrounding individual hyphae (select hyphae are identified with black arrows). The cavities often coalesce in this heavily degraded sample
Soft rot fungi are capable of producing a complete complement of cellulolytic enzymes, including exo-1,4-ß-glucanase, endo-1,4-glucanase, and 1,4-ß-glucosidase [93, 94]. Research using a variety of agar tests shows that a number of cellulase, hemicellulase, pectinases, and laccase enzymes are produced by many of 27 freshwater soft rot ascomycetes tested [95], with several of these species having not been commonly reported in the soft rot literature previously. Earlier work also indicated that laccase was produced by certain soft rot fungi suggesting that there is at least a limited ability to enzymatically [96] degrade wood cell wall lignin. Other research indicates that laccase is the only lignin degrading enzyme produced by at least one soft rot species [97]. Daniel and Nilsson [85] have hypothesized that a radical generating system may also function in conjunction with a system for enzymatically oxidizing lignin, which would suggest laccase mediator involvement. They have also reported that cavity formation may be “flattened” on one side where the fungus encounters more heavily lignified wood that is not as readily degraded as wood that is cellulose-rich. However, Type I soft rot fungi have been observed to lose cavity-formation capability when moving into delignified wood cell walls. This suggests that the orientation of cellulose elementary fibrils, and also the surrounding encapsulating lignin matrix, may play a role in cavity formation.
Type II soft rot attack is more common, with several species reportedly capable of producing Type II soft rot attack alone. Type II soft rot appears similar to white rot at the microscale as the wood cell wall is thinned from the lumen, outward, until only the middle lamella remains [85]. This type of soft rot decay rarely occurs in softwoods; however hardwoods, possessing a combination of syringyl- and guaiacyl-type lignins and generally having lesser amounts of total lignin in the cell walls, are attacked preferentially. Many species of soft rot fungi isolated from wood poles are capable of producing both Type I and Type II soft rot attack with one, Phialophora fastigiata, producing Type II attack followed sporadically by Type I attack with longer exposure in laboratory testing [98].
3.2 Staining and Mold Fungal Degradation of Wood
This section reviews fungi that inhabit the surface of wood or that penetrate into wood but cause only limited amounts of degradation, primarily of the nonstructural wood components. It is important to recognize that beyond fungi, other causes of discoloration and surface coloration of wood and wood products are known, and casual observation, particularly by lay persons, is often not enough to distinguish fungal stain and mold from these other causes. This is highlighted by what has been described as the “largest mold litigation settlement” in North America which was in excess of US $150 million [99]. In this matter, testimony from wood experts was not allowed in a case where a brown to black discoloration had developed on exterior wood that was finished with a clear wood polymer coating, formulated with a high content of iron. A court ruled that the product had been discolored by mold fungi when in fact the iron in the polymeric finish had reacted with extractives in western North American softwood species to cause excessive darkening of the finish (Fig. 4.8). Mold was in fact only a very minor factor in the staining of the wood. Although the cause of the discoloration may not have affected the legal outcome of this particular case, from a wood deterioration perspective, it is important to understand the causal mode so that future problems/judgments of a similar nature can be avoided. In a similar manner, when Mount St. Helens erupted in the US state of Washington in 1980, many standing trees in the path of the hot mud flows created on the slopes of the mountain were damaged. The mud flows were created by the rapid melting of snowpack on the mountain during the eruption. Examination of the trees in the mudflow zones after the eruption revealed that a dark blue-grey stain had developed in the interior sapwood rings of many trees (Fig. 4.9). Initially this blue stain was considered to be a fungal stain until analysis revealed no presence of fungi in the stained wood. However, researchers observed a high level of iron in the interior rings of stained sapwood corresponding with the location of the stain. The iron had been translocated up the interior sapwood via the damaged root system as the trees slowly died after the eruption [100]. These stories reflect the importance of careful analysis of wood to determine whether specific deterioration or defects are caused by biological agents, or by other phenomenon.

A “clear” wood finish that was manufactured using high levels of iron and then used to coat exterior siding made of western North American wood species with typical levels of extractives. Under wet conditions, the iron in the wood finish reacted with the extractives from wood to produce a dark-colored pigmentation. This image was from a structure involved in what has been described as the “largest mold litigation settlement” to date, showing the importance of allowing science to prevail in litigation matters. Mold fungi produced only a limited amount of the dark coloration in the wood finish and on the surface of the wood

“Mount St. Helen’s blue stained wood.” This purple to black stain developed in the interior sapwood of dead and dying Douglas fir trees on the slopes of Mount St. Helens after it erupted in the 1980s. Although it was considered to be a fungal blue stain by local mills, analysis showed that high levels of iron present in the dark zones were responsible for the discoloration
3.2.1 Mold Fungi
Mold fungi typically are classified in the Ascomycota, but where sequencing has not been done and the sexual stage of the mold fungus is unknown, many are still classified as Fungi Imperfecti. Mold fungi inhabit the surface of wood and feed on simple surface sugars and starches or residues that can collect on the surface of wood. Typically, the mycelial growth is hyaline and the spores are pigmented, which discolors the surface of wood resulting in an appearance degrade in finish lumber or other finish products. Most mold fungi grow on a variety of surfaces and they require only surface moisture and a nutrient source for growth. They can grow on inert objects like glass or plastic when those materials are coated with a thin layer of plant or animal residues that contain enough nutrient to support growth of the mold.
When growing on the surface of wood, sugars from subsurface parenchyma will substitute for the sugar residues in other plant or animal residues to permit growth of mold fungi. The presence of mold on structural lumber does not cause any structural damage, and in many areas of the world, structural lumber with mold growth is permitted for commercial sale. For esthetic reasons, and also because of growing concern about mold spores from a human and animal health perspective, it may be desirable to prevent mold from occurring on wood, or remediating any mold growth. Typically mold growth can be remediated by simply brushing pigmented spores from the surface with a stiff brush. Alternately the wood can be planed, or a dilute water solution of sodium hypochlorite or other bleaching solution can be brushed onto the surface to remove the surface spores or hyphal discoloration. Longer term biocidal treatments can also be used if the surface of the wood is anticipated to become rewetted periodically with recurrent mold problems.
As reviewed earlier in the introduction of this section on fungi, some Architectural manuals indicate that fungi, including mold, will grow on wood at only 20% moisture content (MC). Technically this is incorrect as the mold fungi require liquid moisture to survive and sporulate; however, at 20% MC with the cycling of temperatures, water can condense on the surface of wood to permit mold growth. In localized zones of the wood surface, liquid water in the surface capillaries would occur permitting mold growth.
3.2.2 Staining of Wood by Fungi
As noted in the section on soft rot fungi, some stain fungi can intergrade with, and cause, soft rot under certain conditions. Some stain fungi can also intergrade with mold fungi and in early stages of stain fungal growth, when these fungi may grow just on the surface of wood and produce spores, they may be considered to be mold fungi. However, true stain fungi have the ability to penetrate into wood to seek out nutrients such as sugars, starches, and also wood resins and waxes [73] in the parenchyma cells. Some stain fungi also have the ability to remove the more amorphous polysaccharide components such as pectins and hemicellulose. Some stain fungi also produce fine bore holes in wood (Fig. 4.10) although these are smaller diameter bore holes than are seen with wood decay Basidiomycota fungi, and the hyphae are constricted in size as they penetrate the wood cell wall. The penetration of the hyphae may be in part via mechanical action as opposed to enzymatic/catalytic degradation of the wall to produce the bore holes. Daniel points out that wood species or types that are less lignified, such as tension wood in hardwoods, can be more readily attacked by stain fungi that intergrade into soft rot fungi, and hemicellulose is also more readily removed by stain fungi in this type of wood [101, 102].

Fine bore holes produced by blue stain fungi in pine sapwood tracheids. The bore holes (circled in yellow) are much smaller than those produced by Basidiomycota decay fungi, and the fungal hyphae producing the bore holes are constricted in size
Genomic information has separated and reclassified some of the more commonly established wood staining fungi, with fungal genera like Ceratocystis and Ophiostoma being split with new names [103, 104]. Some of the same genera names have continued, with common genera that now include Ceratocystiopsis, Ophiostoma, and Grosmannia. In species that produce a sexual stage, these fungi produce dark colored, flask-shaped, spore-bearing structures known as perithecia that grow on the surface of wood. Spore-containing asci are produced in the base of the perithecia, and the asci and spores are then exuded through the neck of the perithecia for dissemination. Typically, the spore mass is discharged in a moist viscous fluid droplet which rests at the top of the opening in the perithecia neck (Fig. 4.11). Although spores can be dispersed by wind and rain water, many species of stain fungi that produce perithecia have adapted to be disseminated by beetles. The height of the perithecia neck for particular fungal species has evolved to match the height and anatomy of particular beetles or other insects and arachnids (particularly mites). The viscous fluid mass of spores is often described as a sticky slime, which adheres to the body or legs of the insects/arachnids as they feed upon the fungal mycelium and spores. The spores are then disseminated as the vector travels to other sites. Uzunovic et al. [105] provide an excellent review of wood staining by fungi and other agents and report that log harvesting equipment can be a major factor in fungal stain dissemination. Spores will germinate on suitable wood surfaces, but the wood must be at or above the fiber saturation point, at least in localized surface areas, for stain fungi to grow and penetrate into wood.

Blue stain in early stages of development on pine wood. Coloration of the hyphae is developing in the sapwood, but not the heartwood. Similarly, perithecial fruiting body development also is present on the surface of the sapwood and the hyaline/clear spore mass droplets are being exuded at the tip of the perithecia. At the right, the spore droplets have coalesced to form a larger pool of spore exudate
Because stain fungi must first gain a foothold in wood by accessing simple monomeric or oligo sugar substrates, they typically do not grow on or penetrate deeply into heartwood because simple sugars and starches have been depleted and fungistatic or fungitoxic phenolic extractives are present instead. For this reason, these fungi are also sometimes known as sapstain fungi. As the fungal stain hyphae mature in wood, they produce melanin in their cell walls. The melanin is thought to be useful in protection of the fungal hyphae from radical damage from sunlight, and the melanization also occurs in some sporophores, presumably to also protect against sunlight. These fungi may cause the wood to appear many different shades of color, and in some cases, the colors can appear blue, green, yellow, red, or other shades of the spectrum. Blue staining is perhaps most noticeable, and it is caused when fungal hyphae grow in the parenchyma, and especially in the ray parenchyma just under the surface of the wood. As the fungal hyphae melanize, they take on a dark brown or black coloration. When reflected through the crystalline layers of cellulose in the parenchyma, the brown/black coloration is diffracted and it can appear different colors, most often appearing as blue color to the eye. This is an optical illusion caused by the scattering of the reflected light, much in the way that dark soot particles in smoke can appear blue, or other colors, as they scatter and reflect light. Blue or a dark-grey or black coloration is common in stained wood, and this is often described as blue-stained wood.
Stain discoloration can occur under favorable conditions in as little as 3 days in lumber. Some softwood genera with wide sapwood bands and large resin canals such as pines (Pinus sp.) will stain more readily than wood species that have more heartwood and smaller or no resin canals such as spruce (Picea sp.) or fir (Abies sp.). Staining typically occurs at temperatures above 15 °C, and moving wood to a cold environment to arrest stain development is a potential remediation method in some cases. However, staining at lower temperatures has been reported in some areas where the fungi have apparently adapted to these temperatures [106]. Prevention of stain development can also be achieved by keeping logs wet to exclude oxygen needed by the fungi for growth. In mill yards, water sprinklers placed above or on top of log decks are sometimes used for this purpose when temperatures permit stain fungi to be active. Biocidal treatments to protect against wood staining can also be used, although rapid processing to get logs sawn and the lumber then dried to 19% moisture content or less is preferred from both and environmental and expense standpoint whenever possible.
Fungal staining in structural lumber and wood products is allowed in many countries as no change in modulus of rupture or modulus of elasticity occurs in this wood. However, some wood products specifically exclude fungal stained wood. Also, international shipments of wood with fungal stain (and mold) have been limited in some cases [107, 108]. High temperature kiln drying of wood can kill the fungi and for some countries this is a permissible solution if international shipment of wood with fungal stain is a concern. Fungal stain is also a concern in certain wood products that may undergo shock loading as is the case in ladder stock where a misplaced foot may result in a rapid loading (shock loading) of the step below. The removal of hemicellulose in heavily stained wood causes loss of toughness or shock resistance by as much as 15–30% [109], and this wood is not allowed by code in structural applications like ladders that may experience shock loading or that require toughness.
4 Bacteria
The bacteria are a separate domain or branch of life that is distinctly different from the other microorganisms discussed in this chapter. They are prokaryotes, and by their nature they are unicellular, although bacteria can form stable communities leading some researchers to think of them as multicellular communities. Although some of the enzymes and metabolites secreted by bacteria have similarities to those produced by the higher eukaryotic organisms, the degradation of wood solely by bacteria is considered to be a relatively slow process in comparison to degradation by decay fungi, insects, or marine organisms. Bacteria are important symbionts in the degradation of wood by some insects and marine borers, and those symbioses are discussed in the sections on those organisms in this chapter. There are also commensal and antagonistic effects between bacterial and fungi, and this is reviewed below.
Bacterial attack of wood is most often observed in wood or woody debris that has been submerged or buried. Under these conditions, aerobic bacterial attack at low oxygen levels occurs initially on nonlignified or partially lignified tissues including parenchyma and pit membranes in the sapwood. Heartwood that contains extractives is often not aggressively attacked, similarly to attack patterns by fungi. This is in part because of the toxicity of heartwood extractives to microorganism, but also because of the reduced content of starch and sugars in the parenchyma cells of the heartwood being a less rich nutrient source. Daniel [4, 68], Singh and others [110] have observed that bacterial attack increases the permeability of wood because the degradation of parenchyma provides avenues for capillary water uptake. The destruction of both bordered pit membranes and simple pits in the parenchyma opens up channels for water penetration in the tracheids/fibers and rays, respectively.
Bacterial attack of wood can occur in wet aerobic environments, but also in some species when near anaerobic conditions prevail, particularly when wood is wet or submerged. The degradation of wood under these conditions is quite slow and, as noted by Daniel, bacteria found in wood are those “having a tendency” to be aerobic or facultative aerobes [68]. Wood degrading bacteria are primarily gram-negative species. As degradation progresses and bacteria become established in wood, they detoxify, degrade, and metabolize heartwood extractives. With long-term exposure, bacteria significantly degrade heavily lignified tissues and complete degradation of lignocellulose cell walls occurs. Some resistant bacterial species also have the ability to detoxify and metabolize preservative treatment chemicals such as creosote. In addition, research has demonstrated the ability of some bacteria to tolerate wood treated with metals, such as chromated copper arsenate (CCA) and copper-treated wood. In some cases, the bacteria seem to solubilize and then precipitate or compartmentalize some of the metals in their extracellular biofilm matrix as a means of detoxification so that they can then access the nutrients in wood.
Despite the potential importance of fungal-bacterial interactions within wood during degradation, there are relatively few reports on fungal-bacterial commensalism in nature [111]. Fewer still are reports of fungal-bacterial interactions within wood undergoing degradation in the built environment. However, in wet environments, it is common to observe erosion bacteria (below) in association with soft rot fungi in structural members undergoing degradation. In nature, there is an increase in bacterial diversity as fungal decay progresses, with sapwood having a great bacterial abundance and heartwood bacterial communities becoming more diverse. Wood in forest environments hosts nitrogen fixing bacteria that can provide nitrogen to colonizing fungi, and nitrogen fixation increases as decay advances. Bacteria have been observed to migrate across soil with the hyphal growth of wood decay species. Fungi can also have a negative effect on bacterial colonization of wood, with inoculation of stumps in forest environments with the fungus Phlebiopsis inhibiting bacterial growth in the same wood. Conversely, microbial antagonism has been used to beneficial effect in wood processing industries to protect against both stain and decay fungal invasion of freshly felled timber. Just as fungal antagonism with the fungus Trichoderma has been used to protect against fungal decay initiation, some bacteria produce inhibitory substances and antibiotics which can prevent fungal growth in wood. The use of Streptomyces sp. to protect wood against fungal degradation has been patented [112].
Apart from symbiotic or commensal relationships with bacteria, bacterial attack of wood has been classified into two forms: tunneling bacterial attack and erosion bacterial attack. Both of these classifications refer to the level of attack which occurs at the micron and nano-scale and, as such, it is most readily observed using electron microscopy. Bacterial attack has significant implications relative to the degradation of lignocellulose in forested environments [113]; however, this attack takes far longer than fungal degradation.
The tunneling bacteria (TB) cavities are produced by pleomorphic bacteria 1–2 μm long that typically fix to the surface of the wood cell lumen via their extracellular biofilm matrix (ECM), and then bore holes that radiate and branch as the bacterium divides to penetrate the wood cell wall [4, 68]. Tunnels are not oriented with the microfibril angle of the wood cell wall. TB have the capacity to bore through the entirety of the wood cell wall and even into the lignin-rich middle lamella. As TB form cavities, they slough off outer layers of bacterial cell wall and EMC, leaving these deposited as a trail of concentric arcs within the tunnel. Tunneling bacteria in most cases appear to require oxygen for their activity [110] and are not active in submerged or deep burial sites.
Attack of wood by erosion bacteria (EB) is arguably more economically important and more widespread compared to attack by TB. EB can tolerate sites with lower oxygen availability [110] than TB, and although they can also degrade wood in aerobic environments, they are active degraders of submerged wood. EB typically initiate attack of the wood from the lumen surface or the bordered pit region. The rod-shaped bacteria will align to form parallel erosion troughs, which deepen over time to degrade the wood (Fig. 4.12). Cavities can take on an angular appearance resembling Type I soft rot fungal cavities to some extent. Although the wood cell wall layers are degraded, the lignin-rich middle lamella typically is not attacked [4]. In wood submerged for more than 300 years such as found in archeological excavations, EB attack predominates as the primary type of biodegradation. Degradation of foundation piling by EB is also an important issue, although the degradation process is quite slow. However, over several hundred years, extensive attack can develop to initiate the loss of structural properties. For many years, it was assumed by marine archaeologists that submerged wooden relics could be excavated from the mud on the floor of the ocean or rivers for observation, and after study the materials could be re-buried to continue long-term preservation of the wooden objects. It is known that submerged samples above the mudline that receive enough oxygen degrade slowly by erosion bacteria [114, 115] but buried material without oxygen is often well preserved. It is now understood however that disturbance of the mineral layers surrounding buried objects, with concurrent introduction of freshly oxygenated water, promotes the active growth of EB and also soft rot fungi. This increases the rate of wood degradation in reburied wooden objects for a period of years until oxygen and nutrient levels in the areas immediately surrounding the wood are depleted [116]. Re-burial of wooden artifacts is therefore likely to permit enhanced deterioration for a time, at least until oxygen levels can be depleted by the organisms.

Aligned erosion bacterial degrading wood taken from a submerged site. (Image courtesy of Dr. Geoffrey Daniel)
5 Insect Damage to Wood
Invertebrates , and particularly insects in terrestrial environments, can play a major role in wood decomposition in the environment [117]. However, even under ideal infestation conditions, their presence is not guaranteed because they are patchily distributed both in time and geographically. When present, they can have major consequences on the trajectory and timing of wood deterioration through their feeding activities including comminution and the introduction of microbes. Four ways that insects and other invertebrates facilitate wood degradation are: fragmentation and related physical comminution, enzymatic digestion, biotic interactions, for example, ants bringing foraged food back to their nest, and nitrogen fertilization (including N2 fixation) [117]. Such interactions promote the deconstruction of wood and other lignocellulose substrates, and in some cases, the actions of insects accelerate wood decomposition initiated by microorganisms or other animals.
Tunneling activities of insects increase the aeration of wood, often facilitating gas exchange. This fragmentation greatly increases the surface area exposed to the elements and typically also permits entry of microbial inoculum such as spores or propagules from fungi and bacteria. Interestingly, van der Wal et al. [118] found that wood blocks decay faster than the sawdust/shavings produced by such comminution and the authors reasoned that sawdust is a much better substrate for bacteria than it is for wood decay fungi. However, this is more dependent on the culture conditions as the wood samples were buried or placed on top of soil in this work, and these conditions are known to be less favorable for fungal growth [119]. Such openings can also permit penetration deep into wood by rhizophores (cord fungi) in natural ecosystems; however, these are less likely to be present in structural environments.
Five groups of insects are principally involved in wood deconstruction. Some, such as bark beetles, will attack weakened trees, but in this chapter insects such as bark beetles are discussed only peripherally as the chapter focus is on insects that attack wood in structural environments. The insect groups focused on in this chapter section are: Termites -subterranean (principally Rhinotermitidae), -dampwood (Termopsidae), and -drywood (Kalotermitidae), Powderpost beetles (Ptinidae, subfamily Anobiinae; and Bostrichidae, subfamilies Lyctinae and Bostrichinae), Cerambycid wood boring beetles (Cerambycidae), Carpenter ants (Camponotus), and Carpenter bees (Xylocopidae). Globally, the largest group, causing the most damage in attacking wood in service, are the termites [117]. Global damage estimates for termite activity are variable, insufficiently documented, and are largely underestimated [120], but a single study estimates that global damage by the Formosan termite (Coptotermes) alone exceeds 26.7 billion euros per year [120]. In the United States, termites cause billions of dollars in damage to structures annually [121]. On a worldwide basis, based on the author’s experience, the next most important groups in damaging structural wood products are the carpenter ants, followed by the wood boring beetles. Damage by carpenter ants is greater in the higher latitudes, while termite and beetle attack is typically more prominent in warmer temperate to subtropical and tropical zones. Typically, carpenter bees (Xylocopidae) mine wood trim and finish boards and are considered much less consequential; however, they are discussed because of the severe damage they can cause to high-value or historic structures.
All insect taxa discussed in this chapter degrade wood either mechanically (fragmentation by chewing) or through mechanical action combined with enzymatic action as part of digestion. Beetles and termites exhibit both modes, while carpenter ants and carpenter bees only chew/mine and remove wood and eject sawdust-like frass. They also vary in their association/relations with internal and external microorganisms. Lignocellulose degrading endo- and ectosymbionts occurs in beetles and termites; however, carpenter ants and carpenter bees largely appear to lack microbial associations. The effects of these insects on assemblages of wood decomposers (ectosymbionts and others) are not well known at the present time and must be further studied. While some beetles and termites appear to benefit from the presence of lignocellulose degrading fungi, others may lack demonstrable effects. In most cases, ants and bees scrupulously eliminate and inhibit/retard the growth of microbes; however, in nonstructural environments, ant-fungal and ant-microbial mutualism or symbiosis is well described [122]. However, in structural environments, carpenter ants and carpenter bees, like many Hymenopterans, are fastidious housekeepers and secrete substances that can halt microbial growth within their nest galleries. Some though can also engage in symbiotic relationships to foster the growth of beneficial microorganisms that can battle insect pathogens [123].
Although not a focus of this chapter, readers should be aware that some insects that do not directly attack structural wood have the potential to affect structural degradation by other insects or agents. As an example, although not a species that attacks structural wood, ants in the genus Aphaenogaster nest in woody debris in the southeastern United States and change the course of its decomposition by preying on termites and secreting substances that limit the growth of some microbes. This in turn limits the termite population in the local environment which would potentially then reduce the numbers of termites capable of attacking structural wood also in the same environment.
5.1 Termites
Termites are social insects that live in large, complex colonies containing thousands to millions of individuals. Worker termites are responsible for the damage done to wood. Their fragile appearing, creamy-white, and thinly waxed exoskeletons belie their mandibles destructive power. They comprise five families with the center for their species diversity and abundance being pan-tropical (subtropical and tropical). About 2700 described species feed on wood, and perhaps as many as 4000 species of termite exist, but most of these are assumed to be humus or soil-feeders. Approximately 1800 species of termites are wood-feeding lower termites. There is at least one termite species for all terrestrial environments between 45 north latitude and 45 south latitude, covering some 2/3 of the Earth’s terrestrial environments [124]. The greatest species richness is in lowland wet and dry tropical forests, but termites are also common in savannas, including tropical/subtropical grasslands. The savanna termites are largely soil/detritus feeders and they do not attack structural wood. In the northern hemisphere, termite distribution is primarily within North America and Asia; however, they are present in limited numbers in Europe, north of southern France, and the Mediterranean region. Termite migration into northern France and other higher latitude regions, particularly in regions closer to the coast, is however occurring. Termites are more widely distributed in the southern hemisphere, particularly Africa and South America. With increasing human activity and increasing temperatures at higher latitudes, termite populations have gradually increased their range in colder climate regions. Species that nest and feed inside individual pieces of wood (drywood and dampwood termites) are the most widely distributed [125]. Their ability to invade floating woody debris (flotsam) and, for some drywood species, live inside flooring and furniture, means genera of these termites have been widely distributed by humankind, and these insects can now be found on all continents – even distant oceanic islands.
5.1.1 Termite Classification
Dictyoptera is a revised superorder containing cockroaches (Order Blattodea), termites (Order Blattodea – Epifamily Termitoidae), and mantises (Mantodea). Hindgut flagellates which aid in cellulose digestion link the lower termites to the relictual wood-eating cockroaches (Cryptocercus). Lower termites comprise 5–7 families worldwide with the Rhinotermitidae (subterranean) and Kalotermitidae (drywood) families probably the most economically important [126, 127]. In the most derived clade of termites (Termitidae – higher termites), protozoan symbionts are lost and diversified feeding types, including fungal farming, humus/soil feeding, leaf-litter feeding, or an epiphytic lifestyle, are utilized [128]. Among the Termitidae, from an evolutionary perspective, wood-feeding has arisen several times among the ground-dwelling humus feeders [129]. Nasutitermes termite species can attack structural hardwood timber in tropical regions, particularly in the southern hemisphere. Although these are typically tree-dwelling termites, they can attack hardwood sapwood. Coniferous species are generally considered to be resistant to Nasutitermes. Most soil-detritus feeding termites do not typically damage structural wood in-service.
Generally, relative to wood deterioration, the termites are separated into the nonphylogenic categories of subterranean termites, drywood termites and dampwood termites. The first two of these termites are important in structural wood degradation, but the dampwood termite is not, so it is only briefly reviewed here. Dampwood termites are a unique primitive type of termite in the family Termopsidae. They have the most restricted distribution – limited to wet temperate forests, and they typically have very limited capacity to attack in-service structural wood. Eggleton and Tayasu [125] attribute the distribution of dampwood termites to flotsam (floating pieces of wood) dispersing the three subfamilies of this type of termite.
Approximately, three-quarters of all termite species are higher/soil-feeding termites. They are very important from an ecological perspective, and these termites generally either feed on humic material in the soil or they farm Basidiomycota fungi (Termitomyces). These higher termites comprise the greatest amount of insect biomass on the planet. However, the less-species rich and abundant subtropical and temperate, lower termite species account for most primary large woody debris deterioration, including structural wood degradation. These lower termites are commonly known as subterranean termites and drywood termites for the types of wood they attack, and this terminology will be used in this chapter. Drywood termites are by far, less economically important compared to Subterranean termites, although drywood termites can be very destructive in certain situations where they are found.
5.1.1.1 Subterranean Termites
Subterranean termites, including mound and arboreal species, comprise most (80%) of the economically important species of termites. They are the most widely distributed termite, extending into temperate regions and causing the most damage to structural wood of all termite types [130]. Of the 2300 known species, 183 damage structures with 83 species causing what Su and Scheffrahn [130] call “significant damage.” Termites also feed on living trees (e.g., Coptotermes formosanus), but it is unclear how much living vs. damaged/dead tissue is consumed [121]. C. formosanus, the “Formosan termite,” is one of the most destructive subterranean termites, and it alone has been reported to cause billions in dollars of damage in cities such as New Orleans, in North America, and in other locations around the world.
Subterranean termites attack wood in contact with soil or other moisture sources as this contact allows the wood to remain damp or wet, and the moist environment is required for the termite’s growth and reproduction. Reticulitermes spp. is one of the most common subterranean termite genera found in the world, and these termites forage for wood on, or in, the soil or soil litter. Controlling the amount of wood debris buried in the soil around structures is an important way to limit termite activity. The termites also make mud shelter tubes, extending from a water source (typically the soil) to wood, to utilize wood that is not in contact with soil or water. These shelter tubes protect them from desiccation and predation as they travel, and can also help in limiting moisture loss from the termite. This in turn helps to increase the moisture content of the attacked wood. Subterranean termites preferentially consume the lower density earlywood in softwoods, leaving behind the higher density latewood (Fig. 4.13a). In hardwoods, lower density wood is also preferred but attack patterns will vary with different species. Sapwood is preferred to heartwood but both can be attacked and usually softwoods are selected over hardwoods [121] when an option is available. The moisture content of wood undergoing attack by subterranean termites is typically in the range of 28–32%. Termites will bring water into their galleries to moisten the wood, and if the moisture content is below 28–32%, they will transport water in to insure the wood reaches these levels. They defecate watery feces when disturbed or for use when building shelter tubes.

(a) Subterranean termite damage to a “2x4” structural wood member. Note the characteristic “muddy” appearance of the gallery which resembles dried oatmeal and results from termites packing fecal material and soil particles into galleries. Subterranean termites feed with the grain of wood – removing earlywood xylem tissue, leaving behind denser latewood. (b) Subterranean termite damage. (Note the laminated appearance of the wood resulting from consumption of softer earlywood, leaving the denser latewood untouched)
Potter [121] is quick to point out that no wood species is safe from the ravages of these insects; however, wood that is treated adequately with biocidal wood preservatives or that is naturally decay resistant, can be protected. Subterranean termites fill much of their galleries with soil/mud and fecal material. This gives the walls of their galleries a characteristic muddy/dried-oatmeal-coated appearance (Fig. 4.13b). Wood undergoing fungal decay is highly attractive to, and preferred by, many species of termites [121]. The reproductive caste of eastern subterranean termites (R. flavipes) may only develop optimally when fed decayed wood [131]. Conversely, because of the increased porosity, termite-attacked wood can promote moisture penetration in some cases, and fungi can establish in termite-attacked wood if that wood becomes wetted to the fiber saturation point.
Estimates vary on how much wood subterranean termites destroy. Bradford et al. [132] found Reticulitermes spp. to be significant contributors to the mass loss of wood blocks on soils. Large subterranean termite colonies may destroy up to 28 g of structural wood per day, and Formosan termites are reported to have devoured the framing in one wall of a building in 3 months [121]. A large subterranean termite colony can consume an amount of biomass equivalent to that consumed by a grazing animal of similar weight [133]. Millions of worker termites could potentially weigh 4–15 kg, comparable to a large rabbit, hare, or small deer. Grace [133] estimated that a R. flavipes colony of this size could consume 0.5 kg of wood each day. Formosan termites can destroy twice this amount, daily. Researchers have measured wood consumption rates for groups of worker termites in the laboratory. For example, Haverty et al. [134] found that 1000 workers could destroy 83 mg of lignocellulose in a day. Field experiments using over 60 colonies of Reticulitermes termites, each containing somewhat less than 100,000 workers, showed that these termites were able to eat the equivalent of 5 inches of a “2x4” annually for each of the 3 years of the experiment [135]. Similar analyses showed that Reticulitermes colonies removed 3–20% of wood mass within 9 months. Potter [121] cautions that values such as these are variable because of the nature of estimating termite colony size. Even at a particular location, and within a species, there is great variability. Within structures, termites may not attack framing or other structural members, while “for no apparent reason” they will attack adjacent trim material. This may be attributed to multiple food sources, or the lower density of typical trim and molding woods. Termites relentlessly forage for food even after finding and securing large “deposits” of wood. Many biotic and abiotic factors influence consumption rates. Therefore, such study should only be used as a comparative measure for the “destructive potential” of the particular species being studied.
As related to the destruction of woody biomass by subterranean termites in forested environments: As logs or other woody debris decompose and are colonized by microorganisms and insects, nitrogen fixation rates peak during the channelization or tunneling process [136]. Among wood destroying insects, nitrogen fixation was first detected in the guts of termites [124, 137] with the finding that the C:N ratio is reduced when wood is fed upon by termites [138]. Some of this essential nutrient is absorbed by the termite’s gut, but the remainder is excreted in their fecal pellets. Some estimate that Reticulitermes spp. colonies can fix several grams of nitrogen per log per year [139]. In this way, these termites carry out what Ulyshen [117] referred to as nitrogen fertilization. Although it has not been reported in the literature, this accumulation of nitrogen would also occur in termite-attacked wood in structural environments. The increased amounts of nitrogen would enhance the growth of other organisms, including fungi, which could accelerate the rate of deterioration of termite-attacked wood in structural environments.
5.1.1.2 Drywood Termites
As reviewed above, drywood termites are far less destructive overall than their subterranean cousins. However, in certain environments, they can cause considerable damage because their colonization and tunneling can go unnoticed, often for years. Their cryptic nature is in part due to the fact they are able to inhabit wood that is much lower in moisture content than the subterranean termites. For example, following mating flights, drywood termites can establish in infrequently visited attic spaces that typically are quite dry. Although their preferred wood moisture content is higher (10%), unlike subterranean termites they have been reported to have the ability to attack and remain active in wood that is at 2.5–3% MC.
A unique feature of drywood termites is that they are able to remove water from wood that is being digested. As the digested wood is excreted as frass, it is compressed by specialized anal pads of the drywood termite to press out any remaining water. The excreted frass takes on a 6-sided angular appearance which is the impression of the anal pads, visible with a hand lens (Fig. 4.14). The distinct appearance and feel of the frass pellets produced is diagnostic for the presence of a drywood termite infestation. Often the only evidence of drywood termite attack is the presence of a pile of angular pellets which have been pushed out, or fallen out, of a gallery. The angular frass feels “gritty” when rolled between the fingers, as opposed to the frass or wood particles ejected from the galleries of some beetles or ants, which has a smoother feel. In contrast to subterranean termites, drywood termites readily feed across the grain of wood, and they do not coat their gallery walls with soil particles or feces as subterranean termite species do. By ejecting their fecal pellets during the boring process, their galleries appear quite open and “airy” which can also be a diagnostic feature to distinguish their attack from that of subterranean termites.

Drywood termite frass showing the angular nature of the frass pellets. As an aid in field diagnostics, the ejected pellets feel “gritty” when rubbed between the fingers because of this angular characteristic. Scale bar = 1 mm
5.1.2 Social Structure and Life Cycle
Termites typically have a highly organized social structure with queens, workers, soldiers, and alates (reproductives). Understanding of both their social habits and life cycle can be important in understanding wood deterioration by termites and also control of these insects. Queens lay eggs from which emerge larvae (first instar nymphs) and the larvae grow through a series of molts to become nymphs which can produce any of the adult forms, depending on diet and pheromones: workers, soldiers, or reproductive. There is also an age-based division of labor in insect societies. Young nymphs care for eggs and the king and queen, taking on higher risk tasks as they mature. Colonies do not produce alates until they are 5–7 years of age. Most termites reproduce by swarming – the production of winged male and females. In temperate climates, this usually occurs in the spring or summer. In all areas of the world, swarming will occur after warm, humid periods, or rain. Most subterranean termites will swarm with mating flights during the day, but the Formosan subterranean termite has its mating flights in the evening or night hours. The drywood termite has night mating flights. Many mating pairs perish in this process as they are weak flyers and often land in locations unsuitable for survival or nesting. After mating, the insects pair up: one male (the king) and one female (the queen), and if suitable habitat can be found, they build a nest chamber in soil or wood and begin the process anew. Colonies can be initiated in cracks in bark, between bark and wood, and in similar locations.
5.1.3 Foraging and Gallery Making
All termites forage for woody or cellulose-containing (cellulosic) debris. The lower termites use microbial symbionts, principally flagellates and bacteria, in their hindguts to digest these substances: extracting nutrients, water, and energy from cell content remnants (starch and sugars), cellulose and hemicellulose. As a result, their fecal pellets are comprised almost entirely of lignin. Wood moisture content required to support termites varies by species, with drywood termites typically requiring the least moisture (10–13% MC) and dampwood termites the most (from 35 to 50% MC). Subterranean termites (Rhinotermitidae) are intermediate to these and typically prefer wood at a moisture content of 28–32%. Drywood termites eject very dry fecal pellets from their galleries, while dampwood and subterranean species pack their feces into the galleries which may well help maintain wood moisture and reduce air flow [121]. However, subterranean and dampwood termites do not extract water from their feces the way drywood termites do.
Clearly termites are unique among all insects capable of decomposing wood. They not only destroy more lignocellulose than all other arthropods, perhaps combined, and open channels for others to invade, but they alter the chemistry and nutrient status of the wood itself. The fixation of atmospheric nitrogen by their symbionts may contribute to an increase in rates of degradation and promote the activity of other degradation agents, either concurrent with or after termite attack.
5.2 Wood-Boring Beetles
Many beetles are known to attack trees and are present in felled timber; however, this chapter section will focus primarily on beetles that attack seasoned (processed and dried) wood and are capable of re-attacking that wood. Classification of the beetles has changed from prior decades, and the most important of these beetles are known commonly as powderpost beetles in the families Ptinidae (subfamily Anobiinae) and Bostrichidae (subfamilies Lyctinae and Bostrichinae). The Anobiinae are also known by the common name “death watch beetles.” Bostrichinae are also often called false powderpost beetles or horned powderpost beetles. The Cerambycidae species, Hylotrupes bajulus, known commonly as the Old House Borer is the most important cerambycid attacking structural wood worldwide [140]. Other beetles that are less frequently encountered except in localized regions include the Golden Buprestid which can be destructive, but rarely re-infests. The only other beetle commonly observed in heavily deteriorated wood is the Wharf Borer, which does little damage because the wood it attacks is typically already heavily degraded. This chapter section will focus primarily on the powder post beetles because of their destructive nature.
5.2.1 Powder Post Beetles
Powderpost beetles are the most economically important wood destroying beetles [141], and they get their name from the powder-like frass that sifts out of the exit holes produced by the larvae. Two families and three subfamilies are considered: The Bostrichidae family includes both the Lyctinae and Bostrichinae subfamilies; and the Ptinidae family includes the Anobiinae subfamily. Of these, the Bostrichinae (bostrichid beetles) are considered by the common name of the false powderpost beetles in North America because of their larger diameter exit hole and coarser frass texture. In other parts of the world and in other languages, other terms such as the horned powderpost beetle may be used. As such, we consider the bostrichid beetles in the powderpost beetle category because of their ability to re-infest seasoned wood and the fact that they are now placed in the same superfamily with the Lyctinae. The Lyctinae (lyctid beetles – known commonly as the true powderpost beetles) produce fine talcum-like powder (Fig. 4.15). Both the Anobiinae (anobiid beetle – known also in common parlance as the deathwatch beetle) and the bostrichid beetles produce coarse or gritty-feeling powder when the frass is rubbed between the fingers (Table 1).

Powderpost beetle damage by the genus Lyctidae: (a) A lyctid beetle resting in an exposed pupal chamber, after maturation from the larvae into an adult, and just prior to what would have been the emergence flight. (b) A dead adult lyctid beetle on the surface of wood debris. (c) The appearance of infested wood after powderpost (lyctid) beetle larvae has bored through the wood and the adults have emerged through the flight hole. The upper half of the panel has been surfaced with a razor to reveal the galleries of the larvae below the surface of the wood. Comminuted wood powder produced by the larvae within those galleries is also present on the surface of the wood
The adult powderpost beetle will lay its eggs in pores or cracks of wood, with each beetle genus having preference for a particular size of pores or cracks in either hardwoods or softwoods. After the eggs hatch, the larvae will bore through the wood seeking sugars and starches in the wood parenchyma cells, their primary food, before metamorphosing into an adult just beneath the surface of the wood. The Anobiinae are also able to metabolize hemicelluloses in the cell wall [142]. After maturation, the adult will bore an emergence hole to fly from the wood and initiate the life cycle again. Differentiation of powderpost beetle species can be done by analysis of these emergence, or flight holes, and also the texture of the frass produced as overviewed in Table 1.
The sapwood of hardwood species is attacked by the Lyctinae subfamily. In most cases, the Bostrichinae subfamily attacks hardwood, but softwood attack has also been observed in rare cases. The sapwood of hardwoods and softwoods are attacked by the Anobiinae subfamily (Table 1). The larval stage is responsible for almost all of the wood damage, and it is this immature stage of powderpost beetles that feed on wood to cause the damage [143]. In warmer climates, the wood-boring beetles are second only to termites in lignocellulose destruction by insects. Powderpost beetle larvae are highly adapted for xylophagy with highly specialized jaws and hairs to anchor them to their galleries. Because starches are depleted in the heartwood of most wood species, heartwood attack is typically more limited particularly in wood with high levels of heartwood extractives. Zhong and Schowalter [144] found that just 0.6% of the wood volume was bored and ingested by these insects; however, they note that assessment of wood consumption can be instrumental in determining decomposition rates and trajectories. Most anobiid species attack wood, and with the aid of yeast symbionts are able to digest cellulose [145].
To re-infest wood, powderpost beetles also require that adequate levels of moisture be present in that wood. Anobiid beetles can attack wood as low as 15% MC and are often found in unheated wooden structures where the wood moisture content can be elevated to this level. The optimum for their growth and development, however, is 30% [140]. Saunders [140] refers to the true powderpost beetles (Lyctinae subfamily) as the most important destructive agents of hardwoods. He reports that although they can attack woods of 8–32% MC, their greatest activity and development is in the range of 10–20% moisture. Control of powderpost beetles in heated structures can be achieved simply by heating to drive the moisture content down to an ambient 7–8% to prevent re-infestation. In unheated structures, control over moisture content may be more difficult, and if the wood is unfinished (uncoated), then biocides or other methods may be required for control. However, because powderpost beetle require pores of specific dimensions for oviposition, simply coating the surface of the wood with a paint or coating will prevent re-infestation in most cases.
5.2.2 Cerambycid Beetles
The old house borer (Hylotrupes bajulus) is the most important Cerambycid attacking wood in structural environments worldwide [140]. A European species originally, it has gradually spread throughout the world. It was introduced to the USA from Europe and occasionally attacks the sapwood of North American species. This beetle requires a high moisture content, 28%, near or above the fiber saturation point, for optimal growth and spends 2–10 years in its larval stage [140, 146]. Its name is derived from this long larval period with emergence of the adults in homes noted 10+ years after initial construction. It can survive at moisture levels between 15% and 25% [140]. The lower Cerambycids produce their own cellulases, but survive and grow better on wood infested with decay fungi [136]. Like the Anobiinae beetles, some Cerambycids also possess yeast symbionts [147].
Damage to wood and structures by beetles can be significant with less damage occurring in the higher latitudes and colder climates. Wood removed by Coleopterans can have structural (support/engineering) consequences and they can also alter the trajectory of wood decomposition processes. Like termites, some of these can fix nitrogen and have a fertilizing effect which could impact other deterioration processes in structural environments.
5.3 Carpenter Ants and Carpenter Bees
Carpenter ants (Camponotus spp.) and carpenter bees (principally family Xylocopidae) are both members of the insect order, Hymenoptera. Both groups (taxa) are wood fragmenters although, in nature, most Camponotus species nest in soil, under debris, or in plants [148], producing cavities in wood for their nests [117]. These holometabolous (complete metamorphosing insects) are helpless in the larval stage and must be attended (groomed and fed). Therefore, only adults damage wood and do so by mining/fragmenting and removing the resulting frass with their mandibles. More than 900 Camponotus species occur worldwide, the majority of which are beneficial, and only a few are prominent pests of structures. Some 23 species of carpenter ants in North America are structural pests and seven of these species cause extensive damage to wood structures and wood [148, 149]. Carpenter ants are the most important structural insect pests (wood destroying or otherwise) in latitudes greater than 45 degrees. Carpenter bees are considered the least damaging of all wood destroying insects [150]. However, carpenter bee damage is often highly visible, and they can be difficult to control in exterior wood trim and siding of structures. Because of this, they can cause localized high-value damage and be a nuisance.
5.3.1 Carpenter Ants
Carpenter ants damage utility poles, merchantable timber, and even living trees [148, 151]. A structural pest control industry survey estimates that carpenter ants account for over 40% of all ant management activities in the USA [152] at an estimated cost (in 1986) of US$25 million annually [153]. Carpenter ants will nest in many materials other than wood, for example: structural voids, insulation (fiberglass bats & foam), hollow doors and curtain rods, and almost any dark, moist cavity. Although they have the capability of boring into and forming galleries in wood, they do not use the wood as a food source as termites do, but solely as a habitat. As such, carpenter ants can often be found in lumber piles inhabiting cracks of narrow width between boards, and they do not burrow into the wood if cracks such as these provide suitable habitat that can be kept at a suitable moisture content/humidity. Carpenter ants have been observed to re-establish colonies in locations that have better moisture/humidity environments if they can locate wood with better moisture conditions or wood undergoing decay that fungi have kept moist [154]. Carpenter ants will also bring water into their nest to enhance the moisture content by carrying droplets between their mandibles. Wet and decayed wood is softer and easier to mine, and soft bodied ant larvae cannot tolerate dry conditions. However, in some species such as Camponotus vicinus [155], the survival of a founding queen and associated brood rearing from eggs to adult-worker ants has been reported to occur in wood of low moisture content (10–13% MC). Carpenter ants can form satellite colonies containing workers, late-stage larvae, and pupae. Satellite nests can occur in drier areas if the parent colony was established in a wetter site, and in these cases, larval development/maturation may be enhanced in these warm, but less moist sites.
Like termites, carpenter ants also have winged reproductives and mating flights. In temperate climates, these flights occur when the weather warms and is more favorable for the active growth of vegetation. Mating occurs during the mating flight, and after landing, the female will discard her wings to initiate a new nest while the male dies within a day or two. Ants, like termites, are not strong flyers and tend to drift with air currents. As such, the female may not land on a suitable site to seek shelter and initiate a new nest. This queen swarmer is subject to predation by birds and animals until protecting habitat can be found or created. Once the chamber is excavated and sealed against intruders, the queen lays her first batch of eggs. She rears the resulting larvae through the pupal stage using secretions from her fat reserves and degenerating wing muscles. The nascent workers then open the nest chamber and venture out to forage. Subsequent broods, and the queen, are tended (and fed) by this first worker generation. Like termite colonies, carpenter ant societies take 5–7 years to mature and start producing their own swarmers.
Carpenter ants have much smaller colonies than the one million or so individuals of subterranean termite societies. However, their numbers are greater than originally thought. Hansen and Akre [156] found Camponotus vicinus colonies, including satellite nests, contain 100,000 individuals and a C. modoc colony to contain about half that number. Fowler [153] reported C. pennsylvanicus nests (not including satellite nests) to contain about 20,000 workers. If each worker occupied (on average) about 0.1 cc then large colonies could occupy a substantial amount of space (10 liters), requiring considerable wood removal for their nests, especially in winter when they huddle together. Shields et al. [157] found that utility poles can be almost completely hollowed out by carpenter ants to a height of 2 meters within 5–7 years after they are put into service. Sanders [158] referred to this region where only annual rings were left unmined as the “laminated zone.” Such poles were prone to failure due to excavation by carpenter ants. Similar lamellar damage is seen in some cases in dimensional lumber (Fig. 4.16).

Carpenter ant damage to a “2x6” support and plywood sheathing (cross-sectional view). The “2x6” pressure treated member was poorly treated with preservative in the interior heartwood and extensive carpenter ant damage occurred in this relatively untreated heartwood. The attack occurs in a lamellar manner, with the denser latewood remaining as the lower density early is removed. (Note removal of wood with the “sand-paper smooth” appearance of galleries and also the lack of fecal material and soil particles (mud). The sapwood of the “2x6” member at the top left and right is penetrated better than the heartwood by the pressure treatment, and hence the sapwood has not been attacked)
Because carpenter ants do not digest wood, and they can survive in cracks between boards, this can allow them to exploit wood that is preservative treated to lower levels in some cases. For this reason, preservative treatments can often be ineffective at controlling carpenter ants. This is particularly true in larger exterior-use timbers that are untreated in a portion of the wood core such as the interior regions of rail ties (sleepers) or utility poles. As these timbers age and dry, seasoning checks develop into interior untreated or poorly treated wood. Carpenter ants enter the interior wood through these cracks or checks and form galleries in the untreated or poorly treated interior wood. Carpenter ants keep clean galleries and do not coat the walls of their galleries with their feces as termites do. They eject both wood shavings and debris, including dead colony members, from their nests. Piles of these wood particles and debris, or debris found in crevices, are tell-tale evidence of the presence of a carpenter ant nest in infested wood (Fig. 4.17). It has been observed previously that soft rot fungi in some cases may be active in wood immediately adjacent to the galleries of a Camponotus infestation [85], perhaps suggesting a symbiotic relationship between carpenter ants and soft rot-causing fungi. Softening of the wood would presumably aid excavation by the ants, and ant foraging would then aid in the dissemination of the fungus. But it was not reported how frequent this association may occur.

Wood shavings and debris from the interior of a carpenter ant gallery caught in a check at the surface of a preservative treated wood pole. Poles typically are treated with preservatives such that only the outer circumference of the wood (sapwood) is treated. As drying checks develop, carpenter ants will exploit those openings to access the interior untreated wood. The interior wood is then colonized and wood shavings and other debris ejected from the galleries will be found either in piles on the surrounding ground, or more often, caught in cracks and checks such as shown here
Carpenter ants can cause significant damage to structures and structural wood despite not digesting the wood. Carpenter ants survive in colder climates where termites cannot, and the ants are therefore of greater concern to wood in service in those areas compared to termites. Wood galleries produced solely through fragmentation have the same impact on structural load bearing timbers as wood galleries produced by fragmentation and ingestion (as with termites). Therefore, the early detection of carpenter ant activities, and control efforts, is as important for this group as it is for termites [148].
5.3.2 Carpenter Bees
Unlike ants, carpenter bees are solitary and do not live in colonies. Females return to their native galleries, typically excavated in rough sawn and unpainted wood, over several seasons. Because these bees can also inhabit trees and tree branches, the population can build up in woodland areas around structures, and it can be difficult to eliminate carpenter bees just by eliminating an individual infestation. The females are known to use the same gallery for several years and expand their tunnels to rear their offspring (Fig. 4.18). Because there is no queen as in social insects, each female rears their own offspring. The bee’s mining activities result in 12 mm diameter, cylindrical channels in wood 10–15 cm in length (Fig. 4.19) [143]. A two-meter section of log, pole, or tree might contain the galleries of 10 or 12 carpenter bees. These insects expand their galleries much more slowly than their social cousins, the termites, and carpenter ants. Although the exit hole is typically the only portion of the gallery visible to observers, birds such as woodpeckers may become aware of the gallery presence and they are known to excavate into finished wood in structures to access the bee larvae as food. Gallery proximity to the exterior of wood may lead to greater damage and loss of integrity from woodpecker attack and foraging bird attack [155].

Carpenter bee damage to wood that was exposed by sawing longitudinally through the nest chamber. (Note “bee bread” pellet of pollen and nectar at the end of the branching gallery section, and entrance hole in the center of the upper section)

Carpenter bee damage to wood. (a) Carpenter bee entry holes (arrows). (b) Carpenter bee male protecting the gallery and harassing intruders. (c) Entrance holes on rough-sawn siding, with wood shavings below the galleries. (d) Entry holes to a carpenter bee gallery with the hole on the right plugged with caulk by a homeowner. Yellow streaking below the entry hole is pollen-stained fecal residue and is commonly observed below these holes. Plugging holes without killing the bee and the larvae is an ineffective control as the bees will bore out by producing other holes
Similarly to other bees, females collect pollen and nectar from flowers, but they deposit this as “bee-bread” in cells at the end of each tunnel. A bee-bread pellet will be produced over time until it is about the size of a peanut, at which time the female will deposit an egg on it and seal off the cavity [159]. In the higher latitudes, 5–6 cells are produced by each female. Carpenter bees make their nests typically in rough sawn window frames, wooden siding and shingles, eaves, fences, railings, doors, wooden ceilings (outdoor covered porches), outdoor furniture, and other wood elements. The male bee has no stinger, but will typically be present during the day outside the nest to aggressively harass intruders near the nest.
Carpenter bees do not digest the wood they excavate, and therefore, they will even bore into preservative treated wood. They often excrete near the entrance to their gallery, and yellow streaks of the pollen feces extending downward from this hole will be apparent (Fig. 4.19). Plugging carpenter bee holes is typically an ineffective control and the bees will tunnel into, or out of, the wood from an adjacent area. Killing the female as well as the larvae, followed by plugging of the gallery hole, is often necessary to eliminate site activity. Because of the carpenter bee’s preference for roughened wood surfaces, colonization can be discouraged by using sanded or planed wood that is then finished with paint or a coating.
6 Marine Borer Attack of Wood
Marine borer attack is most commonly observed on wooden structures in marinas and coastal waters, but these organisms are also active in the deep ocean as well, with attack by Xylophaga pholads having been recorded in seawater more than 1.5 km deep. In addition to the molluscan shipworms and pholads that attack wood, a group of crustaceans that are known as gribbles can attack wood and slowly erode wood from the surface. The gribbles produce some of their most visible destruction in intertidal region, where wood piling can take on an hour-glass shape with the zone of wood between low and high tide being most heavily attacked. Although the destructive capacity of marine borers is of great concern, it is also important to recognize the tremendous ecological importance of these lignocellulose-degrading organisms on a global scale. It has been shown that 70% of the dead wood biomass in mangrove swamps is digested by shipworms [160]. In terrestrial environments, it has been estimated that more than 75% of the above-ground carbon sequestered is in forest and woody biomass [161]. It currently is unknown how much of that terrestrial carbon is ultimately washed into the world’s oceans, but even if only a small fraction of that debris is processed by marine borers, this verifies their role as important cyclers of carbon on earth.
Shipworms, pholads, and gribbles have been a scourge to humankind for thousands of years. The ancient Romans wrote about damage to wooden ships, and how their ships were liable to the action of Teredo or shipworm. The children’s story of the “little Dutch Boy” plugging leaks in the wooden dikes of the Netherlands with his finger to prevent inundation of the country with seawater was based on the very real problem of deterioration of the dikes by shipworms and the galleries that they make in wood. The Netherlands resolved the issue at the time by replacing some dikes with expensive, imported stone, but attack of untreated wood around the world by marine borers continues as a common marine problem. As another measure of the importance of marine borers in history, it has been argued that the Spanish Amada, intent on the invasion of England in 1588 to overthrow Queen Elizabeth I, may well have been successful if not for the weakening of the Spanish fleet by shipworms. Delays in launching the Spanish attack on England were caused for numerous reasons including the need to mount a counter offensive against the British in Cadiz harbor, Spain, in 1587. This attack required the Spanish ships to sail or be anchored in warmer southern waters for more than a year before the Amada could initiate their attack on England. Many of these ships were previously part of the King of Spain’s fleet from India, Portugal, and the Caribbean, with several having unsound hulls because of this prior service in warm ocean waters [162]. Although many factors and individual battles occurred, ultimately the attack by the Spanish Armada failed, with more than 1/3 of the Armada’s ships being broken up and lost in storms as they retreated. Ships with sound hulls, free of marine borer attack, would more likely have survived these storms. The defeat of the Spanish Armada saw a shift in European power from a dominant feudalistic empire, to northern Europe which continued to dominate militarily after that time [163]. Although it is not possible to know if the severity of marine borer damage alone could have turned the tide of this important battle, many other instances of these organisms impacting the decisions or the fate of humans have been documented.
6.1 Shipworms
Shipworms are not actually worms but are marine mollusks that feed on wood and woody debris. They are classified in the superfamily of Pholadacea along with their cousins the pholads and with the shipworm being in the Teredinidae. More than 70 of these teredinid marine mollusks are known [164]. Representative genera include Teredo, Bankia, and Lyrodus. Shipworms typically bore into wood, using it not only as a food source but as protective housing for their soft, elongated bodies. They are active in wood from shallow intertidal regions down as deep as 200 m in the ocean [165]. As noted in the introduction to this section, shipworm activity has thwarted the marine activities of humans for thousands of years. Christopher Columbus had two of his ships destroyed when shipworms bored though the thick oak planking of his ship’s hulls during his voyages to explore the New World. Many reports of damage to ships extend back through the centuries when wooden ships were commonplace, and this has also totaled to significant monetary damages. Combined with the activities of the pholads, shipworms have been estimated to cause more than one billion US dollars damage each year globally.
Shipworms can be found in all oceans of the world [165] but different species have preferred environmental conditions. Species can survive across a wide range of water salinities, with some species able to survive in water with only nine parts per thousand (ppt) salinity and others able to survive quite brackish waters. Teredinid larvae have been reported to survive in 5 ppt salinity waters. Maximal salinity for the euryhaline species Teredo navalis has been reported at 45 ppt [166] and even 60 ppt [167]. For reference, ocean salinity averages are about 35 ppt (3.5% salt), whereas river water salinity is 0.5 ppt or less. Shipworms appear to have broad tolerance for oxygenation of water. Prior reports on shipworms from Baltic waters [168] indicate that oxygen levels for survival of Teredo adults and larvae needed to be greater than 4 mg/l and they also reported that temperature tolerance ranged from 1.4 °C to 30 °C. Turner reported that shipworms could withstand freezing temperatures. Temperatures from 4 °C to 16 °C were considered appropriate for “intense” Bankia attack on wood [169]. In New Zealand waters, Bankia was found to require temperatures of 19–20 °C for breeding, whereas Lyrodus has a lower temperature tolerance of 10 °C [170]. Nototeredo species have been reported to survive 72 h in air, and Teredo species have been reported to withdraw their siphons entirely for up to 6 weeks [167] for survival when exposed to air.
Protandry occurs in shipworms, and a female shipworm may produce and release 100,000 eggs or more. Some species of shipworm fertilize their eggs in a brood pouch near the gills. In other species, the eggs develop into larvae in the burrow of the female. The planktonic larval stage is released allowing the shipworm to disperse its young, which may travel with currents thousands of kilometers. Larvae are found both in coastal estuaries and throughout the oceans, with research suggesting that shipworm larvae release chemical cues in seawater once they have established on suitable wood which attracts other free-swimming larvae to the same location [171]. Once established on a suitable piece of wood, the shipworm can bore into the wood and grow to the adult stage to complete the life cycle.
Shipworms use a pair of modified valves (shell-like rasps) and move these with powerful adductor and foot muscles to rasp away at wood and create a burrow (Fig. 4.20). Except in early stages of larval development, the valves do not enclose the entire animal as occurs with many other mollusks. As the larvae grow, the valves develop to fit around just the anterior region of the adult borer. The outer surfaces of the valves are grooved and the grooves also have serrations to more effectively reduce the wood to minute shavings and produce a smooth burrow. Boring rates of 3 cm/week have been recorded. Typically, the borers will attach to an available wood surface and bore in at an angle to that surface. Over time as the borer matures and becomes larger, it will turn the burrow to orient along the wood grain. In some cases, the density of animals attacking the wood is so great, that the boring orientation cannot be changed or the burrow of one borer would intersect with that of another (Fig. 4.21). As they bore, the shavings of wood are ingested through the mouth. As the tube is extended, the borer secretes a calcareous material which it uses to coat the interior of the burrow in thin shell-like material (Fig. 4.22). Although it has been suggested that the shell-like coating may serve a structural purpose to prevent collapse of the burrow, and this may be true for some species of shipworm that do not live in wood, the coating is typically quite thin in wood-boring species and does not provide significant structural support. Instead it is more likely that the calcareous lining helps to regulate the conditions of the burrow by limiting diffusion of nutrients or metabolites into the wood, and similarly, it helps to allow the borer to maintain a hydrated environment when tidal conditions or drought leave the shipworm burrow above the waterline for short periods of time. The posterior end of the shipworm remains in the same location where the larvae initiated the burrow, and incurrent and excurrent siphon extend from the surface of the wood to permit nutrient and water exchange (Fig. 4.23). Sperm is secreted and eggs released from this site as well. The siphons can be withdrawn and a pair of hardened calcareous pallets extended to seal off the burrow to the exterior.

Shipworm rasps as viewed under scanning electron microscopy. (a) The valves (shells) of the shipworm are modified to form rasp-like structures around the anterior region of the animal. Higher magnification ((b) and (c)) of the valves shows the aligned serrations that allow the animal to effectively bore into the wood to produce the shipworm galleries. (Image by Dr. Reuben Shipway, and used with permission)

Tightly packed galleries of shipworms in oak wood piling. This piling in the cold coastal waters of Maine USA was attacked and completely riddled by shipworms over a one-year period until failure occurred. Typically, after entering the wood wherever access is permitted, shipworms will re-orient their galleries to align with the wood grain. In this case, however, the number of animals boring into the wood did not permit gallery realignment, and the shipworms burrowed directly to the center of the piling until the piling was weakened enough to fail

A pine wood plank attacked by both shipworms and gribble. The author’s finger (Goodell) points to the calcareous tube extending from the wood, which originally housed the shipworm. This calcareous material lines the entire shipworm gallery inside wood, completely surrounding the animal. The wood surface is fragmented because of the action of gribble

Schematic cartoon of the shipworm attack of wood. The shell-like rasps at the anterior end of the animal enlarge and lengthen the burrow of the shipworm over time. Digestion of the wood shavings is reported to occur via secretion of CAZymes secreted by the both the shipworm and by the bacterial symbionts living in the gills
It should be noted that shipworms include species that do not bore into wood and do not derive their nutrition from wood. These include the “giant shipworm” Kuphus polythalamia a relatively primitive shipworm that matures in wood, but then migrates to live in the mud of mangrove swamps. This Kuphus species has been reported to employ chemotrophic bacterial symbionts in its gills to oxidize hydrogen sulfide and use this as an energy source instead of organic compounds [172]. A soft limestone-boring, river-inhabiting shipworm, Lithoredo abatanica, has also been recently reported [173]. The mode of nutrition for this shipworm has not yet been described, but as it does not bore in wood yet has gill symbionts, a mode of chemolithotrophy is suggested.
At one time, it was thought that shipworms obtained their nutrition primarily from ingestion of plankton in the sea; however, it is now known that considerable amounts of cellulose are digested by these borers. Therefore, in addition to rasping wood to bore a burrow that they inhabit, the wood residue in the form of shavings is ingested to provide the bulk of needed nutrients by the shipworm. Based on recent work and analysis of a single species Lyrodus pedicellatus, the digestion of wood in the gut of the shipworm occurs in part through the action of cellulase enzymes secreted by the digestive organs of shipworms themselves [160]. This action is aided by the production of bacterial symbionts that grow within with the gill of the shipworm [174]. Even though a symbiotic relationship with bacteria to digest wood may seem similar in some ways to the termite’s lifestyle in the insect world, the location of the cellulase-producing symbionts located in the gills of the shipworm rather than the gut is quite unique in the Animalia kingdom. However, microbial symbionts are known to be present in the gills of other bivalve species where they contribute essential amino acids, fix carbon, and provide energy to these mollusks.
Sabbadin et al. (2018) [160] indicate that cellulase enzymes, predominantly glycohydrolase (GH)1 and ß-glucosidase, are secreted from a specialized digestive gland in the shipworm gut. These enzymes are then collected in the caecum (Fig. 4.23), a relatively large food storage/digestive organ in the shipworm, where the majority of wood residues are collected and digestion occurs. It is understood that the caecum is virtually sterile with microbial symbionts largely being absent in that organ. The symbiont cellulases (produced in the gills) represent less than 15% of the total cellulase load observed in the caecum, but still representing an interesting case of cellulase enzymes produced by both organisms involved in the symbiosis. Although the production of cellulolytic enzymes produced by the digestive organs of the shipworm and by symbiont bacteria residing the shipworm gills is very important, additional factors are required for complete digestion of the lignocellulose. In other wood biodegradation systems and in biorefinery applications, it is well known that cellulases alone are insufficient to deconstruct wood, even when that wood has been comminuted to small particles. This suggests therefore that other low molecular weight degradative agents or other undiscovered mechanisms may also play a role in the digestion of wood in the shipworm digestive system, and more research is needed to better understand this important phenomenon.
Shipworms can be difficult to control because the larvae can travel great distances, and population reduction is normally not possible. Nonchemical controls for mobile wooden structures such as boats, log rafts, and similar structures include moving infested materials to fresh water for several weeks to months to kill both adult and larval stages. Wooden boats can also be dry-docked or brought to shore for several days to weeks to limit establishment of borers in the wood. Because shipworms require oxygen from flowing seawater, stagnation of the water immediately surrounding submerged wood such as wood piling can be an effective treatment. In the fifteenth century, ships with copper or other cladding began to appear with the cladding functioning both as a physical barrier and as a layer to prevent oxygenated water from reaching the wood with copper ions also likely contributing as a biocidal component when copper sheet was used. Loss of cladding from a portion of a ship’s bottom typically resulted in attack of that portion of the exposed wooden bottom. In wooden waterfront structures such as wood piling, rigid or flexible wraps that stagnate a thin layer of water around the wood below the waterline have been effective in controlling shipworm attack. However, if holes develop in the wrap that are large enough for water to circulate, water will become oxygenated at least in localized regions, and borer larvae can penetrate very small openings. Antifouling paints, where permitted, can be effective if maintained and biocidal pressure treatments in sapwood are also effective in many cases.
6.2 Pholads
Wood boring organisms commonly known as pholads or piddocks are also mollusks, and are similar in some ways to the shipworms. Pholads are in the superfamily Pholadacea, and they are further divided into the family of Xylophagainae and the subfamily of Martesiinae [167, 175]. The Xylophagainae have the ability to bore into wood, and they are similar to the shipworms in that they are capable of digesting wood particles that are rasped from wood substrates as the burrow is produced. Because they rely on wood as a source of nutrients, pholads in the Xylophagainae continue burrowing until their death. The Martesiianae are not wood digesting borers, and although they will aggressively attack wood, some species will also bore into other available substrates such as soft rock that serve to provide anchored protection (Fig. 4.24). The two primary valves of the pholads are modified into rasping tools similarly to that seen in the shipworms, but in the pholads the valves enclose much of the animal. In some species, the valves combine with a series of supplemental plates, and the pholad body can be enclosed completely within a somewhat flexible multicomponent valve.

Burrows of rock-boring pholads collected from Vancouver Island, British Columbia, Canada. The remains of pholad shell-rasps can be observed in two of the bore holes
Like some shipworms, pholads have a free-swimming larval stage that is released by the female. The larval stage finds a suitable substrate and attaches to begin boring. Pholad burrows are relatively short compared to that of shipworms and typically are 6–7 cm long with some species reported to have burrows 15–18 cm long. The burrow shape may be ovoid to cylindrical and depending on the species may be less than 1 cm in diameter up to more than with 2.5 cm diameter. The burrow entrance may be more restricted in size.
Xylophagainae pholads inhabit deep ocean waters around the globe and have been found at depths from 150 m to greater than 7000 m. At higher latitudes, some Xylophagainae borers can occupy shallower offshore sites, but not typically near-shore shallows. They have been observed to bore into, and cause damage to, the sacrificial wooden skids on traps used by fisherman catching bottom dwelling species. More limited information is available on the life cycle and breeding in the Xylophagainae pholods because of the challenges of studying them in deep water environments. In the Xylophaga, some research suggests that dwarf males are associated with females, and rather than brooding the larvae, eggs are released which are then fertilized by these dwarf males.
The Martesiinae pholads are not obligate wood feeders as are the Xylophagainae. The Martesia genus has free-swimming larvae and it causes extensive damage of submerged wood in tropical and subtropical regions. They have a world-wide distribution in tropical and warm temperate seas and are known as an invasive species. Hawaii, southern England, and Florida have all reported Martesia as its range has expanded by inadvertent human distribution and the warming of ocean waters. The first report of Martesia striata in the Mediterranean Sea was recorded in 2015 [176], and global distribution is expanding as this species has been observed on driftwood. Even though they can be aggressive wood borers, Martesiinae pholads do not digest the wood they bore into, and therefor, biocidal pressure treatments of wood are often less effective on this type of marine borer. A dual treatment of creosote and CCA is typically recommended for protection of submerged wooden marine structures in countries where use of these chemicals is still permitted.
6.3 Gribbles
Gribbles are small crustaceans in the Isopoda order and the family Limnoriidae that have a high capacity for wood and lignocellulose consumption. Representative genera include Limnoria and Sphaeroma. Colloquially, in North America, the gribble are often known as “limnoria” no matter what genera they are from. The bodies of the adult gribble are curved like pill-bugs, their Isopod cousins, but are only 3–6 mm in length. Their color is yellow to white and they are translucent under bright light. The head of the animals is not broadly attached to the thoracic plates of the body, so that the body and head can flex to some extent as the animal bores into wood. The adult Limnoriidae has seven pairs of legs and female and male will pair within a burrow and can remain paired for several months. The adult female has been reported to carry “around 22 large eggs” in a pouch between the front legs, with the eggs being about 1/4 the body width [177, 178]. Juveniles will inhabit the adult burrow and can produce side burrows. The adult burrows typically will extend up to a centimeter or more into wood (Fig. 4.25) and the wood shavings produced are ingested by the gribble for digestion. The wood they attack is typically colonized by microorganisms and it is known that they ingest some wood degrading microorganisms, despite having a sterile gut [179]. Limnoriidae are found in full salinity waters from subpolar to tropical regions [180] and are reduced in number where fresh water mixes along shorelines and bays. However, some species can be found in stenohaline shallow waters of inlets and bays, with others species in the ocean at depths of greater than 1000 m. With the exception of few seagrass-feeding species, the Limnorid gribble utilizes wood as a major part of their food source. Sphaeroma are found in temperate and tropical areas, where they tolerate a broad range of salinity waters, primarily in intertidal zones. The Sphaeroma are aggressive wood borers but unlike the Limnorid gribble it has been reported they have mouthparts that are not suited to the further processing of wood for digestion, with masticatory structures that are present in other herbivores [181]. Genomic analysis to assess production of cellulase enzymes has not been conducted, but one report suggests higher survival rate of Sphaeroma juveniles when fed with pure cellulose [182]. Additional work is needed to assess whether wood feeding is a requirement in Sphaeroma species, at least in juveniles.

Gribble burrows at the surface of wood piling (Fig. 4.26) that has been razor-sectioned to expose the length of the burrows. The yellow circle highlights the action of a single gribble at the end of one burrow
In some parts of the world, gribble are often associated with Chelura, which are also crustaceans in the Amphipoda, and which can live in gribble burrows to compete for oxygen and nutrients. Chelura also bore into wood on their own, and although not classified as a Limnorid gribble, they are included in this section for convenience. Chelura competition has been reported to retard the rate of wood damage by gribble because of this competition. Both the Limnorid gribble and Chelura are known to produce carbohydrate active enzymes from their digestive glands, and like the molluscan borers, the digestive system contains no symbiotic microorganisms that could function to partially or fully deconstruct lignocellulose. Similarly to the molluscan shipworms, CAZymes alone could not be the sole digestive agent responsible for the deconstruction of lignocellulose, so other digestive components or systems would be needed. In this regard, a nonenzymatic mechanism that works in synergy with CAZymes secreted by limnorids has been demonstrated by Besser et al. [183] to function in the digestion of wood by these animals. In this mechanism, hemocyanin secreted into the hindgut of limnorids is proposed to function similarly to phenoxidase enzymes by modifying lignin and allowing hexose polysaccharides like cellulose to be more readily digested by CAZymes. This helps explain how some types of gribble that secrete no classically defined peroxidases into their gut can effectively circumvent the lignin barrier that otherwise would prevent digestion of wood by cellulases alone.
The gribble and Chelura have been distributed globally by oceanic travel, originally by wooden ship. However, even today larvae of marine boring organisms are taken up in bilge water which is released thousands of kilometers distant to disseminate these species. Additionally, driftwood travels thousands of miles distributing adults and early stage animals. Gribble attack is most prominent in the intertidal zone and occurs 0–30 m deep [179, 180]. Generally, the limnorid gribble are not considered to be strong swimmers and they are distributed by ocean currents and attachment to floating wood or boats. One of the most destructive species, L. lignorum, is primarily found in northern hemisphere temperate oceans. It has been reported to occur sporadically in reports from the southern hemisphere, but it is unknown if the identifications are accurate, nor why the species has not established in the southern hemisphere. L. quadripunctata is also an aggressive species that is thought to have disseminated from southern hemisphere water to the northern hemisphere. However, it does not occur in tropical waters. As oceanic temperatures have risen in the last 50 years, the range of L. quadripunctata has shifted to higher latitudes in the northern hemisphere. This species is more resistant to some types of pressure preservative wood treatment, particularly creosote. Because of this and its high feeding rate, L. quadripunctata has become known as one of the most damaging of gribble species in North American harbors.
Gribble attack occurs from the surface of the wood, and in harbors with wooden piling, these piling take on an “hour-glass” or “pencil-pointed” shape when they are aggressively attacked. Unlike shipworm and pholad attack, which can be hidden by the growth of algae and other marine fouling organisms, intertidal damage by gribble is more readily observed. Gribble attack can go unnoticed though, particular when associated with damage to submerged foundation piling and other submerged wooden structures. In this regard, wooden foundation piling is still in wide use throughout the world. Many current and historic structures are supported by wooden foundation piling. One example in Boston harbor (Boston, Massachusetts, USA) highlights the importance of monitoring for gribble attack in hidden locations including foundation structures such as the ones in the Back Bay region of that city. Figure 4.26 shows the condition of submerged structural support piling removed from beneath an 8-story brick and stone building, originally built around the turn of the nineteenth century as a wool storage warehouse, and converted now to upscale business suites. Approximately 100 years after construction, engineers noted that one corner of the building had sunk by a few millimeters. Typically, submerged piling would be surrounded by granite blocks and/or soil/rock fill, which is impervious to marine borer attack. This was the case originally with this structure. To investigate the sinking of the building though, crews were brought in to excavate under the building, below sea level, removing granite and fill so that inspection of the wooden piling could be done by the first author and engineering crews. It became apparent that tidal action over the years had washed away fill material in between some of the granite block and wood piling, in some locations for a distance of greater than 35 meters horizontally under the structure. Submerged wooden piling typically shows no structural problems for hundreds of years, but in this case where the fill materials had been removed by tidal action, gribble were able to travel with tidal action for more than 30 meters horizontally under the building to attack the piling in the newly exposed locations (Fig. 4.26). The damage was identified in piling from 8 to 10 meters below sea level, and pumps had to be used continually to permit excavation, inspection, and repair.

(a) Wood foundation piling during excavation under a building that shows evidence of selective gribble attack. After decades of tidal washing, soil was washed away from the piling and channels opened up to seawater, which allowed the gribble access to (b) select areas of the foundation piling where damage was caused
7 Summary and Future Perspective
This chapter provides an overview of agents involved in wood deterioration including fungi, bacteria, insects, and marine organisms. Although by no means a complete compendium of wood degrading organisms, these microorganisms and animals are associated with the bulk of damage/deconstruction caused by living organisms to wood and other lignocellulose substrates, such as bamboo, used in structural applications. Understanding the biology and habits of degradative agents is important from several perspectives. First, they all contribute to the cycling of carbon in the environment, and with much current focus on carbon sequestration as related to climate change, it is important to have a broad understanding of the organisms overviewed in this chapter as they are involved in the cycling of the largest reserve of terrestrial fixed carbon on earth – wood and related lignocellulose materials. Additionally, from an economic perspective, destruction amounting to billions of dollars annually is associated with the damage caused by these organisms as well as the resources expended on the remediation and protection of wooden structures.
Wood protection with biocides and nonbiocidal systems is covered in other chapters of this book. The purpose of this chapter has been to provide the reader an understanding of why protection is needed, and also how new types of wood protection systems may be developed in the future based on our knowledge of wood deconstruction mechanisms employed by a variety of organisms. Knowledge of the life-cycle of termites, for example, led to the development of insect growth regulators and chitin synthesis inhibitors that now are widely used to slowly disrupt the breeding cycle of termite colonies. Use of this type of inhibitor results in the decline and eventual collapse of termite colonies as the worker population is reduced, resulting in effective control. A better understanding of the biochemical mechanisms and the biology of the other organisms reviewed in this chapter will provide the foundation for the development of better control and protection systems and reduce reliance on more toxic, broad-spectrum pesticides and wood preservatives.
Relative to future research needs, fundamental research will continue to uncover new mechanisms that the organisms reviewed in this chapter used to deconstruct and metabolize lignocellulose materials. In the last 25 years, new understanding of how some fungi attack wood has completely changed our thinking about how decay mechanisms evolved. New mechanisms involved in wood digestion by other organisms reviewed in this chapter should also be expected in the future. New findings on the relationships present between bacterial symbionts in both insects and marine borers are likely to spawn new thinking on wood degradation mechanisms in these organisms – and new types of targeted and environmentally friendly control practices. Further the role of an entire microbial domain of life, the archaea, is virtually unexplored relative to their function in symbiotic relationships with wood degrading animals such as marine borers and insects. It is likely that important roles for these microorganisms in symbiotic relationships, or as sole agents of wood deterioration in select environments, will be discovered in the future. Although considerable progress has been made in our understanding of decay and degradation mechanisms, additional work is needed to open new knowledge frontiers and to allow the development of both better systems for control of wood degradation and better systems for harnessing the biotechnological power of these organisms to benefit future sustainable bioprocessing of renewable lignocellulose materials.
References
Cragg, S.M., Beckham, G.T., Bruce, N.C., Bugg, T.D.H., Distel, D.L., Dupree, P., Etxabe, A.G., Goodell, B.S., Jellison, J., McGeehan, J.E., McQueen-Mason, S.J., Schnorr, K., Walton, P.H., Watts, J.E.M., Zimmer, M.: Curr. Opin. Chem. Biol. 29, 108–119 (2015)
Geib, S.M., Filley, T.R., Hatcher, P.G., Hoover, K., Carlson, J.E., Jimenez-Gasco, M.d.M., Nakagawa-Izumi, A., Sleighter, R.L., Tien, M.: Proc. Natl. Acad. Sci. 105, 12932–12937 (2008)
Encinas, O., Daniel, G.: Mater. Org. 30, 239 (1996)
Daniel, G.: In: Nicholas Darrel, D., Goodell, B., Schultz Tor, P. (eds.) Deterioration and Protection of Sustainable Biomaterials Acs Symposium Series Vol 1158, pp. 23–58. American Chemical Society, Washington, DC (2014)
Goodell, B., Zhu, Y., Kim, S., Kafle, K., Eastwood, D., Daniel, G., Jellison, J., Yoshida, M., Groom, L., Pingali, S.V., O’Neill, H.: Biotechnol. Biofuels. 10, 179 (2017)
Zabel, R.A., Morrell, J.J.: In Press, 2nd edn. Academic Press, San Diego (2020)
Worrall, J.J., Anagnost, S.E., Wang, C.J.K.: Can. J. Microbiol. 37, 869–874 (1991)
Williams, S.: Cazypedia carbohydrate-active enzymes. https://www.cazypedia.org/index.php/Glycoside_Hydrolase_Families (2017). Accessed 30 Dec 2017
Bissaro, B., Várnai, A., Røhr, Å.K., Eijsink, V.G.H.: Microbiol. Mol. Biol. Rev. 82, e00029–e00018 (2018)
Arantes, V., Jellison, J., Goodell, B.: Appl. Microbiol. Biotechnol. 94, 323–338 (2012)
Suzuki, M.R., Hunt, C.G., Houtman, C.J., Dalebroux, Z.D., Hammel, K.E.: Environ. Microbiol. 8, 2214–2223 (2006)
Riley, R., Salamov, A.A., Brown, D.W., Nagy, L.G., Floudas, D., Held, B.W., Levasseur, A., Lombard, V., Morin, E., Otillar, R., Lindquist, E.A., Sun, H., LaButti, K.M., Schmutz, J., Jabbour, D., Luo, H., Baker, S.E., Pisabarro, A.G., Walton, J.D., Blanchette, R.A., Henrissat, B., Martin, F., Cullen, D., Hibbett, D.S., Grigoriev, I.V.: Proc. Natl. Acad. Sci. 111, 9923–9928 (2014)
Floudas, D., Binder, M., Riley, R., Barry, K., Blanchette, R.A., Henrissat, B., Martínez, A.T., Ortillar, R., Spatafora, J.W., Yadav, J.S., Aerts, A., Benoit, I., Boyd, A., Carlson, A., Copeland, A., Coutinho, P.M., de Vries, R.P., Ferreira, P., Findley, K., Forest, B., Gaskell, J., Glotzer, D., Górecki, P., Heitman, J., Hesse, C., Hori, C., Igarashi, K., Jurgens, J.A., Kallen, N., Kersten, P., Khajamohiddin, S., Kohler, A., Kues, U., Kumar, A., Kuo, A., LaButti, K., Larrondo, L.F., Lindquist, E., Ling, A., Lucas, S., Lundell, T., Martin, R., McLaughlin, D.J., Morgenstern, I., Morin, E., Murat, C., Nolan, M., Ohm, R., Patyshakuliyeva, A., Petersen, R.H., Rokas, A., Ruiz-Dueñas, F.J., Sabat, G., Salamov, A., Samejima, M., Schmutz, J., Slot, J.C., St, F., John, J., Stenlid, H., Sun, S., Sun, A., Tsang, A., Wiebenga, D., Young, A., Pisabarro, D.C., Eastwood, F., Martin, D., Cullen, I.V., Grigoriev, D.S.H.: Science. 336, 1715–1719 (2012)
Eastwood, D.C.: In: Deterioration and Protection of Sustainable Biomaterials. American Chemical Society – Oxford University Press, Book Series 1158 (2014)
Ding, S.-Y., Liu, Y.-S., Zeng, Y., Himmel, M.E., Baker, J.O., Bayer, E.A.: Science. 338, 1055–1060 (2012)
Stone, J.E., Scallan, A.M.: Tappi. 50, 496–501 (1967)
Tepfer, M., Taylor, I.E.P.: Science. 213, 761–763 (1981)
Kleman-Leyer, K., Agosin, E., Conner, A.H., Kirk, T.K.: Appl. Environ. Microbiol. 58, 1266–1270 (1992)
Highley, T.L., Illman, B.L.: Biodeterior. Abstr. 5, 231–244 (1991)
Eastwood, D.C.: In: Deterioration and Protection of Sustainable Biomaterials Acs Symposium Series Vol 1158, pp. 93–112. American Chemical Society, Washington, DC (2014)
Wu, B., Gaskell, J., Zhang, J., Toapanta, C., Ahrendt, S., Grigoriev, I.V., Blanchette, R.A., Schilling, J.S., Master, E., Cullen, D., Hibbett, D.S.: ISME J. 13, 1391–1403 (2019)
Wilcox, W.W.: Wood Fiber. 9, 252–257 (1978)
Barr, D.P., Aust, S.D.: Environ. Sci. Technol. 28, 79A–87A (1994)
Goodell, B., Jellison, J., Liu, J., Daniel, G., Paszczynski, A., Fekete, F., Krishnamurthy, S., Jun, L., Xu, G.: J. Biotechnol. 53, 2–3, 133–162. (1997). ISSN 0168-1656. (https://www.sciencedirect.com/science/article/pii/S0168165697016817)
Yelle, D.J., Wei, D., Ralph, J., Hammel, K.E.: Environ. Microbiol. 13, 1091–1100 (2011)
Cowling, E.B.: In: Cote Jr., W.A. (ed.) Cellular ultra-structure of woody plants. Syracuse Univ. Press, Syracuse, N. Y. pp. 341–368 (1964)
Pettersson, G., Cowling, E.B., Porath, J.: Biochim. Biophys. Acta. 67, 1–8 (1963)
Liu, J., Zhu, Y., Wang, C., Goodell, B., Esker, A.: Int. J. Biol. Macromol. 153, 443–440 (2020)
Kent, M.S., Zeng, J., Rader, N., Avina, I.C., Simoes, C.T., Brenden, C.K., Busse, M.L., Watt, J., Giron, N.H., Alam, T.M., Allendorf, M.D., Simmons, B.A., Bell, N.S., Sale, K.L.: Green Chem. 20, 3024–3037 (2018)
Kido, R., Takeeda, M., Manabe, M., Miyagawa, Y., Itakura, S., Tanaka, H.: Biocontrol. Sci. 20, 105–113 (2015)
Hatakka, A., Hammel, K.E.: In: Hofrichter, M. (ed.) Industrial Applications. The Mycota (a Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research), vol. 10, pp. 319–340. Springer, Berlin/Heidelberg (2011)
Jellison, J., Goodell, B., Fekete, F., Chandhoke, V.: In: Proceedings of 21st Annual Meeting of the International Research Group on Wood Preservation Rotorua, New Zealand. IRG/WP 1442. 16pp. (1990) https://www.irg-wp.com/search-irg-docs.html
Fekete, F.A., Chandhoke, V., Jellison, J.: Appl. Environ. Microbiol. 55, 2720–2722 (1989)
Jellison, J., Chandhoke, V., Goodell, B., Fekete, F., Hayashi, N., Ishihara, M., Yamamoto, K.: In: Proceedings of 22nd Annual Meeting of the International Research Group on Wood Preservation May 20-24 Kyoto, Japan. IRG-WP-1479. 16pp. (1991) https://www.irg-wp.com/search-irg-docs.html
Jellison, J., Chandhoke, V., Goodell, B., Fekete, F.A.: Appl. Microbiol. Biotechnol. 35, 805–809 (1991)
Chandhoke, V., Goodell, B., Jellison, J., Fekete, F.A.: FEMS Microbiol. Lett. 90, 263–266 (1992)
Goodell, B., Liu, J., Jellison, J., Jun, L., Paszczynski, A., Fekete, F.: Chelation activity and hydroxyl radical production mediated by low molecular weight compounds phenolate isolated from Gloeophyllum Trabeum. (Eds.) E. Srebotnik and K. Messner. International Conference on Biotechnology in the Pulp and Paper Industry (ICBPPI). TAPPI Sixth Intrnl. Conf. on Biotech in Pulp and Paper. Facultas-Universitätsverlag, Berggasse 5, A-1090 Wien, Austria. 661 pp. (1996)
Arantes, V., Milagres, A.: Biotechnol. 81, 413–419 (2006)
Salgado, P., Melin, V., Durán, Y., Mansilla, H., Contreras, D.: Environ. Sci. Technol. 51, 3687–3693 (2017)
Nurika, I., Eastwood, D., Bugg T., Barker G., J. Ind. Microbiol. Biotechnol. 47(1), 145–154 (2020)
Hammel, K., Kapich, A., Jensen, K., Ryan, Z.: Enzym. Microb. Technol. 30, 445–453 (2002)
Eastwood, D.C., Floudas, D., Binder, M., Majcherczyk, A., Schneider, P., Aerts, A., Asiegbu, F.O., Baker, S.E., Barry, K., Bendiksby, M., Blumentritt, M., Coutinho, P.M., Cullen, D., de Vries, R.P., Gathman, A., Goodell, B., Henrissat, B., Ihrmark, K., Kauserud, H., Kohler, A., LaButti, K., Lapidus, A., Lavin, J.L., Lee, Y.-H., Lindquist, E., Lilly, W., Lucas, S., Morin, E., Murat, C., Oguiza, J.A., Park, J., Pisabarro, A.G., Riley, R., Rosling, A., Salamov, A., Schmidt, O., Schmutz, J., Skrede, I., Stenlid, J., Wiebenga, A., Xie, X., Kuees, U., Hibbett, D.S., Hoffmeister, D., Hogberg, N., Martin, F., Grigoriev, I.V., Watkinson, S.C.: Science. 333, 762–765 (2011)
Zhuang, L., Guo, W., Yoshida, M., Feng, X., Goodell, B.: RSC Adv. 5, 104043–104047 (2015)
Tamaru, Y., Yoshida, M., Eltis, L.D., Goodell, B.: Int. J. Biol. Macromol. 128, 340–346 (2019)
Zhu, Y., Mahaney, J., Jellison, J., Cao, J., Gressler, J., Hoffmeister, D., Goodell, B. . J. Ind. Microbiol. Biotechnol. 44(3):329–338 (2017)
Zhang, J., Presley, G.N., Hammel, K.E., Ryu, J.-S., Menke, J.R., Figueroa, M., Hu, D., Orr, G., Schilling, J.S.: Proc. Natl. Acad. Sci. 113, 10968–10973 (2016)
Paszczynski, A., Crawford, R., Funk, D., Goodell, B.: Appl. Environ. Microbiol. 65, 674–679 (1999)
Kerem, Z., Jensen, K.A., Hammel, K.E.: FEBS Lett. 446, 49–54 (1999)
Boddy, L.: In: The Fungi, 3rd edn, pp. 293–336. Academic Press, Boston (2016).
Dashtban, M., Schraft, H., Syed, T.A., Qin, W.: Int. J. Biochem. Mol. Biol. 1, 36–50 (2010)
Baba, Y., Tanabe, T., Shirai, N., Watanabe, T., Honda, Y., Watanabe, T.: Biomass Bioenergy. 35, 320–324 (2011)
Otjen, L., Blanchette, R.A.: Can. J. Bot. 64, 905–911 (1986)
Otjen, L., Blanchette, R., Effland, M., Leatham, G.: Holzforschung. 41, 343 (1987)
Blanchette, R.A., Burnes, T.A., Leatham, G.F., Effland, M.J.: Biomass. 15, 93–101 (1988)
Wolfaardt, F., Taljaard, J.L., Jacobs, A., Male, J.R., Rabie, C.J.: Bioresour. Technol. 95, 25–30 (2004)
Eriksson, K.-E., Kirk, T.K.: In: Moo-Young, M. (ed.) Comprehensive Biotechnology, vol. 4, pp. 271–294. Pergamon Press, Toronto (1985)
Capolupo, L., Faraco, V.: Appl. Microbiol. Biotechnol. 100, 9451–9467 (2016)
Sonoki, T., Takahashi, K., Sugita, H., Hatamura, M., Azuma, Y., Sato, T., Suzuki, S., Kamimura, N., Masai, E.: ACS Sustain. Chem. Eng. 6, 1256–1264 (2018)
Shinoda, E., Takahashi, K., Abe, N., Kamimura, N., Sonoki, T., Masai, E.: J. Ind. Microbiol. Biotechnol. 46, 1071–1080 (2019)
Aust, S.D.: Environ. Health Perspect. 103, 59–61 (1995)
Abdel-Hamid, A.M., Solbiati, J.O., Cann, I.K.O.: In: Sariaslani, S., Gadd, G.M. (eds.) Advances in Applied Microbiology, vol. 82, pp. 1–28. Academic Press, San Diego (2013)
Baldrian, P., Valaskova, V.: FEMS Microbiol. Rev. 32, 501–521 (2008)
Rytioja, J., Hildén, K., Yuzon, J., Hatakka, A., de Vries, R.P., Mäkelä, M.R.: Microbiol. Mol. Biol. Rev. 78, 614–649 (2014)
Kim, J.S., Gao, J., Daniel, G.: Int. Biodeterior. Biodegradation. 105, 30–40 (2015)
Tuor, U., Winterhalter, K., Fiechter, A.: J. Biotechnol. 41, 1–17 (1995)
Leonowicz, A., Matuszewska, A., Luterek, J., Ziegenhagen, D., Wojtaś-Wasilewska, M., Cho, N.-S., Hofrichter, M., Rogalski, J.: Fungal Genet. Biol. 27, 175–185 (1999)
Vasina, D.V., Moiseenko, K.V., Fedorova, T.V., Tyazhelova, T.V.: PLoS One. 12, e0173813 (2017)
Daniel, G.: In: Wood Deterioration and Preservation Acs Symposium Series Vol 845, pp. 34–72. American Chemical Society, Washington, DC (2003)
Arantes, V., Milagres, A.M.F., Filley, T., Goodell, B.: J. Ind. Microbiol. Biotechnol. 38, 541–555 (2011).
Messner, K., Fackler, K., Lamaipis, P., Gindl, W., Srebotnik, E., Watanabe, T.: Where We Are Today ACS Symposium Series, Vol. 845, pp. 73–96 (2003).
Fackler, K., Gradinger, C., Schmutzer, M., Tavzes, C., Burgert, I., Schwanninger, M., Hinterstoisser, B., Watanabe, T., Messner, K.: Food Technol. Biotechnol. 45, 269–276 (2007)
Fernandez-Fueyo, E., Ruiz-Duenas, F.J., Miki, Y., Martinez, M.J., Hammel, K.E., Martinez, A.T.: J. Biol. Chem. 287, 16903–16916 (2012)
Mäkelä, M.R., Marinović, M., Nousiainen, P., Liwanag, A.J.M., Benoit, I., Sipilä, J., Hatakka, A., de Vries, R.P., Hildén, K.S.: In: Sariaslani, S., Gadd, G.M. (eds.) Advances in Applied Microbiology, vol. 91, pp. 63–137. Academic Press, San Diego (2015)
Pérez-Boada, M., Ruiz-Dueñas, F.J., Pogni, R., Basosi, R., Choinowski, T., Martinez, M.J., Piontek, K., Martínez, A.T.: J. Mol. Biol. 354, 385–402 (2005)
Qin, X., Sun, X., Huang, H., Bai, Y., Wang, Y., Luo, H., Yao, B., Zhang, X., Su, X.: Biotechnol. Biofuels. 10, 103 (2017)
Hofrichter, M.: Enzym. Microb. Technol. 30, 454–466 (2002)
Knop, D., Levinson, D., Makovitzki, A., Agami, A., Lerer, E., Mimran, A., Yarden, O., Hadar, Y.: Appl. Environ. Microbiol. 82, 4070–4080 (2016)
Floudas, D., Binder, M., Riley, R., Barry, K., Blanchette, R.A., Henrissat, B., Martínez, A.T., Otillar, R., Spatafora, J.W., Yadav, J.S., Aerts, A., Benoit, I., Boyd, A., Carlson, A., Copeland, A., Coutinho, P.M., de Vries, R.P., Ferreira, P., Findley, K., Foster, B., Gaskell, J., Glotzer, D., Górecki, P., Heitman, J., Hesse, C., Hori, C., Igarashi, K., Jurgens, J.A., Kallen, N., Kersten, P., Kohler, A., Kües, U., Kumar, T.K.A., Kuo, A., LaButti, K., Larrondo, L.F., Lindquist, E., Ling, A., Lombard, V., Lucas, S., Lundell, T., Martin, R., McLaughlin, D.J., Morgenstern, I., Morin, E., Murat, C., Nagy, L.G., Nolan, M., Ohm, R.A., Patyshakuliyeva, A., Rokas, A., Ruiz-Dueñas, F.J., Sabat, G., Salamov, A., Samejima, M., Schmutz, J., Slot, J.C., St, F., John, J., Stenlid, H., Sun, S., Sun, K., Syed, A., Tsang, A., Wiebenga, D., Young, A., Pisabarro, D.C., Eastwood, F., Martin, D., Cullen, I.V., Grigoriev, D.S.H.: Science. 336, 1715–1719 (2012)
ExPASy: Enzyme Entry Ec 1.11.1.19. Dye decolorizing peroxidase. https://enzyme.expasy.org/EC/1.11.1.19 (2018)
Lauber, C., Schwarz, T., Nguyen, Q.K., Lorenz, P., Lochnit, G., Zorn, H.: AMB Express. 7, 164 (2017)
Mate Diana, M., Alcalde, M.: Microb. Biotechnol. 10, 1457–1467 (2017)
Camarero, S., Ibarra, D., Martínez, M.J., Martínez, Á.T.: Appl. Environ. Microbiol. 71, 1775–1784 (2005)
Niku-Paavola, M.L., Anke, H., Poppius-Levlin, K., Viikari, L.: In: Applications of Enzymes to Lignocellulosics Acs Symposium Series Vol 855, pp. 176–190. American Chemical Society, Wasington, DC (2003)
Greaves, H.: Holzforschung. 31, 71–79 (1977)
Daniel, G., Nilsson, T.: In: Bruce, A., Palfreyman, J. (eds.) Forest Products Biotechnology, pp. 235–250. Taylor and Francis Publishers, London (1998)
Zabel, R.A., Lombard, F.F., Wang, C.J.K., Terracina, F.: Wood Fiber Sci. 17, 75–91 (1985)
Koppers, I.: Company: Koppers Pressure Treatments Help Wood Serve You Longer. Koppers Company, Pittsburgh (1977)
Resch, H., Arganbright, D.G.: For. Prod. J. 21, 38–43 (1971)
Davies, D.L.: Proc. Am. Wood- Pres. Assoc. 67, 37–42 (1971)
Blanchette, R.A., Held, B.W., Jurgens, J.A., McNew, D.L., Harrington, T.C., Duncan, S.M., Farrell, R.L.: Appl. Environ. Microbiol. 70, 1328–1335 (2004)
Unligil, H.H., Chafe, S.C.: Wood Sci. Technol. 8, 27–32 (1974)
Hale, M.D., Eaton, R.A.: Mycologia. 77, 447–463 (1985)
Tanaka, H., Fuse, G., Enoki, A.: Mater. Org. 27, 157–170 (1992)
Goodell, B., Qian, Y., Jellison, J.: In: Development of Commercial Wood Preservatives Acs Symposium Series Vol 982, pp. 9–31. American Chemical Society, Washington, DC (2008)
Simonis, J.L., Raja, H.A., Shearer, C.A.: Fungal Divers. 31, 135–146 (2008)
Tanaka, H., Itakura, S., Enoki, A.: Holzforschung. 54, 463–468 (2000)
Liers, C., Arnstadt, T., Ullrich, R., Hofrichter, M.: FEMS Microbiol. Ecol. 78, 91–102 (2011)
Morrell, J., Zabel, R.A.: Wood Fiber Sci. 17, 132–143 (1985)
Heckman, C.: Tech. Rep., Seattle post-intelligencer reporter Seattle, Washington, DC. https://www.seattlepi.com/news/article/Big-wood-stain-lawsuit-settled-1099677.php (2002)
Goodell, B.S., Resch, H., Bendtsen, B.A.: Wood Fiber Sci. 17, 377–381 (2007)
Encinas, O., Daniel, G.: Pat. Mater. Org. 29, 255–272 (1995)
Daniel, G.: In: Kim, Y.S., Funada, R., Singh, A.P. (eds.) Secondary Xylem Biology, pp. 131–167. Academic Press, Boston (2016)
De Beer, Z., Seifert, K., Wingfield, M.: CBS Biodivers. Ser. 12, 1–19 (2013)
De Beer, Z.W., Seifert, K., Wingfield, M.: Utrecht, CBS-KNAW Fungal Biodiversity Centre. CBS Biodivers. Ser. 12, 261–268 (2013)
Uzunovic, A., Byrne, T., Gignac, M., Yang, D.-Q.: Tech. Rep., N. R. C. FPInnovations-FORINTEK FPInnovations, Canada. https://fpinnovations.ca/media/publications/Documents/wood-discoloration.pdf (2008)
Miller, D.J., Goodell, B.: For. Prod. J. 31, 54–55 (1981)
NeLMA: Beware of using blue-stained wood in wood packaging materials. Tech. Rep. http://www.nelma.org/beware-of-using-blue-stained-wood-in-wood-packaging-materials/ (2016)
FAO: Regulation of wood packaging material in international trade. Tech. Rep., Food and Agriculture Organization of the United Nations: International Plant Protection Convention, Viale delle Terme di Caracalla, Rome. https://www.ippc.int/static/media/files/publication/en/2016/06/ISPM_15_2013_En_2016-06-07.pdf (2016)
Forest-Products-Laboratory: Wood Handbook – Wood as an Engineering Material. General Technical Report Fpl-Gtr-190, p. 508. U.S. Department of Agriculture, Forest Service, Forest Products Laboratory, Madison (2010)
Singh, A., Kim, Y., Singh, T.: Bacterial Degradation of Wood (2016).
Johnston, S.R., Boddy, L., Weightman, A.J.: FEMS Microbiol. Ecol. 92, fiw179 (2016)
Croan, S.C., Highley, T.: Biological treatment for controlling wood deteriorating fungi. A method for retarding the growth of wood-degrading fungi by treating the wood with an effective amount of viable, Nonsporulating Streptomyces Rimosus Sc-36 Nrrl 21063., Patent (1993)
Melillo, J.M., Frey, S.D., DeAngelis, K.M., Werner, W.J., Bernard, M.J., Bowles, F.P., Pold, G., Knorr, M.A., Grandy, A.S.: Science. 358, 101–105 (2017)
Bjordahl, C., Daniel, G., Nilsson, T.: Int. Biodeterior. Biodegradation. 45, 15–26 (2000)
Powell, K.L., Pedley, S., Daniel, G., Corfield, M.: Inter. Biodeter. Biodegrad. 47, 165–173 (2001)
Riess, W., Daniel, G.: Int. J. Naut. Archaeol. 26, 330–338 (1997)
Ulyshen, M.D.: Biol. Rev. 91, 70–85 (2016).
van der Wal, A., de Boer, W., Smant, W., van Veen, J.A.: Plant Soil. 301, 189–201 (2007)
Anon: Awpa E10, standard method of testing wood preservatives by laboratory soil-block cultures. 2015 AWPA book of standards E10–15, 418 21 (2015)
Bradshaw, C.J.A., Leroy, B., Bellard, C., Roiz, D., Albert, C., Fournier, A., Barbet-Massin, M., Salles, J.-M., Simard, F., Courchamp, F.: Nat. Commun. 7, 12986 (2016)
Potter, M.F.: In: Handbook of Pest Control: The Behavior, Life History, and Control of Household Pests, pp. 293–441. Mallis Handbook, Richfield (2011)
Currie, C.R.: Annu. Rev. Microbiol. 55, 357–380 (2001)
Warren, R., Bradford, M.: Insect. Soc. 59, 215–221 (2012)
Breznak, J.A., Brune, A.: Annu. Rev. Entomol. 39, 453–487 (1994)
Eggleton, P., Tayasu, I.: Ecol. Res. 16, 941–960 (2001).
Grimaldi, D.A., Engel, M.S.: Cambridge University Press, Cambridge/New York (2005)
Engel, M.S., Grimaldi, D., Krishna, K.: Am. Mus. Novit. 3650, 1–27 (2009)
Stokland, J.N., Siitonen, J., Jonsson, B.G.: Biodiversity in Dead Wood, Ecology, Biodiversity, and Conservation. Cambridge University Press, New York (2012)
Inward, D., Beccaloni, G., Eggleton, P.: Biol. Lett. 3, 331–335 (2007)
Su, N.-Y., Scheffrahn, R.H.: In: Abe, T., Bignell, D.E., Higashi, M. (eds.) Termites: Evolution, Sociality, Symbioses, Ecology, pp. 437–453. Springer, Netherlands (2000). http://springerlink.bibliotecabuap.elogim.com/5410.1007/5978-5494-5017-3223-5479_5420
Lenz, M., Amburgey, T.L., Dai, Z.-R., Mauldin, J.K., Preston, A.F., Rudolph, D., Williams, E.R.: Sociobiology. 18, 203–254 (1991)
Bradford, M.A., Warren II, R.J., Baldrian, P., Crowther, T.W., Maynard, D.S., Oldfield, E.E., Wieder, W.R., Wood, S.A., King, J.R.: Climate Change. 4, 625 (2014)
Grace, J.K.: In: Proceedings of the National Conference on Urban Entomology, pp. 67–76. University of Maryland, College Park (1992)
Haverty, M.I., Nutting, W.L., Lafage, J.P.: Insect. Soc. 23, 175–178 (1976)
Forschler, B.T., Townsend, M.L.: Environ. Entomol. 25, 952–962 (1996)
Ausmus, B.S.: Ecol. Bull. 25, 180–192 (1977)
Benemann, J.R.: Science. 181, 164–165 (1973)
Kitchell, J.F., O’Neill, R.V., Webb, D., Gallepp, G.W., Bartell, S.M., Koonce, J.F., Ausmus, B.S.: BioScience. 29, 28–34 (1979)
Yamada, A., Inoue, T., Wiwatwitaya, D., Ohkuma, M., Kudo, T., Sugimoto, A.: Ecosystems. 9, 75–83 (2006)
Saunders, D.: In: Handbook of Pest Control: The Behavior, Life History, and Control of Household Pests, pp. 463–513. Mallis Handbook, Richfield (2011)
Anonymous: Methods of laborayory testing of wood preservatives against fungi and borers (powder post beetles): part 1 determination of threshold values of wood preservatives against fungi. Tech. Rep., Bureau of Indian Standards, New Dehli (2008)
Parkin, E.A.: J. Exp. Biol. 17, 364–377 (1940)
Zabel, R.A., Morrell, J.J.: Academic Press, New York (1992)
Zhong, H., Schowalter, T.D.: Can. J. For. Res. 19, 943–947 (1989)
Noda, H., Kodama, K.: Appl. Environ. Microbiol. 62, 162–167 (1996)
Becker, G.: Mater. Org. 12, 141–160 (1977)
Suh, S.-O., Blackwell, M.: FEMS Yeast Res. 5, 87–95 (2004)
Akre, R.D., Hansen, L.D., Myhre, E.A.: Am. Entomologiste. 41, 221–226 (1995)
Buczkowski, G., Wang, C.: In: Dhang, P. (ed.) Urban Insect Pests: Sustainable Management Strategies. CABI, Oxfordshire (2014)
Barrette, J., Thiffault, E., Saint-Pierre, F., Wetzel, S., Duchesne, I., Krigstin, S.: Dynamics of dead tree degradation and shelf-life following natural disturbances: can salvaged trees from boreal forests ‘fuel’ the forestry and bioenergy sectors? Forestry. 88, 275–290 (2015)
Fowler, H.G., Roberts, R.B.: Zeitschrift für Angewandte Entomologie. 94, 149–152 (2009)
NuVentures: The us structural pest control industry – the 1991 season. Tech. Rep., NuVentures Consultants, La Jolla (1992)
Fowler, H.: In: Vinson, S.B. (ed.) Economic Impact and Control of Social Insects, pp. 272–289. Praeger, New York (1986)
Mankowski, M.E.: Biology of the carpenter ants camponotus vicinus (Mayr) and camponotus modoc (wheeler) in Western Oregon. PhD PHD thesis, FOREST PRODUCTS, Oregon State, Corvallis (2001)
Mankowski, M.E., Morrell, J.J.: J. Econ. Entomol. 104, 740–744 (2011)
Hansen, L.D., Akre, R.D.: In: Meer, R.K.V., Jaffe, K., Cedeno, A. (eds.) Applied Myrmecology: A World Perspective, p. 741. Westview Press, Boulder (1990)
Shields, E.J., Jacobs-Lorena, V.Y., Blagen, M., Testa, A.M.: Am. Entomol. 46, 50–55 (2000)
Sanders, C.J.: Can. Entomol. 96, 894–909 (1964)
Sims, S.R.: In: Handbook of Pest Control: The Behavior, Life History, and Control of Household Pests, pp. 823–881. Mallis Handbook, Richfield (2011)
Sabbadin, F., Pesante, G., Elias, L., Besser, K., Li, Y., Steele-King, C., Stark, M., Rathbone, D.A., Dowle, A.A., Bates, R., Shipway, J.R., Cragg, S.M., Bruce, N.C., McQueen-Mason, S.J.: Biotechnol. Biofuels. 11, 59 (2018)
Watson, R.T., Noble, I.R., Bolin, B., Ravindranath, N.H., Verardo, D.J., Dokken, D.J. (Eds.): Land use, land-use change and forestry. Tech. Rep., Intergovernmental Panel on Climate Change (IPCC): C/O World Meteorological Organization, IPCC. World Meteorological Organization. 7bis Avenue de la Paix, C.P. 2300, CH- 1211 Geneva 2, Switzerland. http://www.ipcc.ch/ipccreports/sres/land_use/index.php?idp=3 (2000)
Nolan, C.J.: The Age of Wars of Religion, 1000–1650: An Encyclopedia of Global Warfare and Civilization, 1. Greenwood Press, Westport (2006)
Dodson, E.J.: The Discovery of First Principles. Vol. I. iUniverse.com. Bloomington, Indiana. ISBN: 9780595214327. 668p (2002)
Treneman, N., Borges, L., Shipway, R., Raupach, M., Altermark, B., Carlton, J.: Aquatic Invasions. 13, 101–112 (2018)
Distel, D.L.: In: Wood Deterioration and Preservation Acs Symposium Series Vol 845, pp. 253–271. American Chemical Society, Washington, DC (2003)
Tuente, U., Piepenburg, D., Spindler, M.: Helgol. Mar. Res. 56, 87–94 (2002)
Voight, J.R.: J. Molluscan Stud. 81, 175–186 (2015)
Manders, M.R.: Guidelines for predicting decay by shipworm in the Baltic Sea, Tech. Rep. https://www.machuproject.eu/documenten/Wreck_protect_Guidelines_Predicting.pdf (2011)
Quayle, D.B.: Marine Wood Borers in British Columbia: Canadian Special Publication of Fisheries and Aquatic Sciences. Department of Fisheries and Oceans, Ottawa (1992)
McKoy, J.L.: N. Z. J. Mar. Freshw. Res. 14, 263–275 (1980)
Toth, G.B., Larsson, A.I., Jonsson, P.R., Appelqvist, C.: PLoS One. 10, e0124950 (2015)
Distel, D.L., Altamia, M.A., Lin, Z., Shipway, J.R., Han, A., Forteza, I., Antemano, R., Limbaco, M.G.J.P., Tebo, A.G., Dechavez, R., Albano, J., Rosenberg, G., Concepcion, G.P., Schmidt, E.W., Haygood, M.G.: In: Proceedings of the National Academy of Sciences (2017)
Shipway, J.R., Altamia, M.A., Rosenberg, G., Concepcion, G.P., Haygood, M.G., Distel, D.L.: Proc. R. Soc. B Biol. Sci. 286, 20190434 (2019)
O’Connor, R.M., Fung, J.M., Sharp, K.H., Benner, J.S., McClung, C., Cushing, S., Lamkin, E.R., Fomenkov, A.I., Henrissat, B., Londer, Y.Y., Scholz, M.B., Posfai, J., Malfatti, S., Tringe, S.G., Woyke, T., Malmstrom, R.R., Coleman-Derr, D., Altamia, M.A., Dedrick, S., Kaluziak, S.T., Haygood, M.G., Distel, D.L.: Proc. Natl. Acad. Sci. U. S. A. 111, E5096–E5104 (2014)
WoRMS-Editorial-Board: World register of marine species. Tech. Rep. Available from http://www.Marinespecies.Org at Vliz. (2018). Accessed 4 Apr 2018
Çevik, C., Ozcan, T., Gündoğdu, S.: BioInvasions Rec. 4, 277–280 (2015)
Cragg, S.M., Pitman, A.J., Henderson, S.M.: Int. Biodeterior. Biodegradation. 43, 197–205 (1999)
Cowles, D.: Invertebrates of the Salish Sea. Rosario Beach Marine Laboratory, Fidalgo Island, Anacortes, WA; a campus of Walla Walla University. https://inverts.wallawalla.edu/index.html (2002). Accessed 5 Apr 2018
Cragg, S.M.: In: Wood Deterioration and Preservation Acs Symposium Series Vol 845, pp. 272–286. American Chemical Society, Washington, DC (2003)
E. Shalaeva: Cabi.: In: Invasive Species Compendium. CAB International, Wallingford (2018)
Si, A., Bellwood, O., Alexander, C.G.: J. Zool. 256, 463–471 (2002)
Benson, L.K., Rice, S.A., Johnson, B.R.: Florida Acad. Sci. 62, 128–144 (1999)
Besser, K., Malyon, G.P., Eborall, W.S., Paro da Cunha, G., Filgueiras, J.G., Dowle, A., Cruz Garcia, L., Page, S.J., Dupree, R., Kern, M., Gomez, L.D., Li, Y., Elias, L., Sabbadin, F., Mohamad, S.E., Pesante, G., Steele-King, C., Ribeiro de Azevedo, E., Polikarpov, I., Dupree, P., Cragg, S.M., Bruce, N.C., McQueen-Mason, S.J.: Communications. 9, 5125 (2018)
Acknowledgments
Professor Goodell’s work on this chapter was supported by the National Institute of Food and Agriculture, U.S. Department of Agriculture, the Center for Agriculture, Food and the Environment, and the Microbiology department at University of Massachusetts Amherst, under project number S1075 - MAS00503. The contents are solely the responsibility of the authors and do not necessarily represent the official views of the USDA or NIFA.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2023 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Goodell, B., Nielsen, G. (2023). Wood Biodeterioration. In: Niemz, P., Teischinger, A., Sandberg, D. (eds) Springer Handbook of Wood Science and Technology. Springer Handbooks. Springer, Cham. https://doi.org/10.1007/978-3-030-81315-4_4
Download citation
DOI: https://doi.org/10.1007/978-3-030-81315-4_4
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-81314-7
Online ISBN: 978-3-030-81315-4
eBook Packages: Chemistry and Materials ScienceChemistry and Material Science (R0)