
Abstract
The phytohormone cytokinin was initially believed to be only involved in the process of cell division and thus it was named so, but recent studies have shown that cytokinin also plays an important role in protection of plants against various biotic and abiotic stress developed due to inappropriate environmental conditions. Cytokinin is a purine-based compound having a substitution at N6 position. Cytokinin plays a vital role during abiotic stress conditions such as drought, salinity, high light intensity, temperature extremes, heavy metal toxicity and nutrient deficient conditions. Interaction of cytokinin-mediated signalling system with other stress hormones enable plants to enhance the function of endogenous protective machineries such as enzymatic antioxidants which ultimately protect plants and maintain their cellular structure and homeostatic condition under severe environmental stress. In case of biotic stress, plants reduce their cytokinin level in order to defend themselves against pathogen infection. Higher level of cytokinin induces cell division and accumulation and allocation of nutrients and reduces the rate of senescence of leaves. Thus, it can be concluded that during biotic stress, cytokinin acts a negative regulator, and by decreasing the level of cytokinin, the damages caused during biotic stress can be restricted in plants. In this chapter, we mainly focus on the biosynthesis and signalling pathway of cytokinin in plants and then demonstrate the role of cytokinin and its interaction with different stress hormones under several biotic and abiotic stress conditions.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Cytokinin
- Histidine kinase
- Histidine phosphotransferase
- Response regulator
- Environmental stress
- Isopentenyl transferases
- Cytokinin dehydrogenases
6.1 Introduction
Plants are sessile living organisms which cannot escape unfavourable environment by shifting their positions. Harsh environmental conditions are negative factors which adversely affect the growth, biomass and yield of plants. Stress faced by plants may be due to the invading pathogens or infestations of insects collectively known as biotic stress and abiotic stress which occurs due to hostile environmental conditions like drought, salinity, extreme temperature or heavy metal toxicity (Roychoudhury et al. 2013). Harsh conditions lead to oxidative burst in plant tissues which ultimately enhances the formation of reactive oxygen species (ROS) like hydrogen peroxide (H2O2), superoxide (.O2−) and hydroxyl radicals (.OH). These ROS lead to cellular membrane damages and formation of toxic products like malondialdehyde (MDA) and methylglyoxal (Banerjee and Roychoudhury 2018). Plants have several mechanisms which help them adapt and survive under unfavourable conditions. Production of various endogenous signalling molecules helps to coordinate and enhance the functions of the internal defensive pathways which ultimately induce their survival capability under severe environmental conditions.
In addition to the action of other stress hormones such as abscisic acid (ABA), jasmonate and salicylic acid, cytokinin (CK) also plays an important role in regulating the action of protective machineries of plants (Ha et al. 2012). CKs were initially believed to be only involved in cell division and in the regulation of the cell cycle (Schaller et al. 2014), but later it was found that CKs also govern various other functions like inhibition of root growth, maintenance of apical dominance, formation of shoot meristem, growth of lateral buds, expansion and senescence of leaves and nitrogen signalling pathways (Frebort et al. 2011; Giulini et al. 2004; Miyawaki et al. 2004; Nooden et al. 1990). Analysis of the interactive pathways involved between CKs and environmental stress has recently begun and is now well characterised (O’Brien and Benkova 2013; Ha et al. 2012).
CKs are derivatives of adenine residue with a substitution at N6 position by isoprenoid or aromatic acid residues (Sakakibara 2006). Based on the reduction or hydroxylation of the side chains, a wide variety of derivatives of CKs are found in nature such as cis-zeatin, trans-zeatin, isopentenyladenine and dihydrozeatin known as isoprenoid CKs . N6-(meta-hydroxybenzyl) adenine is an aromatic CK found in less quantity in plants (Faiss et al. 1997). CKs in plants are mostly found in immature seeds, meristematic regions and young leaves. Endogenous level of CK in plants is maintained by the activity of various enzymes which are mainly responsible for biosynthesis of CKs or alteration of one member of CK family to other members (Letham 1994). Expression of targeted genes can be regulated by CK through the action of a two-component system where transfer of phosphate unit between Asp and His residues helps to transduce the signal which ultimately facilitates plants to provide suitable response against stimuli (Huang et al. 2012).
In this chapter, we summarise the emerging role of CKs in plant systems when exposed to different stress conditions. We begin with the metabolism and signalling pathway of CKs in plants and then cover the different aspects of CKs under various abiotic stresses like drought, salinity, extreme temperature, light, nutrient deficiency and heavy metal stress. We also provide an overview of protective role of CKs under different biotic stresses. For each of these stress conditions, we present an extensive analysis on the role of CK signalling pathway and its interaction with other stress hormones and metabolites which are involved in enhancing the tolerance limit of plants against harsh environmental conditions.
6.2 Metabolic Pathway of CKs
An interesting feature of the CKs is that they are made of purine and differ from adenine and other members of CK family by the presence of various chemical groups at N6 position which also determines their biological activity, transportation and presence in the cells or tissues of plants (Hirose et al. 2008). The two major forms of CKs present in Arabidopsis are trans-zeatin and isopentenyladenine (Kiba et al. 2013) which shows acropetal and basipetal movement, respectively, during long-range transportation (Matsumoto-Kitano et al. 2008). CK receptors also have distinctive binding affinity to different CKs involved in the signalling pathway in Arabidopsis (Stolz et al. 2011; Romanov et al. 2006). Cis-zeatin, an isoform of trans-zeatin, is less active and is found in some monocots and dicots, and its expression is mostly dependent on the growth stage of the plants (Frebort et al. 2011). Dihydrozeatin which is formed in Phaseolus vulgaris by the enzymatic action of zeatin reductase on trans-zeatin (Martin et al. 1989) was found in apical buds and seeds (Mok et al. 1990). In addition, aromatic CKs are also present in plants such as tomato (Nandi et al. 1989), Arabidopsis (Tarkowska et al. 2003), pea (Gaudinova et al. 2005) and Populus (Strnad 1997) but are less studied, and their presence and activity need to be analysed in other crops. Inactive forms of CKs are synthesised by de novo pathway, and phosphoribohydrolase activity by Lonely Guy (LOG) enzyme is required to make free active base CKs (Sakakibara 2010). Interconversion between active and inactive forms of CKs can be done by the application of adenosine kinase (Schoor et al. 2011). Plants can regulate the activity of N- and O-glucosyltransferases which are involved in the glucosylation of CKs, producing their bioactive forms (Veach et al. 2003).
It was earlier considered that CKs were formed in the roots of plants and transported upward to the shoot, but recent studies have shown that CKs are also locally synthesised in the plant cells. Different members of CKs are present in different plant tissues and in different species which affirms that different CKs have different functions (Hirose et al. 2008), for example, members of isopentenyladenine family of CKs are more found in the seeds of oil palm (Huntley et al. 2002), whereas dihydrozeatin and zeatin are mostly found in soybean seeds (Singh et al. 1988). In addition, members of trans-zeatin family are mostly found in xylem sap, and the members of isopentenyladenine and cis-zeatin family are predominantly present in phloem sap (Hirose et al. 2008). The concentration of CKs is also mediated by the interaction of plants with extrinsic factors such as biotic and abiotic stress (Schafer et al. 2014; Miyawaki et al. 2004; Hashem 2013).
The metabolic pathway of CKs can be divided into three phases: synthesis of CKs by isopentenyltransferase (IPT) enzymes, activation of the synthesised inactive forms by the activity of LOG enzymes and degradation by CK dehydrogenases (CKX) . Miyawaki et al. (2004) reported that the presence of AtIPT in Arabidopsis is required for the production of tRNA- and ATP/ADP-IPTs. IPT produces isopentenyladenine using AMP, ADP or ATP as isoprenoid acceptors for isopentenylation and dimethylallyl pyrophosphate (DMAPP) as the side chain donor generated from methylerythritol phosphate and mevalonate pathways (Sakakibara 2006). Synthesis of isopentenyladenine is initiated from its nucleoside and nucleotide precursors, whereas trans-zeatin are produced either using isopentenyladenine directly or indirectly using the immediate precursors of isopentenyladenine. Sakakibara (2010) reported another pathway involved in the synthesis of trans-zeatin by the activity of isopentenyltransferase using hydroxylated terpenoid as a side chain donor. Isopentenylation of tRNA by AtIP2 and AtIP9 leads to the formation tRNA-IPTs (Miyawaki et al. 2006). Decomposition of isopentenylated tRNAs leads to the formation of cis-zeatin in a very low amount due to longer half-life of tRNAs (Klambt 1992). Bassil et al. (1993) reported that isomerisation of trans-zeatin to cis-zeatin in the presence of a catalyst occurs in beans. Members of the dihydrozeatin family are synthesised from zeatin by zeatin reductase (Gaudinova et al. 2005).
LOG enzyme plays a major role in the activation of isopentenyladenine and trans-zeatin. Phosphoribohydrolase activity of LOG enzymes on the nucleotide 5′-monophosphates unit of CKs is essential for their function (Frebort et al. 2011). CKs may be activated by the LOG-dependent pathway which is considered as the major pathway for CK activation or by the two-step pathway which is independent of LOG enzyme (Tokunaga et al. 2012). LOG independence is not well characterised, and very few studies have been conducted which can elaborate the mechanism of this pathway (Thu et al. 2017). Tokunaga et al. (2012) reported that seven genes encoding LOG enzymes are present in Arabidopsis, where most of the functions are performed by LOG7 enzymes. Other homologs of LOG enzymes have been recently discovered in plants like Prunus persica and Populus trichocarpa (Immanen et al. 2013).
Catabolism of CKs takes place by addition of a sugar moiety by glycosyltransferase reaction or by irreversible dehydrogenation (Zalabak et al. 2013). CKXs cleave the N6 moiety of trans- and cis-zeatin and isopentenyladenine, whereas the aromatic side chain of CKs is cleaved at a lower rate; dihydrozeatin are non-cleavable by this enzyme (Galuszka et al. 2007). CKXs belong to a small family, containing only seven members which are species-specific, and play an essential role in maintaining the homeostasis of the cells (Bilyeu et al. 2001). In Arabidopsis, cis-zeatin is mostly cleaved by AtCKX 1 and AtCKX 7, whereas AtCKX 2, AtCKX 3 and AtCKX 4 cannot hydrolyse it (Gajdosova et al. 2011). Another way to maintain the level of CKs in the cells is by glycosylation. O-glucosyltransferases and O-xylosyltransferases are involved in reversible glycosylation of CK residues to O-glucosylated and O-xylosylated forms, respectively. N-glucosyltransferases produce N9-glucosylated or N7-glucosylated CK residues which cannot be degraded by β-glucosidases (Thu et al. 2017).
6.3 CK-Mediated Signalling Pathway and Its Components
6.3.1 Signal Transduction by CKs
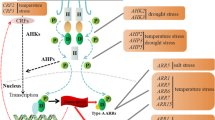
CK signalling pathway is a multistep process which involves the phosphorylation of histidine kinase (HK) protein acting as a receptor, followed by the transfer of signal by histidine phosphotransfer (HP) protein and finally receiving the signal and accordingly responding by an ultimate response regulator (RR) (Thu et al. 2017). The signalling pathway starts when stimuli leads to the formation of the CK molecules which binds to the cyclase/histidine kinase associated sensory extracellular (CHASE) domain of the HK receptor. This binding leads to the transfer a phosphoryl group from the histidine residue to the aspartate molecules present in the receptors. The histidine residue present in HP protein then receives the phosphoryl group which ultimately transfers the residue to the aspartate residue of the RR which helps to initiate the transcription of the targeted genes (Fig. 6.1). The signalling pathway is dependent on the HK receptors because it has been reported in Arabidopsis that HK receptors are highly specific to the substituted side chain residues of the CKs (Hwang et al. 2012).

Two-component CK signalling system. The HK receptor for CKs which is activated in plants due to external stimuli is mainly located on the membrane of cells and endoplasmic reticulum. The pathway is initiated by the binding of CKs to the CHASE domain of HK receptor (1), which then gets phosphorylated at the histidine residue and ultimately transfers it to the aspartate residue at the C-terminal of HK receptors (2). The phosphate molecule is then transferred to the HP protein (3) which enters into the nucleus (4). Within the nucleus, it transfers the phosphate molecule to aspartate residue present on the receiver domain of either type-A or type-B RR (5). Type-A RR acts as inhibitor of type-B RR and negatively regulates the HK receptors, whereas type-B RR via its effector domain regulates the gene transcription (6) or other responses mediated by CK signalling (7)
6.3.2 Components of the Signalling Pathway
6.3.2.1 Histidine Kinases (HK)
HK are multi-domain complex receptors of CKs. A CHASE domain present at the N-terminal acts as a sensor domain for CKs which is followed by two or more transmitter domain and lastly two receiver domains having HK activity (Lomin et al. 2012). CKs bind to the CHASE domain which leads to the conformation change of the sensors that initiate the autophosphorylation of the transmembrane domain and finally transferring the phosphoryl group to the receiver domain (Lomin et al. 2012).
6.3.2.2 Histidine Phosphotransfer (HP)
HP proteins are 17 kDa phosphotransmitter proteins which act as a transmitter of signal from HK to RR by transferring the phosphate groups (Shi and Rashotte 2012). Five Arabidopsis HPs have been identified by Suzuki et al. (1998). Imamura et al. (2001) reported that HPs are present in cytoplasm and also demonstrated their phosphotransfer activity in vitro. Formation of protoxylem is mediated by another type of HP which acts as an inhibitor of the CK signalling pathway (Mahonen et al. 2006).
6.3.2.3 Response Regulator (RR)
There are 23 RRs present in Arabidopsis which can be categorised under four types, type-A, type-B, type-C and pseudo-RRs, based on their domain structure and protein sequences (Mizuno and Nakamichi 2005). Addition of a phosphate group to type-A RR is necessary to stabilise it, whereas phosphorylation of type-B RR is required to enhance their binding ability to DNA that ultimately regulates the expression of target genes and also induces the stability of type-A RR (To et al. 2007). Type-A RR negatively regulates the pathway due to the presence of only a receiver domain (To et al. 2004), whereas type-B RR consists of both an output domain having a glutamine-rich domain which enables them to bind to DNA and a receiver domain (Sakai et al. 2001). Gupta and Rashotte (2012) reported that in Arabidopsis, some of the genes which are targeted by type-B RR are under the regulation of type-A RR which shows that signalling mediated by CKs is self-regulated.
6.4 Role of CK Signalling System in Response to Various Abiotic Stresses
Researches differ regarding their view about the role of CKs in plants against stress conditions. Several studies have shown that CKs act as negative regulators in plants in initiating tolerance mechanisms against environmental stress (Ghanem et al. 2008; Hansen and Dorffling 2003; Albacete et al. 2008; Kudoyarova et al. 2007), whereas several other studies have reported a short-term induction in the level of CKs against stress conditions (Alvarez et al. 2008; Pospisilova et al. 2005; Walker and Dumbroff 1981). Nishiyama et al. (2011) have reported higher activity of IPT in Arabidopsis for 1–2 hours, whereas the activity of CKXs was lowered for 1 hour after plants were exposed to salt stress. Overall, it can be concluded that initial exposure of plants to stress conditions leads to higher production of CKs which again gradually decrease to the normal level in the course of time.
6.4.1 Drought Stress
Drought stress is experienced by plants due to inadequate or insufficient supply of water which leads to chlorophyll degradation and enhanced production of ROS resulting in higher membrane damage (Nadeem et al. 2019). In addition, drought stress also reduces growth of plants by reducing cell division which results in lower turgor pressure caused by limited water uptake from the surrounding environment (Bhatt and Rao 2005).
Separate studies have reported that drought stress reduces the endogenous content of CKs in plants (Todaka et al. 2017; Nishiyama et al. 2011). Similar effect was noted in Arabidopsis where drought stress reduces the expression of IPT and induces the expression of CKXs (Le et al. 2012; Ramireddy et al. 2014). Lowered formation of CKs and thus reduced signalling under drought enhance the formation and sensitivity of plants to ABA (Tran et al. 2007), since ABA is antagonistic to CKs. In addition, reduced level of CKs lowered the shoot length (Werner et al. 2003). Higher synthesis of ABA and reduced shoot length are two known adaptive responses of plants against drought stress (Roychoudhury and Paul 2012).
Plants in which CK signalling pathway was suppressed by mutating the HK receptors or lowering the transfer of phosphate group to HP or by lowering the function of RR showed higher tolerance to drought stress (Nguyen et al. 2016). They also reported that CKs also control the transcription of a large number of ABA-responsive, drought/dehydration genes which enhances the tolerance capacity of plants under water-deficit conditions. Contradictory to these studies, Tran et al. (2007) and Susan et al. (2013) reported that expression of HK in Arabidopsis and maize plants, respectively, was upregulated under drought stress. The expression of RR6 was also upregulated in response to drought stress or exogenous application of CKs, suggesting that RR6 may play an important role in improving the yield of rice plants under drought stress (Panda et al. 2018). In addition to the negative role of RR22, recent studies have also demonstrated the positive role of RR22 by maintaining the integrity of the cell membrane and enhancing the expression of drought-responsive gene in Arabidopsis (Kang et al. 2013). Higher expression of IPT gene enhances the endogenous content of CKs which reduces the effect of drought stress in plants like cotton (Kuppu et al. 2013), tropical maize (Bedada et al. 2016), eggplant (Xiao et al. 2017) and bentgrass (Xu et al. 2016). CKs also negatively affect the water content of plants by lowering root length and increasing stomatal density and conductance. Overexpression of DREB6.2 genes ultimately induces the expression of CKX4a which reduces the endogenous content of CKs to produce transgenic apples which were resistant against drought stress (Liao et al. 2017). Production of ROS is one of the major damaging effects of drought stress in plants. Mytinova et al. (2011) reported that overexpression of CKX2 from Arabidopsis in tobacco plants reduces the endogenous content of CKs in the cells which results in a higher activity of superoxide dismutase, glutathione reductase and ascorbate peroxidase lowering the level of ROS in transgenic plants. Contradictory results were observed in bentgrass and eggplants where overexpression of IPT gene induces the endogenous content of CKs which reduces the level of ROS formed in the cells by enhancing the activity of superoxide dismutase, ascorbate peroxidase, dehydroascorbate reductase and catalase (Xu and Huang 2017). Thus, the actual signalling process of CKs varies in different plant species when exposed to water-deficit conditions.
6.4.2 Salinity Stress
High salt concentration in the soil is one of the major abiotic stresses experienced by plants. Cell membrane damage reduces the ability to detoxify the ROS content. ROS accumulation and nutrient imbalance are some of the initial symptoms observed in plants when exposed to salt stress (Rahnama et al. 2010). During salt stress, excess deposition of Na+ ions in the tissues of plants affects the ion balance which may interfere with signalling process (James et al. 2011; Roychoudhury and Chakraborty 2013; Roychoudhury et al. 2013). High salt concentration also interferes with the nutritional uptake from the soil, especially uptake of K+ required for growth and productivity of plants.
Salinity stress in plants is mostly regulated by ABA. Since ABA acts antagonistically to CKs, it can be concluded that CKs act as a negative regulator of salt stress in plants (Cortleven et al. 2019). Reduced expression of IPT gene leads to lower formation of CKs in Arabidopsis which was able to adapt more efficiently against salt-stressed environment as compared to wild-type plants (Nishiyama et al. 2011). Plants better adapted against salt stress also showed altered expression of genes which were regulated by ABA. Similar observation was also reported by Wang et al. (2015), who showed that overexpression of IPT8 gene in Arabidopsis leads to higher formation of CKs which ultimately enhances the symptoms of damage in plants maintained under high salt concentration. High-affinity K+ transporter 1;1 (HKT1;1) is present in Arabidopsis which is responsible for removal of Na+ ions during salt stress in plants. CKs negatively regulates the activity of HKT1;1. Mason et al. (2010) reported that reduced endogenous content of CKs enhanced the expression of HKT1;1 which removed excess Na+ ions accumulated in the cells during salinity stress ultimately improving the survival capability of plants.
Along with the negative role , positive effect of CKs is also reported against salt stress. Keshishian et al. (2018) reported that salt stress in tomato plants enhanced the formation of CKs which positively affects the tolerance capability of plants. A similar effect in tomato was also demonstrated by Aremu et al. (2014) where they showed that application of INCYDE which acts as a potent inhibitor of CKX and thus increases the CK content of the cells results in better production of flowers and also protected the photosynthetic apparatus. Salt-sensitive cultivars of rice lacking CKX2 showed higher tolerance against salt stress which was ultimately reflected in their grain yield (Joshi et al. 2018).
6.4.3 Extreme Temperature
Plants require optimum temperature for their growth and maximum productivity. Any deviation from this optimal range may cause molecular and physiological damage and can also reduce the yield of plants. Cold stress leads to retardation in development, chlorosis and necrosis of cells and also causes sterility of flowers as noted in rice plants (Jiang et al. 2002). Low temperature causes shortage of water in plants affecting the integrity of plasma membrane (Steponkus et al. 1993). Temperature above the optimal range induces the formation of ROS causing oxidative stress in plants. Collapse of cellular structure in plants may occur when exposed to very high temperature which leads to cell death (Ahuja et al. 2010). High temperature also interferes with the metabolic process by altering the protein structures of the enzymes (Lobell et al. 2011).
The effect of CKs in plants is still uncertain. Maruyama et al. (2014) reported that cold stress decreases the endogenous level of CKs and CK signalling, which reduces the size of plants. Arabidopsis containing mutant receptors such as ahk2, ahk3 and ahk3 cytokinin response element (cre)1 was highly adapted against cold stress, proving that CKs play a negative role during regulation of cold stress (Cortleven et al. 2019). Jeon et al. (2010) reported the activation of CK receptors, and Jeon and Kim (2012) observed that several HP and RR can be activated in the absence of CKs. Higher level of cold tolerance in Arabidopsis having mutant rr5, rr6 and rr7 was observed (Jeon et al. 2010). In contrast to the above observation, Shi et al. (2012) reported that overexpression of RR genes such as RR22, RR15, RR7 and RR5 enhanced the cold adaptability of Arabidopsis plants. Along with the classical signal pathway of CKs, several CK response factors (CRFs) have also been linked with protection of plants against cold stress. Jeon et al. (2016) reported that CRF2 expression can initiate the formation of lateral roots which is directly controlled by RR1 in Arabidopsis. Zwack et al. (2016) reported that the expression of CRF4 can be induced by cold stress, and it acts as a positive regulator of freeze tolerance. Thus, from the above observation, it can be concluded that the activation of the two-component signalling system either in presence or absence of CKs may play an important role in providing protection against cold stress in plants.
Exogenous application of CKs enhances the adaptability of plants against high temperature, whereas excess heat reduces the endogenous level of CKs in the cells (Cortleven et al. 2019). External application of CKs reduces the effect of heat in plants by protecting their photosynthetic machineries (Liu and Huang 2002), increasing the thermo-tolerance of the reproductive tissues in plants like rice, passion fruit and maize (Sobol et al. 2014; Wu et al. 2016; Cheikh and Jones 1994) and enhancing the activity of heat shock protein and antioxidative system in bentgrass (Xu et al. 2010). Skalak et al. (2016) reported that high temperature induces the degradation of endogenous CKs of the cells which helps in the stomatal closure in Arabidopsis.
6.4.4 Light Stress
Light is responsible for the survival of life on earth, but excess light also causes damage to the photosynthetic apparatus and bleaching of chlorophyll. Solar radiation is the only source of natural light on earth which can be utilised by plants for photosynthesis, but it also contains ultraviolet light which can damage or mutate the DNA of the cells (Jansen et al. 1998). Excess light can inhibit photosynthesis which leads to excess production of ROS causing damage to the cellular membrane and its components.
Recent studies have demonstrated a direct link between CKs and light through photoreceptor phy B (Chi et al. 2016; Sweere et al. 2001). Plants having a higher level of CKs show better adaptability against light stress and protect the D1 reaction centre of photosynthesis which is severely damaged by excess light (Cortleven et al. 2014). They also reported that CK-mediated signalling system is mostly controlled by HK3 and HK2 and type-B RRs such as RR1 and RR12 in Arabidopsis. Other investigations observed that the components of antioxidative system protect photosynthetic apparatus under different stresses like drought (Rivero et al. 2009). This was used as evidence by Cortleven and Schmulling (2015) to support their observation that CKs can protect photosynthetic apparatus in plants when exposed to high light. Low light also acts as a stress stimulus causing an induction in response mechanism in plants. In tobacco and Arabidopsis, CKs act as an important signalling molecule transported across the xylem tissue which helps plants adapt in low light environment (Boonman et al. 2009). Carabelli et al. (2007) reported that CKX induces the degradation of CKs in plants which inhibits the formation of leaf primordia, ultimately promoting growth of hypocotyl towards light.
6.4.5 Nutrient Stress
Besides carbon , oxygen and hydrogen which plants easily accumulate from CO2 and H2O present in the surrounding environment, plants also require several micronutrients (boron, silicon, iron and selenium) and macronutrients (potassium, nitrogen and phosphorus) for their normal growth and development. CK plays a pivotal role during uptake of these nutrients from the soil.
Boron is an essential micronutrient whose deficiency can lead to the downregulation of genes involved in CK-mediated signalling (Abreu et al. 2014). Pavlu et al. (2018) observed that boron transporter (BOR)4, a gene which codes for boron transporter, has an expression pattern similar to that of RR1 and LOG7 genes. In oilseed rape, a strong correlation was observed between boron concentration and formation and activation of CKs (Eggert and von Wiren 2017). Silicon is the major soil component, and its deposition leads to enhanced activation of various defensive machineries in plants; it also provides structural support to various tissues of plants. Markovich et al. (2017) reported that silicon deposition in the tissues of Arabidopsis and sorghum enhanced the synthesis of CKs by upregulating the expression of IPT7 gene and reduced the process of leaf senescence induced by dark stress via activating the CK signalling pathway. Selenium induces the stress tolerance capability of plants when present in lower concentration (Pilon-Smits et al. 2009). The two major forms of selenium, viz. selenite and selenate, are absorbed by plants through phosphate and sulphate transporters (Schiavon and Pilon-Smits 2017). Thus, it can be said that the pathway responsible for phosphate and sulphate uptake in plants, mediated by CK signalling, acts as a common pathway for the uptake of selenium from the soil.
Nitrogen acts as a major element during the synthesis of CKs. In Arabidopsis, the enzymatic activity of IPT3 and IPT5 which catalyses the rate-limiting step of CK synthesis is controlled by the availability of nitrogen during the production of cytochrome P450 and trans-zeatin (Kiba et al. 2013; Kieber and Schaller 2014). CKs transported to the shoot via the root control the nitrate response and key traits such as activity-related traits of meristem (Muller et al. 2015) and leaf size (Rahayu et al. 2005). The activity of IPT3 is also mediated by the presence of inorganic phosphate (Pi), and thus the expression of IPT3 is downregulated in the absence of Pi (Hirose et al. 2008). Supply of Pi in plants, after facing a situation of phosphorus deficiency, leads to enhanced expression of CRF5, CRF6 and IPT3 (Woo et al. 2012) Exogenous application of CKs lowers the uptake and response of Pi deficiency in Arabidopsis and rice (Franco-Zorrilla et al. 2002) by utilising the stored Pi from the internal sources which ultimately reduces the Pi deficiency signalling and symptoms (Lai et al. 2007). Potassium is one of the major macronutrients for plants, and it is added externally in the soil as fertilizer along with nitrogen and phosphorus. Potassium deficiency in Arabidopsis lowers the endogenous CK level which results in inhibition of CK-mediated signalling system, ultimately reducing the root length and inducing potassium uptake from the soil (Nam et al. 2012). They also reported that CK deficiency leads to higher accumulation of ROS and enhanced root hair growth along with higher expression of HAK5 gene which codes for a transporter involved in the uptake of potassium from the surroundings. Hirose et al. (2008) showed that CK formation by IPT3 is directly associated with sulphur deficiency in soil. They also reported that exogenous application of CK enhanced the expression of genes which were regulated by the presence of sulphur, whereas Maruyama-Nakashita et al. (2004) reported that the expression of sulphate transporter in roots was downregulated by CKs .
6.4.6 Heavy Metal Stress
Tight regulation is important for transport and absorption of heavy metals , since higher deposition of heavy metal in plant tissues damages the cell structure and leads to oxidative stress. Along with desired element such as macronutrients and micronutrients, plants also uptake several metals and metalloids like mercury, cadmium, chromium, arsenic and lead which are toxic for plants. Bruno et al. (2017) and Yang et al. (2017) reported that Arabidopsis adapt themselves against cadmium and aluminium toxicity by enhancing the level of CKs along with signalling system mediated by CKs which hamper the growth of the roots. Werner et al. (2010) reported that plants which are deficient in CKs accumulate a higher amount of cadmium in their tissues, whereas a contrasting result was observed in the case of arsenate toxicity where plants were able to tolerate arsenate stress in CK-deficient conditions (Mohan et al. 2016). CK enhances the activity of glutathione-S-transferase (Brenner and Schmulling 2015), which induces the production of thiol compounds which can be utilised by plants against arsenate toxicity (Mohan et al. 2016).
6.4.7 Biotic Stress
Biotic stress in plants is mostly caused by the infection of pathogens like virus, fungus and bacteria or by rodent and insects which severely affect the yield of plants. In recent times, broad studies have been conducted on the role of CKs with regard to such biotic stresses (Albrecht and Argueso 2017). Walters and McRoberts (2006) reported that during biotrophic fungal infection, CKs help the pathogens to multiply within the infected host. CK helps in the formation of green islands which cause the assimilation of nutrition at the infected site and also delay senescence of the leaf. Not only that, CK at the infected site also inhibits photosynthetic metabolism by enhancing the activity of invertase. Infection of plants with geminiviruses hinders the activity of various adenosine kinases (Baliji et al. 2010). The viral infection got boosted up due to higher expression of genes involved in primary CK formation. Increased level of CK helps geminiviruses by promoting cell division, availability of resources and inhibiting senescence of infected leaves. Radhika et al. (2015) reported that Rhodococcus fascians, an actinomycete which produces a wide range of methylated CKs, is more stable against plant CKX and helps the pathogen grow more quickly by inducing the cell division of host plants. Kind et al. (2018) observed that the virulence of Claviceps purpurea can be reduced by expressing the CK synthesis and CKX gene in the pathogen. Various other studies conducted by several groups such as Siemens et al. (2006), Spallek et al. (2017) and Siddique et al. (2015) have also shown the negative effect of CK in plants under different biotic stresses.
6.5 Conclusion and Future Perspectives
It is evident from our discussion that CKs play an important role in abiotic and biotic stress signalling in plants. Manipulating the signalling process involved during stress condition can be beneficial for sustainable growth of plants mostly in the case of biotic stress where lower level of CK helps host plants to better inhibit the growth of the pathogen infection by either degrading the endogenous content of CKs via enhancing the activity of CKX or by lowering the production of CKs via downregulating the activity of the enzymes like IPT. CKs behave differently for different abiotic stresses, and thus it becomes necessary in the near future to further elaborate CK pathways for better understanding the role of CKs against harsh environmental conditions. Though the interaction of CKs with different stress hormones such as ABA or salicylic acid is known, further studies will help to expand our knowledge about the interaction between these hormones which can be applied in genetic engineering to develop stress-tolerant crop plants with better yield. Thus, it can be concluded that CKs are essential signalling molecules which enhance co-ordination and interaction among different pathways under various biotic and abiotic stresses.
References
Abreu I, Poza L, Bonilla I et al (2014) Boron deficiency results in early repression of a cytokinin receptor gene and abnormal cell differentiation in the apical root meristem of Arabidopsis thaliana. Plant Physiol Biochem 77:117–121
Ahuja I, de Vos RCH, Bones AM et al (2010) Plant molecular stress responses face climate change. Trends Plant Sci 15:664–674
Albacete A, Ghanem ME, Martinez-Andujar C et al (2008) Hormonal changes in relation to biomass partitioning and shoot growth impairment in salinized tomato (Solanum lycopersicum L.) plants. J Exp Bot 59:4119–4131
Albrecht T, Argueso CT (2017) Should I fight or should I grow now? The role of cytokinins in plant growth and immunity and in the growth-defence trade-off. Ann Bot 119:725–735
Alvarez S, Marsh EL, Schroeder SG et al (2008) Metabolomic and proteomic changes in the xylem sap of maize under drought. Plant Cell Environ 31:325–340
Aremu AO, Masondo NA, Sunmonu TO et al (2014) A novel inhibitor of cytokinin degradation (INCYDE) influences the biochemical parameters and photosynthetic apparatus in NaCl-stressed tomato plants. Planta 240:877–889
Baliji S, Lacatus G, Sunter G (2010) The interaction between geminivirus pathogenicity proteins and adenosine kinase leads to increased expression of primary cytokinin-responsive genes. Virology 402:238–247
Banerjee A, Roychoudhury A (2018) Abiotic stress, generation of reactive oxygen species, and their consequences: an overview. In: Singh VP, Singh S, Tripathi D, Mohan-Prasad S, Chauhan DK (eds) Revisiting the role of reactive oxygen species (ROS) in plants: ROS boon or bane for plants? Wiley, Hoboken, pp 23–50
Bassil NV, Mok DW, Mok MC (1993) Partial purification of a cis-trans-isomerase of zeatin from immature seed of Phaseolus vulgaris L. Plant Physiol 102:867–872
Bedada LT, Seth MS, Runo SM et al (2016) Drought tolerant tropical maize (Zea mays L.) developed through genetic transformation with isopentenyltransferase gene. Afr J Biotechnol 15:2447–2464
Bhatt RM, Rao NKS (2005) Influence of pod load response of okra to water stress. Indian J Plant Physiol 10:54–59
Bilyeu KD, Cole JL, Laskey JG et al (2001) Molecular and biochemical characterization of a cytokinin oxidase from maize. Plant Physiol 125:378–386
Boonman A, Prinsen E, Voesenek LACJ et al (2009) Redundant roles of photoreceptors and cytokinins in regulating photosynthetic acclimation to canopy density. J Exp Bot 60:1179–1190
Brenner WG, Schmulling T (2015) Summarizing and exploring data of a decade of cytokinin-related transcriptomics. Front Plant Sci 6:29
Bruno L, Pacenza M, Forgione I et al (2017) In Arabidopsis thaliana cadmium impact on the growth of primary root by altering SCR expression and auxin-cytokinin cross-talk. Front Plant Sci 8:1323
Carabelli M, Possenti M, Sessa G et al (2007) Canopy shade causes a rapid and transient arrest in leaf development through auxin-induced cytokinin oxidase activity. Genes & Development 21:1863–1868
Cheikh N, Jones RJ (1994) Disruption of maize kernel growth and development by heat stress (role of cytokinin/abscisic acid balance). Plant Physiol 106:45–51
Chi W, Li J, He B et al (2016) DEG9, a serine protease, modulates cytokinin and light signaling by regulating the level of Arabidopsis Response Regulator 4. Proc Natl Acad Sci U S A 113:3568–3576
Cortleven A, Schmulling T (2015) Regulation of chloroplast development and function by cytokinin. J Exp Bot 66:4999–5013
Cortleven A, Nitschke S, Klaumunzer M et al (2014) A novel protective function for cytokinin in the light stress response Is mediated by the Arabidopsis Histidine Kinase 2 and Arabidopsis Histidine Kinase 3 Receptors. Plant Physiol 164:1470–1483
Cortleven A, Leuendorf JE, Frank M et al (2019) Cytokinin action in response to abiotic and biotic stresses in plants. Plant Cell Environ 42:998–1018
Eggert K, von Wiren N (2017) Response of the plant hormone network to boron deficiency. New Phytol 216:868–881
Faiss M, Zalubilova J, Strnad M et al (1997) Conditional transgenic expression of the ipt gene indicates a function for cytokinins in paracrine signaling in whole tobacco plants. Plant J 12:401–415
Franco-Zorrilla JM, Martin AC, Solano R et al (2002) Mutations at CRE1 impair cytokinin-induced repression of phosphate starvation responses in Arabidopsis. Plant J 32:353–360
Frebort I, Kowalska M, Hluska T et al (2011) Evolution of cytokinin biosynthesis and degradation. J Exp Bot 62:2431–2452
Gajdosova S, Spichal L, Kaminek M et al (2011) Distribution, biological activities, metabolism, and the conceivable function of cis-zeatin-type cytokinins in plants. J Exp Bot 62:2827–2840
Galuszka P, Popelkova H, Werner T et al (2007) Biochemical characterization of cytokinin oxidases/dehydrogenases from Arabidopsis thaliana expressed in Nicotiana tabacum L. J Plant Growth Regul 26:255–267
Gaudinova A, Dobrev PI, Solcova B et al (2005) The involvement of cytokinin oxidase/dehydrogenase and zeatin reductase in regulation of cytokinin levels in pea (Pisum sativum L.) leaves. J Plant Growth Regul 24:188–200
Ghanem ME, Albacete A, Martinez-Andujar C et al (2008) Hormonal changes during salinity-induced leaf senescence in tomato (Solanum lycopersicum L.). J Exp Bot 59:3039–3050
Giulini A, Wang J, Jackson D (2004) Control of phyllotaxy by the cytokinin-inducible response regulator homologue ABPHYL1. Nature 430:1031–1034
Gupta S, Rashotte AM (2012) Down-stream components of cytokinin signaling and the role of cytokinin throughout the plant. Plant Cell Rep 31:801–812
Ha S, Vankova R, Yamaguchi-Shinozaki K et al (2012) Cytokinins: metabolism and function in plant adaptation to environmental stresses. Trends Plant Sci 17:172–179
Hansen H, Dorffling K (2003) Root-derived trans-zeatin riboside and abscisic acid in drought-stressed and rewatered sunflower plants: interaction in the control of leaf diffusive resistance? Funct Plant Biol 30:365–375
Hashem HA (2013) Cadmium toxicity induces lipid peroxidation and alters cytokinin content and antioxidant enzyme activities in soybean. Botany 92:1–7
Hirose N, Takei K, Kuroha T et al (2008) Regulation of cytokinin biosynthesis, compartmentalization and translocation. J Exp Bot 59:75–83
Huang GT, Ma SL, Bai LP et al (2012) Signal transduction during cold, salt, and drought stresses in plants. Mol Biol Rep 39:969–987
Huntley RP, Jones LH, Hanke DE (2002) Cytokinins and gibberellins in sap exudate of the oil palm. Phytochemistry 60:117–127
Hwang I, Sheen J, Muller B (2012) Cytokinin signaling networks. Annu Rev Plant Biol 63:353–380
Imamura A, Yoshino Y, Mizuno T (2001) Cellular localization of the signaling components of Arabidopsis his-to-asp phosphorelay. Biosci Biotech Bioch 65:2113–2117
Immanen J, Nieminen K, Silva HD et al (2013) Characterization of cytokinin signaling and homeostasis gene families in two hardwood tree species: Populus trichocarpa and Prunus persica. BMC Genomics 14:885
James RA, Blake C, Byrt CS et al (2011) Major genes for Na+ exclusion, Nax1 and Nax2 (wheatHKT1;4 and HKT1;5), decrease Na+ accumulation in bread wheat leaves under saline and waterlogged conditions. J Exp Bot 62:2939–2947
Jansen MAK, Gaba V, Greenberg BM (1998) Higher plants and UV-B radiation: balancing damage, repair and acclimation. Trends Plant Sci 3:131–135
Jeon J, Kim NY (2012) Arabidopsis response Regulator1 and Arabidopsis histidine phosphotransfer Protein2 (AHP2), AHP3, and AHP5 function in cold signaling. Plant Physiol 161:408–424
Jeon J, Kim NY, Kim S et al (2010) A subset of cytokinin two-component signaling system plays a role in cold temperature stress response in Arabidopsis. J Biol Chem 285:23371–23386
Jeon J, Cho C, Lee MR et al (2016) CYTOKININ RESPONSE FACTOR2 (CRF2) and CRF3 regulate lateral root development in response to cold stress in Arabidopsis. Plant Cell 28:1828–1843
Jiang QW, Kiyoharu O, Ryozo I (2002) Two novel mitogen-activated protein signalling components, OsMEK1 and OsMAP1, are involved in a moderate low-temperature signalling pathway in rice. Plant Physiol 129:1880–1891
Joshi R, Sahoo KK, Tripathi AK et al (2018) Knockdown of an inflorescence meristem-specific cytokinin oxidase - OsCKX2 in rice reduces yield penalty under salinity stress condition. Plant Cell Environ 41:936–946
Kang NY, Cho C, Kim J (2013) Inducible expression of Arabidopsis response regulator 22 (ARR22), a type-C ARR, in transgenic Arabidopsis enhances drought and freezing tolerance. PLoS One 8:e79248
Keshishian EA, Hallmark HT, Ramaraj T et al (2018) Salt and oxidative stresses uniquely regulate tomato cytokinin levels and transcriptomic response. Plant Direct 2:e00071
Kiba T, Takei K, Kojima M et al (2013) Side-chain modification of cytokinins controls shoot growth in Arabidopsis. Dev Cell 27:452–461
Kieber JJ, Schaller GE (2014) Cytokinins. Arabidopsis Book 12:e0168
Kind S, Hinsch J, Vrabka J et al (2018) Manipulation of cytokinin level in the ergot fungus Claviceps purpurea emphasizes its contribution to virulence. Curr Genet 64:1303–1319
Klambt D (1992) The biogenesis of cytokinins in higher plants: our present knowledge. In: Kaminek M, Mok DWS, Zazimalova E (eds) Physiology and biochemistry of Cytokinins in plants. SPB Academic Publishing, The Hague
Kudoyarova GR, Vysotskaya LB, Cherkozyanova A et al (2007) Effect of partial root zone drying on the concentration of zeatin-type cytokinins in tomato (Solanum lycopersicum L.) xylem sap and leaves. J Exp Bot 58:161–168
Kuppu S, Mishra N, Hu R et al (2013) Water-deficit inducible expression of a cytokinin biosynthetic gene IPT improves drought tolerance in cotton. PLoS One 8:e64190
Lai F, Thacker J, Li Y et al (2007) Cell division activity determines the magnitude of phosphate starvation responses in Arabidopsis. Plant J 50:545–556
Le DT, Nishiyama R, Watanabe Y et al (2012) Identification and expression analysis of cytokinin metabolic genes in soybean under normal and drought conditions in relation to cytokinin levels. PLoS One 7:e42411
Letham DS (1994) Cytokinins as phytohormones: sites of biosynthesis, translocation, and function of translocated cytokinin. In: Mok D, Mok M (eds) Cytokinins: chemistry, activity, and function. CRC Press, Boca Raton, Florida, USA
Liao X, Guo X, Wang Q et al (2017) Overexpression of MsDREB6.2 results in cytokinin-deficient developmental phenotypes and enhances drought tolerance in transgenic apple plants. Plant J 89:510–526
Liu X, Huang B (2002) Cytokinin effects on creeping bentgrass response to heat stress: II. Leaf senescence and oxidant metabolism. Crop Sci 42:466–472
Lobell DB, Schlenker W, Costa-Roberts J (2011) Climate trends and global crop production since 1980. Science 333:616–620
Lomin S, Krivosheev D, Steklov MY et al (2012) Receptor properties and features of cytokinin signaling. Acta Nat 4:31
Mahonen AP, Bishopp A, Higuchi M et al (2006) Cytokinin signaling and its inhibitor AHP6 regulate cell fate during vascular development. Science 311:94–98
Markovich O, Steiner E, Kouˇril Š, Tarkowski P, Aharoni A, Elbaum R (2017) Silicon promotes cytokinin biosynthesis and delays senescence in Arabidopsis and Sorghum. Plant Cell Environ 40:1189–1196
Martin RC, Mok MC, Shaw G et al (1989) An enzyme mediating the conversion of zeatin to dihydrozeatin in Phaseolus embryos. Plant Physiol 90:1630–1635
Maruyama K, Urano K, Yoshiwara K et al (2014) Integrated analysis of the effects of cold and dehydration on rice metabolites, phytohormones, and gene transcripts. Plant Physiol 164:1759–1771
Maruyama-Nakashita A, Nakamura Y, Yamaya T et al (2004) A novel regulatory pathway of sulfate uptake in Arabidopsis roots: implication of CRE1/WOL/AHK4-mediated cytokinin-dependent regulation. Plant J 38:779–789
Mason MG, Jha D, Salt DE et al (2010) Type-B response regulators ARR1 and ARR12 regulate expression of AtHKT1;1 and accumulation of sodium in Arabidopsis shoots. Plant J 64:753–763
Matsumoto-Kitano M, Kusumoto T, Tarkowski P et al (2008) Cytokinins are central regulators of cambial activity. Proc Natl Acad Sci U S A 105:20027–20031
Miyawaki K, Matsumoto-Kitano M, Kakimoto T (2004) Expression of cytokinin biosynthetic isopentenyltransferase genes in Arabidopsis: tissue specificity and regulation by auxin, cytokinin, and nitrate. Plant J 37:128–138
Miyawaki K, Tarkowski P, Matsumoto-Kitano M et al (2006) Roles of Arabidopsis ATP/ADP isopentenyltransferases and tRNA isopentenyltransferases in cytokinin biosynthesis. Proc Natl Acad Sci U S A 103:16598–16603
Mizuno T, Nakamichi N (2005) Pseudo-response regulators (PRRs) or true oscillator components (TOCs). Plant Cell Physiol 46:677–685
Mohan TC, Castrillo G, Navarro C et al (2016) Cytokinin determines thiol-mediated arsenic tolerance and accumulation. Plant Physiol 171:1418–1426
Mok DWS, Mok MC, Shaw G et al (1990) Genetic differences in the enzymatic regulation of zeatin metabolism in Phaseolus Embryos. In: Pharis RP, Rood SB (eds) Plant Growth Substances 1988. Springer, Berlin, Heidelberg
Muller D, Waldie T, Miyawaki K et al (2015) Cytokinin is required for escape but not release from auxin mediated apical dominance. Plant J 82:874–886
Mytinova Z, Motyka V, Haisel D et al (2011) Antioxidant enzymatic protection during tobacco leaf ageing is affected by cytokinin depletion. Plant Growth Regul 65:23–34
Nadeem M, Li J, Yahya M et al (2019) Research progress and perspective on drought stress in legumes: a review. Int J Mol Sci 20:2541
Nam YJ, Tran LSP, Kojima M et al (2012) Regulatory roles of cytokinins and cytokinin signaling in response to potassium deficiency in Arabidopsis. PLoS One 7:e47797
Nandi S, Palni L, Letham D et al (1989) Identification of cytokinins in primary crown gall tumours of tomato. Plant Cell Environ 12:273–283
Nguyen KH, Ha CV, Nishiyama R et al (2016) Arabidopsis type B cytokinin response regulators ARR1, ARR10, and ARR12 negatively regulate plant responses to drought. Proc Natl Acad Sci U S A 113:3090–3095
Nishiyama R, Watanabe Y, Fujita Y et al (2011) Analysis of cytokinin mutants and regulation of cytokinin metabolic genes reveals important regulatory roles of cytokinins in drought, salt and abscisic acid responses, and abscisic acid biosynthesis. Plant Cell 23:2169–2183
Nooden LD, Singh S, Letham DS (1990) Correlation of xylem sap cytokinin levels with monocarpic senescence in soybean. Plant Physiol 93:33–39
O’Brien JA, Benkova E (2013) Cytokinin cross-talking during biotic and abiotic stress responses. Front Plant Sci 4:11
Panda BB, Sekhar S, Dash SK et al (2018) Biochemical and molecular characterisation of exogenous cytokinin application on grain filling in rice. BMC Plant Biol 18:89
Pavlu J, Novak J, Koukalova V et al (2018) Cytokinin at the crossroads of abiotic stress signalling pathways. Int J Mol Sci 19:2450
Pilon-Smits EA, Quinn CF, Tapken W et al (2009) Physiological functions of beneficial elements. Curr Opin Plant Biol 12:267–274
Pospisilova J, Vagner M, Malbeck J et al (2005) Interactions between abscisic acid and cytokinins during water stress and subsequent rehydration. Biol Plant 49:533–540
Radhika V, Ueda N, Tsuboi Y et al (2015) Methylated cytokinins from the phytopathogen Rhodococcus fascians mimic plant hormone activity. Plant Physiol 169:1118–1126
Rahayu YS, Walch-Liu P, Neumann G et al (2005) Root-derived cytokinins as long-distance signals for NO3-induced stimulation of leaf growth. J Exp Bot 56:1143–1152
Rahnama A, James RA, Poustini K et al (2010) Stomatal conductance as a screen for osmotic stress tolerance in durum wheat growing in saline soil. Funct Plant Biol 37:255–263
Ramireddy E, Chang L, Schmulling T (2014) Cytokinin as a mediator for regulating root system architecture in response to environmental cues. Plant Signal Behav 9:e27771
Rivero RM, Shulaev V, Blumwald E (2009) Cytokinin-dependent photorespiration and the protection of photosynthesis during water deficit. Plant Physiol 150:1530–1540
Romanov GA, Lomin SN, Schmulling T (2006) Biochemical characteristics and ligand-binding properties of Arabidopsis cytokinin receptor AHK3 compared to CRE1/AHK4 as revealed by a direct binding assay. J Exp Bot 57:4051–4058
Roychoudhury A, Chakraborty M (2013) Biochemical and molecular basis of varietal difference in plant salt tolerance. Annu Res Rev Biol 3:422–454
Roychoudhury A, Paul A (2012) Abscisic acid-inducible genes during salinity and drought stress. In: Berhardt LV (ed) Advances in medicine and biology, vol 51. Nova Science Publishers, New York, pp 1–78
Roychoudhury A, Paul S, Basu S (2013) Cross-talk between abscisic acid-dependent and abscisic acid-independent pathways during abiotic stress. Plant Cell Rep 32:985–1006
Sakai H, Honma T, Aoyama T et al (2001) ARR1, a transcription factor for genes immediately responsive to cytokinins. Science 294:1519–1521
Sakakibara H (2006) Cytokinins: activity, biosynthesis, and translocation. Annu Rev Plant Biol 57:431–449
Sakakibara H (2010) Cytokinin biosynthesis and metabolism. In: Davies P (ed) Plant hormones. Springer Netherlands
Schafer M, Meza-Canales ID, Navarro-Quezada A et al (2014) Cytokinin levels and signaling respond to wounding and the perception of herbivore elicitors in Nicotiana attenuata. J Integr Plant Biol 57:198–212
Schaller GE, Street IH, Kieber JJ (2014) Cytokinin and the cell cycle. Curr Opin Plant Biol 21:7–15
Schiavon M, Pilon-Smits EA (2017) The fascinating facets of plant selenium accumulation –biochemistry, physiology, evolution and ecology. New Phytol 213:1582–1596
Schoor S, Farrow S, Blaschke H et al (2011) Adenosine kinase contributes to cytokinin interconversion in Arabidopsis. Plant Physiol 157:659–672
Shi X, Rashotte AM (2012) Advances in upstream players of cytokinin phosphorelay: receptors and histidine phosphotransfer proteins. Plant Cell Rep 31:789–799
Shi Y, Tian S, Hou L et al (2012) Ethylene signaling negatively regulates freezing tolerance by repressing expression of CBF and type-A ARR genes in Arabidopsis. Plant Cell 24:2578–2595
Siddique S, Radakovic ZS, De La Torre CM et al (2015) A parasitic nematode releases cytokinin that controls cell division and orchestrates feeding site formation in host plants. Proc Natl Acad Sci U S A 112:12669–12674
Siemens J, Keller I, Sarx J et al (2006) Transcriptome analysis of Arabidopsis clubroots indicate a key role for cytokinins in disease development. Mol Plant Microbe Interact 19:480–494
Singh S, Letham DS, Jameson PE et al (1988) Cytokinin biochemistry in relation to leaf senescence IV. Cytokinin metabolism in soybean explants. Plant Physiol 88:788–794
Skalak J, Cerny M, Jedelsky P et al (2016) Stimulation of ipt overexpression as a tool to elucidate the role of cytokinins in high temperature responses of Arabidopsis thaliana. J Exp Bot 67:2861–2873
Sobol S, Chayut N, Nave N et al (2014) Genetic variation in yield under hot ambient temperatures spotlights a role for cytokinin in protection of developing floral primordia. Plant Cell Environ 37:643–657
Spallek T, Melnyk CW, Wakatake T et al (2017) Interspecies hormonal control of host root morphology by parasitic plants. Proc Natl Acad Sci U S A. 114:5283–5288
Steponkus PL, Uemura M, Webb MS (1993) A contrast of the cryostability of the plasma membrane of winter rye and spring oat-two species that widely differ in their freezing tolerance and plasma membrane lipid composition. In: Steponkus PL (ed) Advances in low-temperature biology, vol 2. JAI Press, London
Stolz A, Riefler M, Lomin SN et al (2011) The specificity of cytokinin signalling in Arabidopsis thaliana is mediated by differing ligand affinities and expression profiles of the receptors. Plant J 67:157–168
Strnad M (1997) The aromatic cytokinins. Physiol Plant 101:674–688
Susan J, Fatemeh R, Latifeh P (2013) Effect of abiotic stresses on histidine kinases gene expression in Zea mays L. cv. SC. 704. J Stress Physiol Biochem 9:124–135
Suzuki T, Imamura A, Ueguchi C et al (1998) Histidine-containing phosphotransfer (HPt) signal transducers implicated in His-to-Asp phosphorelay in Arabidopsis. Plant Cell Physiol 39:1258–1268
Sweere U, Eichenberg K, Lohrmann J (2001) Interaction of the response regulator ARR4 with phytochrome B in modulating red light signaling. Science 294:1108–1111
Tarkowska D, Dolezal K, Tarkowski P et al (2003) Identification of new aromatic cytokinins in Arabidopsis thaliana and Populus × canadensis leaves by LC-(+) ESI-MS and capillary liquid chromatography/frit–fast atom bombardment mass spectrometry. Physiol Plant 117:579–590
Thu NBA, Hoang XLT, Truc MT et al (2017) Cytokinin signaling in plant response to abiotic stresses. In: Pandey GK (ed) Mechanism of plant hormone Signaling under stress
To JPC, Haberer G, Ferreira FJ et al (2004) Type-a ARRs are partially redundant negative regulators of cytokinin signaling in Arabidopsis. Plant Cell 16:658–671
To JPC, Deruere J, Maxwell BB et al (2007) Cytokinin regulates type-A Arabidopsis response regulator activity and protein stability via two-component phosphorelay. Plant Cell 19:3901–3914
Todaka D, Zhao Y, Yoshida T et al (2017) Temporal and spatial changes in gene expression, metabolite accumulation and phytohormone content in rice seedlings grown under drought stress conditions. Plant J 90:61–78
Tokunaga H, Kojima M, Kuroha T et al (2012) Arabidopsis lonely guy (LOG) multiple mutants reveal a central role of the LOG-dependent pathway in cytokinin activation. Plant J 69:355–365
Tran LS, Urao T, Qin F et al (2007) Functional analysis of AHK1/ATHK1 and cytokinin receptor histidine kinases in response to abscisic acid, drought, and salt stress in Arabidopsis. Proc Natl Acad Sci U S A 104:20623–20628
Veach YK, Martin RC, Mok DW et al (2003) O-glucosylation of cis-zeatin in maize. Characterization of genes, enzymes, and endogenous cytokinins. Plant Physiol 131:1374–1380
Walker MA, Dumbroff EB (1981) Effects of salt stress on abscisic acid and cytokinin levels in tomato. Zeitschrift Fur Pflanzenphysiologie 101:461–470
Walters DR, McRoberts N (2006) Plants and biotrophs: a pivotal role for cytokinins? Trends Plant Sci 11:581–586
Wang Y, Shen W, Chan Z et al (2015) Endogenous cytokinin overproduction modulates ROS homeostasis and decreases salt stress resistance in Arabidopsis thaliana. Front Plant Sci 6:1004
Werner T, Motyka V, Laucou V et al (2003) Cytokinin-deficient transgenic Arabidopsis plants show multiple developmental alterations indicating opposite functions of cytokinins in the regulation of shoot and root meristem activity. Plant Cell 15:2532–2550
Werner T, Nehnevajova E, Kollmer I et al (2010) Root-specific reduction of cytokinin causes enhanced root growth, drought tolerance, and leaf mineral enrichment in Arabidopsis and tobacco. Plant Cell 22:3905–3920
Woo J, MacPherson CR, Liu J et al (2012) The response and recovery of the Arabidopsis thaliana transcriptome to phosphate starvation. BMC Plant Biol 12:62
Wu C, Cui K, Wang W et al (2016) Heat-induced phytohormone changes are associated with disrupted early reproductive development and reduced yield in rice. Sci Rep 6:34978
Xiao XO, Zeng YM, Cao BH et al (2017) PSAG12-IPT overexpression in eggplant delays leaf senescence and induces abiotic stress tolerance. J Horticul Sci Biotech 92:349–357
Xu Y, Huang B (2017) Transcriptional factors for stress signaling, oxidative protection, and protein modification in ipt-transgenic creeping bentgrass exposed to drought stress. Environ Exp Bot 144:49–60
Xu Y, Gianfagna T, Huang B (2010) Proteomic changes associated with expression of a gene (ipt) controlling cytokinin synthesis for improving heat tolerance in a perennial grass species. J Exp Bot 61:3273–3289
Xu Y, Burgess P, Zhang X et al (2016) Enhancing cytokinin synthesis by overexpressing ipt alleviated drought inhibition of root growth through activating ROS-scavenging systems in Agrostis stolonifera. J Exp Bot 67:1979–1992
Yang Z, Liu G, Liu J et al (2017) Synergistic action of auxin and cytokinin mediates aluminum-induced root growth inhibition in Arabidopsis. EMBO Rep 18:1213–1230
Zalabak D, Pospisilova H, Smehilova M et al (2013) Genetic engineering of cytokinin metabolism: prospective way to improve agricultural traits of crop plants. Biotechnol Adv 31:97–117
Zwack PJ, Compton MA, Adams CI et al (2016) Cytokinin response factor 4 (CRF4) is induced by cold and involved in freezing tolerance. Plant Cell Rep 35:573–584
Acknowledgements
Financial assistance from the Science and Engineering Research Board, Government of India, through grant EMR/2016/004799 and the Department of Higher Education, Science and Technology and Biotechnology, Government of West Bengal, through grant 264(Sanc.)/ST/P/S&T/1G-80/2017 to Dr. Aryadeep Roychoudhury is gratefully acknowledged.
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2021 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this chapter

Cite this chapter
Singh, A., Roychoudhury, A. (2021). Cytokinin-Mediated Signalling During Environmental Stress in Plants. In: Aftab, T., Hakeem, K.R. (eds) Plant Growth Regulators. Springer, Cham. https://doi.org/10.1007/978-3-030-61153-8_6
Download citation
DOI: https://doi.org/10.1007/978-3-030-61153-8_6
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-61152-1
Online ISBN: 978-3-030-61153-8
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)