
Abstract
Uniting arthropods and the deep sea as a research focus allows us to combine the largest habitat on Earth with the most abundant and successful animal group on our planet. To set the stage for this volume, we briefly introduce the deep-sea environment, the history of its exploration and gaps in our knowledge in terms of global distributions of biodiversity, before we discuss the current understanding of the Crustacea and Pycnogonida within the phylum Arthropoda. The ecological context in the deep sea of the Americas is addressed and briefly outlined, including the geological history, biogeographic boundaries, and the effect of a large oxygen minimum zone in the region. Deep-water (>200 m) areas of the Americas represent roughly 72% of the national exclusive economic zones (EEZ) in the west Atlantic Ocean and 86% in the east Pacific Ocean. As in other regions of the world, our knowledge of the diversity and ecology of deep-water arthropod communities of the Americas is still very limited. Comprehensive research programs aimed at exploring the vast offshore areas off the Americas must increase rapidly to study the deep-water fauna before it is substantially, and possibly, irreversibly impacted by human activities and global climate change.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Pancrustacea
- Multicrustacea
- Astrobiology
- Biogeography
- Oxygen minimum zone
- Atlantic ocean
- Pacific ocean
- Caribbean sea
- Pelagic
- Benthic
- Bathyal
- Abyssal
- Exclusive economic zone
1.1 Introduction
1.1.1 Our Watery World and the Beginnings of Life
Look out across the ocean on a calm day, from the shore or from the deck of a ship. The vista is daunting in its scale yet innocuous in its features. But beneath this tranquil skin lies a teeming horde of organisms, from the tiniest of viruses to the mightiest whales, all of which are continually influenced by the physical features of the seawater within which they move—and by which they are moved. (Herring 2002)
Pycnogonids and crustaceans play important biological roles in all the Earth’s oceans, and in many habitats they dominate in numbers and/or biomass. Considering the deep sea, however, large gaps in our knowledge remain, and before a range of chapters present diversity, distribution, and significance of crustaceans in the American regions, let us expand our focus: How much do we know about our ocean? Do we still “know less about the bottom of our ocean than of the surface of the Moon”? What do we consider the deep sea to be and how diverse is this environment? And of course, the complexity of the question: What is an arthropod, and what place do crustaceans and pycnogonids hold within it?
The deep sea, with its unique chemical properties, remains a hot contender for the origin of all life on Earth around 4 billion years ago (Dodd et al. 2017). Most recently, Jordan et al. (2019) added to evidence that the complex organic compounds necessary for basic cell formation can be formed around deep-sea hydrothermal vents. This prompted the headline news “Life on Earth probably originated in deep-sea vents and aliens could be growing the same way now, scientists suggest” (Cockburn 2019). Could complex life have similarly evolved in oceans of other solar system bodies?
The Earth may not be the only world in our solar system to contain deep oceans. Many of its celestial bodies show the presence of water either as traces of vapor in their atmospheres or as ice on the surface. The Moon itself has strong indications of ice at its South Polar regions, making it a site considered for a permanently occupied base. Despite a long distance from the Sun in the coldness of space, deep, salty liquid oceans are suspected to exist beneath the icy surface of three of the Galilean moons of Jupiter, Europa, Ganymede, and Callisto, and similarly on Enceladus, a moon of Saturn (Paganini et al. 2020; Schmidt 2020; Cable et al. 2020). Tidal flexing from the gravitational pull of the nearby planet generates sufficient internal heat, and presumably hydrothermal vents, on these moons to form liquid oceans tens of kilometers deep below an icy surface, itself several kilometers thick. Despite their relatively small size in comparison to the Earth, there are strong suggestions that there is significantly more water in liquid oceans of some of these worlds than exists on Earth. Naturally, the question that comes to mind is: Are these ocean worlds hospitable for life to exist?
Proposed astrobiology programs are set to explore the conditions in and under the ice with melt probe missions and deployment of ocean gliders, using the Arctic and Antarctic sea ice as analogs (Schmidt 2020). Thus, before extrapolation to any extraterrestrial conditions can be made, it is first critical to have a sound knowledge of our own watery worlds. As such, the contributions to our understanding of ice-ocean interactions on Earth and the life it can support, provided by the initiatives of the planetary analog research, are significant. Together, they represent progress toward a better understanding of our own oceans under the ice here on Earth while pushing the boundaries of how data from other ocean worlds can be interpreted.
1.2 The Deep Sea
1.2.1 How Little Do We Know of Earth’s Deep Ocean?
In 1818, British explorer John Ross reported that he had collected a basket star from a depth of 1600 m during his search of the Northwest Passage – the first true deep-sea faunal record. Despite 200 years of subsequent intensive deep-sea research, a common quote persists: “95 % of the ocean remains unexplored and we know more about the surface of the Moon than the ocean floor.” Does this statement still hold in the twenty-first century?
The exploration of both the deep sea and space is relatively new, but true deep-sea discoveries predate the Russian and US space programs by nearly 140 years with the first satellite Sputnik 1 launched into Earth’s orbit by the Soviet Union only in 1957. Also, the surface of the Moon is about one-tenth of the area covered by ocean on Earth, i.e., 38 compared to 362 million km2 (Gregersen 2010; Charette and Smith 2010), and the combined total of physical samples available from the Moon are about 380 kg (Zeigler et al. 2019), compared to the swathes of deep-sea geological samples and biological specimens collected over the last two centuries. Twelve men have walked on the Moon, and, as of July 2020, thirteen people have visited the deepest point on Earth, the Challenger Deep of the Mariana Trench (Lobner 2020). Notably, the dive of the Bathyscaphe Trieste took Jacques Piccard and US Navy Lieutenant Don Walsh to a depth of around 10,900 m in 1960, just one year before Soviet Air Force Pilot Yuri Gagarin was the first person in space in 1961.
Yet, the deep sea is more than the deepest point on Earth. Every year, hundreds of manned submersible dives are conducted into the deep sea, taking scientists to bathyal and abyssal depths to study underwater features and faunal communities. In fact, several manned submersibles are rated to dive more than 1000 m, including the 6500 m rated American DSV Alvin and Japanese DSV Shinkai 6500 and the 7000 m rated Chinese DSV Jiaolong (Kelley et al. 2016). At even greater depths, the ability to research at hadal depths has substantially increased in 2018 and 2019 with the Five Deeps Expedition, which completed 39 dives with the submersible DSV Limiting Factor to the deepest point in every ocean (Jamieson 2020). In addition to manned submersibles, many research expeditions every year deploy a variety of sampling equipment and sensors, such as CTDs, sediment traps, baited landers, epibenthic sleds, and remotely operating and autonomous vehicles. They survey, image, and collect specimens in order to study the deep sea. There are also long-term research stations, like the Monterey Bay Aquarium Research Institute’s “Station M” which has been sampling the abyssal zone since 1989 (Smith et al. 2017). Arguably, there is a greater continued presence of scientists in the deep sea than in space, and the International Space Station (ISS) has been continuously occupied only since November 2000, with a total of 240 individuals from 19 countries who have visited the ISS since then (Garcia 2020).
The metrics that are applied to compare what we know of the bottom of the ocean and the surface of the Moon, however, are those of remote sensing and mapping. Detailed maps of the surfaces of the Moon, Mars, and Venus so far exceed the resolution available for our ocean floor. A near-global map of the lunar surface was compiled in 2011 by the NASA’s Lunar Reconnaissance Orbiter (LRO) to a scale of 100 m, which included the second ever image of the far side of the Moon. In contrast, the entire ocean floor has now been mapped to a maximum resolution of around 5 km (Sandwell et al. 2014), which allows for the identification of large features such as underwater mountains, ridges, or trenches. However, less than 18% of the seafloor has actually been measured directly (Mayer et al. 2018). While indeed the surface of our nearest celestial neighbors is better mapped than the deep ocean, this might change in the near future.
With an ambitious target, the Nippon Foundation-GEBCO Seabed 2030 Project is a focused global initiative for mapping the entire ocean floor to a resolution of about 140 m (at 4000 m depth), similar to those provided by the LRO, by the year 2030 (Mayer et al. 2018). Of course, accurate bathymetry maps are crucial for understanding the geomorphology that impacts many seafloor processes and habitat heterogeneity and, in turn, their biological communities (Stewart and Jamieson 2018). However, in situ images and specimens are what many scientists are interested in, but considering the global scale and the cost and effort to sample the deep sea (Clark et al. 2016), the distribution of deep-sea biological specimens remains very scarce globally. This is exemplified using the Ocean Biogeographic Information System (OBIS) database, the world’s largest repository of marine biological data. The distribution of over 4.4 million sample records from depths ≥200 m (per degree latitude and longitude) across the surface of all oceans is shown (Fig. 1.1). It is immediately evident that some areas (e.g., parts of continental Americas, Europe, and Asia) are relatively densely sampled, but the majority, particularly of the oceanic areas, have not been sampled at all.

OBIS records for all Animalia, depth ≥200 m, 4,440,609 records, 1° grid colored by counts of records from 0 (white) to 1055 (dark blue). Projection: Equal Earth (sphere) Americas, ArcGIS Pro 2.4.2. (The Esri Third-Party Software Acknowledgments are available online at https://links.esri.com/open-source-acknowledgments.) (Extracted 28 November 2019)
The same pattern emerges when one examines the global distribution of samples with depth. Webb et al. (2010) revealed “biodiversity’s big wet secret,” using OBIS records to clearly show that the number of biological records sharply declined from a depth of >200 m and then again between 5000 and 6000 m (Fig. 1.2). More than 50% of all OBIS records are from the continental shelf, which constitutes <10% of the ocean surface. The global distribution of records through the water column further highlights the paucity of records from the deep pelagic ocean. Thus, we can put to rest the oversimplistic and nonsensical comparison between our presumed knowledge of the deep ocean and the Moon. Over the past century, much work, time, and innovation have allowed us to reach deeper toward a fuller understanding of the deep ocean. Yet, there is much work in the next century to be done.

Global distribution within the water column of recorded marine biodiversity. The horizontal axis splits the oceans into five zones on the basis of depth, with the width of each zone on this axis proportional to its global surface area. The vertical axis is ocean depth, on a linear scale. This means that area on the graph is proportional to volume of ocean. The number of records in each cell is standardized to the volume of water represented by that cell and then log10-transformed. The inset shows in greater detail the continental shelf and slope, where the majority of records are found. (Reproduced from Webb et al. (2010), https://doi.org/10.1371/journal.pone.0010223.g002)
1.2.2 The Deep-Sea Environment
“Ninety per cent of the two-thirds of the surface of the Earth covered by the sea lies beneath the shallow margins of the continents; and most lies under 2 km or more of water. We may, therefore, with some justification, speak of the deep-sea bottom as constituting the most typical environment, and its inhabitants as the typical lifeforms of the solid face of our planet. Yet, because of the remoteness of this habitat and the difficulties in observing and sampling these organisms, they are known to only few scientists; and as living rather than pickled specimens, to less than a handful. Yet the possibility of life existing at these great depths, and a curiosity about the nature of these life forms in what appears to be one of the most ‘difficult’ of environments has fascinated Man since the early days of oceanic exploration” Gage and Tyler (1991: xi).
This Preface of what has become a standard textbook on deep-sea biology still rings true. The sheer scale of the three-dimensional space, that is, the open ocean, remains as daunting today as it was three decades ago. The average depth of the ocean is around 3700 m and covers an estimated area of 362 million km2 (Charette and Smith 2010), holding nearly all the space that can be inhabited by life on Earth. The deep sea as defined herein is considered as the area deeper than the continental shelf (on average to 200 m depth), reaching to nearly 11,000 m at the Challenger Deep, Mariana Trench, Pacific Ocean. This includes the world’s largest mountain range (the Mid-Atlantic Ridge) and the deepest canyons. Within the deep sea, there are three zones: the bathyal extending down the continental slope from 200 to 3000 m; the abyssal, from 3000 to 6000 m, covering 54% of the Earth’s surface with a vast network of abyssal plains that are punctuated by ocean ridges and trenches (Smith et al. 2008); and the hadal, from 6000 to 11,000 m, accounting for the deepest 45% of the ocean and consisting of subduction trenches, fracture zone, troughs, and depressions (Jamieson et al. 2009).
The world’s five oceans are connected by powerful global currents. Paradoxically, however, instead of being a single, uniform system, the ocean is complex and heterogeneous. There are myriad communities with changing interactions and relationships, and many of the species that inhabit these communities are adapted to the extreme environmental conditions of increasing hydrostatic pressure with depth, no light, low temperature, and limited food inputs (Perrone et al. 2003; Downing et al. 2018). Some deep-sea communities are highly specialized, such as the hydrothermal vent assemblages. These hydrothermally active habitats have only been recently discovered, the first in 1977 around the Galapagos Rift (Lonsdale 1977). Other distinct deep-sea communities, including prominent crustaceans, inhabit only the extreme depths of the hadal subduction trenches (Beliaev 1989; Jamieson 2015). In contrast, some species inhabit all oceans, with cosmopolitan species among pelagic crustaceans not uncommon (see review by Halsband et al. 2020). Increasingly powerful genetic tools, however, have revealed that many supposedly widespread species are mosaics of several cryptic or pseudocryptic species, in both pelagic (e.g., Andrews et al. 2014; Cornils and Held 2014; Cornils et al. 2017) and benthic shallow-water and deep-sea crustaceans (e.g., Garlitska et al. 2012; Havermans et al. 2013). In most cases, though, some general boundaries around regional assemblages can be drawn based on submarine topography, stable environmental characteristics, and geotectonic history. For example, Longhurst (2007), refined by Reygondeau and Dunn (2019), proposed nested global biogeographic partitions for pelagic ecosystems. Spalding et al. (2007), with Marine Ecoregions of the World (MEOW), did the same for the world’s coastal and shelf areas (depths <200 m), and Watling et al. (2013) refined the UNESCO’s (2009) Global Open Oceans and Deep Seabed (GOODS) classification with a delineation of bathyal, abyssal, and hadal provinces. While these marine biogeographic boundaries are rarely strict, the classifications in all cases are proposed to be of use as management tools and for analyses (such as predictive modelling) or to guide establishment of marine protected areas and minimize the negative impact of commercial activities in marine environments. The delineations of these global classifications were substantially guided by knowledge of deep-sea fauna distributions, so a common limitation is that of the fauna itself, which continues to bring surprises.
1.2.3 The Deep Sea of the Americas
1.2.3.1 The Ecological Context
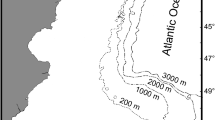
The Americas stretch in a continuous landmass from the Arctic Ocean in the north to the Antarctic Ocean in the south and covers nearly 30% of the Earth’s land area. The continent delimits the Atlantic Ocean from the Pacific Ocean and entirely encompasses the Caribbean Sea. To the north, narrow and shallow passages between Alaska and Russia (~80 km distance) and Canadian Ellesmere Island and Greenland (16 km distance) limit the present movement of water between deep oceans (Fig. 1.3). Instead, the only connection of deep oceanic waters remains the relatively small gap of the Drake Passage, between the southern tip of South America (56°S) and the northern tip of Palmer Peninsula in west Antarctica (62°S, Fig. 1.4). Here, waters join the Antarctic Circumpolar Current that fuels the thermohaline circulation or the global conveyor belt (Iudicone et al. 2008). “Engines” of this global circulation, where warm surface water sinks downward to join the cold bottom waters, are located near the Americas in the northern Atlantic Ocean (Labrador Sea) and on either side of South America (Ross Sea and Weddell Sea; Kuhlbrodt et al. 2007).

Geophysical and bathymetric map of North America and surrounding shelf and seas, showing the 200m bathymetric contour line (dark lines) and the national exclusive economic zones (white lines). The Gulf of Mexico is abbreviated (GOM). Projection: North America Lambert Conformal Conic. Credit: Esri, Garmin, GEBCO (General Bathymetric Chart of the Oceans), NOAA (National Oceanic and Atmospheric Administration) National Geophysical Data Center, and other contributors

Geophysical and bathymetric map of South America and surrounding shelf and seas showing the 200 m bathymetric contour line (dark lines) and the national exclusive economic zones (white lines). Projection: South America Lambert Conformal Conic. Credit: Esri, Garmin, GEBCO (General Bathymetric Chart of the Oceans), NOAA (National Oceanic and Atmospheric Administration) National Geophysical Data Center, and other contributors
In geological history, the continental movements changed underwater topography and exchange of water and associated fauna, with biological signals of these events still detectable in regional faunas (e.g., Feldmann and Schweitzer 2006). The breakup of Gondwana and Laurasia about 200 million years ago (mya) resulted in the formation of the separate Pacific, Indian, and Atlantic oceans which was completed during the Eocene Epoch (~50 mya). The formerly widespread Tethyan fauna began to regionally differentiate, facilitating lineage divergence. The eastern Pacific Ocean fauna remained connected to that of the Atlantic until relatively recently, being separated by the formation of the Isthmus of Panama during the Pliocene; strong evidence indicates a progressive shoaling and that a deep-water connection was shut off around 10 mya (Bacon et al. 2015; O’Dea et al. 2016). The linking of North and South America by the Isthmus of Panama had major impacts on global climate, oceanic and atmospheric currents, and biodiversity, yet the timing of this critical event remains contentious. In 1914, a new connection between the Pacific Ocean and the Caribbean Sea had been established by the Panama Canal with a number of channels and locks transiting through the freshwater Gatun Lake. Surprisingly little is known about the effect of the Panama Canal on the distribution of marine biota, but crustaceans appear to be some of the most successful hitchhikers that have been transported in both hull-fouling and ballast waters (Cohen 2006; Ros et al. 2014). The discussions typically consider shallow-water species, but it is probable that some vertically migrating planktonic stages of deep-water taxa are transported in ballast waters and survive the interoceanic transit, particularly in light of a significant expansion of the capacity of the Panama Canal (Brierley 2014; Muirhead et al. 2015).
The modern distributions of marine organisms around the Americas generally follow global latitudinal clines. The range boundaries of crustaceans and other marine invertebrates generally coincide with transitions between oceanic domains with distinct biogeochemical properties and planktonic and benthic communities (Watling et al. 2013; Reygondeau and Dunn 2019). In general, little is known about distribution pattern of deep-water pycnogonids of the Americas. Some species of Colossendeis feature a cosmopolitan or very wide distribution (Staples 2007; Hendrickx this volume). American species of Pycnogonida have been sparsely studied, and this volume contributes to their general knowledge (Soler et al. this volume).
In the western Atlantic Ocean, the biogeographic boundaries for marine species are often related to convergence regions of currents, where the cold Labrador Current meets the Gulf Stream in the north or the Falkland Current meets the Brazil Current in the south. These regions often show marked changes in regional species assemblages in both shallow and deep water (e.g., see Boschi (2000) for shallow-water crabs and Schnabel et al. (2011) for deep-water squat lobsters). While the Caribbean and the Gulf of Mexico communities appear to be distinct and diverse based on continental shelf species (Boschi 2000; Briggs 2003), these distinctions are less pronounced at pelagic bathyal and abyssal depths (Watling et al. 2013; Sutton et al. 2017). As a result, no single pattern is evident.
The separation between biogeographical provinces in the eastern Pacific Ocean is similarly related to major oceanographic processes. This includes areas of upwelling off Peru, Chile, and California and the presence of an extensive oxygen minimum zone that creates a large median wedge between a generally northern and southern fauna (see below). These biogeographic regions have been reported for shallow-water decapods (Wicksten 1989), bathyal squat lobsters (Macpherson et al. 2010), and mesopelagic plankton (Sutton et al. 2017). In all cases, however, studies point to extensive caveats, sampling gaps, and general patchy knowledge that often confound and preclude extrapolations to large regional and global scales.
1.2.3.2 The Oxygen Minimum Zone
Striking features affecting the composition and abundance of the deep-water pelagic and benthic communities worldwide are oxygen minimum zones (OMZ). While many small coastal areas are occasionally affected by short-term or smaller-scale oxygen depletions (Rabalais and Turner 2001; Gooday et al. 2009), at the oceanic level, OMZs are particularly ubiquitous and persistent in the East Pacific, the northern part of the Indian Ocean, and the East Atlantic (Diaz and Rosenberg 1995; Levin et al. 2000) (Fig. 1.5). Of these, the East Pacific OMZ is by far the widest of its kind (Fig. 1.5), ranging almost uninterrupted from Alaska to central Chile (Helly and Levin 2004), over about 90° of latitude and up to about 7000 km offshore (Diaz and Rosenberg 1995). The widest OMZ cores are found off western Mexico, Central America, and Peru. The core narrows considerably toward higher latitudes (the northern part of the Baja California Peninsula to Oregon and further north) as well as south of Peru (Helly and Levin 2004). In these areas, dissolved oxygen concentration rapidly decreases with depth and reaches severe hypoxic values or even close to anoxic conditions in what is known as the OMZ core (Helly and Levin 2004, Fig. 1.6). The depth at which the upper and lower limits of the OMZ core occur and the degree of intensity of the OMZ depend on geographic location. Thus, the bathymetric range (interval) corresponding to hypoxic (either mild or severe) or anoxic conditions will vary significantly from one locality to another (Helly and Levin 2004; Serrano 2012; Papiol et al. 2016).

Worldwide distribution of oxygen minimum zones. Data from the World Ocean Atlas

Oxygen depletion in the water column strongly affects pelagic communities not adapted to hypoxic or near-anoxic conditions. Likewise, in areas where the OMZ intercepts the continental shelf and slope, the composition and size of benthic communities will depend upon their tolerance to oxygen depletion. Additionally, the amount of organic matter available for consumption by detritus feeders is another critical driver (Levin et al. 2000, 2010; Papiol et al. 2016). Of paramount importance for local fisheries is the shallowest depth at which the OMZ core occurs, and shallow habitat compression has been predicted for hypoxia-intolerant taxa (Stramma et al. 2010). Indeed, the shallowest the depletion of dissolved oxygen starts, the highest the impact on fishery activities. In most of the East Pacific Ocean, fishing activities have traditionally taken place on the continental shelf (Méndez 1982; Hendrickx 1995; Rojas-Morales et al. 2000; Álvarez-León 2002; Tabash Blanco 2007; Hendrickx and Serrano 2010; Díaz Merlano et al. 2011; Mendo and Wosnitza-Mendo 2014; Reyes et al. 2014; Villalobos-Rojas et al. 2014; Vieira et al. 2016). Considering the entire East Pacific Ocean and with a few local exceptions, the upper boundary of the OMZ found between 40 and 60°N is located much deeper (between 650 and 550 m depth) than in other areas. Indeed, it is very shallow between 25°N and 5°N (200 m depth or less) and between 5°S and about 20°S (100 m depth or less; Fig. 1.6). Similarly, the depth at which the OMZ core lower boundary is found on average varies with latitude. In the higher latitudes (40–60°N), it is located at 1100–1200 m depth; in the intermediate latitudes (30–5°N), it occurs at 800–1000 m; and in the lower latitudes (0–25°S), it is much shallower (400–500 m; Helly and Levin 2004). Consequently, as for other groups of invertebrates, deep-water crustaceans tolerant of (some) oxygen depletion, or living permanently below the OMZ core, will be found at different depths depending on the latitudinal range they inhabit (Papiol et al. 2016).
Along the OMZ, the deep-water fauna occurring below the lower boundary of the core is almost entirely distinct from the fauna found on the shelf or above the upper boundary of the core (Kameya et al. 1997; Quiroga et al. 2009; Hendrickx and Serrano 2010). Migration of adult stages of benthic species through the OMZ, from shallow to deep water or inversely, appears to be strongly limited due to their physiological intolerance to oxygen depletion (Quiroga et al. 2009; Hendrickx and Serrano 2014). In a similar manner, dispersal and vertical migrations of pelagic forms (including the larval stages of benthic species) are also perturbed or limited (Judkins 1980; Saltzman and Wishner 1997; Hidalgo et al. 2005), particularly in localities where the upper boundary of the OMZ is very shallow (e.g., less than 25 m depth; Criales-Hernández et al. 2008). Consequently, this might interfere with latitudinal and longitudinal distribution and dispersion patterns of many planktonic and nektonic species in areas where the OMZ is wide and strong (Judkins 1980; Morales et al. 1999; Escribano et al. 2000).
1.2.3.3 Political Boundaries and Deep Territorial Seas
All but two of the 35 countries of the Americas (namely, Bolivia and Paraguay) adjoin at least one of the three major oceans. Combined, in the Atlantic Ocean, the total area covered by national exclusive economic zones (EEZ), the territorial sea extending from the coastline to 200 nautical miles offshore (outlined in white in Figs. 1.3 and 1.4), covers about 14.6 M km2, of which 10.5 M km2 (72%) are deeper than 200 m. Comparatively, in the Pacific Ocean, the EEZs cover about 13.9 M km2, of which 11.9 M km2 (86%) are deeper than 200 m (Figs. 1.3 and 1.4; FMI 2018, 2019, GEBCO 2020). Higher proportion of deep sea in the Pacific is linked to the presence of several faraway offshore islands, e.g., Clarion, Malpelo, Galápagos, and Easter Islands.
The extent of the deep sea (>200 m depth) in these American countries is extraordinarily variable, depending on two factors: (1) the size of their EEZ and (2) the steepness of the continental margin. For the Americas, the largest EEZ is that of the USA (not including Hawaii), but this includes the Alaska portion which makes up nearly half (about 2.5 M km2 of a total of 5.5 M km2). In contrast, the smallest EEZ is found around San Cristobal and Nevis of the Lesser Antilles with 9,974 km2; however, this has a proportionally very large deep-water extension, with 93.5% of its EEZ deeper than 200 m. The country with the highest proportion of water deeper than 200 m is Barbados, with 99.8%. By comparison, Argentina with the smallest portion of the EEZ deeper than 200 m (26.1%) is a country with a proportionally large continental shelf. The deepest point in the Americas is Milwaukee Deep, at 8376 m, of the Puerto Rico Trench off the Island of Puerto Rico (Stewart and Jamieson 2019). Given the significant proportion of deep-water habitat lying in the EEZ of countries of the Americas, national tasks of exploring and understanding the structure and functions of these large ecosystems remain colossal. Intensive studies of both the deep-water benthic and pelagic communities that are recognized as diverse and abundant should be considered, as well as monitoring in a permanent manner the environmental conditions prevailing in these ecosystems, particularly deploying deep-water observatories in key areas.
1.2.4 Marine Arthropoda
The phylum Arthropoda comprises insects, spiders, pycnogonids, scorpions, millipedes, and crustaceans. They all share segmented, chitinous exoskeletons and need to molt regularly to grow. Arthropods have a body form so versatile they have been compared to Swiss Army knives and are the most diverse metazoan form of life; with more than 1.3 million known species, they encompass nearly 80% of all known animals (Zhang 2013). However, the relationships among these groups remain hotly debated. Aria and Caron (2017) recently observed that retracing the evolutionary history of arthropods has been one of the greatest challenges in biology. The contributions herein primarily cover Crustacea, as the dominant marine arthropods, but Pycnogonida (sea spiders) are also included as they are one of only two other relatively diverse groups of marine arthropods (the other being the marine mites, e.g., Bartsch 2006). The exclusively marine Pycnogonida are currently placed within the subphylum Chelicerata, which includes the terrestrial spiders and ticks, but the exact phylogenetic placement of pycnogonids remains unclear. Recent genomic-scale analysis of the chelicerates indicated that the marine forms (pycnogonids and the horseshoe crabs) form successive sister groups to terrestrial chelicerates, suggesting a single colonization of land within Chelicerata (Lozano-Fernandez et al. 2019b).
Progressing from the historical view, the Crustacea are now accepted to not be monophyletic inasmuch as insects (Hexapoda) are derived from within the crustaceans (Regier et al. 2010; Schwentner et al. 2017; Lozano-Fernandez et al. 2019a). The crustacean sister group to insects is still debated, but the consistent consensus based on major molecular and morphological datasets points to the remipedes, a small group of highly derived cave dwellers (Lozano-Fernandez et al. 2019a; Ahyong 2020) that were thought by some to be the most “primitive” crustacean form (Schram 1983). The crustaceans and insects are now variously grouped together as Pancrustacea, Tetraconata, or simply Crustacea sensu lato, in which the hexapods are understood as terrestrial crustaceans.
In this volume, the chapters addressing Crustacea are primarily focused on the class Malacostraca. This highly diverse class includes Decapoda (e.g., crabs, prawns, and lobsters), Peracarida (e.g., amphipods, isopods, and cumaceans), and Stomatopoda (mantis shrimps). Other chapters present aspects of the Copepoda, which are united with the Malacostraca in the superclass Multicrustacea (Schwentner et al. 2018; Lozano-Fernandez et al. 2019a). It comprises over 57,100 species, about 85% of all known Crustacea (~67,000) worldwide (Ahyong et al. 2011). Multicrustacea are primarily marine and can be found throughout the world’s oceans and depths. They range in size from a fraction of a millimeter in length for the minute parasitic Tantulocarida Boxshall and Lincoln, 1983 to the Japanese spider crab, Macrocheira kaempferi (Temminck, 1836), with a leg span that can reach 3.8 m (Davie et al. 2015). Moreover, Multicrustacea can be described by a number of superlatives: the aforementioned Japanese spider crab is the largest living arthropod; the coconut crab, Birgus latro (Linnaeus, 1767) is the largest land invertebrate with a weight of up to 4 kg and a leg span of up to 1 m (Drew et al. 2010, Fig. 1.7 center); the deep-sea amphipod of the genus Hirondellea Chevreux, 1889 dominates the scavenging community at extreme depths in the world’s trenches including Challenger Deep (Dahl 1959; Kobayashi et al. 2012; Lacey et al. 2016); planktonic copepods are the dominant mesozooplankton in the marine environment, comprising as much as 80% of its total biomass (Kiørboe 1998). With the diversity in form and number of marine arthropods, it is clear that distributions (vertical and horizontal), lifestyles, and abundances are not uniform throughout the deep sea but differ according to both geological and evolutionary histories (Wilson and Ahyong 2015). Deepwater decapods are most diverse and abundant in the bathyal zone, substantially giving way to peracarids at abyssal depths and to almost entirely relinquish to peracarids at hadal depths. Just as the habitats of the deep sea are not uniform, neither are the faunas, but nearly universally, the ecological significance of Multicrustacea in the marine realm cannot be overstated. They play a fundamental role in exporting, redistributing, and repackaging carbon and nutrients (e.g., Longhurst 1995; Smith et al. 2008; Jamieson et al. 2009), and many species are economically important.

The diversity of shape and form illustrated in the Nouveau Larousse Illustré Ocean Educational Chart by Adolphe Millot (1857–1921), showing examples of various decapods such as crabs, shrimps, and lobsters (1–26), stomatopods (27–28), amphipods and isopods (29–32), notostracan (33), copepods (34–38), and barnacles (39–40)
As marine arthropods constitute a significant portion of the marine biodiversity in nearly all regions and across all depths, they present an excellent model for our understanding of large-scale patterns at global scales. Yet, large areas of the deep sea remain unknown, at the same time as the global climate is undeniably changing.
1.3 Conclusions
The deep ocean surrounding the North and South American continents hosts a wide diversity of arthropods, from microscopic peracarids and benthic copepods to some of the largest decapods such as king crabs. Contributions herein address a wide range of biological aspects related to deep sea crustaceans and pycnogonids, ranging from taxonomic inventories, phylogenetics, and biogeography to community analysis, ecology, reproduction, and fisheries (see Preface).
As in many other regions of the world, our knowledge of the composition and ecology of the deep-water arthropod communities of the Americas is still very limited. Most countries of the Americas, in particular in Central and South America, are yet to implement comprehensive research programs aimed at exploring the vast offshore areas extending off their coastlines. In many cases, limitations are directly linked to the high cost of managing offshore research. Nevertheless, it is noteworthy that despite very limited means, there has been a consistent effort in the last decades to explore the deep-water resources with a view to identify potentially lucrative deep-water fishing grounds or mineral deposits. While this effort is understandable, there is obviously a very high risk that these marine communities are adversely affected when we lack detailed knowledge of the composition and ecology. Returning to the earlier claim, we know more about the surface of the moon than the ocean floor – is it still that case? We certainly have more data, more images, and more samples from the deep sea than the surface of the Moon. However, the sheer scale and complexity of the deep sea and its living systems mean that even in the twenty-first century, the claim remains arguably true.
Unfortunately, it is becoming apparent that depth does not shield the deep ocean and its inhabitants from anthropogenic disturbance (Jamieson et al. 2017, 2019). The state of impact is symbolized by the recent description of Eurythenes plasticus Weston, 2020, a scavenging amphipod from hadal depth of the Mariana Trench named for the presence of microplastic in its gut (Weston et al. 2020). At a time where the global community is increasingly aware of the challenges and risks due to human disturbances and climate change, the immense effects of the deep ocean upon our climate remain a significant piece in the puzzle that is ill-understood (Bindoff et al. in press). Indications are that climate change might threaten the biodiversity of the deep sea proportionately more compared to shallow-water ecosystems (Brito-Morales et al. 2020). Let us remind ourselves that research and the regional and global initiatives to tackle these challenges, through technical advances and protection measures such as marine protected areas and sustainable management of fishing and mining, have to be considered with optimism for the future. A prerequisite for any of these initiatives is detailed knowledge of the fauna and communities in the deep sea. A challenge to all of these initiatives in the Americas is the significant knowledge gap that exists for many parts of the region. Contributions in the present volume are intended to fill some of these gaps.
References
Ahyong ST (2020) Evolution and radiation of Crustacea. In: Thiel M, Poore GCB (eds) The natural history of the Crustacea: evolution and biogeography, vol 8. Oxford University Press, New York, pp 53–79
Ahyong ST, Lowry JK, Alonso M, Bamber RN, Boxshall GA, Castro P, Gerken S, Karaman GS, Goy JW, Jones DS, Meland K, Rogers DC, Svavarsson J (2011) Subphylum Crustacea Brünnich, 1772. In: Zhang Z-Q (ed) Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. Zootaxa 3148:165–191
Andrews KR, Norton EL, Fernandez-Silva I, Portner E, Goetze E (2014) Multilocus evidence for globally distributed cryptic species and distinct populations across ocean gyres in a mesopelagic copepod. Mol Ecol 23:5462–5479. https://doi.org/10.1111/mec.12950
Aria C, Caron J-B (2017) Burgess Shale fossils illustrate the origin of the mandibulate body plan. Nature 545(7652):89–92. https://doi.org/10.1038/nature22080
Álvarez-León R (2002) Aprovechamiento de los crustáceos de importancia comercial del Pacífico colombiano. In: Hendrickx ME (ed) Contributions to the Study of East Pacific Crustaceans 1. [Contribuciones al Estudio de los Crustáceos del Pacífico Este 1]. Instituto de Ciencias del Mar y Limnología, UNAM, Mexico City, pp 1–35
Bacon CD, Silvestro D, Jaramillo C, Smith BT, Chakrabarty P, Antonelli A (2015) Biological evidence supports an early and complex emergence of the Isthmus of Panama. Proc Nat Acad Sci 112(19):6110–6115. https://doi.org/10.1073/pnas.1423853112
Bartsch I (2006) Halacaroidea (Acari): A guide to marine genera. Organ Divers Evol:1–104
Beliaev GM (1989) Deep sea ocean trenches and their fauna. Scripps Institution of Oceanography, San Diego
Bindoff NL, Cheung WWL, Kairo JG, Arístegui J, Guinder VA, Hallberg R, Hilmi N, Jiao N, Karim MS, Levin L, O’Donoghue S, Purca Cuicapusa SR, Rinkevich B, Suga T, Tagliabue A, Williamson P (in press) Changing ocean, marine ecosystems, and dependent communities. In: Pörtner H-O, Roberts DC, Masson-Delmotte V, Zhai P, Tignor M, Poloczanska E, Mintenbeck K, Alegría A, Nicolai M, Okem A, Petzold J, Rama B, Weyer NM (eds) IPCC special report on the ocean and cryosphere in a changing climate
Boschi EE (2000) Species of decapod crustaceans and their distribution in the American marine zoogeographic provinces. Rev Invest Desarr Pesq 13:1–136
Boxshall GA, Lincoln RJ (1983) Tantulocarida, a New Class of Crustacea Ectoparasitic on other Crustaceans. J Crust Biol (1):1–16. https://doi.org/10.2307/1547849
Brierley AS (2014) Diel vertical migration. Current Biol 24(22):R1074–R1076
Briggs JC (2003) Marine centres of origin as evolutionary engines. J Biogeogr 30:1–18
Brito-Morales I, Schoeman DS, Molinos JG, Burrows MT, Klein CJ, Arafeh-Dalmau N, Kaschner K, Garilao C, Kesner-Reyes K, Richardson AJ (2020) Climate velocity reveals increasing exposure of deep-ocean biodiversity to future warming. Nat Clim Chang 10:576–581. https://doi.org/10.1038/s41558-020-0773-5
Cable ML, Neveu M, Hsu H-W, Hoehler TM (2020) Enceladus. In: Meadows V, Des Marais DJ, Arney G, Schmidt B (eds) Planetary astrobiology. University of Arizona Press, Tuscon, pp 217–246
Charette MA, Smith WHF (2010) The volume of Earth’s ocean. Oceanography 23(2):112–114. https://doi.org/10.5670/oceanog.2010.51
Chevreux E (1889) Amphipodes nouveaux provenant des campagnes de “l’Hirondelle” 1887–1888. Bull Soc Zool France 14:283–289
Clark MR, Consalvey M, Rowden AA (2016) Biological Sampling in the Deep Sea. Wiley Blackwell, West Sussex
Cockburn H (2019) Life on Earth probably originated in deep-sea vents and aliens could be growing the same way now, scientists suggest. In: The Independent Digital News & Media Limited. https://www.independent.co.uk/news/science/origin-life-deep-sea-theory-hydrothermal-vents-aliens-planets-oceans-a9185991.html. Accessed 20 May 2020
Cohen AN (2006) Species introductions and the Panama Canal. In: Gollasch S, Galil BS, Cohen AN (eds) Bridging divides: Maritime Canals as invasion corridors. Springer, Dordrecht, p 127e206
Cornils A, Held C (2014) Evidence of cryptic and pseudocryptic speciation in the Paracalanus parvus species complex (Crustacea, Copepoda, Calanoida). Front Zool 11:1–17. https://doi.org/10.1186/1742-9994-11-19
Cornils A, Wend-Heckmann B, Held C (2017) Global phylogeography of Oithona similis s.l. (Crustacea, Copepoda, Oithonidae) – a cosmopolitan plankton species or a complex of cryptic lineages? Mol Phylog Evol 107:473–485. https://doi.org/10.1016/j.ympev.2016.12.019
Criales-Hernández MI, Schwamborn R, Graco M, Ayón P, Hirche HJ, Wolff M (2008) Zooplankton vertical distribution and migration off Central Peru in relation to the oxygen minimum layer. Helgol Mar Res 62(1):85–100
Dahl E (1959) Amphipoda from depths exceeding 6000 meters. Galathea Rep 1:211–241
Davie PJF, Guinot D, Ng PKL (2015) Anatomy and functional morphology of Brachyura. In: Castro P, Davie PJF, Guinot D, Schram FR, Von Vaupel Klein C (eds) Treatise on zoology – anatomy, taxonomy, biology – the Crustacea, complementary to the volumes translated from the French of the Traité de Zoologie, vol 9(C) (I), Decapoda: Brachyura (Part 1). pp 11–163
Díaz Merlano JM, Vieira Betancourt C, Melo Saldarriaga G (2011) Diagnóstico de las principales pesquerías del Pacífico colombiano. Fundación Mar Viva, Bogotá, Colombia
Diaz RJ, Rosenberg R (1995) Marine benthic hypoxia: A review of its ecological effects and the behavioural response of benthic macrofauna. Oceanogr Mar Biol An Rev 33:245–303
Dodd MS, Papineau D, Grenne T, Slack JF, Rittner M, Pirajno F, O’Neil J, Little CTS (2017) Evidence for early life in Earth’s oldest hydrothermal vent precipitates. Nature 543(7643):60–64. https://doi.org/10.1038/nature21377
Downing AB, Wallace GT, Yancey PH (2018) Organic osmolytes of amphipods from littoral to hadal zones: Increases with depth in trimethylamine N-oxide, scyllo-inositol and other potential pressure counteractants. Deep-Sea Res Part I Oceanogr Res Pap 138:1–10. https://doi.org/10.1016/j.dsr.2018.05.008
Drew MM, Harzsch S, Erland S, Hansson BS (2010) A review of the biology and ecology of the Robber Crab, Birgus latro (Linnaeus, 1767) (Anomura: Coenobitidae). Zool Anz 249:45–67. https://doi.org/10.1016/j.jcz.2010.03.001
Escribano R, Marín V, Irribarren C (2000) Distribution of Euphausia mucronata at the upwelling area of Peninsula Mejillones, northern Chile: the influence of the oxygen minimum layer. Sci Mar 64(1):69–77
Escribano R, Hidalgo P, Krautz C (2009) Zooplankton associated with the oxygen minimum zone system in the northern upwelling region of Chile during March 2000. Deep Sea Res Part II Top Stud Oceanogr 56(16):1083–1094
Feldmann RM, Schweitzer CE (2006) Paleobiogeography of Southern Hemisphere decapod Crustacea. J Paleontol 80(1):83–103. https://doi.org/10.1666/0022-3360(2006)080[0083:POSHDC]2.0.CO;2
FMI (2018) Flanders Marine Institute. IHO Sea Areas, version 3. Available online at https://www.marineregions.org/ doi:https://doi.org/10.14284/323
FMI (2019) Flanders Marine Institute. Maritime boundaries geodatabase: Maritime boundaries and exclusive economic zones (200NM), version 11. Available online at https://www.marineregions.org/. https://doi.org/10.14284/386
Gage JD, Tyler PA (1991) Deep-sea biology: a natural history of organisms at the deep-sea floor. Cambridge University Press, Cambridge
Garcia M (2020) International Space Station facts and figures. https://www.nasa.gov/feature/facts-and-figures. Accessed 20 May 2020
Garlitska L, Neretina T, Schepetov D, Mugue N, De Troch M, Baguley JG, Azovsky A (2012) Cryptic diversity of the ‘cosmopolitan’ harpacticoid copepod Nannopus palustris: genetic and morphological evidence. Mol Ecol 21(21):5336–5347. https://doi.org/10.1111/mec.12016
GEBCO (2020) GEBCO Compilation Group 2020 Grid. https://doi.org/10.5285/a29c5465-b138-234d-e053-6c86abc040b9
Gooday A, Jorissen F, Levin L, Middelburg J, Naqvi S, Rabalais N, Scranton M, Zhang J (2009) Historical records of coastal eutrophication-induced hypoxia. Biogeosciences 6:1707–1745
Gregersen E (2010) The inner solar system: the Sun, Mercury, Venus, Earth, and Mars. The Rosen Publishing Group, Inc, New York
Halsband C, Ahyong ST, Brandt A, Kosobokova K, Ward P, Goodall-Copestake WP, Macpherson E (2020) Chapter 6. Biogeography of the oceans. In: Thiel M, Poore GCB (eds) The natural history of the Crustacea: evolution and biogeography of the Crustacea. Oxford University Press, New York, pp 121–154
Havermans C, Sonet G, d’Udekem d’Acoz C, Nagy ZT, Martin P, Brix S, Riehl T, Agrawal S, Held C (2013) Genetic and morphological divergences in the cosmopolitan deep-sea amphipod Eurythenes gryllus reveal a diverse abyss and a bipolar species. PLoS One 8(9):e74218. https://doi.org/10.1371/journal.pone.0074218
Helly J, Levin L (2004) Global distribution of naturally occurring marine hypoxia on continental margins. Deep-Sea Res Part I Oceanogr Res Pap 51:1159–1168
Hendrickx M (1995) Introducción. In: Fischer W, Krupp F, Schneider W, Sommer C, Carpenter K, Niem V (eds) Guía FAO para la identificación de especies para los fines de la pesca. Pacífico centro-oriental Vol. I, Plantas e Invertebrados. FAO, Roma, pp 1–7
Hendrickx ME (this volume) The deepwater Colossendeis tenera Hilton, 1943 (Pycnogonida, Pantopoda, Colossendeidae) Off Western Mexico. In: Hendrickx ME (ed) Deep-sea pycnogonids and crustaceans of the Americas. Springer, Cham
Hendrickx M, Serrano D (2010) Impacto de la zona de mínimo de oxígeno sobre los corredores pesqueros en el Pacífico mexicano. Interciencia 35:12–18
Hendrickx M, Serrano D (2014) Effects of the oxygen minimum zone on squat lobsters distribution in the Gulf of California, Mexico. Central Eur J Biol 9:92–103. https://doi.org/10.2478/s11535-013-0165-6
Herring P (2002) The biology of the deep ocean. Biology of habitats. Oxford University Press, New York
Hidalgo P, Escribano R, Morales C (2005) Ontogenetic vertical distribution and diel migration of the copepod Eucalanus inermis in the oxygen minimum zone off northern Chile (20–21 S). J Plank Res 27(6):519–529
Iudicone D, Speich S, Madec G, Blanke B (2008) The Global Conveyor Belt from a Southern Ocean Perspective. J Phys Oceanogr 38(7):1401–1425. https://doi.org/10.1175/2007jpo3525.1
Jamieson A (2015) The Hadal Zone: life in the Deepest Oceans. Cambridge University Press, Cambridge
Jamieson AJ (2020) The Five Deeps Expeditions and an update of full ocean depth exploration and explorers. Mar Technol Soc J 54(1):6–12. https://doi.org/10.4031/MTSJ.54.1.1
Jamieson AJ, Fujii T, Mayor DM, Solan M, Priede IG (2009) Hadal trenches: the ecology of the deepest places on Earth. Trends Ecol Evol 25(3):190–197. https://doi.org/10.1016/j.tree.2009.09.009
Jamieson AJ, Malkocs T, Piertney SB, Fujii T, Zhang Z-Q (2017) Bioaccumulation of persistent organic pollutants in the deepest ocean fauna. Nat Ecol Evol 1(3):51. https://doi.org/10.1038/s41559-016-0051
Jamieson AJ, Brooks LSR, Reid WDK, Piertney SB, Narayanaswamy BE, Linley TD (2019) Microplastics and synthetic particles ingested by deep-sea amphipods in six of the deepest marine ecosystems on Earth. R Soc open sci 6(2):180667. https://doi.org/10.1098/rsos.180667
Jordan SF, Rammu H, Zheludev IN, Hartley AM, Maréchal A, Lane N (2019) Promotion of protocell self-assembly from mixed amphiphiles at the origin of life. Nat Ecol Evol 3(12):1705–1714. https://doi.org/10.1038/s41559-019-1015-y
Judkins D (1980) Vertical distribution of zooplankton in relation to the oxygen minimum off Peru. Deep-Sea Res Part I Oceanogr Res Pap 27:475–487
Kameya A, Castillo R, Escudero L, Tello E, Blaskovic V, Cordova J, Hooker Y, Gutiérrez M, Mayor S (1997) Localización, distribución y concentración de langostinos rojos de profundidad. Bol IMARPE, Spec Pub (1997):1–47
Kiorboe T (1998) Population regulation and role of mesozooplankton in shaping marine pelagic food webs. Hydrobiologia 363:13–27. https://doi.org/10.1023/a:1003173721751
Kobayashi H, Hatada Y, Tsubouchi T, Nagahama T, Takami H (2012) The hadal amphipod Hirondellea gigas possessing a unique cellulase for digesting wooden debris buried in the deepest seafloor. Plos One 7:e42727
Kuhlbrodt T, Griesel A, Montoya M, Levermann A, Hofmann M, Rahmstorf S (2007) On the driving processes of the Atlantic meridional overturning circulation. Rev Geophys 45(2). https://doi.org/10.1029/2004rg000166
Lacey NC, Rowden AA, Clark MR, Kilgallen NM, Mayor DJ, Linley TD, Jamieson AJ (2016) Community structure and diversity of scavenging amphipods from bathyal to hadal depths in three South Pacific trenches. Deep Sea Res Part I Oceanogr Res Pap 111:121–137. https://doi.org/10.1016/j.dsr.2016.02.014
Levin L, Gage J, Martin C, Lamont P (2000) Macrobenthic community structure within and beneath the oxygen minimum zone, NW Arabian Sea. Deep-sea Res Part II Top Stud Oceanogr 47:189–226
Levin LA, Sibuet M, Gooday AJ, Smith CR, Vanreusel A (2010) The roles of habitat heterogeneity in generating and maintaining biodiversity on continental margins: an introduction. Mar Ecol 31(1):1–5
Lobner P (2020) 60th Anniversary of the First Visit by Humans to the Deepest Point in the Ocean – the Challenger Deep. https://lynceans.org/all-posts/60th-anniversary-of-the-first-visit-by-humans-to-the-deepest-point-in-the-ocean-the-challenger-deep/. Accessed 20 May 2020
Longhurst A (1995) Seasonal cycles of pelagic production and consumption. Progress Oceanogr 36(2):77–167. https://doi.org/10.1016/0079-6611(95)00015-1
Longhurst A (2007) The ecological geography of the sea, 2nd edn. Elsevier Inc, Amsterdam/Heidelberg. https://doi.org/10.1016/B978-0-12-455521-1.X5000-1
Lonsdale P (1977) Clustering of suspension-feeding macrobenthos near abyssal hydrothermal vents at oceanic spreading centers. Deep Sea Res 24(9):857–863. https://doi.org/10.1016/0146-6291(77)90478-7
Lozano-Fernandez J, Giacomelli M, Fleming JF, Chen A, Vinther J, Thomsen PF, Glenner H, Palero F, Legg D, Iliffe T, Pisani D, Olesen J (2019a) Pancrustacean evolution illuminated by taxon-rich genomic-scale data sets with an expanded Remipede Sampling. Genome Biol Evol 11(8):2055–2070. https://doi.org/10.1093/gbe/evz097
Lozano-Fernandez J, Tanner AR, Giacomelli M, Carton R, Vinther J, Edgecombe GD, Pisani D (2019b) Increasing species sampling in chelicerate genomic-scale datasets provides support for monophyly of Acari and Arachnida. Nature Comm 10(1):2295. https://doi.org/10.1038/s41467-019-10244-7
Lu Y, Xia X, Cheung S, Jing H, Liu H (2019) Differential distribution and determinants of ammonia oxidizing Archaea sublineages in the Oxygen Minimum Zone off Costa Rica. Microorganisms 7(10):2–14. (453)
Mayer L, Jakobsson M, Allen G, Dorschel B, Falconer R, Ferrini V, Lamarche G, Snaith H, Weatherall P (2018) The Nippon Foundation—GEBCO Seabed 2030 Project: the quest to see the World’s Oceans Completely Mapped by 2030. Geosciences 8(2):63
Méndez M (1982) Crustáceos comerciales y otras especies comunes en el litoral peruano. Bol Lima 20:1–20
Mendo J, Wosnitza-Mendo C (2014) Reconstruction of total marine fisheries catches for Peru: 1950–2010. Univ British Columbia Working Pap Ser 2014-21, pp 1–23
Morales C, Hormazábal S, Blanco J (1999) Interannual variability in the mesoscale distribution of the depth of the upper boundary of the oxygen minimum layer off northern Chile (18–24S): Implications for the pelagic system and biogeochemical cycling. J Mar Res 57(6):909–932
Mosch T, Sommer S, Dengler M, Noffke A, Bohlen L, Pfannkuche O, Liebetrau V, Wallmann K (2012) Factors influencing the distribution of epibenthic megafauna across the Peruvian oxygen minimum zone. Deep Sea Res Part I Oceanogr Res Pap 68:123–135
Paganini L, Villanueva GL, Roth L, Mandell AM, Hurford TA, Retherford KD, Mumma MJ (2020) A measurement of water vapour amid a largely quiescent environment on Europa. Nat Astron 4:266–272. https://doi.org/10.1038/s41550-019-0933-6
Papiol V, Hendrickx ME, Serrano D (2016) Effects of latitudinal changes in the oxygen minimum zone of the northeast Pacific on the distribution of bathyal benthic decapod crustaceans. Deep-sea Res Part II Top Stud Oceanogr 137:113–130. https://doi.org/10.3897/BDJ.4.e8661
Perrone FM, Della Croce N, Dell’Anno A (2003) Biochemical composition and trophic strategies of the amphipod Eurythenes gryllus at hadal depths (Atacama Trench, South Pacific). Chem Ecol 9(6):441–449. https://doi.org/10.1080/0275754031000095723
Quiroga E, Sellanes J, Arntz W, Gerdes D, Gallardo V, Hebbeln D (2009) Benthic megafaunal and demersal fish assemblages on the Chilean continental margin: the influence of the oxygen minimum zone on bathymetric distribution. Deep-sea Res Part II Top Stud Oceanogr 56(16):1112–1123
Rabalais NN, Turner RE (2001) Hypoxia in the northern Gulf of Mexico: Description, causes and changes. In: Rabalais NN, Turner RE (eds) Coastal hypoxia: consequences for living resources and ecosystems, coastal and estuarine studies, vol 58. American Geophysical Union, Washington, DC, pp 1–36
Regier JC, Shultz JW, Zwick A, Hussey A, Ball B, Wetzer R, Martin JW, Cunningham CW (2010) Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences. Nature 463(7284):1079–1098. https://doi.org/10.1038/nature08742
Reyes C, Rodríguez C, León R (2014) Análisis bioeconómico de las pesquerías de camarón blanco del Pacífico colombiano. Rev Luna Azul 39:153–164
Reygondeau G, Dunn D (2019) Pelagic biogeography. In: Cochran JK, Bokuniewicz HJ, Yager PL (eds) Encyclopedia of ocean sciences, 3rd edn. Academic, Oxford, pp 588–598. https://doi.org/10.1016/B978-0-12-409548-9.11633-1
Rojas-Morales J, Campos-Montero J, Segura-Hernández A, Mug-Villanueva M, Campos R, Rodríguez-Solano O (2000) Shark fisheries in Central America: A review and update. Pesquería de tiburón en América Central: Una revisión y actualización. Uniciencia 17:49–56
Ros M, Ashton GV, Lacerda MB, Carlton JT, Vázquez-Luis M, Guerra-García JM, Ruiz GM (2014) The Panama Canal and the transoceanic dispersal of marine invertebrates: Evaluation of the introduced amphipod Paracaprella pusilla Mayer, 1890 in the Pacific Ocean. Mar Environ Res 99:204–211. https://doi.org/10.1016/j.marenvres.2014.07.001
Saltzman J, Wishner K (1997) Zooplankton ecology in the eastern tropical Pacific oxygen minimum zone above a seamount: 2. Vertical distribution of copepods. Deep-Sea Res Part I Oceanogr Res Pap 44:931–954
Sandwell DT, Müller RD, Smith WHF, Garcia E, Francis R (2014) New global marine gravity model from CryoSat-2 and Jason-1 reveals buried tectonic structure. Science 346(6205):65–67. https://doi.org/10.1126/science.1258213
Schmidt B (2020) The astrobiology of Europa and the Jovian System. In: Planetary astrobiology, pp 185–216
Schnabel KE, Cabezas Padilla P, McCallum A, Macpherson E, Ahyong S, Baba K (2011) Worldwide distribution patterns of squat lobsters. In: Poore GCB, Ahyong ST, Taylor J (eds) The biology of squat lobsters. CSIRO Publishing/CRC Press, Melbourne/Boca Raton, pp 149–182
Schram FR (1983) Remipedia and crustacean phylogeny. In: Schram FR (ed) Crustacean phylogeny, Crustacean Issues Vol 1. A.A. Balkema, Rotterdam, pp 23–28
Schwentner M, Combosch DJ, Nelson JP, Giribet G (2017) A phylogenomic solution to the origin of insects by resolving crustacean-hexapod relationships. Current Biol 27(12):1818–1824.e5. https://doi.org/10.1016/j.cub.2017.05.040
Schwentner M, Richter S, Rogers DC, Giribet G (2018) Tetraconatan phylogeny with special focus on Malacostraca and Branchiopoda: highlighting the strength of taxon-specific matrices in phylogenomics. Proc R Soc B: Biol Sci 285(1885):20181524. https://doi.org/10.1098/rspb.2018.1524
Serrano D (2012) La zona de mínimo oxígeno en el Pacífico mexicano. In: Zamorano P, Hendrickx M, Caso M (eds) Biodiversidad y comunidades del talud continental del Pacífico mexicano. Secretaría del Medio Ambiente y Recursos Naturales (SEMARNAT), Instituto Nacional de Ecología (INE), Mexico, pp 105–119
Smith CR, De Leo FC, Bernadino AG, Sweetman AK, Martinex Arbizu P (2008) Abyssal food limitation, ecosystem structure and climate change. Trends Ecol Evol 23(9):518–528. https://doi.org/10.1016/j.tree.2008.05.002
Smith KLJ, Sherman AD, McGill PR, Henthorn RG, Ferreira J, Huffard CL (2017) Evolution of monitoring an abyssal time- series station in the northeast Pacific over 28 years. Oceanogr 30(4):72–81. https://doi.org/10.5670/oceanog.2017.425
Soler-Membrives A, Lucena RA, Company JB, Rotllant G (this volume) Deep-sea Pycnogonids from Uruguay: every deep cruise adds valuable information. In: Hendrickx ME (ed) Deep-sea pycnogonids and crustaceans of the Americas. Springer, Cham
Spalding MD, Fox HE, Allen GR, Davidson N, Ferdaña ZA, Finlayson M, Halpern BS, Jorge MA, Lombana A, Lourie SA, Martin KD, McManus E, Molnar J, Recchia CA, Robertson J (2007) Marine ecoregions of the world: a bioregionalization of coastal and shelf areas. BioScience 57(7):573–583. https://doi.org/10.1641/b570707
Staples DA (2007) A new species of Colossendeis (Pycnogonida: Colossendeidae) together with records from Australian and New Zealand waters. Memoirs of Museum Victoria 64:79–94
Stewart HA, Jamieson AJ (2018) Habitat heterogeneity of hadal trenches: Considerations and implications for future studies. Prog Oceanogr 161:47–65. https://doi.org/10.1016/j.pocean.2018.01.007
Stewart HA, Jamieson AJ (2019) The five deeps: The location and depth of the deepest place in each of the world’s oceans. Earth-Sci Rev:102896. https://doi.org/10.1016/j.earscirev.2019.102896
Stramma L, Schmidtko S, Levin L, Johnson G (2010) Ocean oxygen minima expansions and their biological impacts. Deep-Sea Res Part I Oceanogr Res Pap 57(4):587–595
Sutton TT, Clark MR, Dunn DC, Halpin PN, Rogers AD, Guinotte J, Bograd SJ, Angel MV, Perez JAA, Wishner K, Haedrich RL, Lindsay DJ, Drazen JC, Vereshchaka A, Piatkowski U, Morato T, Błachowiak-Samołyk K, Robison BH, Gjerde KM, Pierrot-Bults A, Bernal P, Reygondeau G, Heino M (2017) A global biogeographic classification of the mesopelagic zone. Deep Sea Res Part I Oceanogr Res Pap 126:85–102. https://doi.org/10.1016/j.dsr.2017.05.006
Tabash Blanco F (2007) Explotación de la pesquería de arrastre de camarón durante el período 1991–1999 en el Golfo de Nicoya, Costa Rica. Revista de biologia tropical 55(1):207–218
Temminck CJ (1836) Coup-d’oeil sur la faune des iles de la Sonde et de l’empire du Japon: discours préliminaire, destiné à servir d’introduction à la Faune du Japon. https://doi.org/10.5962/bhl.title.119899
UNESCO (2009) Global Open Oceans and Deep Seabed (GOODS) – biogeographic classification. vol IOC Technical Series. UNESCO-IOC, Paris
von Linné C (1767) Systema naturae: per regna tria natura, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis 12.2 edn. Laurentii Salvii, Holmiae [Stockholm]. https://doi.org/10.5962/bhl.title.68927
Vieira C, Diaz MC, Díaz J (2016) Ordenamiento y manejo pesquero en la costa norte del Pacífico colombiano. In: La pesca artesanal en la costa norte del Pacífico colombiano: un horizonte ambivalente. Fundación MarViva, Bogotá, pp 45–57
Villalobos-Rojas F, Herrera-Correal J, Garita-Alvarado C, Clarke T, Beita-Jiménez A (2014) Actividades pesqueras dependientes de la ictiofauna en el Pacífico Norte de Costa Rica. Rev Biol Trop 62(4):119–137
Watling L, Guinotte JM, Clark MR, Smith C (2013) A proposed biogeography of the deep ocean floor. Prog Oceanogr 111:91–112
Webb TJ, Vanden Berghe E, O’Dor R (2010) Biodiversity’s big wet secret: the global distribution of marine biological records reveals chronic under-exploration of the deep pelagic ocean. Plos One 5(8):e10223. https://doi.org/10.1371/journal.pone.0010223
Weston JNJ, Carrillo-Barragan P, Linley TD, Reid WDK, Jamieson AJ (2020) New species of Eurythenes from hadal depths of the Mariana Trench, Pacific Ocean (Crustacea: Amphipoda). Zootaxa 4748(1):163–181. https://doi.org/10.11646/zootaxa.4748.1.9
Wilson GDF, Ahyong ST (2015) Lifestyles of the species-rich and fabulous: the deep-sea crustaceans. In: Watling L, Thiel M (eds) The natural history of the Crustacea: lifestyles and feeding biology. Oxford University Press, New York, pp 279–298
Zeigler RA, Mosie AB, Kent JJ, Krysher CH, Watts LA, McCubbin FM (2019). The Apollo sample collection: 50 years of solar system insight. Presented at the 50th Lunar and Planetary Science Conference, The Woodlands, Texas
Zhang Z-Q (2013) Animal biodiversity: an update of classification and diversity in 2013. Zootaxa 3703(1):005–011
Acknowledgments
Our thanks to Pieter Provoost (OBIS Data Manager and IOC Software Engineer) for a complete OBIS database export and Arne Pallentin and Sally Watson (NIWA) for GIS mapping support. We are grateful to Britt Lonneville, Vlaams Instituut voor de Zee, Oostende, Belgium, for providing data related to the EEZ extension in the Americas. We are grateful to Yanhong Lu (Department of Ocean Science, Hong Kong University of Sciences and Technology) for generously providing original data of Fig. 1.6b. We also thank Ruben Escribano (Departamento de Oceanografía, Universidad de Concepción) and Thomas Mosch (Helmholtz-Zentrum für Ozeanforschung Kiel, GEOMAR) for their assistance with the elaboration of Fig. 1.6c. Our gratitude to Danièle Guinot (Muséum national d’ Histoire naturelle, Paris) and Christine Carrau (Bibliothèque Théodore Carrau, Paris) for tracing the source of the Nouveau Larousse Illustration plate of crustaceans (Fig. 1.7). KS and RP were funded by a New Zealand Strategic Science Investment Fund to the National Institute of Water and Atmospheric Research (NIWA) Coasts and Oceans Research Programme 2 (2019/2020 SCI).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Schnabel, K.E., Ahyong, S.T., Gomez, A.J., Hendrickx, M.E., Peart, R.A., Weston, J.N.J. (2020). The Deep-Water Crustacean and Pycnogonid Fauna of the Americas in a Global Context. In: Hendrickx, M.E. (eds) Deep-Sea Pycnogonids and Crustaceans of the Americas. Springer, Cham. https://doi.org/10.1007/978-3-030-58410-8_1
Download citation
DOI: https://doi.org/10.1007/978-3-030-58410-8_1
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-58409-2
Online ISBN: 978-3-030-58410-8
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)