
Abstract
The Yazd–Kerman massifs include the poorly known highlands of central and southern Iran reaching to 4465 m a.s.l. at the highest peak. This area belongs to the Irano–Turanian phytogeographical region and 68% of the flora belongs to this floristic region. Total flora of the area is composed of 1308 vascular plant species of which 8% are endemics. The richest genera in terms of endemic species are Astragalus (21 species), Acantholimon (13), Cousinia (8), Echinops (8) and Nepeta (7). Hemicryptophytes (36%) and therophytes (32%) are the most dominant life forms. Several vegetation types are distinguishable in various habitats including halophytic vegetation, Stipa-Artemisia steppes, psammophytic vegetation, shrublands and woodlands, Rheum ribes vegetation, wetland communities, rocky cliff vegetation, subalpine umbelliferous communities and alpine–subnival communities. The protected areas hardly correlate with the biodiversity hotspots in the region and therefore many species are threatened.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


5.1 Introduction
The Yazd–Kerman massifs comprise several high and isolated mountains located in central and southern Iran (Fig. 5.1). The highest peaks of the YK massifs are Hezar (4465 m a.s.l.), Lalezar (4351 m a.s.l.), Palvar (4233 m a.s.l.), Jupar (4135 m a.s.l.), Shirkuh (4065 m a.s.l.), Khabr (3845 m a.s.l.) and Jebal Barez (3741 m a.s.l.). These high mountains are separated from other mountains by flat plains and lowland semi–desert and desert areas. The great Kavir Desert is situated on the north and Lut Desert on the east.

Geographical location of the YK massifs and climatic diagrams of five stations in the area. The highest mountain peaks are shown on the map
Even though being partly separated by lowland semi–desert areas, these mountains are strongly connected to each other floristically, and recent studies have recognized them as an integrated unit, either in the form of an area of endemism (Noroozi et al. 2018, 2019b) or as several centres of endemism (Noroozi et al. 2019a). The flora of the YK massifs shows a close phytogeographical connection to other neighboring mountains, especially the Zagros range (Noroozi et al. 2019b). Moreover, they have a remarkable floristic link to the mountains of the Hindu Kush, Central Asia and the Western Himalaya (Noroozi et al. 2008, 2010, 2011; Doostmohammadi and Kilian 2017). High mountains of the YK harbor many species-rich IT genera such as Astragalus, Acantolimon and Nepeta (Zohary 1973). However, the lowland areas are characterized by several psammophytic and halophytic IT species and in the southern territories of the area there is a strong representation of various SS and Somalia–Masai elements (Edmondson et al. 1980; Leonard 1988–1989).
Floristic studies on these mountains date back to the early twentieth century (Bornmüller 1911, 1937, 1938, 1940, 1942), followed by the extensive work of the Flora Iranica (Rechinger 1963–2015) and Flora of Iran (Assadi et al. 1988–2017). Early local floristic studies have been conducted on Jupar Mountain (Freitag and Kuhle 1980) and in Khabr National Park (Edmondson et al. 1980) and more recently on the Bahr–Aseman, Ra’skuh, Riseh, Khabr and Hezar Mountains and the Damgahan valley (Iran Nejad et al. 2001; Zarezadeh et al. 2007; Rajaei et al. 2011; Saberi et al. 2013; Malekpourzadeh et al. 2015; Payande et al. 2016). Since the Flora Iranica, new taxonomic articles have constantly added to the floristic stock of the area, either by reporting range extensions in some species or by introducing new taxa (Ghahreman and Mirtadzadini 2000; Attar et al. 2001; Maassoumi and Vakili 2001; Zarre and Podlech 2001; Mozaffarian 2002; Nordenstam et al. 2002; Jamzad et al. 2003; Podlech and Zarre 2003; Podlech and Maassouni 2003; Mirtadzadini and Attar 2004; Mirtadzadini et al. 2004; Podlech 2004; Ranjbar and Karamian 2004; Ghahremaninejad and Narimisa 2005; Khodashenas et al. 2006; Maasoumi and Mirtadzadini 2006; Mohsenzadeh et al. 2007; Ajani and Ajani 2008; Hamdi et al. 2008; Assadi 2009; Attar and Mirtadzadini 2009; Ajani et al. 2010; Mohsenzadeh et al. 2010; Noroozi et al. 2010; Mahmoodi et al. 2013; Sonboli et al. 2011; Ranjbar 2011; Kilian et al. 2012; Dashti et al. 2014; Mirtadzadini and Attar 2014; Moazzeni et al. 2014; Ranjbar and Mahmoudi 2015; Nejad Falatoury et al. 2016; Moazzeni et al. 2016; Rajaei and Mozaffarian 2016; Doostmohammadi and Kilian 2017; Pahlevani 2017; Doostmohammadi and Mirtadzadini 2019). The numerous reports and articles, especially in the most recent years, imply that these mountains are not yet well–known and new species are still waiting to be discovered (see above mentioned references). The most common vegetation types of the area were recorded by Zohary (1963), as part of two longer transects crossing the YK massifs. The introduced associations, however, were briefly described by mentioning only two dominant species without any coefficient of abundance–dominance (Leonard 1993). More recently, Leonard (1991–1992) investigated the plant associations of the desert areas and lowland plains of the YK region in more detail. Both studies are restricted mainly to lowlands and flat plains between mountain ranges and no vegetation studies have been conducted on highlands of the area.
Here we give a preliminary overview on the floristic diversity, phytogeographical affinity, vegetation structure and conservation status of the YK massifs. More specifically the statistics are provided for the number of species occurring in the area, their chorology and rate of endemism. Major plant communities and vegetation units in the area are also discussed briefly. Finally, Bupleurum aitchisonii (Boiss.) Wolff is reported from the area as an addition to the flora of Iran.
5.2 Geology
The YK massifs are composed of three major tectonic crustal blocks, including the Sanandaj–Sirjan zone, the Central Iranian microcontinent and the Urmia–Dokhtar volcanic belt that are separated mainly by the Nain–Baft fault (Berberian and King 1981; Mehdipour Ghazi and Moazzen 2015). The geological structure and orogeny of the YK massifs is very complicated. These high mountains are mostly composed of Cretaceous limestone and early Tertiary (Eocene) conglomerates (Kerman conglomerate, which consists of limestone, sandstone and basic volcanic components; Freitag and Kuhle 1980; Kuhle 1974). The precursory phases of the Alpine orogeny in Iran lasted from the late Triassic (200 Ma) to the Late Jurassic–Early Cretaceous (ca. 65 Ma; Berberain and King 1981). The subsequent late Cretaceous to middle Tertiary movements constructed the major structural and geographical features of the area and created a continental climate that has lasted up to the present (Stöcklin 1968). After the Eocene period of extensive volcanism, it was mainly during the Oligocene orogenic phase that the mountain ranges of Central Iran took shape. Most of the copper and lead–zinc mineralizations are related to the early Tertiary (Eocene) volcanic activities (Berberian and King 1981; Stöcklin 1968).
Soils of the area can be classified, based on topography, into three main groups including soils of the plains and valleys, soils of the plateaus and soils of the mountains. Valleys and low plains are covered mostly by fine or coarse–textured alluvial soils, halomorphic (solonchak and solonetz) soils and poorly drained saline alluvial soils. Plateaus above 1500 m a.s.l., which constitute a great part of the YK massifs, are mostly composed of desert soils in association with regosols, sand dunes, sierozem and solonchak soils. Soils of mountains and slopes are very shallow and without any clear profile. These soils comprise a high proportion of rock fragments with low rates of weathering. Soils of the dissected slopes and mountains of the YK massifs are mainly formed by complex associations of calcareous lithosols together with desert and sierozem soils, plus lithosols with sierozem and brown soils. Organic matter in these soils is very low (Dewan and Famouri 1964). In some places, like on the Bidkhan volcano (northwest of Lalezar Mountain), volcanic debris also plays a role in the formation of soils in the YK massifs (Khalili Mobarhan and Ahmadipour 2015). The reader is referred to Stöcklin (1968), Dewan and Famouri (1964) and Kuhle (1976) for a detailed survey on the orogeny and geology of the YK massifs.
5.3 Climate
The climate of the YK massifs is characterized by a high degree of continentality, prolonged dry summers and harsh winters. Based on the Bioclimatic Classification System of Iran (Djamali et al. 2011), major parts of the YK massifs are comprised within the Mediterranean desertic continental bioclimate. This bioclimate is characterized by a very low annual precipitation and extended dry season and covers a large part of the Central Iranian deserts. But this bioclimatic system doesn’t give a clear view of the climatic conditions and precipitation of all high mountains, as it suffers from a lack of data from higher elevations. The annual precipitation in the city of Baft (2280 m a.s.l.) is 250 mm (Fig. 5.1) and annual precipitation values of 300–450 mm can be assumed for higher elevations, as the precipitation increases by about 100 mm per 1000 m of elevation. Precipitation falls are mostly in the form of snow at higher elevations and during the winter. According to Djamali et al. (2011), some places of high elevation in the YK massifs belong to the Mediterranean xeric continental bioclimate, which also covers the eastern parts of the Zagros and most of the Kopet Dagh Mountains. The presence of a Mediterranean xeric oceanic bioclimate in the southern parts of the YK massifs is of high interest. This bioclimate covers the mountains and foothills of Jebal Barez and the Khabr National Park. The relatively low continentality and a somewhat higher average of winter temperature minima create an appropriate climate in several microhabitats.
There are no glaciers in the YK massifs, but in some colder years, several snow patches persist throughout the year in the valleys and on the north-facing slopes of the Shirkuh, Hezar and Lalezar mountains. Based on geomorphological studies in the Jupar, Hezar and Lalezar mountains, Kuhle (2008) concluded that two late-Pleistocene mountain glaciations (pre–LGP and LGP) have occurred in the area and that a rather low average annual temperature of 11.2 °C and 10.5 °C can be assumed for these highlands in the late-Pleistocene, about 130 and 60–18 thousand years ago.
5.4 Flora and Phytogeography
5.4.1 Paleobotany
The history of vegetation and climate change in the northwest and central Zagros shows the dominance of a cold and dry Stipa-Artemisia steppe before 14,000 cal. B.P. which gradually turned into an open scrub of Amygdalus–Pistacia plus Quercus brantii woodland. Thickening of these woodlands, starting at ca. 6000 cal. B.P., finally established the current–day oak forests of the Zagros (Van Zeist and Wright 1963; Van Zeist 1967; Van Zeist and Bottema 1977). Climatic oscillations and consequent vegetation dynamics in the YK massifs are probably similar to that in the Zagros range, but it is not clear how long after the Holocene amelioration the Amygdalus-Pistacia shrublands have expanded to the southeastern areas (Miller and Kimiaie 2006). Another difference is that the YK massifs are much drier and Quercus brantii stands haven’t occupied these mountains, so that the present climax situation is Amygdalus-Pistacia shrublands.
An extensive degradation of Amygdalus-Pistacia shrublands around 2700 cal. B.P. has been recorded from the southern Zagros around lake Maharlu (Djamali et al. 2009) and the southern margin of the YK mountains around Jiroft (Gurjazkaite et al. 2018). These vegetation degradations are presumably due to the intensive human activities which coincided with the beginning of the Persian Empire and continued through the Islamic epoch (Djamali et al. 2009; Gurjazkaite et al. 2018). Similar events may also be assumed for the YK massifs, as these mountains are not very far from the two points that have been studied.
5.4.2 Floristic Diversity
According to our database, 1308 vascular plant species occur within the boundaries of the YK massifs, belonging to 496 genera and 93 families. The Dicots are responsible for most of the diversity of the total flora with 1126 species (86%), in 403 genera and 70 families; the Monocots contribute 167 species (13%), in 84 genera and 18 families. Gymnosperms and Pteridophytes occur in the area with 7 species each. The most diverse families in the area are Asteraceae (204 species), Fabaceae (152), Brassicaceae (94), Poaceae (84), Lamiaceae (81), Caryophyllaceae (61), Apiaceae (54), Boraginaceae (46), Chenopodiaceae (45), and Scrophulariaceae (32; Fig. 5.2). The ten largest families comprise more than 66% of the total flora. The largest genera in the YK massifs are Astragalus (Fabaceae; 92 species), Cousinia (Asteraceae; 29), Nepeta (Lamiaceae; 24), Acantholimon (Plumbaginaceae; 22) and Euphorbia (Euphorbiaceae; 19; Fig. 5.3).

Number of species in the 30 most species rich families of the YK massifs vascular flora (the number of endemic and non-endemic species are separated by different shades)

Number of species in the 30 most species rich genera of the YK massifs vascular flora (the number of endemic and non-endemic species are separated by different shades)
Hemicryptophytes are the main life form in the YK massifs with 36% followed by therophytes (32%), chamaephytes (18%), phanerophytes (9%) and geophytes (5%). The high number of hemicryptophytes in the YK massifs is also seen in a similarly high presence in the general life form spectrum of the IT region. It reflects the predominance of a continental climate with harsh winter conditions. The semi–desert lowlands and salty or sandy plains between the mountain blocks of the YK massifs, together with longer dry seasons in these mountains, provide suitable habitats for many IT and SS therophyte species in this area. In fact the proportion of therophytes in the YK massifs is higher than in the other high mountains of the Iranian Plateau (Noroozi et al. 2019b). This should be due to the dry climate of the region. Although large genera of geophytes are important constituents in many IT habitats, the flora of the YK massifs is poor in this life form. Allium, Tulipa and Eremurus are represented in the area by 9, 2 and 2 species, respectively. From the Fritillaria species, only F. persica occurs in the area. Even F. imperialis, with a wide distribution that extends from Anatolia and the Zagros to the Hindu Kush and Kashmir, is absent from the YK massifs.
5.4.3 Phytogeography
The YK massifs are part of the IT phytogeographical region (Zohary 1973; Leonard 1988–1989). Along its southern borders, it gradually passes through a kind of ecotone to the northern borders of the SS regional zone. Doostmohammadi et al. (2018) redefined the territories of the IT region and split it up into an Irano–Turkestanian highland and a Turanian lowland region. According to this concept, the YK massifs belong to the Irano–Turkestanian phytogeographical region and are surrounded by Turanian areas, particularly in the north and east. Many characteristic Irano–Turkestanian genera, such as Astragalus, Cousinia, Acantholimon and Nepeta contribute to the high species richness and endemism in the YK massifs. Besides, several Turanian and Saharo–Sindian species have intruded the lowland and flat plains between the mountain blocks.
The floristic relationships between the YK massifs and the neighboring mountains are shown in (Fig. 5.4). Since the YK massifs are spatially close to the Zagros mountain range, their strong floristic connection is expected. Besides, a remarkable floristic link is observed between the YK massifs and the Hindu Kush, the mountains of Central Asia and even the Western Himalaya. This disjunct pattern of distribution has been postulated to be probably the result of post glacial warming that has pushed the species to higher elevations and separated them in widely disjuncted populations (Noroozi et al. 2008, 2011).

Floristic affinity of the YK massifs with other mountain ranges. TheY-axis is the percentage of species common with the YK massifs
The flora of the YK massifs is composed of a mixture of elements from different phytogeographical regions, which are classified here into seven main categories (Fig. 5.5), as follows:

Phytochoria of the species of the YK massifs. Cosm: Cosmopolite; P.Reg: Pluri-regional
(Sub)Cosmopolites
This group also can be categorized as Pluri–regional and includes Cosmopolitan and Circum–boreal high mountain species, such as Cerastium cerastoides, Gentiana aquatica, Sagina saginoides, together with ruderal species and weeds like Anagallis arvensis, Atriplex canescens, Capsella bursa–pastoris, Chenopodium album, Convolvulus arvensis, Euphorbia helioscopia, Gnaphalium luteoalbum and Lactuca serriola. This group comprises about 5% of the flora of the area.
IT-ES-M
Just over 3% of the total flora is distributed in three phytogeographical regions. Some representatives are Achillea millefolium, Arenaria leptoclados, Chorispora tenella, Crupina vulgaris, Euphorbia falcata, Geranium dissectum, Heliotropium europaeum, Lepidium draba, Parietaria judaica, Sisymbrium loesselii and Trifolium lappaceum.
IT-M
This group of bi–regional elements includes 10% of the species. Some members are Althaea ludwigii, Atractylis cancellata, Ammi majus, Astragalus hamosus, Calendula alata, Carduus arabicus, Centaurea depressa, Chenopodium botrys, Chrysanthemum coronarium, Crupina vulgaris, Eremopyrum orientale, Erodium laciniatum, Erodium malacoides, Euphorbia falcata, Gladiolus atroviolaceus, Gynandriris sisyrinchium, Herniaria cinerea, Matricaria aurea, Melilotus indicus, Thalictrum isopyroides and Vicia monantha.
IT-SS
This category includes 4% of total flora. Examples are Astragalus crenatus, Atriplex dimorphostegia, Brassica tournefortii, Convolvulus oxysepalus, Diplotaxis harra, Erodium deserti, Hyparrhenia hirta, Lappula spinocarpos, Pennisetum divisum, Platychaete aucheri, Poa sinaica, Schismus arabicus, Spergula fallax, Stipagrostis plumosa and Tribulus longipetalus.
IT-ES
With 3% of species, it is the smallest group of bi-regional elements. Some representatives are Carex melanostachya, Lappula heteracantha, Leonurus cardiaca, Luzula taurica, Myricaria germanica, Plantago atrata, Ranunculus arvensis and Scrophularia umbrosa.
SS Elements
About 7% of the flora of the YK is composed of the Saharo–Sindian element. Examples are Alyssum homalocarpum, Andrachne aspera, Argyrolobium roseum, Asteriscus pygmaeus, Astragalus schimperi, A. sparsus, Bassia eriophora, Brassica aucheri, Calligonum comosum, Cutandia memphitica, Dipcadi unicolor, Erucaria hispanica, Euphorbia granulata, Fagonia bruguieri, Farsetia longisiliqua, Forsskaolea tenacissima, Gaillonia calycoptera, Gaillonia crucianelloides, Gymnarrhena micrantha, Helianthemum lippii, Neurada procumbens and Savignya parviflora.
IT Elements
The Irano–Turanian species (68%) are here dealt within two categories according to Doostmohammadi et al. (2018), including Turanian and Irano–Turkestanian elements.
Turanian or Aralo–Caspian Elements
About 2% of the flora is composed of Turanian species. These are mostly psamophytic species occurring in sandy flat plains and include Bassia eriantha, Calligonum amoenum, C. denticulatum, Cithareloma lehmannii, Climacoptera turcomanica, Euphorbia densa, Octoceras lehmannianum, Schumannia karelinii, Spirorhynchus sabulosus and Stipagrostis karelinii.
Irano–Turkestanian
The Irano–Turkestanian elements are classified in the following subgroups:
Omni–Irano–Turkestanian
This group is composed of 154 species with their distribution throughout the Irano–Turkestanian region, from Turkey to Central Asia. Some representatives are Aethionema trinervium, Amberboa nana, Asperula glomerata, Astragalus campylorrhynchus, Atraphaxis spinosa, Berberis integerrima, Biebersteinia multifida, Celtis caucasica, Clypeola aspera, Cymbolaena griffithii, Dactylorhiza umbrosa, Dianthus crinitus, Drabopsis verna, Eremopyrum distans, Eryngium billardieri, Euphorbia szovitsii, Gagea gageoides, Polygonum polycnemoides and Ziziphora clinopodioides.
Irano–Anatolian Region
There are several Irano-Anatolian species with their main distribution from Turkey and Transcaucasus to the Alborz and the Zagros mountains, reaching also the YK massifs from their southeastern ranges. Some representatives are Allium eriophyllum, Anthemis haussknechtii, Arum elongatum, Astragalus caraganae, A. finitimus, Carex oreophila, Colchicum persicum, Cotoneaster kotschyi, Crataegus atrosanguinea, C. meyeri, Eremostachys macrophylla, Fritillaria persica, Heracleum persicum, Medicago caucasica, Nepeta meyeri, Pimpinella eriocarpa, Salvia hydrangea and Tanacetum pinnatum.
Zagros/YK Pattern of Distribution
There are 116 species distributed across the Zagros and the YK massifs. For example Acantholimon nigricans, Acanthophyllum leucostegium, Aethionema umbellatum, Allium cathodicarpum, Amygdalus elaeagnifolia, Anthemis gayana, Arenaria bulica, Arenaria minutissima, Arenaria persica, Astragalus angustistipulatus, A. campylanthus, A. daenensis, A. pseudoshebarensis, A. yazdii, Centaurea ispahanica, Colchicum bakhtiaricum, Colutea persica, Consolida trigonelloides, Cousinia araneosa, C. eriobasis, Dianthus macranthoides, Dionysia revoluta, Dorema aucheri, Echinops ceratophorus, Eremostachys pulvinaris, Euphorbia khabrica, Ferula assa–foetida, Gypsophila farsensis, Jurinea bungei, Linaria remotiflora, Matthiola flavida, Nepeta depauperata, N. natanzensis, Paronychia bungei, Pimpinella dichotoma, Polygonum dumosum, P. khajeh–jamali, Silene daenensis, S. nurensis, Stachys acerosa, Verbascum hasarense and Zerdana anchonioides.
YK/Hindu Kush–Central Asia Disjunct Distribution
There is a remarkable and relatively well known pattern of disjunct distribution from the YK massifs to the Hindu Kush, Central Asia and the Himalaya. This disjunct relationship is more prominent among high alpine and subnival species and has previously been documented for several species by several authors (Hedge and Wendelbo 1978, Noroozi et al. 2008, 2010; Ajani et al. 2010; Sonboli et al. 2011; Rajaei et al. 2011; Doostmohammadi and Kilian 2017). Some representatives are Brachyactis roylei, Delphinium uncinatum, Gagea alexii, Lactuca pumila, Levisticum officinale, Parnassia cabulica, Pedicularis cabulica, Salvia rhytidea, Tanacetum fisherae, Tanacetum pamiricum and Taraxacum chitralense. Besides these, some new species have been introduced from the YK massifs as siblings of their eastern populations, such as Ferula hezarlalehzarica with relatives in the Hindu Kush and Central Asia (Ajani and Ajani 2008) and Inula persica which is related to I. acuminata from the Western Himalaya and Kashmir (Ghahremaninejar and Narimisa 2005). Here we report Bupleurum aitchisonii (Boiss.) Wolff from alpine elevations (3620 m a.s.l.) on Lalezar Mountain, which provides further evidence for such a floristic connection (Fig. 5.6). The general distribution of Bupleurum aitchisonii is in Afghanistan, Tajikistan, Pakistan and India (W Himalaya) and it was not previously reported from Iran (Rechinger and Snogerup 1987; Pimenov and Kljuykov 2015). The material examined is collected in Iran, Kerman province, on the northern slopes of Lalezar Mountain, in a Cousinia multiloba community, 29°25′23”N, 56°46′32″E, ca. 3620 m a.s.l., on 18 July 2018; Doostmohammadi 3118 (MIR).

Bupleurum aitchisonii on Lalezar Mountain (top) and its distribution (bottom; photo MD)
5.5 Endemism
The YK massifs constitute one of the five recognized areas of plant endemism in Iran (Noroozi et al. 2019b). The isolated nature of these mountains together with wide range of elevation have led to a high rate of allopatric speciation and endemism. There are 109 endemic species restricted to the YK massifs, comprising about 8% of the total flora of the area, belonging to 44 genera and 20 families (Noroozi et al. 2019b). Since these mountains are geographically neighboring the Zagros, and especially the lower elevations are well linked, the YK massifs and the Zagros share many species of low and mid elevations. At higher elevations, however, the number of local endemics remarkably increases and the floristic affinity to the other mountains decreases.
Fabaceae (26 species), Asteraceae (24), Plumbaginaceae (13), Lamiaceae (10), Caryophyllaceae (5), Scrophulariaceae (5), Brassicaceae (3), Primulaceae (3) and Ranunculaceae (3) are the richest families in terms of endemics (Fig. 5.2). The newly discovered, monotypic genus Yazdana shirkuhensis (Caryophyllaceae), is endemic to the YK massifs with a restricted distribution in high alpine scree habitats of Shirkuh mountains (Noroozi et al. 2020). There are also some range-restricted genera occur near the borders of the YK massifs. The ditypic genus Karvandarina (Asteraceae) occurs in the south of the YK massifs around Baft by K. aphylla, and the second species, i.e. K. cartilaginea, is distributed in the southwest Iran (Mirtadzadini et al. 2018). The monotypic genus Zerdana (Brassicaceae) is restricted to south and central Zagros and the YK massifs (in the Shirkuh and Khabr mountains). Pseudofortuynia (monotypic, Brassicaceae) with a main distribution in the east of the Zagros reaches also the northwest of the YK massifs (Khosravi 2003).
Astragalus (21 species), Acantholimon (13), Cousinia (8), Echinops (8) and Nepeta (7) have the highest number of endemic species in the area (Fig. 5.3). The general distributions of the endemic–rich genera overlap to some extent and are almost restricted to borders of the Irano–Turkestanian region (Rechinger 1986; Knapp 1987; Doostmohammadi et al. 2018). These species-rich genera have experienced a common climatic and orographic history in Central and SW Asia and share a rather similar historical biogeography (Djamali et al. 2012; Moharrek et al. 2019). In general, it has been suggested that the continued presence of a complex topography since the late Tertiary, which has led to geographical isolation and restricted gene flow, together with a relatively stable continental climate, have had a strong effect on the speciation and current distribution patterns of many species–rich genera of the IT region (Djamali et al. 2012).
Moharrek et al. (2019) have postulated that the center of origin of Acantholimon most probably is located in eastern Iran–Afghanistan, and the high diversity and endemism of this genus in the YK massifs is of high interest. Moharrek et al. (2019) also hypothesized that the predominant speciation factor in Acantholimon in the SW Asia has probably been the allopatric speciation in restricted populations having relatively similar ecological conditions, rather than adaptation to novel environments and different ecological niches. Nepeta is the largest member of the family Lamiaceae in Iran with about 80 species and 60% endemism (Jamzad 2009). In the YK massifs it also has a remarkable number of endemics.
In general, YK has less endemic species compared to the Alborz and the Zagros. The surface area-size of the YK is almost two times bigger than the Alborz (Noroozi et al. 2019b), but the number of species is almost half and the number of endemics is almost a quarter of the Alborz. This is more likely due to the dry climate of the region. The majority of the montane zone is covered with semi-desert steppes which is a unique habitat with poor species diversity. However, with increasing the elevation, especially in high alpine and subninval zone, the situation changes and flora is very isolated with high endemicity (Noroozi et al. 2010, 2019a, b).
As mentioned before many of the endemics are species from high mountains and alpine areas, but there are several endemics distributed in flat plains and lowland habitats (from 1400–2000 m a.s.l.), such as Echinops aucheri, Euphorbia connata, Ferula gabrielii, Linaria iranica, Phlomoides kermanica and Plantago orzuiensis. Some of these species indicate phylogenetic affinities with the SS and/or M floras. The enigmatic Euphorbia connata is closely related to E. calyptrata from north Africa (Riina et al. 2013), Plantago orzuiensis is similar to some SS species like P. ciliata and P. amplexicaulis (Mohsenzadeh et al. 2010) and Linaria iranica belongs to L. sect. Versicolores and is related to the M species L. tenuis (Hamdi et al. 2008).
5.6 Major Vegetation Types
There are no phytosociological studies dealing with the vegetation structure of the region. However, here we briefly present some dominant vegetation types of these mountains based on Zohary (1973) and our own observations.
5.6.1 Psammophytic Vegetation
Since the YK massifs are surrounded by lowland semi-deserts and deserts, it is not surprising to see that several sandy habitats are present in the flat plains of the area. They comprise a wide range of habitats from mobile sand dunes to fixed sand hills, sand–clay alluvial plains and gravel–sand foothills. They extend up to the elevation of 2000 m a.s.l. in some places and receive an average annual precipitation of less than 100 mm. The main shrub species are Haloxylon persicum and different species of Calligonum including C. bungei, C. denticulatum and C. polygonoides. Other characteristic elements of this vegetation type are Astragalus spp. (sect. Ammodendron), Cistanche tubulosa, Euphorbia connata, Nepeta mahanensis, Schumannia karelinii and Spirorhynchus sabulosus.
5.6.2 Halophytic Vegetation
Various halophytic species cover the saline soils in the marginal lowlands and the alluvial plains between the mountain blocks of the area, from 1400 to 2000 m a.s.l. The most prominent halophytic vegetation occurs along the seasonal brackish streams and is composed of Tamarix spp., and mostly annual halophytic species cover the main saline plains, including Climacoptera turcomanica, Halanthium rarifolium, Limonium iranicum, Petrosimonia glauca and Reaumuria persica. Another type of halophytic vegetation in the area is located around Neybid at the deviation of Golbaf–Bam. It is dominated by Nitraria schoberi accompanied by Frankenia pulverulenta, Phragmites australis and Puccinellia sevangensis. Some Central Iranian endemics such as Anabasis haussknechtii and Salsola abarghuensis also occur in some halo–xerophytic communities.
5.6.3 Stipa-Artemisia Steppes
One of the most extensive vegetation types in the area consists of semi–desert and mountain steppes of Stipa-Artemisia (Fig. 5.7). It is a diverse vegetation type, with various accompanying species and it is made up of different pure or intermixed communities depending on elevation, soil properties and climatic conditions at different elevations. This vegetation type is distributed from low montane zone up to the subnival zone (1500 up to 4465 m a.s.l.). Artemisia is the dominant species in most of the stands but the species composition change from low to high elevation gradually. The dominant Artemisia species are A. inculta (based on a wide circumscription including A. sieberi; Podlech 2013) in the lower plains, and A. kermanensis and A. aucheri in higher mountain communities. A. persica is another representative of the mountain steppes, and dominates in wetter areas and around streams. In lower semi–desert plains it is associated or co–dominated by Ajuga chamaecistus, Convolvulus leiocalycinus, Ebenus stellata, Eremostachys macrophylla, Eryngium billardieri, Hertia intermedia, Iris songarica, Launaea acanthodes, Pteropyrum aucheri, Scariola orientalis, Zygophyllum atriplicoides and various annual species. Some representative elements of upper mountains are Acantholimon chlorostegium, A. scorpius, Astragalus anserinifolius, A. huthianus, A. horridus, Centaurea ispahanica, Fibigia umbellata, Jurinea dumulosa, Matthiola alyssifolia, M. flavida, Trachydium depressum, Pterocephalus gedrosiacus, Salvia hydrangea, and perennial grasses like Psathyrostachys fragilis, Stipa haussknechtii and S. hohenackeriana.

Stipa-Artemisia steppes from lower montane up to subnival zone. (a) Artemisia sieberi (Jebal Barez Mts., 1900 m a.s.l.; photo MD); (b) Artemisia aucheri (Hezar Mts., 2700 m a.s.l.; photo JN); (c) Artemisia aucheri (Hezar Mts., 3600 m a.s.l.; photo JN); (d) Artemisia aucheri (Hezar Mts., 4000 m a.s.l.; photo JN)
5.6.4 Riparian, Riverine and Wetland Vegetation
There are several permanent or temporary rivers and streams in the mountains of the area providing suitable habitats for different hygrophilous plant communities. Sekonj valley in Palvar Mountain and Damgahan Valley in Lakhese Mountain are examples of two deep valleys in the area with prominent species on rocky cliffs and hygrophilous vegetation along the rivers. Main species of these riverine communities belong to Berberis, Clematis, Juncus, Rosa and Salix plus Calamagrostis pseudophragmites, Cerasus yazdiana, Dactylorhiza umbrosa, Epipactis veratrifolia, Lonicera nummulariifolia, Nectaroscordum tripedale, Salvia rhytidea and Teucrium melissifolium.
Alpine meadow communities and high-altitude wetlands that in some places reach up to 4000 m a.s.l., are another type of hygrophilous vegetation in the area. One prominent example of these habitats can be seen in Takhte Sartashtak in southwest of Hezar Mountain at 3600 m a.s.l. (Fig. 5.8a). Some characteristic species of these communities are Botrychium lunaria, Cerastium cerastoides, Cirsium rhizocephalum, Gentiana prostrata (Fig. 5.8b), Inula persica, Parnassia cabulica, Pedicularis cabulica, Primula algida (Fig. 5.8c), Taraxacum primigenium and Triglochin palustris.

Wetlands of the YK massifs. (a) High altitude wetland of Takhte Sartashtak (3600 m a.s.l.); (b) Gentiana prostrata; (c) Primula algida, (both from Lalezar, 3500 m a.s.l.; photos MD)
5.6.5 Shrublands and Woodlands
5.6.5.1 Tecomella undulata Communities
At the southern margin of the YK massifs, just at the border of the IT and the SS phytogeographical regions, a community of Tecomella undulata (Fig. 5.9a) trees occurs at the Golparaki plain. These low tree woodlands (2–5 m) at the Golparaki region cover an area of more than 50 hectares with a high cover–abundance, and form the largest population in Iran (Rezanejad and Ganjalikhani Hakemi 2017; Rezanejad et al. 2018). The precipitation is 100–200 mm per year and summer temperature exceeds 45 °C. Acer monspessulanum, Aerva javanica, Amygdalus scoparia, Astragalus fasciculifolius, Convolvulus acanthocladus, Daphne mucronata, Forsskaolea tenacissima, Helianthemum salicifolium, Leysera leyseroides, Otostegia persica, Plantago ovata, Pycnocycla musiformis, Rumex vesicarius and Salvia macilenta are the main accompanying species in this vegetation type.

(a) Tecomella undulata open woodlands in Dalfard valley (1500 m a.s.l.; photo AT); (b) Pistacia atlantica woodlands in Jebal Barez Mountain (photo JN); (c) Juniperus seravshanica woodlands (Galuchar region, 2850 m a.s.l.; photo MD); (d) Rheum ribes community in the Sa’adi Protected Area (2100 m a.s.l.; photo MD)
5.6.5.2 Pistacia–Amygdalus Woodlands
Different species of Amygdalus (in particular A. scoparia, A. elaeagnifolia and A. eburnea) plus Pistacia atlantica and Acer monspessulanum build various types of vegetation in the area while the general composition of each community depends widely on geographical location, soil structure, elevation and water supply. They occur from ca. 1500 to 2900 m a.s.l. with an annual precipitation of 150–350 mm. These communities range from almost pure Pistacia atlantica forests to Amygdalus scoparia woodlands, scrub dominated by A. elaeagnifolia and pure Acer monspessulanum communities (Fig. 5.9b). Intermixed communities with different coverage of various species occur in several areas and further studies are necessary to correctly specify and determine various vegetation types. At some places like Dalfard and the Jebal Barez Mountains, A. scoparia occurs sympatrically with A. eburnea and produce a hybrid, A. × iranshahrii, which contributes to make a remarkable intermixed vegetation unit. Coverage and density of these woodlands vary from place to place and a wide range of species accompany these vegetation stands, such as different species of Astragalus, Acantholimon and Artemisia together with Crataegus meyeri, Colutea persica, Daphne mucronata, Ephedra procera, Ficus johannis, Pistacia khinjuk, Rhamnus persica and Zataria multiflora. Nuts of Pistacia atlantica are much used by local people to prepare a traditional food, and this obviously has a large impact on the soil seed bank of this tree. Additionally, the understory layer is heavily grazed in some places, and that left the soil prone to water erosion. Some representative species of the herb layer are Acanthophyllum glandulosum, Allium cathodicarpum, Biebersteinia multifida, Bunium persicum, Onosma stenosiphon, Picris strigosa, Polygonum spinosum, Senecio glaucus, Steptorhamphus persicus and Tragopogon caricifolius.
5.6.5.3 Juniperus Woodlands
Juniperus communities (Fig. 5.9c) in the YK massifs are represented by J. seravschanica (Adams et al. 2014a; Hojjati et al. 2018). The distribution area of J. seravschanica extends from Kyrgyzstan, Tajikistan and Afghanistan to southeastern Iran (including the YK Mountains) and reaches the mountains of northern Oman over the Persian Gulf (Adams et al. 2014b). It is noteworthy that some populations in the northern blocks of the YK Mountains (Kuhbanan highlands) belong to a probably hybrid species with its main distribution area in central Zagros (Hojjati et al. 2018). Juniperus trees occur on middle to high mountains from 2000–3550 m a.s.l., usually on steep rocky slopes. Very different and variable woody and herbaceous species accompany these communities, such as Acer monspessulanum, Amygdalus elaeagnifolia, Artemisia spp., Acantholimon spp., Astragalus spp., Cotoneaster persicus, Daphne mucronata and Dorema aucheri. Major Juniperus communities in the YK massifs are in the Khabr National park, Dalfard valley, Sardu and Galuchar region.
5.6.6 Rheum ribes Vegetation
The Rheum ribes communities (Fig. 5.9d) occur at several localities including the Sa’adi Protected Area, the plains north of Shahrebabak and south of Mehriz. It covers vast areas of flat plains and foothills at elevations of 2000–2300 m a.s.l. This vegetation type occurs also in the Alborz and on the Azerbaijan Plateau but it dominates in subalpine zone at different habitats and with different species compositions. Species diversity in this vegetation type is very low and woody species are scarce, except for scattered Amygdalus scoparia stands plus Acantholimon spp., Hertia intermedia and Pteropyrum aucheri. At some places, accompanied by Eremurus persicus, it makes beautiful mixed vegetation stands in early spring. Other associated species are mostly ephemerals and include Allium cathodicarpum, Chaenorhinum grossecostatum, Heteroderis pusilla, Holosteum umbellatum, Linaria michauxii, L. micrantha, Minuartia picta, Senecio glaucus, Valerianella szovitsiana and Ziziphora tenuior.
5.6.7 Subalpine Umbelliferous Communities
Giant umbelliferous species associated with various herbaceous and woody species make a very conspicuous tall–herb vegetation in the montane and subalpine zones of the YK massifs (2200–3800 m a.s.l.), in habitats with a high cover of scree and steep inclination. These communities are dominated by variable umbelliferous herbs, such as Dorema aucheri, Ferula assa–foetida (Fig. 5.10a), Ferula hezarlalehzarica (Fig. 5.10b) and Ferulago angulata (Fig. 5.10c), in different areas.

Tall–herb umbelliferous communities. (a) Ferula assa–foetida (Sa’adi Protected Area, 2500 m a.s.l.; photo MD); (b) Ferula hezarlalehzarica (Hezar Mts., 2500 m a.s.l.; photo JN); (c) Ferulago angulata (Hezar Mts., 3300 m a.s.l.; photo JN)
5.6.8 Chasmophytic Vegetation
Rocky walls and cliffs in the area are found from low montane to the subnival zone (2000–4465 m a.s.l.) with different temperature and precipitation regimes. They harbor a considerable and varied diversity of chasmophytic life forms. Rocky communities in the area are mostly formed by Campanula kermanica (Fig. 5.11a), Corydalis rupestris, Crucihimalaya wallichii, Dielsiocharis kotschyi, Dionysia curviflora, D. janthina, D. khatamii, D. oreodoxa, D. revoluta, D. rhaptodes (Fig. 5.11b, c), Graellsia saxifragifolia, Lactuca hazaranensis, L. pumila, Parietaria judaica, Phagnalon nitidum and Rosularia modesta.

Chasmophytic vegetation. (a) Campanula kermanica (Sekonj valley, 2500 m a.s.l.); (b) Close up of Dionysia rhaptodes; (c) Limestone rocks occupied by Dionysia rhaptodes (b & c in Sa’adi Protected Area, 2700 m a.s.l.; photos MD)
5.6.9 Alpine–Subnival Communities
Since the YK Mountains are located in the low latitudes with dry climate, the treeline is higher and the subnival belt starts above 4200 m a.s.l. in Kerman. However, top of the Shirkuh Mts. in Yazd with elevation of 4065 m a.s.l. shows a scattered subnival vegetation. Various alpine and subnival vegetation types can be seen in different areas and at different altitudes. Artemisia aucheri communities are the dominant vegetation types in the subalpine and alpine areas (Fig. 5.7d), with a high species diversity including several species of Astragalus, Euphorbia and Nepeta. Well–developed communities of Cousinia multiloba also can be found. Thorn-cushion communities are located in windswept areas (Fig. 5.12). They are well developed in high alpine zone of the Shirkuh Mts. (Fig. 5.12a, c) but rather scarce in the Hezar-Lalezar (Fig. 5.13b). Dominant species in the Hezar-Lalezar Mountains are Acantholimon haesarensis, Astragalus hezarensis, A. lalesarensis, Linaria remotiflora, Onobrychis cornuta, Polygonum salicornioides, P. spinosum, Semenovia suffrruticosa, Stachys acerosa, together with Nepeta assurgens, Trachydium depressum and Scutellaria multicaulis. Thymus carmanicus occurs in some places on the tops of the mountains and builds communities with high cover–abundance. Erysimum polatschekii (Fig. 5.13a) is one of the recently described species which grows on scree grounds of these habitats. In high alpine zone of the Shirkuh Mts., Artemisia steppes are less developed and real thorn-cushion grasslands are dominant. Remarkable species of these communities are Acantholimon modestum, Alyssum muelleri, Arenaria persica (Fig. 5.12a), Astragalus microphysa, Cousinia lasiolepis, Eremurus persicus (Fig. 5.12b), Helichrysum davisianum, Onobrychis cornuta (Fig. 5.12c), Piptatherum laterale, Scorzonera intricata, Stachys obtusicrena, Thymus carmanicus and Trachydium depressum.

Alpine habitats in the Shirkuh Mts. (a) Arenaria persica (3800–4000 m a.s.l.); (b) Eremurus persicus (3600–3800 m a.s.l.); (c) Onobrychis cornuta (3900 m a.s.l.; photos JN)

Alpine habitats in the Hezar-Lalezar Mts. (a) Erysimum polatschekii (Lalezar Mts., 4100 m a.s.l.; photo MD); (b) Cousinia cf. fragilis, Astragalus lalesarensis (Hezar Mts., 3900 m a.s.l.; photo JN); c & d. Snowbed communities of high alpine zone dominated by Ranunculus eriorrhizus in most of the regions; (c) Lalehzar Mts., 4100 m a.s.l. (photo MD); (d) Shirkuh Mts., 3900 m a.s.l. (photo JN)
Snowbed communities present in these mountains are very scattered in a few depressions, where the snow cover can survive until mid-summer. Ranunculus eriorrhizus (Fig. 5.13c, d) is the most dominant species of these habitats.
Subnival zone is covered mostly by screes and special species adapted to these habitats. Species richness in these communities is low, but there is a high rate of endemism. Allium lalesaricum, Crepis heterotricha subsp. heterotricha, Nepeta lasiocephala (Fig. 5.14a), N. natanzensis, Paracaryum lalezarense, Pseudocamelina camelinae (Fig. 5.14c), Senecio subnivalis, Silene nurensis and Zerdana anchonioides (Fig. 5.14b) are important representative elements of these communities.

Scree communities of subnival zone. (a) Nepeta lasiocephala (Hezar Mts., 4400 m a.s.l.); (b) Zerdana anchonioides (Shirkuh Mts., 4000 m a.s.l.); (c) Pseudocamelina camelinae (Shirkuh Mts., 4000 m a.s.l.; photos JN)
5.7 Conservation
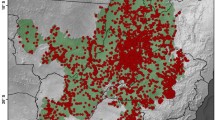
Plant diversity of the YK massifs is facing several threats from both regional and global factors. Grazing has been probably the most important and long–lasting destructive pressure on the flora and vegetation of the Iranian Plateau. It is still a big problem in many parts of Iran, even in the protected areas, and an obstacle for conservation assessments and actions. In the YK massifs as well, all vertical belts are under the impact of grazing herds; low plains in the early spring and high alpine habitats during summer (Fig. 5.15). Over-exploitation of several species, especially traditionally used ones, such as Bunium persicum, Zataria multiflora, Ziziphora clinopodioides and particularly the narrow endemic Dracocephalum polychaetum, is a potential threat for genetic loss and local extinction. Apart from this, the YK flora is under threat of prolonged drought and also global warming. As mentioned before, the rate of endemism is very high at higher elevations due to the strong isolation from other mountains. The impact of global warming on the alpine flora of the YK massifs might in fact be more sever, since these mountains are located at lower latitudes and the high alpine and subnival zones are highly limited and there are no alternative habitats for such species to move up under the impact of climate change. Not only the diversity of plant species, but also the diversity of animal species is very rich in these mountains (Fig. 5.16). Karimi and Jones (2020) estimated that about one quarter of the total area of nature reserves of Iran are under intense human pressure and therefore both expanding protected areas and alleviating anthropogenic activities in the areas under conservation is highly recommended, throughout Iran and particularly in the YK massifs.

Overgrazing (Jebal Barez Mts., 2500 m a.s.l.; photo JN)

Animal diversity of the YK region (Shirkuh Mts.). (a) Polygonia egea (det. Alireza Naderi); (b) Hipparchia parisatis (det. Alireza Naderi); (c) Ochotona rufescens; (d) Paralaudakia microlepis; (e) Laudakia nupta; (f) Laudakia sp.; (g) Ovis orientalis isphahanica (photos JN)
There are one National Park, 11 Protected Areas and two Wildlife Refuges lying in the YK massifs (Fig. 5.17). However, the current nature reserves do not correlate well with plant species hotspots in the region (Noroozi et al. 2019a), and conservation actions in these areas are also very poor, as in many other nature reserves of Iran.

Nature reserves in the YK area (including: 1. Kalmand P.A., 2. Dehaj P.A., 3. Kuhe Bafgh P.A., 4. Kuhe Asiab P.A., 5. Chah Kume P.A., 6. Sa’adi P.A., 7. Bidu P.A., 8. Jupar P.A., 9. Bahr Aseman P.A., 10. Kuhe Shir P.A., 11. Sange Mes P.A., 12. Zaryab W.R., 13. Ruchun W.R. and 14. Khabr N.P.)
Abbreviations
- ES:
-
Euro-Siberian
- IT:
-
Irano-Turanian
- M:
-
Mediterranean
- SS:
-
Saharo-Sindian
- YK:
-
Yazd-Kerman
References
Adams R, Hojjati F, Schwarzbach A (2014a) Taxonomy of Juniperus in Iran: DNA sequences of nrDNA plus three cpDNAs reveal Juniperus polycarpos var. turcomanica and J. seravschanica in southern Iran. Phytologia 96(1):19–25
Adams R, Al-Farsi A, Schwarzbach A (2014b) Confirmation of the southern–most population of Juniperus seravschanica in Oman by DNA sequencing of nrDNA and four cpDNA regions. Phytologia 96(3):218–224
Ajani Y, Ajani M (2008) A new species of Ferula (Umbelliferae) from Southern Iran. Edinb J Bot 65:425–431. https://doi.org/10.1017/S0960428608005052
Ajani Y, Noroozi J, Levichev I (2010) Gagea alexii (Liliaceae), a new record from subnival zone of Southern Iran with key and notes on sect. Incrustatae. Pak J Bot 42:67–77
Assadi M (2009) Four new species of the genus Cousinia Cass. (Asteraceae) from Iran. Iran J Bot 15(1):36–44
Assadi M, Maassoumi AA, Babakhanolou P, Mozaffarian (eds) (1988–2017) Flora of Iran 1–124. Research Institute of Forests and Rangelands Publication, Tehran
Attar F, Mirtadzadini M (2009) Two new species of Cousinia Cass. sect. Pugioniferae Bunge (Asteraceae, Cardueae), from east and southeast of Iran. Iran J Bot 15(2):146–152
Attar F, Ghahreman A, Assadi M (2001) Three new species of the genus Cousinia (Asteraceae, sect. Cynaroideae) from Iran. Nord J Bot 20:697–700
Berberian M, King GCP (1981) Towards a paleogeography and tectonic evolution of Iran. Can J Earth Sci 18:210–265
Bornmüller J (1911) Iter Persico–Turcicum 1892–93. Beitragezur Floravon Persien, Babylonien. Assyrienund Arabien Beihe Bot Central 28(2):89–171
Bornmüller J (1937) Iter Persico–Turcicum 1892–93. Beitragezur Floravon Persien, Babylonien. Assyrienund Arabien Beih Bot Central 57(B):247–294
Bornmüller J (1938) Iter Persico–Turcicum 1892–93. Beitragezur Floravon Persien, Babylonien. Assyrienund Arabien. Beih Bot Centralbl 58(B):252–302
Bornmüller J (1940) Iter Persico–Turcicum 1892–93. Beitragezur Floravon Persien, Babylonien. Assyrienund Arabien. Beih Bot Centralbl 60(B):181–228
Bornmüller J (1942) Iter Persico–Turcicum 1892–93. Beitragezur Floravon Persien, Babylonien. Assyrienund Arabien Beih Bot Centralbl 61(B):72–123
Dashti A, Assadi M, Sharifinia F (2014) A new species of the genus Saponaria L. (Caryophyllaceae) in Iran. Iran J Bot 20(2):146–151
Dewan ML, Famouri J (1964) The soils of Iran. Food and Agriculture Organization of the United Nations, Rome
Djamali M, De Beaulieu JL, Miller N, Andrieu-Ponel V, Ponel P, Lak R, Sadeddin N, Akhani H, Fazeli H (2009) Vegetation history of the SE section of the Zagros Mountains during the last five millennia; a pollen record from the Maharlou Lake, Fars Province, Iran. Veg Hist Archaeobot 18:123–136
Djamali M, Akhani H, Khoshravesh R, Andrieu-Ponel V, Ponel V, Brewer S (2011) Application of the global bioclimatic classification to Iran: implications for understanding the modern vegetation and biogeography. Ecol Meditrr 37:91–114
Djamali M, Baumel A, Brewer S, Jackson ST, Kadereit JW, López-Vinyallonga S, Mehregan I, Shabanian E, Simakova A (2012) Ecological implications of Cousinia Cass. (Asteraceae) persistence through the last two glacial–interglacial cycles in the continental Middle East for the Irano–Turanian flora. Rev Palaeobot Palynol 172:10–20
Doostmohammadi M, Kilian N (2017) Lactuca pumila (Asteraceae, Cichorieae) revisited – additional evidence for a phytogeographical link between SE Zagros and Hindu Kush. Phytotaxa 307:133–140
Doostmohammadi M, Samadi N, Ghorbanalizadeh A (2018) Phytogeography of Genu and Homag; two mountains with an Irano–Turanian flora in the Saharo–Sindian regional zone, S. Iran Nord J Bot e01808. https://doi.org/10.1111/njb.01808
Doostmohammadi M, Mirtadzadini M (2019) Paracaryum lalezarense (Boraginaceae), a new species from high alpine elevations of Lalezar Mountains, in Iran. Phytotaxa 400(5):291–297
Edmondson J, Miller AG, Parris BS (1980) Plants of the Khabr va Rouchun protected areas. Edinb J Bot 38:111–124
Freitag H, Kuhle M (1980) A plant list from the Kuh–e–Jupar (SE. Iran), with some ecological remarks. Willdenowia 10:161–169
Ghahreman A, Mirtadzadini M (2000) Two new species of the genera Jurinea and Scrophularia from Southeast Iran. Iran J Bot 8(2):245–250
Ghahremaninejad F, Narimisa S (2005) Inula persica (Asteraceae: Inuleae), a new species from Kerman province, Iran. Ann Bot Fenn 42:211–213
Gurjazkaite K, Routh J, Djamali M, Vaezi A, Poher Y, Naderi Beni A, Tavakolie V, Kylin H (2018) Vegetation history and human–environment interactions through the late Holocene in Konar Sandal, SE Iran. Quat Sci Rev 194:143–155
Hamdi SM, Assadi M, Vaezi G, Mirtadzadini M (2008) A new species of Linaria Sect. Versicolores (Scrophulariaceae) from Iran. Novon 18(3):340–343. https://doi.org/10.3417/2006010
Hedge I, Wendelbo P (1978) Patterns of distribution and endemism in Iran. Notes Roy Bot Gard Edinburgh 36:441–464
Hojjati F, Kazempour-Osaloo S, Adams R, Assadi M (2018) Molecular phylogeny of Juniperus in Iran with special reference to the J. excelsa complex, focusing on J. seravschanica. Phytotaxa 375(2):135–157
Iran Nejad MH, Shariat Panahi M, Zobeiri M, Marvi Mohajer MR (2001) A Floristic and phytogeographical investigation of Khabr National park and Rouchun wildlife refuge. IJNRR 54(2):111–129. (in Persian)
Jamzad Z (2009) Notes on the genus Nepeta L. (Lamiaceae, Nepetoideae). Iran J Bot 15(2):141–145
Jamzad Z, Ingrouille M, Simmonds MSJ (2003) Three New Species of Nepeta (Lamiaceae) from Iran. Taxon 52(1):93–98
Karimi A, Jones K (2020) Assessing national human footprint and implications for biodiversity conservation in Iran. Ambio. https://doi.org/10.1007/s13280-019-01305-8
Khalili Mobarhan S, Ahmadipour H (2015) Facies studies of Bidkhan stratovolcano (Kerman province). Geosciences 22(95):111–122. (In Persian)
Khodashenas M, Mirtadzadini M, Assadi M (2006) Sisymbrium kermanicum (Brassicaceae: Sisymbrieae), a new species from Iran, Kerman. Iran J Bot 12(1):94–96
Khosravi AR (2003) A reconsideration of Hesperis leucoclada Boiss. (Cruciferae). Iran J Bot 10(1):15–23
Kilian N, Djavadi SB, Eskandari M (2012) Two new mountainous species of Lactuca (Cichorieae, Asteraceae) from Iran, one presenting a new, possibly myrmecochorous achene variant. PhytoKeys 11:61–77. https://doi.org/10.3897/phytokeys.11.2563
Knapp HD (1987) On the distribution of genus Cousinia (Compositae). Plant Syst Evol 155:15–25
Kuhle M (1974) Vorlaufige Ausfuihrungen morphologischer Feldarbeitsergebnisse aus dem SE–Iranischen Hochgebirge am Beispiel des Kuh–i–Jupar. ZfG 18:472–483
Kuhle M (1976) Beitrage zur Quartarmorphologie SE–Iranischer Hochgebirge: Die quartire Vergletscherung des Kuh–i–Jupar. Gottinger Geogrraph Abhandl
Kuhle M (2008) The Pleistocene Glaciation (LGP and pre–LGP, pre–LGM) of SE Iranian mountains exemplified by the Kuh–i–Jupar, Kuh–i–Lalezar and Kuh–i–Hezar Massifs in the Zagros. Polarforschung 77:71–88
Léonard J (1988–1989) Contribution à l’étude de la flore et de la végétation des déserts d’Iran; Fascicule. 8 (étude des aires de distribution les phytochories; les chorotypes) and Fascicule. 9 (Considerapays phytogeographiques sur les phytochories iranotouranienne; saharo–sindienne et de la Somalie–paysMasai). Jardin Botanique National de Belgique, Meise
Léonard J (1991–1992) Contribution a l’etude de la flore et de la vegetation des deserts d’Iran 10, Etude de la vegetation, analyse phytosociologique et phytochorologique des groupements vegetaux. Jardin Botanique National de Belgique, Meise
Léonard J (1993) On the plant "associations" mentioned in Iran by M. Zohary. Bull Jard Bot 62(1):113–118
Maassoumi AA, Mirtadzadini M (2006) Astragalus hotkanensis (Fabaceae, Sect. Microphysa), a new species from Iran. Iran J Bot 12(1):90–91
Maassoumi AA, Vakili SM (2001) New and noteworthy species of the genus Astragalus from Iran. Iran J Bot 9(1):69–70
Mahmoodi M, Maassoumi AA, Noroozi J (2013) A new alpine species and a new record of Astragalus sect. Stereothrix (Fabaceae) from Iran, with comments on the phytogeography of the section. Willdenowia 43:263–270
Malekpourzadeh L, Mirtadzadini M, Nazeri V (2015) The study of flora of Ra’skuh Area in Baft township in Kerman province. TBJ 23:85–94. (In Persian)
Mehdipour Ghazi J, Moazzen M (2015) Geodynamic evolution of the Sanandaj–Sirjan Zone, Zagros Orogen, Iran. Turk JEarth Sci 24:513–528
Miller NF, Kimiaie M (2006) Some plant remains from the 2004 excavations of Tall–e Mushki, Tall–e Jari A and B, an Tall–e Bakun A and B. In: Alizadeh A (ed) The origins of state organizations in prehistoric Highland Fars, Southern Iran. Excavations at Tall–e Bakun, no 128. Oriental Institute Publications, pp 107–118
Mirtadzadini M, Attar F (2004) Cousinia (sect. Spinuliferae) hazarensis (Compositae, Cardueae), a new species from SE Iran. Willdenowia 34:191–194. https://doi.org/10.3372/wi.34.34117
Mirtadzadini M, Attar F (2014) Cousinia kermanensis and C. pulcherantha spp. nov. (Compositae, sect. Congestae and sect. Leiocaules) from southeastern Iran. Nord J Bot 32:279–284
Mirtadzadini M, Attar F, Assadi M (2004) Cousinia subpectinata (Asteraceae: plathyacanthae), a new species from W Kerman province. Iran J Bot 10(2):143–146
Mirtadzadini M, Bordbar F, Parishani MR, Vitek E, Rahiminejad MR (2018) Novelties in Cardueae (Asteraceae). Nova Biologica Reperta 5(3):337–348
Moharrek F, Sanmartin I, Kazempour-Osaloo S, Feliner GN (2019) Morphological innovations and vast extensions of mountain habitats triggered rapid diversification within the species–rich Irano–Turanian genus Acantholimon (Plumbaginaceae). Front Genet 9:698
Mohsenzadeh S, Nazeri V, Mirtadzadini SM (2007) Plantago lachnantha Bge. (Plantaginaceae), a new record for flora of Iran. Iran J Bot 13(2):107–108
Mohsenzadeh S, Nazeri V, Mirtadzadini M (2010) A new species of Plantago (Plantaginaceae) from Iran. Novon 20(3):307–310
Moazzeni H, Zarre S, Assadi M, Joharchi MR, German DA (2014) Erysimum hezarense, a new species and Rhammatophyllum gaudanense, a new record of Brassicaceae from Iran. Phytotaxa 175:241–248. https://doi.org/10.11646/phytotaxa.175.5.1
Moazzeni H, Assadi M, Zare G, Mirtadzadini M, Al-Shehbaz I (2016) Taxonomic novelties in Erysimum for the flora of Iran: E. polatschekii, a new alpine endemic, and E. scabrum, a new record. Phytotaxa 269:47–53. https://doi.org/10.11646/phytotaxa.269.1.6
Mozaffarian V (2002) Studies on the flora of Iran, new species and new records. Pak J Bot 34(4):391–396
Nejad Falatoury A, Assadi M, Ghahremaninejad F (2016) Gypsophila yazdiana (Caryophyllaceae), a New Species from Iran. Novon 24(4):347–351. https://doi.org/10.3417/2015014
Nordenstam B, Moussavi M, Djavadi B (2002) A new annual species of Senecio from Iran. Compos Newsl 38:42–46
Noroozi J, Akhani H, Breckle SW (2008) Biodiversity and phytogeography of the alpine flora of Iran. Biodivers Conserv 17(3):493–521
Noroozi J, Ajani Y, Nordenstam B (2010) A new annual species of Senecio (Compositae–Senecioneae) from subnival zone of Southern Iran with comments on phytogeographical aspects of the area. Compos Newsl 48:43–62
Noroozi J, Pauli H, Grabherr G, Breckle SW (2011) The subnival–nival vascular plant species of Iran: a unique high–mountain flora and its threat from climate warming. Biodivers Conserv 20:1319–1338
Noroozi J, Talebi A, Doostmohammadi M, Rumpf SB, Linder HP, Schneeweiss GM (2018) Hotspots within a global biodiversity hotspot – areas of endemism are associated with high mountain ranges. Sci Rep 8(1):10345. https://doi.org/10.1038/s41598-018-28504-9
Noroozi J, Naqinezhad A, Talebi A, Doostmohammadi M, Plutzar C, Rumpf SB, Asgarpour Z, Schneeweiss GM (2019a) Hotspots of vascular plant endemism in a global biodiversity hotspot in Southwest Asia suffer from significant conservation gaps. Biol Conserv 237:299–307. https://doi.org/10.1016/j.biocon.2019.07.005
Noroozi J, Talebi A, Doostmohammadi M, Manafzadeh S, Asgarpour Z, Schneeweiss GM (2019b) Endemic diversity and distribution of the Iranian vascular flora across phytogeographical regions, biodiversity hotspots and areas of endemism. Sci Rep 9:12991. https://doi.org/10.1038/s41598-019-49417-1
Noroozi J, Pirani A, Moazzeni H, Mahmoodi M, Zare G, Noormohammadi A., Barfuss MHJ, Suen M and Schneeweiss GM (2020) The new locally endemic genus Yazdana (Caryophyllaceae) and patterns of endemism highlight the high conservation priority of the poorly studied Shirkuh Mountains (Central Iran). JSE (Accepted)
Pahlevani AH, Feulner M, Weig A, Liede-Schumann S (2017) Molecular and morphological studies disentangle species complex in Euphorbia sect. esula (Euphorbiaceae) from Iran, including two new species. Plant Syst Evol 303:139–164
Payandeh M, Bordbar F, Mirtadzadini M (2016) Floristic study of Hanza–kuh of Bahr–Aseman protected area (SE Iran). TBJ 28:790–100. (In Persian)
Pimenov M, Kljuykov E (2015) A new species of Bupleurum (Umbelliferae) from Badakhshan, with notes on and a key to the alpine species of High Asia. Phytotaxa 226(3):245–252
Podlech D (2004) New species of Astragalus L. (Leguminosae), mainly from Iran. Ann NatHist Mus Wien 105B:565–596
Podlech D (2013) Some remarks on Artemisia subgenus Seriphidium (Asteraceae) mostly from Afghanistan. Rostaniha 14:48–58
Podlech D, Massoumi AA (2003) New species of Astragalus L. (Fabaceae) from Iran, mainly of sects. Incani and Malacothrix. Feddes Repert 114:320–349
Podlech D, Zarre S (2003) New species of Astragalus sect. Ammodendron (Fabaceae) with a key for the species of the Flora Iranica area. Willdenowia 33(2):341–352
Rajaei P, Maassoumi AA, Mozaffarian V, Nejad Sattari T, Pourmirzaei A (2011) Alpine flora of Hezar mountain (SE Iran). Rostaniha 12(2):111–127
Rajaei P, Mozaffarian V (2016) A new species of Helichrysum (Asteraceae, Gnaphalieae) from Iran. Iran J Bot 22(1):08–10
Ranjbar M (2011) Astragalus baftensis (Fabaceae), a new species of Astragalus sect. Pendulina from Iran. Iran J Bot 17(1):1–5
Ranjbar M, Karamian R (2004) Taxonomic study of Astragalus sect. Erioceras (Fabaceae) in Iran; Additional notes and key to the species. Nord J Bot 22:713–717
Ranjbar M, Mahmoudi C (2015) A revision of Phlomoides sect. Thyrsiflorae (Lamiaceae). Webbia 70(2):19. https://doi.org/10.1080/00837792.2015.1041794
Rechinger KH (ed) (1963–2015) Flora Iranica 1–174. Akademische Druck– u. Verlagsanstalt, Graz; 175. Akademische Verlagsgesellschaft, Salzburg; 176–181. Verlag des Naturhistorischen Museums, Wien
Rechinger KH (1986) Cousinia, morphology, taxonomy and phytogeographical implications. Proc R Soc Edinb 89B:45–58
Rechinger KH, Snogerup M (1987) Bupleurum (pp. 269–297) [Apiaceae]. In: Rechinger KH (ed) Flora Iranica 162. Akademische Druck–und Verlagsanstalt, Graz
Rezanejad F, Ganjalikhani Hakemi F (2017) Studies of Pollen Characteristics in Plants of Fruitless Tecomella undulata (Sm.) Seem. (Bignoniaceae) in Golparaki Region of Jiroft City, Iran. Iran J Sci Technol Trans Sci 41(1):979–988
Rezanejad F, Saberi A, Alimoradi F (2018) Morphology, ecology and propagation of endangered species of Tecomella undulata (Sm.) Seem (Bignoniaceae) in Golparaki plain of Jiroft (Iran). J Devell Biol 10(3):1–14. (In Persian)
Riina R, Peirson JA, Geltman DV, Molero J, Frajman B, Pahlevani A, Barres L, Morawetz JJ, Salmaki Y, Zarre S, Kryukov A, Bruyns PV, Berry PE (2013) A worldwide molecular phylogeny and classification of the leafy spurges, Euphorbia subgenus Esula (Euphorbiaceae). Taxon 62(2):316–342
Saberi A, Hasanabadi Z, Mirtadzadini M, Nazeri V (2013) A study of the flora of Riseh and Paqal’e area Shahrebabak of Kerman, Iran. TBJ 14:67–78. (In Persian)
Sonboli A, Olanj N, Pourmirzaei A (2011) Biosystematics and phylogeny of Tanacetum fisherae, a new record from Iran. Rostaniha 12(2):165–175
Stöcklin J (1968) Structural history and tectonics of Iran: a review. Am Assoc Pet Geol Bull 52:1229–1258
Van Zeist W (1967) Late Quaternary vegetation history of western Iran. Rev Palaeobot Palyn:2301–2311
Van Zeist W, Bottema S (1977) Palynological investigations in western Iran. Palaeohistoria 19:19–85
Van Zeist W, Wright HE (1963) Preliminary pollen studies at Lake Zeribar, Zagros Mountains, southwestern Iran. Science 140:65–67
Zarre S, Podlech D (2001) Taxonomic revision of Astragalus sect Acanthophace (Fabaceae). Sendtnera 7:233–253
Zarezadeh M, Mirvakili SM, Mirhosseini A (2007) Introduction to the flora, Life form and plant geographical distribution of Darrah Damgahan in Mehriz (Yazd province). Pajouhesh-va-Sazandegi 74:129–137
Zohary M (1963) On the geobotanical structure of Iran. Weizman Science Press of Israel, Jerusalem
Zohary M (1973) Geobotanical foundations of the Middle East 1–2. Gustav Fischer Verlag, Stuttgart
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Doostmohammadi, M., Talebi, A., Noroozi, J. (2020). The Yazd–Kerman Massifs. In: Noroozi, J. (eds) Plant Biogeography and Vegetation of High Mountains of Central and South-West Asia. Plant and Vegetation, vol 17. Springer, Cham. https://doi.org/10.1007/978-3-030-45212-4_5
Download citation
DOI: https://doi.org/10.1007/978-3-030-45212-4_5
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-45211-7
Online ISBN: 978-3-030-45212-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)