
Abstract
Recent findings of noticeable changes in diatom preservation in the Holocene record of Pampean shallow lakes evidenced the need for carrying out actualistic studies to decipher the environmental significance of taphonomic signatures. In this chapter, we review a series of field and experimental studies recently conducted, focusing on the effect of contemporary environmental gradients on the dissolution and fragmentation of diatom valves. Field studies signaled salinity, carbonates, and bicarbonates as the main drivers of dissolution of the target taxon Cyclotella meneghiniana in shallow lakes covering wide environmental gradients. Laboratory controlled experiments demonstrated a negative effect of NaCl and HCO3Na on valve preservation, evidenced by an increase in dissolved silica and by the occurrence of valves showing advanced stages of dissolution. Detailed analysis of taphonomic attributes and their relationship with live/dead agreement at a freshwater lake showed that within-lake taphofacies are useful to discriminate between diatom sub-environments. The joint analysis of compositional and taphonomic variations in the recent sedimentary record of the lake demonstrated the usefulness of taphonomic analyses to uncover subtle paleoenvironmental variations, which could be overlooked if only traditional compositional analyses were performed. The strong link between compositional and taphonomic patterns at different working scales highlighted the usefulness of including taphonomic analyses when conducting diatom-based paleoenvironmental studies in environmentally heterogeneous shallow lakes.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
6.1 Introduction
Diatoms (Bacillariophyceae) are among the most ubiquitous bioindicators used to reconstruct paleoenvironmental changes in freshwater environments. These eukaryotic algae are small (2–500 µm) and diverse (>24,000 known species) and can occupy a variety of planktonic, benthic, and subaerial habitats (Julius and Theriot 2010). Since the recognition of diatoms as potential indicators of water pollution and paleoenvironmental changes in the late nineteenth century, diatom analyses of lake sediments became increasingly used as a tool to understand different aspects of lake dynamics, water acidification, eutrophication, and climate change (see Battarbee et al. 2001 and references therein). Although the inner contents of diatom cells are similar to other eukaryotic algae, the uniqueness of diatoms relies on the composition and morphology of their cell walls, which constitute their preservable parts and can thus be incorporated to the fossil record (Battarbee et al. 2001). The cell wall, known as the frustule, is composed of silica (SiO2) arranged in two halves (i.e. valves) and several bands or belts known as the griddle bands. The frustule has many openings allowing the contact between the cytoplasm and the surrounding environment, which can vary considerably in shape and architecture among species, from simple pores to specialized structures of extremely complex micro-architecture (Julius and Theriot 2010). Overall, given its particular chemical composition, small size, and variability in shape and structure, the taphonomic constraints to the preservation of the diatom frustule can differ strongly from those of carbonatic macro-remains (such as mollusks), which have been the main focus of taphonomic research since the establishment of taphonomy as a discipline.
The recognition of taphonomy as a key factor in structuring the diatom sedimentary record has a long history (e.g. Lewin 1961). Removal via outflows, transport from upstream sources, resuspension and reworking of sediments within lake basins, sediment bioturbation, and silica dissolution have been advocated as potential causes of taphonomic biases in continental settings (Battarbee 1986). Among them, dissolution has received most of the attention, given the relevance of this process in saline and alkaline environments (Ryves et al. 2001). Dissolution is enhanced in solutions with low silica concentrations (e.g. Flower and Ryves 2009), elevated temperature, pH (e.g. Lewin 1961; Barker 1992; Flower 1993), and salinity (e.g. Mikkelsen 1977; Barker et al. 1994). These processes can alter not only total diatom abundances in sediments but also assemblages composition (Warnock and Scherer 2015), as species differ in their preservation potential, being delicate and less silicified taxa highly sensitive to cell wall dissolution (Fritz 2007). Therefore, dissolution can lead to an underestimation of the relative abundances of these delicate taxa, biasing the diatom record towards robust species (Ryves et al. 2009). Contrastingly, the causes of fragmentation of diatom frustules have been relatively poorly studied, (e.g. Haberyan 1985; Ryves et al. 2006). Higher fragmentation has been associated with high energy environments, such as littoral and shallow lacustrine zones (Round 1964; Flower and Nicholson 1987). The effect of grazing has also proven to be significant in causing frustule breakage (Haberyan 1985). Therefore, biological and physical environmental variables, such as fetch, depth, wind speed and grazers abundance may play a key role in explaining diatom breakage. Overall, the joint effect of biological, chemical and physical factors in promoting diatom breakage and dissolution can be significant in continental settings, particularly in saline, alkaline and very shallow lakes with high nutrient content, being these changes highly dependent on local taxonomy and variable among environments and regions.
The study of diatoms as a tool to understand Holocene environmental changes in the Pampean Region has increased significantly during the last decade (García-Rodríguez et al. 2009; Stutz et al. 2010; Hassan 2013; Hassan et al. 2011, 2012, 2014). In these studies, the shifts in the composition of sedimentary diatom assemblages during the last ca. 5000 cal. yrs BP were addressed and interpreted as indicative of past changes in salinity and nutrient status of the lakes. Diatom records showed a shift from brackish/saline and low trophic conditions at the mid-Holocene towards freshwater/eutrophic conditions developed since ca. 1500–1200 cal. yrs BP (Hassan et al. 2011, 2014; Hassan 2013). Hassan et al. (2014) observed that these shifts in diatom assemblages composition were also accompanied by noticeable changes in preservation, evidenced by marks on frustule breakage and dissolution. These changes were explained as a consequence of past variations in lake salinity, suggesting the potential use of taphonomic signatures as sources of paleoenvironmental information. These results pointed out the significance of incorporating taphonomic analyses when conducting paleoenvironmental studies and highlighted the need for conducting actualistic studies in Pampean shallow lakes, in order to elucidate the environmental significance of diatom taphonomic signatures.
Under this scenario, we have recently started a research project on actualistic taphonomy of diatoms in Pampean shallow lakes, in order to assess the effect that physicochemical lake characteristics exert on valve preservation. The project included controlled laboratory experiments and field research covering regional and local scales. By conducting this project, we intend to combine the information provided by diatom taphonomy with the classic inferences based on community composition, in order to develop new tools to improve the quality of future paleoenvironmental studies in the area. In this chapter, we review the advances of a series of studies conducted in the context of this project during the last four years and discuss the expected future developments in this field of research.
6.2 Environmental Significance of Modern Diatom Assemblages
The quality of diatom-based paleoenvironmental reconstructions relies on a complete knowledge of the environmental requirements and taphonomic biases of the species found as fossils. Reconstructions assume that environmental requirements of the fossil diatom taxa used as bioindicators have remained constant during the period considered and, consequently, are similar to those of their closest living representatives (taxonomic uniformitarianism, Dodd and Stanton 1990). In this way, the environmental information obtained from living organisms can be used as modern analogs and extrapolated to the fossil record, particularly in Quaternary research (Hassan et al. 2011). Hence, the study of modern diatom-environment relationships constitutes an essential first step to gather ecological information, although the application of this data to paleoenvironmental reconstruction should also consider the biases suffered by the information contained in assemblages during their fossilization.
Pampean diatoms in modern shallow lakes have demonstrated a great potential as proxies for paleoenvironmental reconstructions, as their distribution is strongly related to key environmental variables (Hassan et al. 2009, 2011). Assemblages are mostly dominated by benthic and epiphytic taxa, because of the very shallow depth and high vegetation cover that characterize these environments. Tychoplanktonic taxa (i.e. taxa that are closely related to benthic habitats, occurring intermittently in the water column after being swept up from the bottom; Smol and Stoermer 2010) are also abundant in these wind-stressed shallow lakes, being Cyclotella meneghiniana one of the most widely distributed diatom species in the region (Hassan and De Francesco 2018). In a study covering Pampean shallow lakes and streams, the composition of assemblages was strongly linked to conductivity, pH and nutrient gradients (Hassan et al. 2009). When integrated with datasets from the adjacent Monte and Espinal regions (i.e. semiarid regions located at the west of the Pampean Region, see Fig. 2 in Hassan et al. 2011), the longer gradient of temperature was also significantly linked to diatom composition (Hassan et al. 2011). Overall, the strong relationship between diatom assemblages composition and environmental variables directly (temperature) or indirectly (conductivity and pH) linked to climate enhance their potential application to paleoclimatic reconstruction, provided they were faithfully preserved in the sedimentary record. Moreover, the significant role of nutrients in structuring modern communities also emphasizes their use as biomonitors of anthropogenic impacts in the region (Hassan et al. 2009).
Internal within-lake variability in diatom assemblages composition is high in these lakes, as a consequence of their characteristic environmental heterogeneity (Hassan 2018). The extensive development of the littoral zone relative to the pelagic zone promotes the development of complex lateral environmental gradients, which support diverse and productive periphytic and benthic communities (Wetzel 2001). Consequently, diatom assemblages in these lakes allow to distinguish between littoral and open-waters habitats, as they differ in species composition. Tychoplanktonic taxa, such as Cyclotella meneghiniana , Aulacoseira granulata and A granulata var. angustissima tend to be more abundant in open-waters, whereas epiphytic and benthic taxa usually dominate littoral sediments (Hassan 2018). Moreover, the analysis of epiphytic assemblages inhabiting five macrophyte taxa from Nahuel Rucá lake demonstrated that diatom species can also be used to discriminate between free-floating and submersed/emergent vegetation (Rojas and Hassan 2017). Hence, besides its potential use to reconstruct regional environmental gradients, these assemblages can also be used to discriminate among within-lake subenvironments, providing useful tools for the recognition of subtle environmental changes in the fossil record (Hassan et al. 2018).
This good performance of Pampean diatom assemblages as modern analogs for past environmental conditions opened interrogations about the impact of taphonomic processes in structuring the sedimentary record of these bioindicators, which laid the foundations of our project on actualistic taphonomy. Particularly, the main questions to be addressed were: (1) Is the compositional and environmental information stored by living communities faithfully reflected by death assemblages? (2) Can taphofacies analyses be used to discriminate between the observed local and regional environmental gradients? (3) Which are the environmental causes of diatom taphonomic signatures? and (4) Can diatom-based paleoenvironmental inferences be improved by combining the use of taphonomic and compositional lines of evidence? In the following sections, we will discuss our main advances directed to answer these questions.
6.3 Ecological Fidelity of Death Assemblages
Understanding the faithfulness with which sedimentary diatom assemblages represent the composition of the source communities from which they were derived constitutes a key point in diatom analysis (Battarbee et al. 2001). Diatom-based paleoenvironmental reconstructions are based on the uniformitarian transference of ecological information from modern taxa to the past and assume that death assemblages integrate small-scale temporal and spatial variability of living communities into time-averaged assemblages (Hassan et al. 2008). However, as previously mentioned, diatom assemblages are subjected to a wide array of taphonomic biases in shallow lakes, which may lead to a low compositional agreement between living communities and death assemblages. In fact, if the variability introduced by taphonomic processes exceeds the original biological variability, then the capability of death assemblages to preserve the original ecological information might be completely obliterated. Likewise, these taphonomic processes might shift the record of the responses of diatom assemblages to environmental gradients, thus testing the assumptions of paleoecological and paleoenvironmental analyses (Tomašových and Kidwell 2009b).
Live-dead comparisons are the most common actualistic methods of evaluating the faithfulness of the preservation of ecological information (Kidwell 2013). In live-dead studies, the ecological properties of the life assemblages (LAs) are compared with those of death assemblages (DAs, Kidwell 2013). They have mostly focused on compositional fidelity (i.e., preservation of composition, richness and abundance, Tomašových and Kidwell 2009a, 2011), whereas works on environmental fidelity (i.e., the ability of death assemblages to detect environmental gradients) have been particularly scarce (Tomašových and Kidwell 2009b). In order to assess the degree of ecological fidelity exhibited by death assemblages in Pampean shallow lakes, Hassan (2015) analyzed three live-dead diatom datasets. Two main points were evaluated: (1) the ability of death assemblages to preserve the composition of the original living community (compositional fidelity); and (2) the ability of death assemblages to preserve the original responses of living communities to environmental gradients (environmental fidelity).
6.3.1 Compositional Fidelity
Compositional fidelity of sedimentary diatom assemblages was addressed in three shallow lakes located in the southeastern Pampa Plain: Las Mostazas, Los Carpinchos and Nahuel Rucá (Fig. 6.1, see De Francesco et al. this volume for a description of the Study Area). These are typical Pampean shallow lakes, as they are small (~250 ha), very shallow (maximum depths of 1.5–2 m), hypereutrophic, and surrounded by littoral zones densely populated by emergent and submersed macrophytes, forming a ring that surrounds entirely the lakes and clearly differentiates littoral from open waters areas. The three lakes are located along a north-south gradient covering approximately 60 km, and range from freshwater to slightly brackish salinity (0.3–4.3 mS/cm). A total of 132 surface sediment samples were collected seasonally from each lake’s littoral and open waters subenvironments, each one representing a live-dead pair.

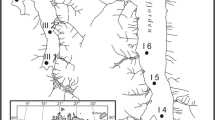
Location map showing all the lakes sampled (yellow circles) in the regional taphofacies analysis conducted by Hassan and De Francesco (2018). The situation of the three lakes used in fidelity, within-lake taphofacies, and modern-fossil comparisons are highlighted (red squares, modified after Hassan and De Francesco 2018). NR: Nahuel Rucá Lake; LM: Las Mostazas Lake; LC: Los Carpinchos Lake
Compositional fidelity was assessed by comparing richness, diversity, abundances and multivariate dispersions among life (i.e. cells containing intact chloroplasts) and death assemblages. These estimates differed significantly between LAs and DAs, indicating a broadly unfaithful representation of diatom communities by death assemblages. The observed differences were interpreted mainly as consequences of (1) differences in the temporal resolution between time-averaged DAs and non-averaged LAs, and (2) differential preservation of diatom taxa related to the intrinsic properties of their valves, specifically the over-representation of robust species and under-representation of fragile ones. The influence of time averaging was expected, however, as DAs constitute time-averaged representations of living communities and integrate dead-valves inputs over long periods of time (Kidwell 2002; Tomašových and Kidwell 2011). Even when diatom living communities are sampled in a seasonal basis, the samples collected in each lake would represent only snapshots of the full seasonal variability, which can be high in diatoms given their very short generation times (from hours to a few days, Baars 1981). Enrichment of death assemblages can also be related to the input of valves from lake subenvironments not included in the sampling strategy, such as epiphytic taxa living upon macrophytes (e.g., Lemnicola hungarica and Cocconeis placentula, Rojas and Hassan 2017). Hence, it was expected that, given longer sampling periods and more intensive sampling of all lake subenvironments, pooled community richness and diversity values would more closely approximate those of the DAs.
Further examination of the differences in the proportional abundances of taxa between LAs and DAs allowed identifying species which were significantly under or over-represented in death assemblages. Most of the taxa over-represented in DAs (i.e., Navicula peregrina , Surirella striatula , Aulacoseira granulata , Cyclotella meneghiniana ) were characterized by highly silicified and robust valves, whereas under-represented taxa (i.e., Nitzschia amphibia , Pseudostaurosira americana , Aulacoseira granulata var. angustissima ) were weakly silicified, and more prone to suffer fragmentation given their elongated shape and denser areolation patterns (Fig. 6.2). Given the very shallow nature of these lakes, their bottom sediments are likely subjected to significant reworking by wind action, being the resulting turbulence a possible cause of breakage and subsequent dissolution for delicate taxa (Ryves et al. 2006). As an example, the under-representation of Aulacoseira granulata var. angustissima and the over-representation of A. granulata in DAs of Nahuel Rucá lake were accompanied by differences in the preservation of both species. Valves of the delicate Aulacoseira granulata var. angustissima showed less fragmentation than A. granulata, as they constitute recently dead remains and tend to remain a shorter time in surface sediments before being more easily destroyed (Fig. 6.2). Hence, as the probability of breakage of diatom valves is strongly related to their length/width ratio and pore row frequency (Haberyan 1985), differential fragmentation of taxa in these wind-stressed shallow lakes is more likely responsible for the observed deviations in proportional abundances.

Compositional fidelity analyses of Nahuel Rucá open waters samples showing a the proportional abundances in life assemblages (LA) versus the proportional abundances in death assemblages (DA) for Aulacoseira granulata var. angustissima (ANGU, white circles) and Aulacoseira granulata (AGRA, black circles). The under-representation of ANGU and the over-representation of AGRA in death assemblages are evidenced by the displacement of samples from the 1:1 expected relationship (dotted line, modified after Hassan 2015). Examination of the Diatom Fragmentation Index (DFI) in the same samples b showed a significantly higher fragmentation of AGRA (Kruskal-Wallis p < 0.001), which tend to remain as identifiable fragments in the sediments long before the destruction of ANGU valves (modified after Hassan et al. 2018). This difference in the preservation of both taxa can be explained by the larger length/width ratio and pore row frequency exhibited by ANGU (d) when compared to AGRAN (c)
6.3.2 Environmental Fidelity
Given the low compositional fidelity exhibited by DAs, the question of whether this unfaithfulness affects the environmental information stored by sedimentary assemblages became a significant issue for diatom-based paleoenvironmental reconstructions in these lakes. If the environmental significance of DAs became biased during its formation, then the inference of environmental parameters from sedimentary diatom composition should incorporate these biases to avoid interpretative errors. Hence, the degree to which DAs capture the original environmental patterns affecting the structure of LAs was evaluated through comparisons at local (within-lake) and regional (between-lake) scales from the same three lake dataset (Hassan 2015).
Results of gradient analyses demonstrated that, despite this low compositional fidelity, DAs were able to capture gradients of conductivity, pH, depth, and vegetation even better than LAs, as the percentage of variance in diatom composition explained by these environmental variables was higher in DAs than in LAs, both at regional and local scales. This higher environmental sensitivity of DAs over LAs has been also mentioned in mollusk-based environmental fidelity studies and related to the capability of DAs to capture community composition (especially rare species) over longer temporal durations (see Tomašových and Kidwell 2009b and references therein). In the case of the studied datasets, it was also related to the structural redundancy exhibited by diatom communities (i.e., more than one mutually exclusive subset of species significantly captures community structure based on the full set of species, Clarke and Warwick 1998). For example, the under-representation of Aulacoseira granulata var. angustissima in DAs of Nahuel Rucá lake was compensated by the over-representation of A. granulata in the same assemblages, as both taxa responded similarly to the studied environmental gradients (Fig. 6.2). In this case, redundancy in environmental information implies that the between-sample relationships of life assemblages can be significantly preserved by DAs even if some taxa are randomly removed by taphonomic processes or are missed due to incomplete sampling (Tomašových and Kidwell 2009b). Overall, this good live-dead agreement implies that past environmental conditions inferred from fossil diatom assemblages would be even more accurate than those based on a single sampling of the original living community in these lakes, supporting one of the main assumptions of diatom-based paleoenvironmental reconstructions. However, the good preservation of environmental gradients observed in diatom assemblages did not imply good preservation of environmental characteristics not considered in that study, such as nutrient status, or longer conductivity and pH gradients, highlighting the importance of conducting more studies to cover new environmental gradients of paleoenvironmental interest.
6.4 Taphofacies Analyses
The low live-dead compositional agreement exhibited by diatom assemblages in Pampean shallow lakes suggested a possible role of taphonomic processes in the structuration of the diatom record. Moreover, the variability in dissolution and fragmentation profiles observed in the fossil record of these lakes pointed valve dissolution and breakage as probable causes of biases in assemblages composition (Hassan et al. 2014), highlighting the importance of analyzing the causes of diatom taphonomic signatures in modern settings. Accordingly, a series of studies intending to assess the environmental significance of diatom dissolution and breakage marks were conducted in a number of Pampean shallow lakes. The purpose was to identify potential taphonomic biases (i.e. postmortem information loss), but also to assess the positive contributions of taphonomic signatures to the interpretation of past environmental changes. Taphofacies analysis (i.e., facies defined on the basis of diagnostic taphonomic traits; Speyer and Brett 1986) constitute a powerful methodology in that direction, as it allows to underscore the non-random relationship between taphonomic signatures and environmental conditions in modern settings, highlighting the feasibility of reconstructing paleoenvironments from taphonomic analyses (Parsons and Brett 1991). Accordingly, the potential use of diatom taphofacies to infer environmental conditions was assessed at (1) regional (i.e. between-lake comparisons; Hassan and De Francesco 2018), and (2) local (i.e. within-lake comparisons; Hassan et al. 2018) scales in modern shallow lakes.
6.4.1 Regional Scales: Between-Lake Taphofacies Comparisons
The impact of long environmental gradients on diatom preservation was assessed in twenty-five lakes of the Pampean Region (Fig. 6.1) representing a strong salinity gradient (0.4–13.2 ppt, Hassan and De Francesco 2018). The preservation was evaluated through the analysis of frustule fragmentation and dissolution on a target taxon, Cyclotella meneghiniana . Given the strong dependency of preservation on valve morphology, the use of a target taxon such as C. meneghiniana allowed to assess changes in preservation independently of the assemblage composition. This taxon had already been used as an indicator of diatom dissolution (e.g. Hassan et al. 2014), not only because of its abundance in modern and past environments but also for its moderate sensitivity to dissolution and its easily recognizable valves, which can be identified even under very poor preservation conditions (Barker 1992). The study of its preservation in Pampean lakes indicated a significant relationship between valve dissolution, salinity, pH, carbonate, bicarbonate and silica concentrations, whereas environmental constraints on valve fragmentation were less clearly identified (Fig. 6.3). According to Generalized Linear Model (GLM) analyses, dissolution was negatively correlated to silica concentration, and positively to the rest of these variables, whereas fragmentation was only marginally correlated to salinity (Fig. 6.3).

Bivariate plots of selected environmental variables (log-transformed) and taphonomic indices along a regional gradient covering 21 lakes (modified after Hassan and De Francesco 2018). Significant relations, according to GLM analyses are highlighted, and significances at p < 0.001 (***), p < 0.01 (**), and p < 0.05 (*) are indicated by asterisks. DDI: Diatom Dissolution Index (average dissolution grade of the assemblage, Ryves et al. 2006), DFI: Diatom Fragmentation Index (average breakage grade of the assemblage, Hassan et al. 2014), Fd: F index of dissolution (ratio of undissolved valves to all valves counted, Ryves et al. 2009), Fb: F index of fragmentation (ratio of unbroken valves to all valves counted, Ryves et al. 2009), TDS: Total Dissolved Solids (ppt)
The application of further direct gradient analyses (Redundancy and Canonical Correspondence Analyses, ter Braak 1994) allowed to assess and compare the percentage of variance in compositional and taphonomical data explained by environmental variables. Salinity explained significant portions of variance both in taphonomic and compositional analyses, although the effect on assemblage composition was only marginal and lower: while it explained a 33.3% of the taphonomic variation (p < 0.001), only an 11.7% of the compositional variance was explained by this environmental variable (p < 0.01). These results were comparable to previous regional studies covering a wider region (Central Argentina), in which log-Conductivity accounted for 4.8% of the variance in diatom assemblage composition (Hassan et al. 2011). These findings indicated that although this environmental factor has an influence in structuring the composition of diatom living communities in Pampean shallow lakes, it plays a stronger and more significant role in the preservation of their frustules after their deposition in death assemblages. Moreover, examination of the multivariate distances among sites showed a significant correlation between taphonomic and compositional distances (based on Manhattan and Bray-Curtis distance metrics, respectively), demonstrating the existence of common causal environmental stressors in the structuration of both patterns. Two causes were suggested to explain this relationship: (1) inter-specific variation in preservation, related to intrinsic properties of frustules, that can cause differential preservation and the consequent changes in species composition along environmental gradients; and (2) the same environmental factors are affecting both the distribution and preservation of the taxa, i.e. by altering the taphonomic signature of frustules and also shifting the species composition of the living communities according to their optima and tolerances.
Whatever the case, these results highlight the potentiality of combining diatom taphonomy and community paleoecology as a promising tool for paleoenvironmental reconstruction. If diatom taphonomic signatures can be used to trace past changes in key environmental variables (such as salinity), then the potential applicability of preservational profiles to the reconstruction of past environmental fluctuations increases significantly. In such a case, taphonomic information can be used to gain paleoenvironmental information, and not simply to assess the loss of information from the fossil record. This was the case of Pampean shallow lakes, where valve dissolution proved to be a more sensitive indicator of salinity differences among lakes than shifts in relative abundances of diatom taxa, enhancing the paleoenvironmental significance of diatom taphonomic signatures at long environmental gradients (Hassan and De Francesco 2018).
6.4.2 Local Scales: Within-Lake Taphofacies Comparisons
The scale of analysis of taphonomic signatures is highly variable, as variability in death assemblages structure and preservation is usually observed at different spatial scales, varying from local changes over centimeters to regional variation over kilometers or more (Staff and Powell 1990). The study of diatom taphofacies at regional scale demonstrated the potential use of this variability in addressing long environmental gradients. At local scales, variability can be promoted by subtle gradients in environmental or sedimentological parameters, but also by taphonomic factors acting during and after deposition (see Hassan et al. 2018 and references therein). In shallow lakes, the extensive development of the littoral zone relative to the pelagic zone provides a wide suite of available microhabitats that promote the growth of periphytic, benthic and pelagic communities (Wetzel 2001), being highly probable for these within-lake subenvironments to imprint distinctive taphonomic signatures on diatom valves. In such cases, recognizing within-lake taphofacies would become a useful tool for identifying subtle environmental differences from fossil diatoms in shallow lakes.
Accordingly, Hassan et al. (2018) conducted a study intending to determine whether diatom taphofacies are useful to discriminate subtle within-lake environmental differences in five sub-environments (free-floating and attached macrophytes, water column, open waters, and littoral bottom sediments) from Nahuel Rucá lake. In that study, compositional (relative abundances, diversity, richness, evenness and relative abundances) and taphonomic (dissolution and fragmentation) variables were obtained from a set of 129 samples (48 of surface sediments, 24 of water and 57 of macrophytes) covering both seasonal and spatial variability in diatom composition. The studied sub-environments differed in depth, macrophyte coverage, and water chemistry, being these differences reflected both by compositional and taphonomic data (Fig. 6.4).

Summary of the taphonomic processes taking place in the different within-lake sub-environments of Nahuel Rucá lake. Reproduced from Hassan et al. (2018), with permission
Assemblages from littoral, open-waters, and water column were dominated by the tychoplanktonic species C. meneghiniana, A. granulata and A. granulata var. angustissima, and differed from epiphytic assemblages, which were dominated by the epiphytic taxa Cocconeis placentula and Lemnicola hungarica. Moreover, diversity and richness were higher in planktonic and sedimentary assemblages, whereas dominance showed higher values in epiphytic samples. This difference was also reflected by taphonomic variables: epiphytic assemblages showed the best preservation, evidenced both by the lowest fragmentation and dissolution indices (Fig. 6.4). This good preservation is related to the conditions of their habitat, as they are protected by the epiphytic biofilm structure and the sheltered areas among macrophytes (Rojas and Hassan 2017), and therefore tend to be less subjected to the taphonomic biases generated by physical and chemical environmental factors than planktonic and sedimentary assemblages. Benthic and planktonic sub-environments, on the other hand, constitute less stable habitats in which sediment reworking pose physical constraints to diatom preservation. Although the compositional analysis prevented the discrimination among planktonic, littoral and open waters sub-environments, the taphonomic analysis was useful to detect preservational differences among them (Fig. 6.4).
Diatom breakage was the main taphonomic attribute identified in Nahuel Rucá assemblages, being marks of dissolution less frequent (Fig. 6.4). As Nahuel Rucá is a very shallow freshwater lake, the prevalence of fragmentation over dissolution was expected and can be advocated as the probable cause of the low live-dead fidelity observed in this lake (Hassan 2015). Fragmentation marks were more frequent in littoral sub-environments. Given the very shallow depths characteristic of this environment (less than 60 cm), frustule breakage is probably enhanced by sediment reworking (Flower and Nicholson 1987). Moreover, as the densely-vegetated areas of the littoral support abundant populations of macroinvertebrates (Tietze and De Francesco 2017), the highest fragmentation found in the littoral sediments can be also explained by biological action through grazing activity. The higher dissolution, on the other hand, was found in open-waters and planktonic habitats, and was related to the within-lake gradients in water chemistry characterizing the lake (Cristini et al. 2017), while in the littoral zone, characterized by lower pH and carbonate concentrations, diatom dissolution was prevented and preservation favored (Barker et al. 1994; Hassan and De Francesco 2018). Open waters and water column, on the other hand, were characterized by higher pH and carbonate/bicarbonate concentrations (Cristini et al. 2017), which could explain the higher frequency of dissolution marks exhibited by diatom assemblages in these sub-environments. Therefore, despite being low, dissolution was significantly higher in open waters and plankton, being probably enhanced by the continuous reworking and mixing exerted by wind action in the lake.
Overall, diatom taphofacies analysis demonstrated to be useful to capture environmental differences also at local scales, being taphonomic signatures related to within-lake differences in depth, macrophyte coverage, and water composition. Under such circumstances, combining taphonomic and compositional analyses would allow to uncover subtle within-lake environmental gradients not identifiable when only compositional analyses are performed. In this case, compositional analyses would allow to discriminate among different microhabitats, leading to paleoenvironmental inferences that would not be possible from taphonomic analyses only.
6.5 Experimental Taphonomy
The patterns of preservation of diatom assemblages observed at both local and regional scales clearly signaled the significant impact that environmental variables exert over the dissolution and breakage of the frustules. Particularly, the influence of salinity and pH can be relevant in Pampean shallow lakes, where the precipitation/evaporation balance plays a significant role in controlling water depth and chemistry. Changes in the hydrological cycle caused by climatic conditions or by anthropogenic activities can cause significant fluctuations in the pH and brine concentration of waters, which can be very alkaline and range from oligo to hypersaline (Fernández Cirelli and Miretzky 2004). Previous hydrochemistry studies in the Southern Pampas indicated that NaCl and HCO3Na are the dominant salts in surface and interstitial waters (Fernández Cirelli and Miretzky 2004; Hassan et al. 2012; Cristini et al. 2017). Hence, diatom silica dissolution caused by high concentrations of these salts, as well as elevated pH, are among the most probable processes explaining the observed patterns of preservation in the Pampean Region. In order to test this hypothesis, a series of short-term experiments on the dissolution of silica from cell walls under controlled laboratory conditions were recently conducted (Diaz and Hassan 2016, 2017).
Experiments were run on artificial assemblages constructed by mixing samples from different subenvironments (epiphytes, plankton, and sediments) from Nahuel Rucá lake. Experimental sets were designed to test for the effect of three concentrations of the two salts, NaCl (0.6, 1.2 and 3 M) and HCO3Na (0.6, 0.9 and 1.2 M), and two pH values (7 and 10) on diatom dissolution. An aliquot of each experimental assemblage (M1, M2, and M3) was thoroughly mixed with each solution to a final volume of 100 ml and stored polyethylene flasks. To avoid biological contamination a drop of mercury (ll) chloride (HgCl2) was added to each flask (Ryves 1994). Each one of the treatments and controls was made by triplicate and kept in an immersion bath under a controlled temperature of 20 °C. Aliquots of the experimental solutions were removed once each 5 days during 20 days and analyzed for changes in dissolved silica concentration (SiDi), relative and absolute abundances of diatoms, and dissolution indices based on the target taxon Cyclotella meneghiniana . All the experimental solutions increased significantly the rate of dissolution of silica from diatom walls, particularly since day 10. The two salts, however, differed in their relative effect, being the final SiDi concentrations obtained with HCO3Na solutions almost twice than those obtained with NaCl (Diaz and Hassan 2016). The effect of high pH was also evident: whereas the pH 7 solution did not produce increased dissolution, the difference in SiDi concentrations between initial and final experimental times was significant at pH 10 (Diaz and Hassan 2017). These increased SiDi values were accompanied by significant changes in the dissolution indices calculated from C. meneghiniana valves, which reached maximum values at pH 10, whereas no significant changes in relative or absolute abundances of diatoms were registered during the experiments. Evidences of dissolution were recorded as increases in the perimeter of individual areolae and decreases in the distances among the conjoining of neighboring areolae, as were observed in many taxa (Fig. 6.5). This effect of early stages of the dissolution process was also signaled by previous articles, both in marine and continental settings (Ryves et al. 2001; Warnock and Scherer 2015), in some cases accompanied by a lack of delicate structures and striae enlargement (Mikkelsen 1977; Ryves et al. 2001).

Scanning Electron Microscope micrographs showing pristine (upper row) and dissolved (lower row) valves of diatom taxa representative of different genera. From left to right: Surirella ovalis, Rhopalodia gibberula, Lemnicola hungarica, Cocconeis placentula and Cyclotella meneghiniana
Overall, these experimental results highlighted the impact that water chemistry exerts on diatom dissolution in Pampean shallow lakes. Given the naturally high pH, NaCl and HCO3Na concentrations characteristic of many of these lakes, these experimental findings can be confidently extrapolated to the interpretation of the dissolution trends found in modern surface sediments. Even as no significant changes in relative or absolute abundances were detected, this can be a consequence of the short exposure times covered by the experiments, instead of a definitive result. Previous studies over longer experimental times found significant changes in diatom abundances (Barker et al. 1994; Ryves et al. 2001), although the results were dependent of temperature and pre-treatments (Barker 1992; Flower 1993). Hence, although this is still pending of being experimentally tested, it is probable that, during longer periods of time, exposure of valves to these salts and increased pHs could bias the composition of Pampean diatom assemblages towards robust taxa.
6.6 Implications for Paleoenvironmental Reconstructions
The strong relationship between diatom preservation and environmental variables observed in field-based and experimental studies enhanced the paleoenvironmental significance of preservation trends along the fossil record of diatoms in Pampean lakes. In the light of these new results, Hassan and De Francesco (2018) reexamined the previously published taphonomic history of C. meneghiniana in Holocene sediments of Nahuel Rucá and San Leoncio (Hassan et al. 2014), suggesting strong fluctuations on past salinity and hardness in these lakes. Increased dissolution values exhibited by C. meneghiniana indicate the occurrence of brackish environments in these lakes by ca. 1500 cal. years BP, being modern conditions developed during the last ca. 200 cal. years BP. (Hassan et al. 2014). Dissolution values for these levels were high, indicating the prevalence of highly destructive environments, suggesting salinity and hardness values lying on the upper reaches of the studied modern gradient. Accordingly, community composition corresponding to these moments of increased salinity should be interpreted with caution, as the inferred destructive conditions could have biased significantly the proportions of taxa towards resistant species. As suggested by Hassan and De Francesco (2018), dissolution data for those levels would be a more confident source of environmental data than community composition, supporting the importance of studying taphonomic histories in these lakes.
In a more detailed study, Hassan et al. (2018) compared the within-lake composition and preservation of modern and Holocene (last ca. 700 years BP) diatom assemblages from Nahuel Rucá lake through multivariate analyses. Comparison of taphonomic indices between contemporary and fossil assemblages allowed to identify modern analogs for past diatom preservation, as several fossil samples showed high similarities to within-lake modern assemblages when dissolution and fragmentation were considered. Among them, samples from the topmost zone of the core overlapped with modern planktonic and littoral samples, being the best preserved of the whole sequence. Samples from the middle of the sequence overlapped with open waters modern samples, as they showed intermediate dissolution and fragmentation values. A number of fossil levels, however, showed dissolution and fragmentation values higher than those recorded in modern assemblages of the lake, lacking analogs in the dataset. These poorly preserved fossil samples were comparable to diatom assemblages from three previously studied Pampean brackish lakes with average salinities between 4 and 13 ppt (Hassan and De Francesco, 2018), which proved to be good analogues of past taphonomic conditions for those levels with no analogues in the Nahuel Rucá within-lake dataset, suggesting the prevalence of brackish conditions towards ca. 700 years BP.
These brackish conditions, however, were only partially suggested when assemblages composition alone was considered, as the dominant taxa were mostly euryhaline species adapted to a wide range of salinity conditions (e.g., C. meneghiniana, C. placentula, N. amphibia) with the consequently reduced value for salinity bioindication (Hassan et al. 2011). In fact, only the presence of rare taxa (<1%), such as Campylodiscus clypeus , Craticula halophila , Navicula salinicola , Nitzschia vitrea , and Synedra platensis , suggested brackish conditions. Hence, as the high salinity conditions suggested by high dissolution values were coincident with the presence of halophilous taxa, the combination of both independent sources of evidence provided reliable data to propose higher salinities in the lake at ca. 700 cal. years BP (Hassan et al. 2018). On the whole, the joint analysis of compositional and taphonomic variations in the recent sedimentary record of Nahuel Rucá demonstrated the usefulness of taphonomic analyses to uncover subtle paleoenvironmental variations, which could be overlooked if only traditional compositional analyses are performed. Accordingly, both taphonomic analyses and a holistic consideration of ecologically meaningful taxa should be considered in order to improve paleoenvironmental reconstructions in environmentally heterogeneous shallow lakes.
6.7 Conclusions and Future Directions
The results reviewed in the present chapter demonstrate the significant role that taphonomic processes play in the structuration of sedimentary diatom assemblages. Overall, these factors can alter the structure and taphonomic signatures of assemblages, leading to biased abundances, lost of delicate taxa and low compositional fidelity. However, these studies also demonstrated the positive contribution that taphonomic signatures can provide to paleoenvironmental studies: as taphofacies were successfully related to local and regional environmental gradients, their potential as paleoenvironmental proxies arise as a useful independent tool of information. Moreover, the experimental demonstration of the role of water chemistry on diatom dissolution also supported the field observations, leading to an empirical demonstration of the processes structuring the patterns of preservation observed.
Accordingly, future directions in this project should tend to provide a more comprehensive picture of unknown aspects of diatom taphonomy. Particularly, long-term experiments on dissolution and fragmentation need to be conducted, in order to clarify the role of both water chemistry and physical factors in structuring the composition of assemblages in natural conditions. Previous studies have demonstrated that diatom preservation is linked to the particular characteristics of the frustule architecture (e.g. row pore frequency, width/length relation, size, the degree of silicification, etc.), being consequently strongly dependent on taxonomy (Haberyan 1985; Barker 1992; Ryves et al. 2009). These particular characteristics have led to the development of rankings of diatom taxa according to their preservation potential in other areas of the world, which proven to be useful in paleoenvironmental reconstructions (Ryves et al. 2009). As this data is still unavailable for Pampean diatom taxa, future research should focus on that direction, as this information would provide useful tools for the environmental interpretation of past diatom assemblages in Pampean shallow lakes. Moreover, experimental designs including fragmentation caused by sediment reworking and bioturbation are particularly necessary, as a way to elucidate the significance of the high fragmentation indices found in these shallow lakes. As the causes of frustule fragmentation have received little attention worldwide, this line of research constitutes an exciting future development that requires further attention. Finally, including new studies on diatom taphofacies that allow to expand the regional and local environmental gradients already studied would increase the applicability of these studies to paleoenvironmental research, allowing the reconstruction of a wider set of paleoenvironmental conditions.
Up to now, the usefulness of these actualistic taphonomic data become evident when applied to the reconstruction of the recent history of the freshwater Nahuel Rucá lake, as it allowed to identify environmental fluctuations not so clearly reflected by the composition of assemblages. Hereafter, if fossil studies are accompanied by local and regional analyses on actualistic taphonomy, the potential information provided by preservational profiles increases significantly. Hence, we conclude that increasing actualistic and experimental data should be a priority in future studies, as taphonomic information can be used to gain paleoecological information, and not simply to assess the loss of information from the fossil record.
References
Baars JWM (1981) Autecological investigations on marine diatoms. 2. Generation times of 50 species. Hydrobiol Bull 15:137–151
Barker P (1992) Differential diatom dissolution in Late Quaternary sediments from Lake Manyara, Tanzania: an experimental approach. J Paleolimnol 7:235–251
Barker P, Fontes JC, Gasse F, Druart J-C (1994) Experimental dissolution of diatom silica in concentrated salt solutions and implications for paleoenvironmental reconstruction. Limnol Oceanogr 39:99–110
Battarbee RW (1986) Diatom analysis. In: Berglund BE (ed) Handbook of Holocene Palaeoecology and Palaeohydrology. Wiley, NewYork, pp 527–570
Battarbee R, Jones V, Flower R, Cameron N, Bennion H, Carvalho L, Juggins S (2001) Diatoms. In: Smol J, Birks HJ, Last W, Bradley R, Alverson K (eds) Tracking environmental change using lake sediments. Springer, Netherlands, pp 155–202
Clarke KR, Warwick RM (1998) Quantifying structural redundancy in ecological communities. Oecologia 113:278–289
Cristini PA, Tietze E, De Francesco CG, Martínez DE (2017) Water geochemistry of shallow lakes from the southeastern Pampa plain, Argentina and their implications on mollusk shells preservation. Sci Total Environ 603:155–166
Diaz MC, Hassan GS (2016) Efecto de diferentes concentraciones de sales sobre la preservación de diatomeas: una aproximación experimental. 11° Congreso de la Asociación Paleontológica Argentina (General Roca), Resúmenes
Diaz MC, Hassan GS (2017) Evaluación experimental de la preservación de diatomeas en lagos someros pampeanos: efecto del pH e implicancias paleoambientales. Reunión de Comunicaciones de la Asociación Paleontológica Argentina, San Luis, 23–25 de noviembre de 2017, Resúmenes
Dodd JR, Stanton RJ (1990) Paleoecology. Concepts and applications., Second Edition edn. Wiley-Interscience Publication, New York, p 502
Fernández Cirelli A, Miretzky P (2004) Ionic relations: a tool for studying hydrogeochemical processes in Pampean shallow lakes (Buenos Aires, Argentina). Quat Intern 114:113–121
Flower R (1993) Diatom preservation: experiments and observations on dissolution and breakage in modern and fossil material. Hydrobiologia 269–270:473–484
Flower RJ, Nicholson AJ (1987) Relationships between bathymetry, water quality and diatoms in some Hebridean lochs. Fresh Biol 18:71–85
Flower RJ, Ryves DB (2009) Diatom preservation: differential preservation of sedimentary diatoms in two saline lakes. Acta Bot Croat 68:381–399
Fritz SC (2007) Salinity reconstructions from continental lakes. In: Elias SA (ed) Encyclopedia of quaternary science. Elsevier, pp 62–71
García-Rodríguez F, Piovano E, Puerto Ld, Inda H, Stutz S, Bracco R, Panario D, Córdoba F, Sylvestre F, Ariztegui D (2009) South American lake paleo-records across the Pampean region. PAGES News 17:115–118
Haberyan K (1985) The role of copepod fecal pellets in the deposition of diatoms in Lake Tanganyika. Limnol Oceanogr 30:1010–1023
Hassan GS (2013) Diatom-based reconstruction of Middle to Late Holocene paleoenvironments in Lake Lonkoy, southern Pampas, Argentina. Diat Res 28:473–486
Hassan GS (2015) On the benefits of being redundant: Low compositional fidelity of diatom death assemblages does not hamper the preservation of environmental gradients in shallow lakes. Paleobiology 41:154–173
Hassan GS (2018) Within vs. between-lake variability of sedimentary diatoms: the role of sampling effort in capturing assemblage composition in environmentally heterogeneous shallow lakes. J Paleolim. https://doi.org/10.1007/s10933-018-0038-8
Hassan GS, De Francesco CG (2018) preservation of Cyclotella meneghiniana Kützing (Bacillariophyceae) along a continental salinity gradient: implications for diatom-based paleoenvironmental reconstructions. Ameghiniana 55:263–276
Hassan GS, Espinosa MA, Isla FI (2008) Fidelity of dead diatom assemblages in estuarine sediments: how much environmental information is preserved? Palaios 23:112–120
Hassan G, Tietze E, De Francesco C (2009) Modern diatom assemblages in surface sediments from shallow lakes and streams in southern Pampas (Argentina). Aquatic Sci 71:487–499
Hassan GS, Tietze E, De Francesco CG, Cristini PA (2011) Problems and potentialities of using diatoms as paleoclimatic indicators in Central Argentina. In: Compton JC (ed) Diatoms: ecology and life cycle. Nova Science Publishers, New York, pp 185–216
Hassan GS, De Francesco CG, Peretti V (2012) Distribution of diatoms and mollusks in shallow lakes from the semiarid Pampa region, Argentina: their relative paleoenvironmental significance. J Arid Env 78:65–72
Hassan GS, Tietze E, Cristini PA, De Francesco CG (2014) Differential preservation of freshwater diatoms and mollusks in late Holocene sediments: paleoenvironmental implications. Palaios 29:612–623
Hassan GS, Rojas LA, De Francesco CG (2018) Incorporating taphonomy into community-based paleoenvironmental reconstructions: can diatom preservation discriminate between shallow lake sub-environments? Palaios 33:376–392
Julius ML, Theriot EC (2010) The diatoms: a primer. In: Smol JP, Stoermer EF (eds) The diatoms: applications for the environmental and earth sciences. Cambridge University Press, Cambridge, pp 8–22
Kidwell SM (2002) Time-averaged molluscan death assemblages: palimpsests of richness, snapshots of abundance. Geology 30:803–806
Kidwell SM (2013) Time-averaging and fidelity of modern death assemblages: building a taphonomic foundation for conservation palaeobiology. Palaeontology 56:487–522
Lewin JC (1961) The dissolution of silica from diatom walls. Geochim Cosm Acta 21:182–198
Mikkelsen N (1977) Silica dissolution and overgrowth of fossil diatoms. Micropaleontology 23:223–226
Parsons KM, Brett CE (1991) Taphonomic processes and biases in modern marine environments: an actualistic perspective on fossil assemblage preservation. In Donovan SK (ed) The Processes of Fossilization: Columbia University Press, New York, pp 22–65
Rojas LA, Hassan GS (2017) Distribution of epiphytic diatoms on five macrophytes from a Pampean shallow lake: host-specificity and implications for paleoenvironmental reconstructions. Diat Res 32:263–275
Round FE (1964) The diatom sequence in lake deposits, some problems of interpretation: Verhandlungen Internationale Vereinigung für Theoretische und angewandte Limnologie, vol 15, pp 1012–1020
Ryves DB (1994) Diatom dissolution in saline lake sediments: an experimental study in the Great Plains of North America. Ph.D. dissertation, Univ. College London, 307 p.
Ryves DB, Juggins S, Fritz SC, Battarbee RW (2001) Experimental diatom dissolution and the quantification of microfossil preservation in sediments. Palaeogeogr Palaeocl Palaeoecol 172:99–113
Ryves DB, Battarbee R, Juggins S, Fritz SC, Anderson NJ (2006) Physical and chemical predictors of diatom dissolution in freshwater and saline lake sediments in North America and West Greenland. Limnol Oceanogr 51:1355–1368
Ryves DB, Battarbee RW, Fritz SC (2009) The dilemma of disappearing diatoms: Incorporating diatom dissolution data into palaeoenvironmental modelling and reconstruction. Quat Sci Rev 28:120–136
Smol JP, Stoermer EF (2010) The diatoms: applications for the environmental and earth sciences. Cambridge University Press, Cambridge, p 667
Speyer SE, Brett CE (1986) Trilobite taphonomy and Middle Devonian Taphofacies: palaios, vol 1, pp 312–327, https://doi.org/10.2307/3514694
Staff GM, Powell EN (1990) Local variability of taphonomic attributes in a parautochthonous assemblage: can taphonomic signature distinguish a heterogeneous environment? J Paleontol 64:648–658
Stutz SM, Borel M, Fontana SL, Del Puerto L, Inda H, García-Rodríguez F, Tonello MS (2010) Late Holocene climate and environment of the SE Pampa grasslands, Argentina, inferred from biological indicators in shallow, freshwater Lake Nahuel Rucá. J Paleolim 44:761–775
ter Braak CJF (1994) Canonical community ordination. Part I: Basic theory and linear methods. Écoscience 1:127–140
Tietze E, De Francesco CG (2017) compositional fidelity and taphonomy of freshwater mollusks from three pampean shallow lakes of Argentina. Ameghiniana 54:208–223
Tomasových A, Kidwell SM (2009a) Fidelity of variation in species composition and diversity partitioning by death assemblages: time-averaging transfers diversity from beta to alpha levels. Paleobiology 35:94–118
Tomasových A, Kidwell SM (2009b) Preservation of spatial and environmental gradients by death assemblages. Paleobiology 35:119–145
Tomasových A, Kidwell SM (2011) Accounting for the effects of biological variability and temporal autocorrelation in assessing the preservation of species abundance. Paleobiology 37:332–354
Warnock JP, Scherer RP (2015) Diatom species abundance and morphologically-based dissolution proxies in coastal Southern Ocean assemblages. Cont Shelf Res 102:1–8
Wetzel R (2001) Limnology: lake and river ecosystems. Academic Press, San Diego, 1006 p
Acknowledgements
Financial support for this research was granted by the Agencia Nacional de Promoción Científica y Tecnológica (PICT 2727/2013), the Consejo Nacional de Investigaciones Científicas y Técnicas (PIP 0063/2010), and the Universidad Nacional de Mar del Plata (EXA 775/16). Special thanks to Pedro Urrutia (Estancia Nahuel Rucá), Santiago González Aguilar (Las Mostazas), Nicolás Bustamante (Camping Ruca Lauquen), and Héctor Sanabria (Los Carpinchos), for permission to access private land where the lakes are located. We also thank Gustava Bernava for performing the chemical analyses. D. Navarro, S. Stutz and M. Tonello helped at some stage of the research. Experimental studies are part of the Doctoral Thesis of M.C. Díaz at the University of Mar del Plata. G. Hassan and C.G. De Francesco are members of the Scientific Research Career of the Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Hassan, G.S., De Francesco, C.G., Díaz, M.C. (2020). Actualistic Taphonomy of Freshwater Diatoms: Implications for the Interpretation of the Holocene Record in Pampean Shallow Lakes. In: Martínez, S., Rojas, A., Cabrera, F. (eds) Actualistic Taphonomy in South America. Topics in Geobiology, vol 48. Springer, Cham. https://doi.org/10.1007/978-3-030-20625-3_6
Download citation
DOI: https://doi.org/10.1007/978-3-030-20625-3_6
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-20624-6
Online ISBN: 978-3-030-20625-3
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)