
Abstract
This book chapter contributes to identifying some core areas inside the oil industry as potential targets for biotechnology, motivated by the increasing global demand of fuels in addition to the reduction of conventional crude oil reserves that have produced a greater interest on the exploitation of unconventional crude reserves. In parallel with enlarged global environmental concerns, it is mandatory the developing and improving clean-alternative fuel technologies with enhanced bioremediation strategies for unconventional crude. These efforts include the application of petroleum biotechnology with promissory microorganisms, especially extremophilic hydrocarbonoclastic fungi, a broad group of cultivable fungi which live optimally under extreme conditions and are characterized by having a high ability to grow using hydrocarbons as sole carbon source and energy. Few publications are focused on petroleum biotechnology and applications of fungal degradation or bioconversion of extra-heavy crude oil (EHCO), a type of crude that contains elevated amounts of asphaltenes, high-molecular-weight compounds with low bioavailability, and limited susceptibility to being biotransformed. We have included an enriching discussion on the biotechnological strategies applied to the study of cultivable fungal biodiversity inhabiting extreme environments to obtain powerful biocatalysts, following a simple and fast screening to determine both their hydrocarbonoclastic abilities and tolerance of growing in the presence of high concentrations of EHCO and hydrocarbons polycyclic aromatics compounds (HPAs). The potential applications of these promissory extremophilic hydrocarbonoclastic fungal strains in mycoremediation and EHCO-bioupgrading processes to promote the sustainable development of the petroleum industry will be discussed.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
In spite of the laudable financial, technical, and scientific efforts focused on the development of new environmental friendly energy sources, it is unquestionable that the world is running on petroleum. In fact, the present world oil requirements to sustain the actual growth behavior of the global population are estimated in more than 95 million barrels per day (OPEC, Annual Statistical Bulletin 2017).
The increasing global demand for fuel and the reduction of conventional crude reserves have generated a great interest on the exploitation of unconventional crude reserves worldwide. These unconventional sources of fossil fuel cannot be extracted, transported, and refined by conventional methods. The certified crude oil reserves worldwide are estimated in more than 1.5 billion barrels, of which 70% are constituted by unconventional crudes, such as the Orinoco Oil Belt (OOB ), the biggest certified reserve of EHCO worldwide located in Venezuela with over 300 billion barrels (OPEC, Annual Statistical Bulletin 2017).
The enormous reserves of unconventional crudes contain high concentrations of toxicity pollutants and high-molecular-weight compounds, such as asphaltenes, which are heterogeneous and complex mixtures insoluble in n-heptane or n-pentane, and soluble in benzene or toluene, which contain heteroatoms (nitrogen, sulfur, and oxygen) and heavy metals such as nickel and vanadium in their structure (Waldo et al. 1991; Strauz et al. 1992; Uribe-Álvarez et al. 2011; Ayala et al. 2012; Naranjo et al. 2007; León et al. 2007).
Unconventional crude oils have been characterized as recalcitrant (very low availability and degradation by microorganisms), polar, and water insoluble, and contain sulfur and/or heavy metals in association; hence there is a high demand for the development of technologies that aim to alleviate environmental impacts during the extraction, production, and refinement of unconventional crude oils.
The sustainable development of the oil industry requires the development of novel environmental friendly technologies, which could offer both higher economic income with environmental remediation of anthropogenic intervened ecosystems and conservation of natural ecosystems. Complementary use of modern biotechnology in the oil industry provides new tools to improve their processes and products in the production chain, decreasing operational costs and increasing productive capabilities with minimum environmental impact (Naranjo et al. 2007, 2008, 2013).
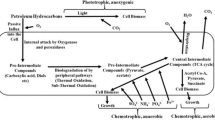
Aromatic hydrocarbons ranging from the single benzene ring to the high-molecular-weight polycyclics are generally biodegraded via one or more of the three independent enzymatic systems. The intracellular P450 monooxygenases that detoxify harmful chemicals are universally present in the microsomes of eukaryotic cells, while lignin-degrading fungi specifically produce extracellular peroxidases and laccases that biodegrade aromatic hydrocarbons (Prenafeta-Boldú et al. 2018). The low functional specificity and high redox potential of peroxidases and laccases enable the oxidation of a broad range of aromatic hydrocarbons and other recalcitrant contaminants (Prenafeta-Boldú et al. 2018).
In this sense, the study and application of the powerful extracellular oxidative lignin-degrading enzyme system (LDS ) secreted by fungi have a great potential as biocatalysts (as whole cells or enzymatic catalyst) in mycoremediation and EHCO bioupgrading processes, through three main pathways: (1) de-aromatization of high-molecular-complexity compounds into more soluble compounds with concomitant reduction of viscosity and enhanced bioavailability by microorganisms; (2) biodesulfurization of sulfur heteroatoms; and (3) de-metallization of heavy metals such as nickel (Ni) and vanadium (V), among others.
Here, we want to contribute and promote the sustainable development of the petroleum industry with the complementary use of the biotechnology of fungi with respect to their ability to degrade and/or transform hydrocarbons, laying special emphasis on the fungal biodiversity associated to extreme environments and the selection of promissory extremophilic and hydrocarbonoclastic fungi.
The term extremophile was first proposed by MacElroy in 1974 to describe a broad group of organisms which lived optimally under extreme conditions (MacElroy 1974; Zhang et al. 2018). Their taxonomic range has been expanded from prokaryotes to all three domains Archaea, Bacteria, and Eukarya (Zhang et al. 2018). An extreme environment is a place that contains conditions that are hard to survive in for most known life forms. These conditions may be extremely high or low temperature (extremely hot or cold), high concentration of a salt (hypersaline), high acidity or alkalinity (acidic or alkaline), desiccation (without water), extremely high pressure (under pressure), high or low content of oxygen or carbon dioxide in the atmosphere (with or without O2 or CO2), high levels of radiation (UV emission, radioactivity), and places anthropogenically impacted, such as soil, sediment, or water contaminated by petroleum or other toxic and contaminant substances (altered by humans).
According to different extreme habitats, extremophiles are classified into seven categories (Arulazhagan et al. 2017; Zhang et al. 2018). Organisms whose optimal growth temperature ranges from 50 to 80 °C or exceeds 80 °C are called thermophiles or hyperthermophiles, respectively. Psychrophiles are organisms that grow at low temperatures ranging from 0 to 15 °C. Halophiles require >3% of NaCl to grow and are classified as halotolerant or slight halophiles (2–5% NaCl), moderate halophiles (5–20% NaCl), and extreme halophiles (20–35% NaCl). Acidophiles or alkaliphiles show optimal growth at pH values 1–5 and pH >9, respectively. Piezophiles or barophiles reside under high hydrostatic pressure which have been isolated from the deep-sea sediments (>3000 m depth and pressures of up to 110 MPa). Finally, xerophiles are organisms that grow under low water content (aw 0.60–0.90). In addition, these organisms are normally polyextremophiles and are adapted to live in habitats where various physicochemical parameters reach extreme values (Rampelotto 2013).
In extreme habitats, microorganisms require a large adaptation process until reaching optimal growth and reproduction. This evolutional redesign involves novel morphophysiological characteristics and modifications of genes and proteins, with subsequent changes in regulatory and metabolic pathways until epigenetic modifications, which have a great interest for biotechnological purposes. In fact, Zhang et al. (2018) reported a total of 314 new bioactive fungal natural products from 56 Ascomycota extremophilic fungi (asexual stage), including terpenoids/steroids, alkaloids/peptides/amides, quinones/phenols, esters/lactones, xanthones, and polyketides. Likewise, this is particularly true for their enzymes, which remain catalytically active under extremes of temperature, salinity, pH, and solvent conditions. Interestingly, some of these enzymes display polyextremophilicity (i.e., stability and activity in more than one extreme condition) making them widely functional in industrial biotechnology (Rampelotto 2013).
The exploitation, production, refining, and transportation of oil and its derivatives occasionally lead to technical and operational accidents with serious harm to the environment, some with irreversible destruction (León et al. 2009; Pernía et al. 2012, 2018). Polycyclic aromatic hydrocarbons are generated from both natural and anthropogenic processes, and are ubiquitous environmental pollutants with cytotoxicity, mutagenicity, and carcinogenicity capabilities. Due to their hydrophobic nature, they persist in the environments. More than two decades ago, the United States Environmental Protection Agency (USEPA ) considered that some PAHs are toxic and possibly human carcinogens (Nadon et al. 1995).
There is wide fungal biodiversity with diverse enzymatic mechanisms that transform different hydrocarbon chemical structures, from short-chain aliphatics to heavy-weight polycyclic aromatics (Prenafeta-Boldú et al. 2018). The hydrocarbonoclastic fungi are a fascinating group of microorganisms with the unique ability to metabolize hydrocarbons as a sole source of carbon and energy, despite their low biodegradability due to their littler solubility and high hydrophobicity that limit their transport into microbial cells (Arulazhagan et al. 2017).
The term extremophilic hydrocarbonoclastic fungi is proposed here to describe a large and heterogeneous group of cultivable fungi which live optimally under extreme conditions, as well as are characterized by having a high ability to grow using hydrocarbons as the sole carbon source and energy. Usually, these fungi are isolated from soils, sediments, fluids, vapor, or water impregnated by petroleum or its derivatives. These extreme environments are mainly the consequence of anthropogenic activities, and usually have hard conditions to survive in for most known forms of life, such as high concentration of salt (hypersaline); high acidity or alkalinity (acidic or alkaline); high concentrations of high-molecular-weight compounds and toxicity pollutants (i.e., asphaltenes); heavy metals and heteroatoms such as sulfur; and high levels of PAHs with high toxicity. Thus, extremophilic hydrocarbonoclastic fungi are normally polyextremophiles, adapted to live in habitats under pressure of various physicochemical conditions considered “extreme.”
Besides their polyextremophiles characteristics, these fungi have the unique ability to use hydrocarbons as a sole source of carbon and energy, despite their high toxicity and hydrophobicity properties; hence extremophilic hydrocarbonoclastic fungi play a pivotal role in the degradation/transformation of petroleum and its derivatives. This is the reason why the use of extremophilic hydrocarbonoclastic fungi as biocatalysts at great scale requires the maintenance and vegetative reproduction in the laboratory and industrial levels in the absence of a sexual stage. The asexual reproduction by mitotic division in fungi is commonly used to produce fungal mycelium to colonize the environments, which generates new identical individuals by remaining haploid, resulting in a progeny with the same genetic information as its own parental inoculum (Moore et al. 2011). Hence, the mitotic division guarantees researchers a simple, fast, and profuse vegetative reproduction of the fungal biocatalysts at the same time keeping their original hydrocarbonoclastic and polyextremophile characteristics.
The scope of this chapter is to demonstrate the application of the fungal biotechnology in the degradation and bioconversion of unconventional crude and its possible applications for the sustainable development of the petroleum industry. Contrary to other publications that are focused on bioremediation of light crude oils, here we present promising fungi with degradation potential of recalcitrant crudes like EHCO, a type of crude oil that is considered nonbiodegradable by microorganisms. At the same time, we demonstrate for the first time the potential of hydrocarbonoclastic fungi isolated from extreme environments (EHCO, hydrocarbon pits, crude distribution pipes, and the natural asphaltene Lake of Guanoco) to tolerate high concentration of EHCO, dibenzothiophene (DBT), phenanthrene, naphthalene, and pyrene in vitro conditions. We also described the relationship between lignin-degrading enzyme system (LDS) and the EHCO bioconversion by several extremophilic hydrocarbonoclastic fungi studied.
2 Biotechnology as the Measurement of Human Being and Biodiversity: Potential Applications for Sustainable Development of the Petroleum Industry
Since ancestral times, and based on its own essence, biotechnology started from the logic of human being, the observation, learning, and exploiting of nature to a benefit, pursuing a better quality life. This initiative was originally contemplative and through the centuries, based on success and error testing, formidably was ahead of its time and was evolving in the same way that knowledge evolved through the course of time. Thus, it could be considered that biotechnology occurs empirically from the activities of daily living of our ancestors, becoming implicit to the human being as a thinking and dynamic element of nature, which blooms before his eyes, building a magical altar of incommensurable bio-possibilities, helping to solve their most basic needs. Therefore, biotechnology dates from the same origin of human evolution as a thinking creature that expands human possibilities. Likewise, biotechnology could be considered as the oldest technology ever practiced by humans. In this process of singular complexity, the human being as subjects and from its knowledge obtains benefits from nature and its biodiversity, while, in the same act, nature as subject and not as an object is observed, acquired, and learned by humans. In this unique process, the bio-possibilities were greater and greater as the knowledge of the human kind was approaching to nature, and its exuberance biodiversity.
The harmonizing use of conventional oil production technologies with petroleum biotechnology emerges as an innovative strategy to help to ensure a sustainable development and the conservation of the environment. However, very little has been described concerning the medullary areas inside the petroleum industry where the biotechnology could have potential applications (Foght 2004). To identify medullary areas not only the products but also the operational processes associated to the petroleum industry in its whole-value chain should be known and the clear increase of the exploitation, production, and processing of unconventional crude oils should be considered. However, it is also important to understand the essence that fundamentally supports any biotechnological development: a specific problem or requirement to be solved, knowledge and technology, and biodiversity.
In principle, the petroleum biotechnology could be defined as the study and rational use of the biodiversity, its genetic, enzymatic, and metabolic resources, to give high-added value to the products and processes of the petroleum industry in its whole-value chain, and to contribute to mitigate the environmental impact of its operational activities. Major aims of the possible contribution of the complementary use of biotechnology in the petroleum industry would be focused on (1) the improvement of its productive capabilities (efficiency and productivity); (2) a comprehensive security of its operational processes (sustainability, safeguarded processes, and productivity); and (3) the remediation of its associated environmental liabilities (environmental sanitation, conservation, and sustainability).
Prenafeta-Boldú et al. (2018) commented that applied research on hydrocarbonoclastic fungi includes dedication to preventing biodeterioration as well as the potential application of fungal enzymatic capabilities for bioremediation purposes. Although there are numerous benefits and applications of hydrocarbonoclastic fungi, in this chapter only four main research and development areas are discussed (Fig. 28.1):
-
1.
Improvement of the physicochemical properties of unconventional crude oil and its derivatives using enzymatic biocatalysts, nanostructured support, and other coupled processes of bioconversion (bioupgrading)
-
2.
Improvement of the recovery factor of oil in reservoirs by using promissory extremophile microorganisms and biodegradable polymers (MEOR, microbial enhanced oil recovery ) (biorecovery)
-
3.
Bioremediation of soil and water contaminated with hydrocarbons, oil pits, drilling waste, and industrial pollutants (bioremediation)
-
4.
Contribution to the reduction of operating costs, ensuring the continuous operation of processes by the detection, monitoring, and control of microbial influenced corrosion, biofouling, and biodeterioration in oil facilities (biocontrol)

Some contributions of the complementary use of biotechnology in the petroleum industry
3 Fungal Biodiversity with Hydrocarbonoclastic Potential: From the Meta-analysis to the Tangible Reality Show
The work of Pernía et al. (2012) represents the first study where a meta-analysis was applied to the study of fungal diversity isolated from crude oil, its derivatives, and hydrocarbon-contaminated environments, including their hydrocarbonoclastic capabilities, through a detailed review of scientific literature published during the last century (from 1900 to 2012) in this passionate field of science. The results of this meta-analysis showed that the substrates with the highest fungal diversity were soils impacted with crude and natural asphalt, obtaining the lowest diversity in soil contaminated with diesel and gasoline in agreement with their high toxicity. The total fungal diversity isolated was constituted mainly by phylum Ascomycota (83%) followed by Zygomycota (10%), Basidiomycota (1%), and others (6%). In the phylum Ascomycota, the most common orders obtained were the Eurotiales (54%), followed by the Hypocreales (18%), Microascales (6%), and Saccharomycetales (5%), and the most predominant genera were Penicillium (18%), Aspergillus (17%), and Fusarium (6%).
Based on the percentage of degradation of total petroleum hydrocarbons (TPH), the group that presented greater degradation, TPH: 52 ± 3.53%, was formed by the genera Aspergillus, Bjerkandera, Coriolus, Emericella, Phanerochaete, Pleurotus, Rhizopus, and Trametes; with respect to degradation rate of the saturated and aromatic fractions, two groups were obtained by the genera: (1) Beauveria, Coriolopsis, Emericella, Fusarium, Phanerochaete, Pleurotus, and Trametes, (74.43 ± 3.40%), and (2) Coriolopsis, Fusarium, Pleurotus, and Trametes (97.75 ± 2.25 %). Likewise, evidence of degradation of resin and asphaltene fractions (10–28% and 10–40%, respectively) was found by the genera Aspergillus, Candida, Emericella, Eupenicillium, Fusarium, Graphium, Neosartorya, Paecilomyces, and Meyerozyma (Pernía et al. 2012).
3.1 Exploring Culturable Extremophilic Fungal Biodiversity with Potential Applications in the Sustainable Development of the Petroleum Industry: Knowing the Bio-possibilities
Fungi have essential roles in natural ecosystems through diverse forms of lifestyles, such as parasitism, mutualistic symbiotic associations with plant roots (e.g., arbuscular mycorrhizal fungi, ectomycorrhizal fungi, Orchid- and Ericoid-mycorrhizal fungi) and other organisms, as well as decomposers of the organic matter; hence fungi directly contribute to biogeochemical cycles (e.g., carbon, nitrogen, and phosphorous nutrients) and influence the greenhouse gas balance in the atmosphere on the global scale. In fact, the incorporation of mycorrhizae in global carbon cycle models is crucial in order to accurately predict ecosystem responses and feedbacks to climate change (Terrer et al. 2016).
The fungal decomposers can downgrade complex organic molecules such as cellulose, hemicellulose, lignin, pectin, starch, and non-synthesized xenobiotic compounds by natural metabolic processes, whose toxicity lies in their chemical nature and is persistent in the environment due to their low bioavailability (Dávila and Vázquez-Duhalt 2006, Naranjo et al. 2007, Pernía et al. 2018). The metabolism of organic compounds to less structurally complex products can directly or indirectly affect the growth of other surrounding microorganisms (Amund et al. 1987, Bartha and Atlas 1977, Brock 2015, Obire 1993, Odokuma and Opokwasili 1993, Pernía et al. 2018).
In the oil production value chain (processes of exploitation, production, refining, transportation of petroleum and its derivatives) there are occasional technical and operational accidents that release xenobiotic compounds into the environment promoting a selective pressure on the microbiota (Atlas and Bartha 1972; Calomiris et al. 1986; Pernía et al. 2018). The fungal catabolic activity (intracellular or extracellular) modifies bio-availability features of the xenobiotic compounds and derivatives, affecting the dynamics of autochthonous microbial communities (Coyne 2000). Subsequently, a selective enrichment process occurs for certain degraders and/or tolerant species, and the disappearance of those that do not have these capabilities (Benka-Coker and Ekundayo 1997).
In the study of the autochthonous fungal communities associated with oil-polluted environments it’s essential to describe their microbial and functional biodiversity. However, for the selection of powerful extremophilic fungi as biocatalysts, the studies of both hydrocarbonoclastic potential and their tolerance to xenobiotic and toxic compounds abilities are crucial. Despite the unquestionable importance of the microbial and functional diversity studies that include the cultivable and non-cultivable microorganisms by means of metagenomics tools, in this work our effort was focused on the analyses of the cultivable extremophilic fungi from the extreme environments due to their potential use as promising biocatalysts for the sustainable development of the petroleum industry.
In this way, our research team started an exhaustive study on the fungal diversity associated with different types of samples: (1) Carabobo EHCO (samples taken directly from an oil well), (2) oil-polluted soil adjacent to an oil pit, (3) the natural asphalt Lake of Guanoco, and (4) petroleum naphtha distribution system (Fig. 28.2). The tolerance to EHCO and polyaromatic hydrocarbon (PAH) compound models, such as naphthalene, phenanthrene, DBT, and pyrene, was also investigated at inter- and intraspecific species levels. The isolation, maintenance, growth, and molecular identification of the fungal strains were performed according to Naranjo et al. (2007, 2013). The descriptive analysis into the functional group of fungi was carried out according to Pernía et al. (2012).

Sampling sites and biotechnological strategy proposed. The samples were taken from (a) the natural asphalt lake of Guanaco, Sucre State, Ven; (b) the oil-discharge area of the oil pit DED-3-F1, Operational District San Tomé, Anzoátegui State, Ven; (c) the naphtha pipeline distribution system, San Diego Cabrutica, Anzoátegui State, Ven; (d) Carabobo EHCO from the CIB254 oil well, O16 Flow station, District of Morichal, Monagas State, Venezuela
3.2 Isolation of Cultivable Fungal Biodiversity from Extreme Environments: Identifying the Cultivable Fungal Communities
At first, the relative frequencies of the different phylum and orders of the fungal strains isolated from the extreme environments were determined as mentioned above. All sites or samples studied were considerate extreme environments because they showed various physicochemical conditions unfavorable for survival of most known life forms.
In the case of the natural samples of asphalt and EHCO, these contain a heterogeneous mixture of organic compounds with a high concentration of asphaltenes and toxicity pollutants such as heavy metals and sulfur. Asphaltene’s recalcitrancy is explained by the high degree of aromaticity combined with the presence of short alkyl chains (Naranjo et al. 2013). Carabobo EHCO assays have heavy American Petroleum Institute (API) gravity of 8.5°, sulfur concentration of 3.9%, and heavy metal Ni and V concentration of 480 mg/L (Pernía et al. 2018).
Oil-polluted soil from oil pits possesses a significant risk to wildlife and humans, due to its enormous mixture of organic compounds and chemical pollutants with high toxicity levels. During extraction and production processes in the Orinoco Oil Belt in Venezuela, oil pits generally store high volumes of sludge, oil spill, chemical treaters, formation water (brine), and drilling waste impregnated with EHCO and highly salinized water-based drilling fluids, which are mainly constituted by a wide range of corrosive compounds such as sodium bicarbonate, sodium carbonate, potassium chloride, potassium hydroxide, glycol, sodium hydroxide, thickeners, and lubricants, among others, xenobiotics that radically increase the alkalinity and salinity levels of the soil destroying its structure (Arellano et al. 2008, Naranjo et al. 2013).
Petroleum naphtha is a highly volatile and flammable intermediate hydrocarbon liquid stream derived from the refining of crude oil that contains paraffins, naphthenes (cyclic paraffins), and aromatic hydrocarbons. In this case, we refer to olefin-containing naphtha derived from the fluid catalytic cracking, visbreakers, and coking processes used in many refineries (named cracked naphtha) that is broadly used by the Venezuelan petroleum industry as a solvent that decreases the high viscosity levels of the EHCO from OOB. According to the National Institute for Occupational Safety and Health (NIOSH), petroleum naphtha is immediately dangerous to life and health (CDC-NIOSH. 2015).
The biotechnological strategy proposed for identification of fungal communities from extreme environments is indicated in Fig. 28.2. Results showed, in terms of biodiversity, the most diverse substrate was the natural asphalt Lake of Guanoco, where 11 different fungal genera were isolated (including one basidiomycete fungus), followed by the oil-polluted soil (7 genera), and EHCO and naphtha, where only 3 genera and 1 genus were isolated, respectively (Table 28.1).
As reported by Pernía et al. (2012), the most isolated fungal group was phylum Ascomycota (91.66%) and only 8.33% belong to phylum Basidiomycota (Fig. 28.3a). In the phylum Ascomycota, the most common orders obtained were the Eurotiales (61.36%), followed by the Hypocreales (20.45%), Saccharomycetales (6.81%), Dothidiales (2.27%), Xylariaceae (2.27%), Pleosporales (2.27%), Microascales (2.27%), and incertae sedis (2.27%), which in this case correspond to the genus Staphylotrichum (Fig. 28.3b). Likewise, the Ascomycota genera with a higher number of species (in parenthesis) were Aspergillus (11), Penicillium (7), Fusarium (6), Neosartorya (4), Trichoderma (3), Candida (2), and Byssochlamys (2). The rest of the fungal strains were represented by only one species. The phylum Basidiomycota was represented by two orders Sporidiobolales (75%) and Polyporales (25%), which were comprised by the yeast-like fungi Rhodotorula (3) and Pycnoporus sanguineus (1), respectively (Fig. 28.3c).

Percentage of fungi isolated from extreme environments according to phylum and order
From the list of isolated fungi (Table 28.1), some species have been reported as cosmopolitan fungi with worldwide distribution, such as Aspergillus fumigatus and Aspergillus terreus, Penicillium glabrum (formerly Penicillium frequentans) and Penicillium oxalicum, Neocosmospora (=Fusarium) solani, Cladosporium sphaerospermum, Trichoderma viride, and Trichoderma inhamatum (Domsch et al. 1980). Penicillium oxalicum was isolated from crude oil-impacted soil in the Borneo Islands (Chaillan et al. 2004). Aspergillus fumigatus has been widely reported as an inhabitant of soils polluted with hydrocarbons such as crude oil, kerosene, and diesel (April et al. 1998; Bento and Gaylarde 2001; Chaillan et al. 2004; Gesinde et al. 2008; Hemida et al. 1993; Oudot et al. 1993). In addition, A. fumigatus was reported as capable of degrading 20% of crude oil (Oudot et al. 1993). Aspergillus terreus was isolated from oil-polluted soils, gasoline, and kerosene (Chaillan et al. 2004; Colombo et al. 1996; Hemida et al. 1993; Uzoamaka et al. 2009), and possesses the capability of degrading up to 30% of crude oil (Algounaim et al. 1995). Neocosmospora solani was isolated from oil-polluted soils, asphalt, and kerosene (Colombo et al. 1996; Hemida et al. 1993; Naranjo et al. 2007, 2008) and could degrade up to 19% of the crude oil in 30 days (Chaineau et al. 1999).
Interestingly, only yeast-like fungi were isolated from EHCO (sample taken directly from an EHCO well), such as Candida tropicalis, Candida viswanathii, Rhodotorula mucilaginosa, and Cyberlindnera (=Williopsis) saturnus. Candida tropicalis was isolated previously from oil-polluted soils (April et al. 1998; Chaillan et al. 2004), in agreement with a prior report in which the yeasts Candida palmioleophila and Meyerozyma (=Pichia) guilliermondii were capable of the degradation of resins and asphaltenes (Chaillan et al. 2004).
Surprisingly, the common species isolated from all petroleum substrates was R. mucilaginosa which, as further discussed, is one of the most tolerant species to EHCO and PAHs. Rhodotorula mucilaginosa was also isolated by Gallego et al. (2007), reporting that it could degrade linear alkanes, C11-C33, branched alkanes, isoprenoids, and cycloalkanes. Other investigations have described R. mucilaginosa as halotolerant (it’s able to grow in concentrations higher than 2.5 M of NaCl); it tolerates a wide pH range of 2–10, and is capable of degrading nitrobenzene (Gross and Robbins 2000; Lahav et al. 2002; Turk et al. 2011; Zheng et al. 2009; Urbina and Aime 2018). More recently, Naranjo et al. (2015) described the isolation and molecular identification of fungal strains from naphtha systems in the oil industry, where the naphtha-tolerant R. mucilaginosa was the most predominant yeast-like fungal species.
The rest of the fungal species, such as Byssochlamys lagunculariae (formerly Byssochlamys nivea), Fusarium venenatum, Trametes coccinea, and Staphylotrichum sp., are new reports for science as species capable of inhabiting EHCO or oil-polluted soils that also have a great potential to be used for mycoremediation or EHCO bioconversion purposes.
3.3 Fungal Screening to Determine Hydrocarbonoclastic Potential and Tolerance to EHCO and HPAs: Obtaining the Powerful Biocatalysts
Our in vitro results show that all isolated fungi were tolerant to 1% EHCO. The most toxic compounds for fungi were DBT and phenanthrene, which showed a growth inhibition of more than 50%. These findings are presented in a cladogram where fungal species are discriminated by functional groups according to their tolerance to EHCO and PAHs (Fig. 28.4) as follows:

Dendrogram of tolerance groups of fungal species according to their tolerance to PAHs
Group No. 4 is the most tolerant capable of growth at 100% rate in all hydrocarbons studied in comparison to the control medium. Interestingly, this group is constituted by the filamentous fungi species B. lagunculariae 87, Penicillium (=Eupenicillium) javanicum, and Penicillium sp. 37, and the unicellular fungi C. tropicalis, C. viswanathii, R. mucilaginosa (three strains), and Ci. saturnus. All these unicellular fungi were isolated from EHCO and, in the case of R. mucilaginosa, were also isolated from oil-polluted soils and naphtha distribution system. These results strongly suggested that the fungal strains with greater hydrocarbonoclastic and tolerance capabilities are associated with EHCO wells and belong to unicellular fungi.
Group No. 1, the second most tolerant group, comprised of fungi that could grow at 95.81 ± 2.35% rate in EHCO, 92.62 ± 2.32% rate in naphthalene, and 80.14 ± 3.52% rate in pyrene. However, their growth was inhibited at approximately 50% rate in DBT and phenanthrene. This group clustered the major amount of fungal strains studied and was constituted by Aspergillus sp. (1–6), A. fumigatus, A. terreus, B. lagunculariae 89, N. solani, Fusarium venenatum 70, A. fischeri (formerly Neosartorya pseudofischeri and Neosartorya spinosa), Paecilomyces sp., Talaromyces (=Penicillium) aculeatum, Penicillium glabrum (formerly Penicillium frequentans), Penicillium oxalicum, Trametes coccinea, Staphylotrichum sp., and Trichoderma inhamatum. The rest of the tolerance groups (Nos. 2 and 3) were the less tolerant, and their growth was inhibited to more than 50% rate in the same conditions.
According to ranges of HPA tolerance, Group No. 4 has a 100% tolerance for all PAHs studied. Group No. 3 tolerates CEP and DBT 100% but it is less tolerant to phenanthrene (12–45%), naphthalene (38–57%), and pyrene (38–54%). Group No. 2 tolerates CEP in 19–61%, DBT (0–57%), phenanthrene (0–60%), and pyrene (0–100%). Finally, Group No. 1 showed a low tolerance to CEP (11.25–62%) but with a wide range of tolerance to the rest of the PAHs studied: DBT (0–100%), phenanthrene (9–100%), naphthalene (75–100%), and pyrene (50–100%).
The discrimination of the different tolerance groups of fungi according to their tolerance rate to EHCO and PAH compounds was also evaluated (Fig. 28.5). Group No. 4 clustered fungi with the same proportion tolerance for all the hydrocarbons, while Group No. 3 showed a greater tolerance for EHCO, and finally fungal species in Groups No. 1 and 2 had greater tolerance to naphthalene alone. Likewise, this analysis corroborates the fact that DBT and phenanthrene were the most toxic PAHs for the fungi isolated in this study.

Proportion of tolerance to the different PAHs by fungal group corresponding to the dendrogram. PYR Pyrene, NAPH Naphthalene, PHEN Phenanthrene, DBT Dibenzothiophene, EHCO Extra-heavy crude oil
Another important result observed at the present work was the difference observed in tolerance to hydrocarbons between species of the same genus, including the difference between strains of the same species (Fig. 28.6). It is known that a functional group is composed of microorganisms that, regardless of their taxonomic classification, present an identical pattern of biochemical responses in the use and transformation of organic substrates.

Percentages of growth in the presence of PAHs compared with the control of fungal species of the genera: (a) Aspergillus, (b) Fusarium, and (c) Penicillium. PYR Pyrene, NAPH Naphthalene, PHEN Phenanthrene, DBT Dibenzothiophene, EHCO Extra-heavy crude oil
In the case of the genus Aspergillus, any of its species could tolerate DBT and phenanthrene, and their growth was inhibited to more than 50% rate (Fig. 28.6a). Among Aspergillus species, the most tolerant were Aspergillus sp. 4–6, which were isolated from an oil-polluted soil adjacent to an oil pit. As can be appreciated, the species isolated from this site also showed a higher tolerance for DBT, phenanthrene, and naphthalene, in comparison to the species isolated from the natural asphalt Lake of Guanoco.
In the case of Fusarium species, the most tolerant species was N. solani, widely reported as a hydrocarbon degrader (León et al. 2007; Naranjo et al. 2007, 2008), and F. venenatum, which has not been previously reported as a hydrocarbon degrader (Fig. 28.6b). Interestingly, different strains of F. venenatum isolated from natural asphalt Lake of Guanoco showed different degrees of tolerance to EHCO as strain 70 grew at 100% rate while strain 69 only grew at 19% rate, compared to the control medium.
In the case of Penicillium species, the most tolerant species were Penicillium sp. 37 and Penicillium javanicum, both able to grow at 100% rate in all hydrocarbons tested (Fig. 28.6c). However other species of this same genus (such as P. oxalicum) were not able to tolerate hydrocarbons with growth inhibitions greater than 50% rate.
The results showed here strongly suggested that the fungal strains belonging to the same species do not necessarily have the same hydrocarbonoclastic abilities and characteristics to tolerate xenobiotic compounds derived from petroleum. Likewise, fungal strains belonging to the same species but isolated from different places do not necessarily have the same characteristic and behavior.
4 Relationship Between the LDS and EHCO Bioconversion in Mitosporic Extremophilic Hydrocarbonoclastic Fungi: Inducing a Powerful Exoenzymatic System
The LDS includes a large range of oxidoreductases and hydroxylases, such as laccases (LACs) and high-redox-potential ligninolytic peroxidases like lignin peroxidase ( LIPs), manganese peroxidases (MNPs), versatile peroxidases ( VEPs), and others. Lignin peroxidases are able to directly oxidize non-phenolic units, while MNPs and LACs oxidize preferentially phenolic units, but also act on non-phenolic units when mediators (e.g., 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid), commonly known as ABTS) are present in the reaction mixture, whereas VEPs are able to combine the catalytic properties of LIPs and MNPs (Martínez et al. 1996, 2005; Ruiz-Dueñas et al. 1999; Saparrat et al. 2002; Gianfreda and Rao 2004).
Concurrently, research is focused on the conversion of heavy oils to lighter oils mediated by biocatalyst. A few of the biotechnological strategies to improve the EHCO physical-chemical properties have promoted an increase in their commercial value. Our research team proposed 13 years ago the use of the extracellular oxidative enzymes ( EOE) of the LDS present in extremophilic fungi as biocatalysts (Naranjo et al. 2007, 2008; León et al. 2007) based on the following facts: (1) lignin is a complex biopolymer structurally similar to asphaltenes, molecules responsible for the high viscosity of EHCO (Fig. 28.7), and (2) ligninolytic fungi have a unique and powerful LDS that includes a wide range of unspecific oxidoreductases and hydrolases which are involved in the transformation and degradation processes of polymeric substances in partially degraded and oxidized soluble products that can be easily assimilated by microorganisms (Gianfreda and Rao 2004, Naranjo et al. 2007, 2013). Studies dealing with the interactions between extremophilic microorganisms and crude oil have led to the identification of biocatalysts which, through multiple biochemical pathways, carry out the desulfurization, denitrogenation, and demetallation reactions in oils.

Representative molecular structures of asphaltenes and lignin. (a) A single condensed polycyclic aromatic core; (b) multiple smaller polycyclic aromatic cores with aliphatic bridges; (c, d) lignin structures
The efficiency of the degradation process by biological systems depends on the susceptibility of these compounds to be degraded and converted into less toxic compounds; therefore, supposed susceptibility is directly related to the chemical structure, concentration, and physical properties of the hydrocarbon (Foght 2004, Gianfreda and Rao 2004; Naranjo et al. 2007, 2013). If the hydrocarbons are soluble, they can be assimilated by the microorganisms more easily and, if they are insoluble they must be transformed into soluble compounds so that they can be used later. However, the first step in the transformation of insoluble compounds is usually catalyzed by extracellular oxidative enzymes of microorganisms, which are released by the cells to the surrounding environment (Foght 2004).
The fungal mineralization and transformations of EHCO and/or its asphaltene fraction by fungi have been documented previously; however they are still controversial. The ability of the fungi Aspergillus flavipes, Aspergillus (=Emericella) nidulans, P. javanicum, and Parascedosporium putredinis (=Graphium) putredinis to degrade resins (15–28%) and asphaltenes (15–40%) was described by Oudot et al. (1993). Chaillan et al. (2004) reported the ability of filamentous fungi Albonectria rigidiuscula (formerly Fusarium decemcellulare) and Paecilomyces variotii, and the yeasts Candida palmioleophila and M. guilliermondii to degrade a range of 10–15% of resins and asphaltenes. Transformations of petroporphyrins and asphaltenes by chloroperoxidase (CPO) of Caldariomyces fumago, a protein with high peroxidase activity and versatility that can halogenate aromatic molecules like polycyclic aromatic hydrocarbons (PAHs), were described by Fedorak et al. (1993).
Later, García-Arellano et al. (2004) showed through Fourier-transform infrared spectroscopy (FT-IR ) that a chemically modified cytochrome C catalyzed the oxidation of carbon and sulfur atoms in the rich fraction of petroporphyrins of the asphaltenes, with 74% and 95% Ni and V removal, respectively. The chemical modification of the surface of the protein with polyethylene glycol resulted in the formation of a conjugate polymer-protein soluble in organic solvents, and the methyl esterification of the heme group increasing the hydrophobicity of its active site. In fact, the highest activity was detected in a tertiary mixture of solvents with 5% water. In this case, the enzymatic biotreatment of asphaltenes represents an interesting alternative for the elimination of heavy metals and the reduction of poisoning catalysts after cracking and hydrocracking of crude oils.
Recently, Uribe-Álvarez et al. (2011) described for the first time the ability of A. fischeri, isolated from natural asphalt Lake of Guanoco in Venezuela, of growing on asphaltenes as the sole carbon and energy source and to mineralize 13.2% of asphaltenes. Ayala et al. (2012) described the biotransformation of porphyrin-free asphaltene fraction catalyzed by CPO-based biocatalyst to reduce coke formation during thermal decomposition in the oil industry. On the other hand, Hernández-López et al. (2016) evaluated the capacity of A. fischeri to transform high-molecular-weight polycyclic aromatic hydrocarbons (HMW-PAHs ) and asphaltenes as the sole carbon source by reverse-phase high-performance liquid chromatography (HPLC), nano-LC mass spectrometry, and IR spectrometry, together with a comparative microarray study and the complete genome of A. fischeri. The formation of hydroxy and ketone groups on the PAH molecules and the internalization of aromatic substrates into fungal cells suggested oxidation of these recalcitrant compounds mediated by the cytochrome P450 system.
Finally, Pourfakhraei et al. (2018) described using saturate, asphaltene, resin, and aromatic (SARA ) analysis chromatograms that the wood-decaying fungus Daedaleopsis sp. can degrade asphaltene and aromatic fraction with a reduction of 88.7% and 38%, respectively, with an increase of 44.8% in the saturate fraction. They also reported that LAC-, LIP-, and MNP-specific activities from LDS were induced in the presence of heavy crude oil (HCO) as the sole carbon source and energy.
In order to confirm the relationship between LDS and bioconversion of EHCO claimed by Naranjo et al. (2013), the study of the enzymatic activities of specific LACs, LIPs, and MNPs induced in minimal medium Czapek supplemented with EHCO as the sole source of carbon and energy was performed. The fungi Fusarium proliferatum (BM-02), Pestalotiopsis palmarum (BM-04), Aspergillus terreus (BM-36), and Pseudallescheria angusta (BM-39) were randomly selected from the list of fungal strains isolated from extreme environments studied (Table 28.1). At first, a screening was carried out to confirm their hydrocarbonoclastic capabilities against several PAH compounds (naphthalene, phenanthrene, dibenzothiophene, and pyrene) according to Naranjo et al. (2013). The phenotypic results showed that all fungi were able to grow using EHCO and all PAHs with the exception of F. proliferatum (BM-02) and Ps. angusta (BM-39) that were unable to grow using phenanthrene (Table 28.2, Fig. 28.8). Regarding the study of the enzymatic activities, the results showed that all fungi induced the enzymatic activities studied in the presence of EHCO compared with the control (Fig. 28.9a, b and c). However, interestingly, the lignin peroxidase was strongly induced with EHCO in the fungi Pestalotiopsis palmarum (BM-04), Aspergillus terreus (BM-36), and Pseudallescheria angusta (BM-39), with the exception of Fusarium proliferatum (BM-02), which showed a few inductions in all activities studied (Fig. 28.9c).

Growth in minimal modified culture medium Czapek supplement with several PAHs as the sole carbon source, such as naphthalene, phenanthrene, pyrene, dibenzothiophene (DBT), and extra-heavy crude oil (EHCO). The fungi Fusarium proliferatum (BM-02), Pestalotiopsis palmarum (BM-04), Aspergillus terreus (BM-36), and Pseudallescheria angusta (BM-39) were randomly selected from the list of extremophilic fungi isolated from extreme environments

Relationship between the LDS and the bioconversion of EHCO in the fungi Fusarium proliferatum (BM-02), Pestalotiopsis palmarum (BM-04), Aspergillus terreus (BM-36), and Pseudallescheria angusta (BM-39)
5 Conclusions and Future Perspectives
It is unquestionable that the modern world was created to the image of petroleum with a concomitant reduction of conventional crude oil reserves and a rising of global demand for fuels and oil derivatives. Consequently, with the increasing of exploitation of unconventional crude reserves, the development and improvement of clean-alternative fuel technologies are required with the mandatory establishment of innovative protocols for unconventional hydrocarbon exploitation. The studying and application of the petroleum biotechnology with promissory microorganisms, specially extremophilic hydrocarbonoclastic fungi and their powerful LDS, in the whole oil industry production chain, is a pathway for improving products and processes, guaranteeing the decreasing of operational costs with increased productivity capabilities with minimum environmental impact. Here, we introduced the term extremophilic hydrocarbonoclastic fungi as a large and heterogeneous group of cultivable fungi which can live optimally under extreme conditions and are characterized by having a high ability to grow using hydrocarbons as the sole carbon source and energy.
Treatment of the unconventional hydrocarbons under extreme conditions plays a vital role to save the ecosystems for anthropogenic intervention which leads to sustainable development. This book chapter shows, through a sequential and comprehensible explanation, a biotechnological strategies to study cultivable fungal biodiversity inhabiting in extreme environments for the isolation of powerful biocatalysts, following a simple and fast screening to determine both their hydrocarbonoclastic potential and tolerance to EHCO and HPAs.
The exploration of cultivable fungal communities from the extreme environments studied allows us to identify phylum Ascomycota as the most isolated fungal group, being Eurotiales the most common order obtained, followed by Hypocreales and Saccharomycetales, and the genera with the most number of species Aspergillus (11), Penicillium (7), Fusarium (6), Neosartorya (4), Trichoderma (3), Byssochlamys (2), and Candida (2), respectively.
Only a few strains of the phylum Basidiomycota were isolated, composed mainly by the orders Sporidiobolales followed by the Polyporales, which are constituted by the yeast-like fungi Rhodotorula (3) and Pycnoporus sanguineus (1), respectively. However, surprisingly the more common species isolated from all of the extreme environments studied was R. mucilaginosa.
It is well understood that the study of microorganisms with potential in oil biotechnology requires research of both their hydrocarbonoclastic potential and tolerance abilities to EHCO and HPAs. These abilities have been pointed out here as crucial for the appropriated selection of powerful hydrocarbonoclastic extremophilic fungi as biocatalysts. In our research, the fungal strains with greater hydrocarbonoclastic and tolerance capabilities were isolated from Carabobo-EHCO wells and belonging mainly to unicellular fungi, such as C. tropicalis, C. viswanathii, Ci. saturnus, and R. mucilaginosa. These yeast-like fungi are the most tolerant species to EHCO and PAHs.
Another important result observed during our investigations was the difference obtained in tolerance to hydrocarbons between species of the same genus, including the difference between strains of the same species, such as Aspergillus, Fusarium, and Penicillium. Likewise, fungal strains belonging to the same species but isolated from different extreme environments do not necessarily have the same hydrocarbonoclastic characteristic and behavior.
On the other hand, the relationship between the powerful LDS and EHCO bioconversion in extremophilic hydrocarbonoclastic fungi was confirmed, where a strong induction of the lignin peroxidase activity and a low induction of the laccase activity were obtained repetitively, in all the fungal strains studied.
Further research is required to develop new visions and perspectives, novel clean technologies, confident alternatives, and strategies to ensure an economically profitable development with social justice, and ecological sustainability, safeguarding the future and quality of life for the next generations. The perspective of this chapter is the potential application of the promissory extremophilic hydrocarbonoclastic fungi to be used as biocatalysts for mycoremediation or EHCO bioupgrading process under stressed conditions to increase the revenues for the industry dedicated to the exploitation of unconventional crudes.
References
AlGounaim MY, Diab A, AlAbdulla R, AlZamin N (1995) Effect of petroleum oil pollution on the microbiological populations of the desert soil of Kuwait. Arab Gulf J Sci Res 13(3):653–672
Amund O, Adebowale AA, Ugoji EO (1987) Occurrence and characteristics of hydrocarbon-utilizing bacteria in Nigerian soils contaminated with spent motor oil. Indian J Microbiol 27:63–67
April TM, Abbott SP, Foght JM, Currah RS (1998) Degradation of hydrocarbons in crude oil by the ascomycete Pseudallescheria boydii (Microascaceae). Can J Microbiol 44:270–278
Arellano T, Infante C, Naranjo L (2008) Manejo integral de fosas de hidrocarburos generadas por la actividad petrolera venezolana. Thesis for Magister in Environmental Management, UNEFA 1–250
Arulazhagan P, Mnif S, Rajesh Banu J, Huda Q, Jalal MAB (2017) HC-0B-01: biodegradation of hydrocarbons by extremophiles. In: Heimann K, Karthikeyan O, Muthu S (eds) Biodegradation and bioconversion of hydrocarbons. Environmental footprints and eco-design of products and processes. Springer, Singapore, pp 137–162
Atlas RM, Bartha R (1972) Degradation and mineralization of petroleum by two bacteria isolated from coastal waters. Biotechnol Bioeng 14:297–308
Ayala M, Hernández-López EL, Perezgasga L, Vázquez-Duhalt R (2012) Reduced coke formation and aromaticity due to chloroperoxidase-catalyzed transformation of asphaltenes from Maya crude oil. Fuel 92:245–249
Bartha R, Atlas RM (1977) The microbiology of aquatic oil spills. Adv Appl Microbiol 22:225–226
Benka-Coker MO, Ekundayo JA (1997) Applicability of evaluating the ability of microbes isolated from an oil spill site to degrade oil. Environ Monit Assess 45:259–272
Bento FM, Gaylarde CC (2001) Biodeterioration of stored diesel oil: studies in Brazil. Int Biodeter Biodegr 47:107–112
Calomiris JJ, Austin B, Walker JD, Colwell RR (1986) Enrichment for estuarine petroleum-degrading bacteria using liquid and solid media. J Appl Bacteriol 42:135–144
CDC-NIOSH (2015) Pocket guide to chemical hazards-petroleum distillates (naphtha). www.cdc.gov
Chaillan F, Flèche AL, Bury E, Phantavong Y, Grimont P, Saliot A, Oudot J (2004) Identification and biodegradation potential of tropical aerobic hydrocarbon-degrading microorganisms. Res Microbiol 155:587–595
Chaineau CH, Morel J, Dupont J, Bury E, Oudot J (1999) Comparison of the fuel oil biodegradation potential of hydrocarbon-assimilating microorganisms isolated from a temperate agricultural soil. Sci Total Environ 227:237–247
Colombo JC, Cabello M, Arambarri AM (1996) Biodegradation of Aliphatic and aromatics hydrocarbons by natural soil microflora and pure cultures of imperfect and lignolytic fungi. Environ Pollut 94:355–362
Coyne M (2000) Microbiología del Suelo: un enfoque exploratorio, 1ª Edición edn. Editorial Paraninfo SA, Madrid, Spain, pp 1–440
Dávila A, Vázquez-Duhalt R (2006) Enzimas ligninolíticas fúngicas para fines ambientales. Mensaje Bioquímico 30:29–55
Domsch KH, Gams W, Anderson TH (1980) Compendium of soil fungi, vol 1, pp 1–860
Fedorak PM, Semple KM, Vazquez-Duhalt R, Westlake DWS (1993) Chloroperoxidase mediated modifications of petroporphyrins and asphaltenes. Enzyme Microb Technol 15:429–437
Foght JM (2004) Whole-cell bio-processing of aromatic compounds in crude oil and fuels. In: Petroleum biotechnology: developments and perspectives. Elsevier Science, Amsterdam, pp 145–175
Gallego JLR, García-Martınez MJ, Llamas JF, Belloch C, Pelaez AI, Sanchez J (2007) Biodegradation of oil tank bottom sludge using microbiol Consortia. Biodegradation 18:269–281
García-Arellano H, Buenrostro-Gonzalez E, Vazquez-Duhalt R (2004) Biocatalytic transformation of petroporphyrins by chemical modified cytochrome c. Biotechnol Bioeng 85:790–798
Gesinde AF, Agbo EB, Agho MO, Dike EFC (2008) Bioremediation of some Nigerian and Arabian crude oils by fungal isolates. Int J Pure Appl Sci 2:37–44
Gianfreda L, Rao MA (2004) Potential of extra-cellular enzymes in remediation of polluted soils: a review. Enzyme Microb Technol 35:339–354
Gross S, Robbins EI (2000) Acidophilic and acid-tolerant fungi and yeasts. Hydrobiologia 433:91–109
Hemida SK, Bagy MMK, Khallil AM (1993) Utilization of hydrocarbons by fungi. Cryptogamie Mycologie 14:207–213
Hernández-López EL, Perezgasga L, Huerta-Saquero A, Vazquez-Duhalt R (2016) Biotransformation of petroleum asphaltenes and high molecular weight polycyclic aromatic hydrocarbons by Neosartorya fischeri. Environ Sci Pollut Res Int 23:10773–10784
Lahav R, Nejidat A, Abeliovich A (2002) The identification and characterization of osmotolerant yeast isolates from chemical wastewater evaporation ponds. Microb Ecol 43:388–396
León V, Córdova J, Muñoz S, De Sisto A, Naranjo L (2007) Process for the upgrading of heavy crude oil, extra-heavy crude oil or bitumens through the addition of a biocatalyst. United States Patent Application 20070231870
León Y, De Sisto A, Inojosa Y, Malaver N, Naranjo-Briceño L (2009) Identificación de biocatalizadores potenciales para la remediación de desechos petrolizados de la Faja Petrolif́ era del Orinoco. RET 1:12–25
Macelroy RD (1974) Some comments on the evolution of extremophiles. BioSystem 6:74–75
Madigan MT, Martinko JM, Bender KS, Buckley DH, Stahl DA (2015) Brock biology of microorganisms, 14th edn. Pearson, Boston
Martínez MJ, Ruiz-Dueñas FJ, Guillén F, Martínez AT (1996) Purification and catalytic properties of two manganese-peroxidase isoenzymes from Pleurotus eryngii. Eur J Biochem 237:424–432
Martínez AT, Speranza M, Ruiz-Duenas FJ, Ferreira P, Camarero S, Guillen F, Martınez MJ, Gutierrez A, del Río JC (2005) Biodegradation of lignocellulosics: microbial, chemical, and enzymatic aspects of the fungal attack of lignin. Int Microbiol 8:195–204
Moore D, Robson GD, Trinci AP (2011) 21st century guidebook to fungi with CD. Cambridge University Press, New York, pp 1–640
Nadon L, Siemiatycki J, Dewar R, Krewski D, Gérin M (1995) Cancer risk due to occupational exposure to polycyclic aromatic hydrocarbons. Am J Ind Med 28(3):303–324
Naranjo L, Urbina H, De Sisto A, Leon V (2007) Isolation of autochthonous non-white rot fungi with potential for enzymatic upgrading of Venezuelan extra-heavy crude oil. Biocatal Biotransformation 25:341–349
Naranjo L, Urbina H, González M, Córdova J, Muñoz S, León V. (2008) Potential of autochthonous non-white rot fungi for partial enzymatic conversion (PEC-IDEA Technology) of Venezuelan extra-heavy crude oil. In: Proceeding of the 6th international symposium on fuels and lubricants (ISFL). New Delhi, India. Paper No. 128
Naranjo-Briceño L, Perniá B, Guerra M, Demey JR, González M, De Sisto A, Inojosa Y, Fusella E, Freites M, Yegres JF (2013) Potential role of oxidative exoenzymes of the extremophilic fungus Pestalotiopsis palmarum BM-04 in biotransformation of extra-heavy crude oil. Microb Biotechnol 6(6):720–730
Naranjo L, Pernía B, Inojosa Y, Rojas D, Sena D’Anna L, González M, De Sisto A (2015) First evidence of fungal strains isolated and identified from naphtha storage tanks and transporting pipelines in Venezuelan oil facilities. Adv Microbiol 5:143–154
Obire O (1993) The suitability of various Nigerian petroleum fractions as substrate for bacterial growth. Discov Innov 5:45–49
Odokuma LO, Okpokwasili GC (1993) Seasonal ecology of hydrocarbon-utilizing microbes in the surface waters of a river. Environ Monit Assess 27(3):175–191
Oudot JP, Dupont J, Haloui S, Roquebert MF (1993) Biodegradation potential of hydrocarbon-degrading fungi in tropical soil. Soil Biol Biochem 25:1167–1173
OPEC (2017) OPEC annual statistical bulletin 2017. http://www.opec.org/opec_web/en/
Pernía B, Demey JR, Inojosa Y, Naranjo L (2012) Biodiversidad y potencial hidrocarbonoclástico de hongos aislados de crudo y sus derivados: un meta-análisis. Latinoam Biotecnol Amb Algal 3:1–40
Pernía B, Rojas-Tortolero D, Sena L, De Sisto A, Inojosa Y, Naranjo L (2018) Fitotoxicidad de HAP, crudos extra pesados y sus fracciones en Lactuca sativa: una interpretación integral utilizando un índice de toxicidad modificado. Rev Int Contam Ambient 34:79–91
Pourfakhraei E, Badraghi J, Mamashli F, Nazari M, Saboury AA (2018) Biodegradation of asphaltene and petroleum compounds by a highly potent Daedaleopsis sp. J Basic Microbiol:1–14
Prenafeta-Boldú FX, de Hoog GS, Summerbell RC (2018) Fungal communities in hydrocarbon degradation. In: McGenity T (ed) Microbial communities utilizing hydrocarbons and lipids: members, metagenomics and ecophysiology. Handbook of hydrocarbon and lipid microbiology. Springer, Cham, pp 1–36
Rampelotto PH (2013) Extremophiles and extreme environments. Life 3:482–485
Ruiz-Dueñas FJ, Martínez MJ, Martínez AT (1999) Molecular characterization of a novel peroxidase isolated from the lignolytic fungus Pleurotus eryngii. Mol Microbiol 31:223–235
Saparrat MCN, Guillén F, Arambarri AM, Martínez AT, Martínez MJ (2002) Induction, isolation, and characterization of two laccases from the with rot basidiomycete Coriolopsis rigida. Appl Environ Microbiol 68:1534–1540
Strausz OP, Mojelsky TW, Lown EM (1992) The molecular structure of asphaltenes: an unfolding story. Fuel 71:1355–1363
Terrer C, Vicca S, Hungate BA, Phillips RP, Colin Prentice I (2016) Mycorrhizal association as a primary control of the CO2 fertilization effect. Science 353:72–74
Turk M, Plemenitaš A, Gunde-Cimerman N (2011) Extremophilic yeasts: plasma-membrane fluidity as determinant of stress tolerance. Fungal Biol 115:950–958
Urbina H, Reyes A, Fusella E, González M, León V, Naranjo L (2007) Pycnoporus sanguineus IDEA, a laccase-overproducing fungi with high potential in partial enzymatic conversion (PEC-Technology) of Venezuelan extra-heavy crude oil. J Biotechnol 131(2 Supplement 1):S94–S95
Urbina H, Aime MC (2018) A closer look at Sporidiobolales: ubiquitous microbial community members of plant and food biospheres. Mycologia 110:79–92
Uribe-Álvarez C, Ayala M, Perezgasga L, Naranjo L, Urbina H, Vazquez-Duhalt R (2011) First evidence of mineralization of petroleum asphaltenes by a strain of Neosartorya fischeri. J Microbial Biotechnol 4:663–672
Uzoamaka GO, Floretta T, Florence MO (2009) Hydrocarbon degradation potentials of indigenous fungal isolates from petroleum contaminated soils. J Phys Nat Sci 3:1–6
Waldo GS, Carlson RM, Moldowan JM, Peters KE, Penner-Hahn JE (1991) Sulfur speciation in heavy petroleums: information from X-ray absorption near-edge structure. Geochim Cosmochim Acta 55:801–814
Zhang X, Li SJ, Li JJ, Liang ZZ, Zhao CQ (2018) Novel natural products from extremophilic fungi. Mar Drugs 16(6):4
Zheng C, Zhou J, Wang J, Qu B, Wang J, Lu H, Zhao H (2009) Aerobic degradation of nitrobenzene by immobilization of Rhodotorula mucilaginosa in polyurethane foam. J Hazard Mater 168:298–303
Acknowledgments
This work was supported by the Projects FONACIT No. 2005000440, Sub-Project 3: MISIÓN CIENCIA No. 2007001401 and FONACIT No. G- 2011000330. The authors recognize Dr. V. León for being a pioneer of the Petroleum Biotechnology in Venezuela, and dedicate this work to the memory of Dr. J. Demey, rest in peace. The authors thank Judith Nyisztor and Aitana Naranjo for grammatical support.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2019 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Naranjo-Briceño, L. et al. (2019). Potential Role of Extremophilic Hydrocarbonoclastic Fungi for Extra-Heavy Crude Oil Bioconversion and the Sustainable Development of the Petroleum Industry. In: Tiquia-Arashiro, S., Grube, M. (eds) Fungi in Extreme Environments: Ecological Role and Biotechnological Significance. Springer, Cham. https://doi.org/10.1007/978-3-030-19030-9_28
Download citation
DOI: https://doi.org/10.1007/978-3-030-19030-9_28
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-19029-3
Online ISBN: 978-3-030-19030-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)