
Abstract
5-Hydroxymethylcytosine (5-hmC) has a critical role in the epigenetic regulation of many cellular processes, including differentiation, cytokine responses, and memory formation. This modified base is generated by the TET enzymes through oxidizing 5-methylcytosine and can be further oxidized to 5-formylcytosine and 5-carboxylcytosine. The level and distribution of 5-hmC in the genome is tightly controlled through transcription of TET genes, post-translational modifications on the TET enzymes, and various interacting partners of TET enzymes. Deregulation of 5-hmC is often found in pathological conditions and plays crucial roles in disease progression. This chapter will discuss recent findings regarding 5-hmC functions, TET proteins, and their gene regulations, as well as their relationships to cancer.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
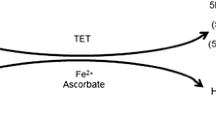
In mammalian DNA, cytosine bases found within the context of a 5′-CpG-3′ dinucleotide are substrates for covalent modification to 5-methylcytosine (5-mC) by the DNA methyltransferases DNMT1, DNMT3A, and DNMT3B, which then can be oxidized sequentially by Ten-Eleven Translocation methylcytosine dioxygenases TET1, TET2, and TET3 to produce 5-hydroxymethylcytosine (5-hmC), 5-formylcytosine (5-fC), and 5-carboxylcytosine (5-caC) (Wu and Zhang 2017) (Fig. 1a). With the discovery of TET catalytic activity (Iyer et al. 2009; Tahiliani et al. 2009), there have been great advances in understanding the role of 5-hmC in various biological processes. Emerging evidence indicates that 5-hmC plays important roles in many cellular processes, including differentiation, cytokine responses, and memory formation (Cao 2016; Vogel Ciernia and LaSalle 2016; An et al. 2017). In this chapter, we discuss the regulation of TET enzymes and their roles in modulating the 5-hmC landscape as well as the impact of TET and 5-hmC deregulation in human disease.

Regulation and functions of 5-hmC. (a) Biochemical pathways that control cytosine modifications. Unmodified cytosine is methylated to 5-mC by DNMTs using SAM as methyl donor. TETs oxidize 5-mC to 5-hmC, 5-fC, and 5-caC stepwise using 2-OG and O2 as co-substrates and Fe2+ as a co-factor (green arrows). 5-hmC and 5-caC can be converted to unmodified cytosine in vitro by direct removal of 5-modifications through DNMT3A/B (purple arrows). 5-fC and 5-caC can be excised by TDG and/or NEIL1 and replaced with unmodified cytosine through base-excision repair (blue arrows). (b) 5-hmC promotes passive demethylation through replication. Newly synthesized DNA contains no modified cytosines. When the CpG dinucleotide on the old strand is methylated, DNMT1 can bind and methylate the complementary CpG dinucleotide on the newly synthesized strand, maintaining methylation over DNA replication and cell division. When the CpG dinucleotide in the old strand is hydroxymethylated, however, if DNMT1 fails to bind, the newly synthesized strand will remain unmodified, leading to loss of modification through multiple rounds of DNA replication. (c) Stable 5-hmC promotes open chromatin and gene transcription. Abbreviations: 2-OG 2-oxoglutarate, 5-mC 5-methylcytosine, 5-hmC 5-hydroxymethylcytosine, 5-fC 5-formylcytosine, 5-caC 5-carboxylcytosine, 5-mod 5-modified cytosine, BER base-excision repair, SAM S-adenosyl methionine, SAH S-adenosyl homocysteine, TSS transcription start site
2 TET and 5-hmC Functions
2.1 Mammalian Cytosine Modifications
In mammalian genomes, cytosines present within 5′-CpG-3′ dinucleotides can be methylated to 5-mC by DNMTs using S-adenosylmethionine (SAM) as the methyl group donor. DNMT1 has optimal activity in methylating hemimethylated cytosines and thus functions mainly to maintain DNA methylation patterns after DNA replication (Lee et al. 2014). In contrast, DNMT3A and DNMT3B function mainly as de novo methyltransferases (Lee et al. 2014) (Fig. 1b). Many studies published since the 1980s have established that 5-mC is a base that regulates chromatin function: When found within gene bodies, 5-mC is associated with gene expression (Jjingo et al. 2012; Jones 2012; Varley et al. 2013), and when concentrated in CpG islands that overlap gene promoters, the base represses gene transcription, a phenomenon commonly observed in cancer cells (Feinberg and Vogelstein 1983; Gama-Sosa et al. 1983; Baylin et al. 1986; Feinberg and Tycko 2004).
The first oxidized product of 5-mC, 5-hmC, was originally identified as a specialized base in T2, T4, and T6 bacteriophages (Wyatt and Cohen 1953). Although the presence of 5-hmC was also observed in mammalian DNA, it was believed that this base was the result of oxidative damage (Tardy-Planechaud et al. 1997; Valinluck and Sowers 2007). 5-hmC did not receive major attention as a chromatin regulating base until it was discovered that 5-hmC is the oxidized product of TET dioxygenase activity using 5-mC as a substrate (Iyer et al. 2009; Tahiliani et al. 2009). Since then, multiple 5-hmC specific assays have been developed (Table 1), which have facilitated the elucidation of 5-hmC functions.
2.2 Roles of 5-hmC
5-hmC is a relatively rare base in mammalian genomes, and its abundance is highly tissue dependent. For example, 5-hmC consists of 0.01–0.3% of all cytosine species in myeloid cells (Ko et al. 2010; Madzo et al. 2014); constitutes around 1% of all cytosines in cerebral tissue (Kriaucionis and Heintz 2009; Wen et al. 2014; Field et al. 2015); and is present at levels as high as 5% of all cytosines in undifferentiated embryonic stem (ES) cells (Tahiliani et al. 2009). Currently, two major functions of 5-hmC are proposed: the base is a component of the demethylation process, and the base can also function as a stable epigenetic mark with distinct roles in chromatin structure and transcriptional regulation.
5-hmC facilitates demethylation both within passive and active demethylation pathways (Fig. 1). When DNA is replicated, a newly inserted cytosine on the daughter strand is unmodified at first. If the cytosine base within the parent 5′-CpG-3′ dinucleotide is methylated, then DNMT1 methylates the cytosine within the newly synthesized strand since its highest activity is on hemi-methylated CpG dinucleotides, and DNMT1 sits at the replication fork, thereby maintaining the DNA methylation pattern (Fig. 1b). However, if the cytosine base within the parent DNA strand is hydroxymethylated, then maintenance of this base in the face of DNA replication requires two enzymes, first a DNMT to produce 5-mC, and then a TET to convert that base to 5-hmC. Therefore, in the face of DNA replication, if a DNMT fails to recognize a 5-hmC, that base will result in an unmodified cytosine in the daughter strand. This process is referred to as passive demethylation (Valinluck and Sowers 2007; Seiler et al. 2018). A more active and cell-cycle independent demethylation pathway involves further oxidation of 5-hmC by TETs to 5-fC and 5-caC (Ito et al. 2011), both of which can be replaced by unmodified cytosines through base-excision repair (BER) by thymine-DNA glucosylase (TDG) (He et al. 2011; Maiti and Drohat 2011; Coey et al. 2016; Pidugu et al. 2016) (Fig. 1a). 5-caC, but not 5-fC, can also undergo BER by Nei-like 1 (NEIL1) excision (Slyvka et al. 2017) (Fig. 1a). In addition to BER-mediated demethylation, some studies have also found that DNMT3A/3B have the ability to remove the 5-modification from 5-hmC and 5-caC directly, but not from 5-mC or 5-fC, resulting in unmodified cytosine (Chen et al. 2012; Liutkevičiūtė et al. 2014) (Fig. 1a).
5-hmC can also function as a stable and distinct epigenetic mark associated with active gene transcription. Many reports have found that 5-hmC is strongly associated with promoters, enhancers, and transcription factor binding sites (Madzo et al. 2014; Mariani et al. 2014; Vasanthakumar and Godley 2015) (Fig. 1c). Similarly, 5-hmC was shown to be associated with active transcription and histone modifications that mark open chromatin (Ficz et al. 2011; Szulwach et al. 2011; Madzo et al. 2014; Lin et al. 2017) (Fig. 1c). Evidence for the role of 5-hmC as a stable and actively maintained epigenetic mark is most apparent in the context of cell differentiation, where gene expression and chromatin structure undergo major changes. For example, in the course of erythropoiesis, 5-hmC at certain genomic loci remains highly enriched despite a global decrease in total 5-hmC level and multiple rounds of DNA replication (Madzo et al. 2014). To maintain 5-hmC during consecutive rounds of DNA replication, both DNMTs and TETs must be involved, indicating a functional role of 5-hmC at these loci.
In addition to its role in regulating gene expression and chromatin structure, recent studies have revealed potential involvement of 5-hmC in DNA damage repair. During murine embryonic development, Tet1 loss-of-function oocytes have a much higher rate of defective meiosis and unresolved double strand breaks compared to those with wild-type Tet1 (Yamaguchi et al. 2012). In addition, DNA damage in mouse Purkinje cells leads to increased 5-hmC levels (Jiang et al. 2015). Further studies in mouse ES cells showed that knock-out (KO) of all three Tet genes led to a high level of mitotic defects, as measured by the presence of chromatin fragments in anaphase, which resulted from a higher sensitivity to replication stress and delayed DNA repair (Kafer et al. 2016). In human cancer cell lines, 5-hmC foci co-localize with DNA damage foci marked by γH2AX and 53BP1 upon induced DNA damage (Kafer et al. 2016). When TET2 is knocked-down with shRNA, 5-hmC foci no longer form, suggesting that TET2 is directly involved the DNA damage response (Kafer et al. 2016). It is possible that 5-hmC promotes local chromatin remodeling or serves as an epigenetic mark to recruit additional DNA damage repair machinery (Kafer et al. 2016).
2.3 TET Protein Structure and Function
TET genes are present in all metazoan genomes. In mammals and other gnathostomata, the TET gene underwent triplication to generate TET1, TET2, and TET3. During this process, TET2 likely underwent a local chromosome inversion causing the 5′ end of the inversion to create a distinct gene, CXXC4 (or IDAX) (Iyer et al. 2009; Ko et al. 2013). All three TET genes are transcribed and translated into catalytically functional TET enzymes.
All three TET proteins share a C-terminal catalytic domain, whereas TET1 and TET3 share an N-terminal CXXC domain (Fig. 2a). The catalytic domains consist of a cysteine-rich domain and a double-stranded β-helix (DSBH) domain, together forming a globular structure with the core DNA-binding DSBH in the center (Hu et al. 2013, 2015) (Fig. 2b). TET catalytic domains specifically recognize and bind CpGs and flip the modified cytosine base into the catalytic cavity, with no preference for flanking sequences (Hu et al. 2013) (Fig. 2b). This base flipping mechanism appears to be conserved evolutionally, as the same mechanism is found in the Naegleria gruberi Tet enzyme (Hashimoto et al. 2014, 2015).

TET protein domains and binding with DNA. (a) Human TET proteins. All three TETs have conserved C-terminal catalytic domain, which consists of a cysteine-rich domain (Cys) and two double-stranded beta-helix (DSBH) DNA binding domains. The two DSBH domains are separated by a low complexity (LC) insert. Both TET1 and TET3 have a CpG binding CXXC domain near the N-terminus. During evolution, the sequences encoding the CXXC domain of TET2 became an independent gene: CXXC4/IDAX. (b) The crystal structure of TET2:DNA binding (Hu et al. 2013). The TET2 catalytic domain forms a double-stranded beta-helix (DSBH, yellow) structure that binds DNA modified CpG (purple). The DSBH structure is stabilized by several surrounding alpha-helices (red). The modified cytosine base is flipped out from the DNA double-helix and inserted in the active site along with 2-oxoglutarate and oxygen. Protein Data Bank (PDB) ID: 4NM6 (Hu, L., Li, Z., Cheng, J., Rao, Q., Gong, W., Liu, M., Shi, Y.G., Zhu, J., Wang, P., and Xu, Y. (2013). Crystal structure of TET2-DNA complex: insight into TET-mediated 5mC oxidation. Cell 155, 1545–1555.), image created with Jmol (Jmol: an open-source Java viewer for chemical structures in 3D. http://www.jmol.org/)
TET proteins are 2-oxoglutarate (2OG) and oxygen (O2) dependent dioxygenases that use Fe(II) as a co-factor. TET catalytic domains share high homology to the previously discovered J-binding proteins (JBP1/2) (Iyer et al. 2009). JBP1/2 are dioxygenases in kinetoplastids that catalyze the oxidation of thymine at the methyl group to carboxyuracil, which is the first catalytic step towards base J. Based on this homology, the catalytic function of oxidizing 5-mC to 5-hmC was identified for the TETs (Tahiliani et al. 2009). Additional studies found that TETs can oxidize 5-hmC further to 5-fC and 5-caC in a stepwise fashion (He et al. 2011; Ito et al. 2011). In mammalian cells, the levels of 5-fC and 5-caC are 2–3 orders of magnitude lower than those of 5-hmC (Ito et al. 2011).
Enzyme kinetic studies showed that the K m values of TETs for oxygen, Fe(II), and 2OG are about 30 μM, 4 μM, and 60 μM, respectively (Laukka et al. 2015). This study also showed that TETs bind modified CpGs (5-mC, 5-hmC, or 5-fC) at around 100 nM. At a minimum, TET enzymes are two- to threefold more efficient at oxidizing 5-mC compared to 5-hmC or 5-fC due to differences in the conformation of the active sites (Hu et al. 2015), which leads to faster 5-hmC creation than its removal through further oxidation. The catalytic differences, together with TDG/NEIL1 mediated removal of 5-fC and 5-caC, may explain the relative abundance of 5-hmC over 5-fC and 5-caC.
The affinity of TETs for oxygen is much higher than that of the HIF prolyl-hydroxylases, which are 2OG/O2 dependent dioxygenases like TETs that hydroxylate hypoxia-inducible factor α (HIFα) to induce HIFα degradation (Laukka et al. 2015). Although both of these enzyme families use O2 as a substrate, the higher affinity for O2 in TETs suggests that the TETs may remain functional in hypoxic environments. Later studies showed that TETs retain catalytic activity under physiological hypoxia, at O2 concentrations at which the HIF prolyl-hydroxylases no longer function. The TETs may act with less efficiency or may lose their activities under the pathological hypoxia found in solid tumors (Thienpont et al. 2016).
2OG is produced from isocitrate by the isocitrate dehydrogenases (IDH1/2) in the TCA cycle. It is usually abundant in cells and is not limiting for TET activities. However, 2OG analogs can accumulate in pathological conditions as a result of mutations in genes encoding TCA cycle enzymes (Pollard et al. 2005; Dang et al. 2009). The IDH1 R100/R132 and IDH2 R140/R172 missense mutations commonly found in cancers catalyze the production of R-2-hydroxyglutarate (R-2-HG) (Dang et al. 2009; Losman and Kaelin 2013). R-2-HG competes with 2OG for TET binding, but cannot be used in the oxidation reaction and thus acts as an inhibitor of TETs (Xu et al. 2011a). Consistent with this finding, glioblastomas and leukemias with these IDH mutations display global hypermethylation (Figueroa et al. 2010; Ko et al. 2010; Turcan et al. 2012). In addition, IDH and TET mutations are almost always mutually exclusive in leukemias, strongly suggesting that the two enzymes function in the same cellular processes (Gaidzik et al. 2012; Patel et al. 2012). Two additional TCA cycle metabolites, fumarate and succinate, were found to accumulate in solid tumors with mutated fumarate hydratase (FH) or succinate dehydrogenase (SDH) (Pollard et al. 2005). Fumarate and succinate are also able to inhibit TET activity by competing with 2OG for TET binding (Laukka et al. 2015). This was demonstrated in SK-N-BE(2) neuroblastoma cells, where 5-hmC levels decreased 10–40% upon fumarate or succinate treatment (Laukka et al. 2015).
TET activity is enhanced by ascorbate (vitamin C) by maintaining the Fe co-factor in a reduced (II) state (Kuiper and Vissers 2014). Ascorbate is essential for the function of collagen prolyl-4-hydroxylase, another 2OG/O2 dependent dioxygenase similar to the TETs. Ascorbate deficiency causes failure of prolyl hydroxylation and leads to scurvy (Gorres and Raines 2010). The addition of ascorbate to cultured mouse ES cells or mouse embryonic fibroblasts leads to a rapid increase in 5-hmC levels that is Tet dependent (Blaschke et al. 2013; Minor et al. 2013; Yin et al. 2013). Other studies showed demethylation in ascorbate treated human ES cells and mouse embryonic fibroblasts as well as significant changes in the transcriptome (Chung et al. 2010; Blaschke et al. 2013; Chen et al. 2013).
3 Normal Transcription of TET Genes and Its Impact on the 5-hmC Landscape
Highly regulated expression of TET genes is essential for proper maintenance of the 5-hmC landscape in steady state or epigenetic reprogramming. Despite the functional redundancy, expression of each TET gene is distinct depending on the developmental context and tissue type, and disrupted expression patterns can give rise to pathogenic phenotypes. Here, we highlight the regulation of TET expression and its impact on 5-hmC by examining ES cells and adult neuronal cells, two cell types where 5-hmC is highly abundant and undergoes dynamic changes. It should be noted that the majority of embryonic development studies were done in murine systems, but there are supporting data obtained from studies in human cell lines. The species differences are denoted by mouse Tet and human TET.
3.1 TET Transcription and TET Protein Regulation of 5-hmC in Embryonic Development
Embryonic development is characterized by extensive epigenetic remodeling. Immediately after fertilization, 5-mC in the paternal zygotic genome is oxidized to 5-hmC and is further converted to unmodified cytosine through the demethylation pathways (Fig. 1a) (Mayer et al. 2000; Oswald et al. 2000). Mammalian ES cells have high levels of 5-hmC, which is enriched in enhancers, gene bodies, exons, transcription start sites, and 5′-untranslated regions of genes (Tahiliani et al. 2009; Pastor et al. 2011; Stroud et al. 2011; Wu and Zhang 2011). It is also enriched in the promoters of genes that feature both H3K27me3 and H3K4me3 marks, which are associated with open chromatin (Pastor et al. 2011; Stroud et al. 2011; Wu and Zhang 2011). These enrichments are often associated with elevated gene transcription (Ficz et al. 2011). Furthermore, throughout the development of the embryo, there are dynamic gains and losses of 5-hmC at specific regions of the genome, especially in the primordial germ cells (PGCs). Murine PGCs undergo their own demethylation events, between e9.5 and e10.5, in which there is global erasure of 5-mC (Hajkova et al. 2002; Seki et al. 2005). The TET enzymes are critical for 5-hmC changes in embryonic development and PGCs, as loss of all three TET genes is lethal and loss of one or combinations of two has distinct phenotypes (Ficz et al. 2011; Wu and Zhang 2011). During epigenetic remodeling of the developing embryo, expression of TET1, TET2, and TET3 is tightly controlled.
3.2 Normal Tet1 Transcription in ES Cells and PGCs
In mammalian ES cells, normal TET1 expression is very high and plays a role in maintaining the pluripotent state (Koh et al. 2011). TET1 expression in ES cells is promoted by canonical ES factors, such as OCT3/4, MYC, and NANOG, and is reduced when these factors are absent (Koh et al. 2011; Neri et al. 2015). TET1 expression decreases as cells differentiate (Neri et al. 2015). The decrease in expression is mediated by the polycomb complex, which epigenetically silences the transcription start sites of TET1 through deposition of the repressive histone mark, H3K27me (Neri et al. 2015). Although it is known that Tet1 is expressed as early as e9.25 in PGCs, it is not known what transcription factors control expression of Tet1 (Vincent et al. 2013).
Truncated TET1 RNAs have been identified in mouse and human embryonic systems. In mouse ES cells, there is a Tet1 isoform switch after e14.5. Full length Tet1 is expressed in early embryos, ES cells, and PGCs, but only a Tet1 RNA that does not code for part of the N-terminus, including the CXXC domain (Tet1s), is expressed in adult tissues (Zhang et al. 2016). Despite having a lower chromatin affinity, Tet1s is still capable of binding DNA and its binding pattern is similar to that of full-length Tet1 (Zhang et al. 2016). Nonetheless, when homozygous Tet1s male mice were crossed with WT female mice, the resulting pups featured higher levels of perinatal lethality and lower birth weights compared to pups from WT crosses (Zhang et al. 2016). These experiments indicate that full-length Tet1 is important for normal embryonic development in mice, but not necessary in adult mice. The regulation of 5-hmC by Tet1s has yet to be studied in mice. A similar mechanism has been hypothesized in human systems: full-length TET1 is present in ES cells, and an alternate truncated protein that lack the CXXC domain is present in adult tissues (Good et al. 2017). This smaller isoform of TET1 has been implicated in many cancers, including breast, uterine, and ovarian cancers (Good et al. 2017). This isoform is catalytically active, but does not have as great an impact on gene expression as the full-length TET1 (Good et al. 2017). The mechanism behind this is unclear, although it has been shown that the TET1 short-isoform binds outside of CpG islands, a distinction from full-length TET1 (Good et al. 2017).
3.3 Loss of Tet1 Disrupts 5-hmC Levels and Distribution in ES Cells and PGCs
Loss of Tet1 in ES cells has a measurable impact on total levels and distribution of 5-hmC. When Tet1 was knocked down in mouse ES cells, there is a decrease of about 35–50% of total 5-hmC levels (Tahiliani et al. 2009; Dawlaty et al. 2011; Hon et al. 2014). Correspondingly, there is an increase in total 5-mC levels (Ito et al. 2010; Dawlaty et al. 2011). Loss of 5-hmC in Tet1 knock-down (KD) ES cells has been localized to promoters, gene bodies, and a few intergenic regions (Wu and Zhang 2011; Huang et al. 2014). Similarly, Tet1 KD in PGCs results in an increase in global 5-mC levels and a 45% reduction in 5-hmC (Yamaguchi et al. 2012).
In addition to aberrant cytosine modification patterns, loss of Tet1 in ES cells and PGCs leads to deregulation of gene expression (Dawlaty et al. 2011). Specifically, there is reduced expression of pluripotent factors Nanog, Oct4, and Sox2, as well as other genes associated with pluripotency (Ito et al. 2010; Ficz et al. 2011). In PGCs, loss of Tet1 was found to reduce expression of some meiotic genes (Yamaguchi et al. 2012).
Loss of Tet1 in ES cells and PGCs results in reduced growth and up-regulation of differentiation-associated genes (Ito et al. 2010; Pastor et al. 2011). When mouse ES cells that lack Tet1 undergo differentiation, there is upregulation of genes encoding primary trophectoderm factors (Cdx2, Eomes, and Elf5) and downregulation of genes encoding neuroectoderm factors (Pax6 and NeuroD2) (Ito et al. 2010; Dawlaty et al. 2011; Koh et al. 2011; Xu et al. 2011b). Further, as mentioned previously, depletion of Tet1 in PGCs leads to defective meiotic prophase that includes DSBs and formation of univalent chromosomes (Yamaguchi et al. 2012).
Tet1 KO mouse ES cells are able to differentiate and form living mice (Dawlaty et al. 2011). However, a number of defects have been reported in mice that lack Tet1 (Dawlaty et al. 2011; Rudenko et al. 2013; Zhang et al. 2013; Kumar et al. 2015). Some Tet1 deficient mice have a smaller body size compared to WT (Dawlaty et al. 2011). Mice that lack Tet1 have memory extinction impairment and abnormally enhanced hippocampal long-term depression, indicating that the downregulation of neuroectoderm factors impacts normal brain function (Rudenko et al. 2013). Others have reported impaired spatial memory, object location memory, and threat recognition memory in Tet1 deficient mice (Zhang et al. 2013; Kumar et al. 2015). The meiosis defects mentioned above in mouse PGCs lead to loss of oocytes and consequent decrease in mouse litter size (Yamaguchi et al. 2012).
3.4 Loss of Tet2 Transcription and Its Impact in ES Cells and PGCs
Tet2 is expressed about fivefold less than Tet1 in ES cells, and its expression has been shown to be under the control of Oct4 (Koh et al. 2011; Sohni et al. 2015). Loss of Tet2 results in about a 50–90% reduction in 5-hmC (Koh et al. 2011; Hon et al. 2014). This loss of 5-hmC in Tet2 deficient cells is localized to the gene bodies and exon-intron boundaries of highly expressed genes (Huang et al. 2014). An additional report found Tet2 deficient cells to have increased 5-hmC around the Nanog promoter, which is believed to be the result of compensatory Tet1 expression (Huang et al. 2014; Langlois et al. 2014). Similar to Tet1 deficient cells, there is an increase in 5-mC in reaction to the loss of Tet2, but specifically at enhancers under this condition (Hon et al. 2014).
Overall, loss of Tet2 has a less dramatic impact on downstream gene expression than the loss of Tet1 (Koh et al. 2011; Huang et al. 2014). Broadly, there is delayed gene induction in Tet2 deficient ES cells during differentiation (Hon et al. 2014). In one study, there was only a modest increase in neuroectoderm markers Pax6, NeuroD1, Lefty1, and Lefty2, the opposite effect of Tet1 KD (Koh et al. 2011). When Tet2 deficient cells were allowed to differentiate, there was a bias towards neuroectoderm differentiation in one mouse and one human model (Koh et al. 2011; Langlois et al. 2014). However, according to another group, loss of Tet2 has no impact on gene expression or ES cell morphology (Ito et al. 2010). Differentiation of Tet2 deficient cells resulted in a decrease in both number and clonogenic capability of hematopoietic progenitor cells (Langlois et al. 2014). In PGCs, Tet2 is heterogeneously expressed at e9.5 and e10.5 and is generally regarded as redundant with Tet1 (Hackett et al. 2013; Vincent et al. 2013). When Tet1 and Tet2 are silenced together, demethylation of genes in PGCs is inhibited (Hackett et al. 2013).
The evidence for the roles of Tet2 in ES cells is not always consistent across studies. First, as previously stated, the loss of Tet2 has highly variable impacts on the level of 5-hmC (50–90% reduction) (Koh et al. 2011; Hon et al. 2014). There are also differing reports on the impact of Tet2 on gene expression. Loss of Tet2 promoted the expression of neuroectoderm markers in two studies (Koh et al. 2011; Langlois et al. 2014), but two other groups found that loss of Tet2 negatively regulated Lefty1 and had no impact on global gene expression (Ito et al. 2010; Hon et al. 2014). In addition, Tet2 is frequently studied within the context of Tet1 loss in embryonic development (Ficz et al. 2011; Koh et al. 2011; Vincent et al. 2013). Few studies have looked at the role of Tet2 independent of Tet1 in ES cells and PGCs.
3.5 Tet3 Transcription and Its Epigenetic Role in the Zygote
Tet3 expression is essential for normal 5-hmC epigenetic remodeling of the zygote, but is nearly undetectable during very early embryonic development before the gene gets re-expressed in specific adult tissues (Gu et al. 2011; Koh et al. 2011; Wossidlo et al. 2011). Tet3 expression is high in oocytes and zygotes and is significantly decreased in two-cell embryos (Gu et al. 2011; Wossidlo et al. 2011). When Tet3 is knocked down, the conversion of 5-mC to 5-hmC in the paternal DNA is abrogated, and there is little 5-hmC (Gu et al. 2011; Wossidlo et al. 2011). Consequently, the demethylation of key paternal embryonic genes (Oct4, Lemd1, and Nanog) is impaired (Gu et al. 2011). The pattern of 5-hmC enrichment has yet to be investigated at other regions of the genome. As a result, the Oct4 gene activation is delayed in the early embryo (Gu et al. 2011). The loss of Tet3 from the maternal genome causes developmental failure in mouse embryos, due to retention of paternal 5-mC (Gu et al. 2011).
Although Tet3 expression is low in ES cells, it increases during neuronal differentiation (Li et al. 2015b). Tet3 KO cells are capable of expressing neuronal differentiation markers but often undergo apoptosis (Li et al. 2015b). Tet3 is also critical for Xenopus eye and neural development (Xu et al. 2012).
3.6 Loss of Multiple Tets Has Serious Functional Consequences in Embryonic Development
Double and triple KO (DKO and TKO) of TET genes in embryonic systems has dramatic effects on 5-hmC maintenance and cell phenotype. When Tet1 and Tet2 are silenced together, there is loss of 75–80% of 5-hmC in ES cells, greater than those of the effects of either single KD (Koh et al. 2011). Tet3 is upregulated in the double silenced cells, presumably to compensate the loss of Tet1 and Tet2 (Koh et al. 2011). In Tet1/Tet2 DKO cells, the changes in gene expression are similar to Tet1 KD alone, suggesting that Tet1 has a dominant role on transcription over Tet2 in ES cells (Koh et al. 2011). During differentiation of mouse embryos, Tet1/Tet2 DKO cells remain depleted of 5-hmC, and there are developmental defects in chimeric embryos consistent with perinatal lethality (Dawlaty et al. 2013). The defects Tet1/Tet2 DKO embryos exhibit include exencephaly, hemorrhages in the head, or profound growth retardation. However, some Tet1/Tet2 DKO mice are viable (Dawlaty et al. 2013). Although both sexes of the DKO mice are fertile, females have smaller ovaries and reduced fertility (Dawlaty et al. 2013).
Tet1/Tet3 DKO in embryonic systems also has severe biological consequences, including abnormal mitosis and nuclear blebbing in the two-cell stage (Kang et al. 2015). Although not well studied in ES cells, gestation of Tet1/Tet3 DKO mouse embryos had been assessed in one study (Kang et al. 2015). In these embryos, 5-hmC was undetectable by antibody staining, and only about 70% of them survived to eight-cell embryos. By e3.5, 60% of the remaining embryos had aberrant Nanog expression. During the entire process, the transcriptomes of the embryos featured very high variability. Ultimately, embryos were not viable beyond e10.5.
tet2/tet3 DKO has only been examined in the context of embryonic development in zebrafish (Li et al. 2015a). In this model, tet2 and tet3 are the predominant 5-hmC regulators in embryos. tet2/tet3 DKO resulted in a 30-fold loss of 5-hmC, although the distribution of 5-hmC was not characterized. These embryos featured abnormal differentiation, including altered brain morphology, smaller eyes, and a lack of a definitive hematopoietic stem cell population, and ultimately did not survive beyond the larval stage. Gene expression was not assessed, so it is not known how loss of tet2 and tet3 impacted the transcriptome of these animals.
Tet1/Tet2/Tet3 TKO is invariably lethal and leads to complete depletion of 5-hmC in ES cells (Dawlaty et al. 2014; Hu et al. 2014; Lu et al. 2014). Promoters and enhancers are hypermethylated, and many genes are deregulated (Lu et al. 2014). However, studies disagree about whether the majority of genes are upregulated or downregulated in TKO cells (Dawlaty et al. 2014; Lu et al. 2014). One study found that when the TKO cells underwent differentiation, there was reduced expression of mesodermal and endodermal markers (Dawlaty et al. 2014). In addition, the cells formed fewer differentiated structures. When injected into embryos, there was a lower incidence of chimeric embryos formed from TKO cells compared to controls, indicating that the Tets are essential for normal embryonic development.
3.7 Cocaine Stimulation Remodels the 5-hmC Landscape Through Tet1 Expression
The brain is particularly enriched with high levels of 5-hmC (Kriaucionis and Heintz 2009; Globisch et al. 2010), where the base is thought to play a major role in neuronal adaptation to stimuli, which requires epigenetic remodeling. Once considered static epigenetic marks, we now appreciate that covalent cytosine modifications in the brain are reversible and dynamic, with frequent remodeling of 5-mC and 5-hmC, due in part to TET1 expression (Reik 2007; Guo et al. 2011a, b). Changes to the 5-hmC landscape occur in response to neuronal activity, indicating specific stimuli have the potential to remodel 5-hmC level and distribution (Guo et al. 2011a). One such stimulus that has been studied within the context of brain tissue is cocaine exposure (Feng et al. 2015). This study found repeated cocaine administration resulted in downregulation of Tet1 expression and increased 5-hmC levels within the nucleus accumbens of mice. The increases of 5-hmC were enriched around the enhancers and gene bodies of genes known to be involved in drug addiction. The downregulation of Tet1 and altered 5-hmC appeared to enhance behavioral responses to cocaine. Nonetheless, the mechanism of how Tets are regulated in the brain is yet to be identified. Despite being a tissue enriched in 5-hmC, very little is known about the expression and function of the Tets in this system.
Epigenetic reprogramming of the 5-hmC landscape is highly dependent on rigorous regulation of expression from the TET genes. It is clear that each TET enzyme has a unique role in each process, and the loss of one enzyme can only be partially compensated by the other two, indicating non-overlapping functions.
4 Mutations of TET Genes in Cancers
Epigenetic abnormalities such as global hypomethylation and site-specific hypermethylation have been known to be characteristics of cancers since the 1980s (Feinberg and Vogelstein 1983; Gama-Sosa et al. 1983; Baylin et al. 1986; Feinberg and Tycko 2004). Since the discovery of TET function, the role of 5-hmC in pathologic conditions has become a new focus in epigenetics. However, many techniques used to study 5-mC, including sodium bisulfite treatment and several methylation-sensitive restriction enzymes, cannot distinguish 5-mC from 5-hmC. For this reason, many interpretations of ‘methylcytosine’ should be more properly described as ‘5-mC or 5-hmC’. The recent development of 5-hmC specific techniques (Table 1), such as TAB-seq (Yu et al. 2012) and hMe-SEAL (Song et al. 2011), allow specific detection of 5-hmC. Through the use of these new techniques, widespread 5-hmC alterations were found in blood and solid tumor cancers (Vasanthakumar and Godley 2015). Here, we will discuss some of the most common causes of 5-hmC deregulation in cancer, including mutations of TETs and changes in TET expression.
4.1 TET1 Mutations and Transcriptional Changes in Cancer
The TET1 gene was originally discovered as a fusion partner of KMT2A (MLL) in cases of t(10;11)(q22;23) acute myeloid leukemia (AML) (Ono et al. 2002; Lorsbach et al. 2003). MLL encodes a histone methyltransferase, and leukemia-specific chromosomal fusions involving this gene frequently result in production of MLL-fusion proteins. The MLL-TET1 fusion is found in about 0.3% of all MLL-fusion leukemias, resulting in the fusion of the N-terminus of MLL with the C-terminal catalytic domain of TET1 (Lee et al. 2013; Meyer et al. 2013). The MLL-TET1 fusion has also been found in some acute lymphoblastic leukemia (ALL) cases and has the capability to drive a phenotypic switch from ALL to AML (Burmeister et al. 2009; Ittel et al. 2013). MLL-TET1 fusion causes translocation of the TET1 catalytic domain, which is normally absent in myeloid systems, therefore promoting AML progression (Huang et al. 2013). In fact, TET1 is a direct target of all MLL-fusion proteins and is found to be significantly upregulated in MLL-rearranged leukemia samples compared to normal bone marrow samples (Huang et al. 2013). This alteration results in increased global level of 5-hmC (Huang et al. 2013, 2016). However, genomic enrichment of 5-hmC in MLL-rearranged leukemia cells has yet to be studied.
TET1 mutations not involving chromosomal fusion are rare in hematopoietic malignancies, especially compared to TET2 mutations. A study of 83 patients revealed only 2 cases with TET1 mutations in myelodysplastic syndrome (MDS) or chronic myelomonocytic leukemia (CMML) patients (Lasho et al. 2018) (Fig. 3), but it remains unclear whether these are driver or passenger mutations.

Mutation effects in TET proteins. TET1 and TET3 are rarely mutated in human diseases. Most of the mutations identified in TET1 and TET3 are missense mutations or in-frame insertion/deletion (indel), leading to single amino acid substitutions in the protein. All of these mutations occur outside of the core catalytic domain or the CXXC domain, likely maintaining the catalytic function of the protein. A notable exception is a nonsense mutation in TET3, which truncates the protein before the catalytic domain, leading to complete loss of the catalytic domain in the protein. Numerous TET2 mutations have been described over the last decades, most of which lead to loss of function in TET2 protein. In contrast to TET1 and TET3 mutations, TET2 missense mutations often directly impact the catalytic domain and cause impaired catalytic activities. Nonsense and frameshift mutations are commonly found throughout the entire coding sequence, also leading to loss of TET2 catalytic activity
The role of TET1 in the pathogenicity of cancers is usually linked to its gene expression. The 5-hmC levels in solid tumors are often low compared to surrounding normal tissue in solid tumors, including liver, brain, kidney, lung, colorectal, and gastric cancers (Kudo et al. 2012; Vasanthakumar and Godley 2015). Knock-down of Tet1 alone in 3T3 fibroblasts is sufficient to replicate the 5-hmC loss seen in tumors, but not enough to induce malignant transformation (Kudo et al. 2012). Although many of the earlier studies on 5-hmC did not provide mechanistic links between 5-hmC alterations and pathogenic phenotypes, more recent studies began to provide better insight into the roles of 5-hmC regulation in cancers.
For example, we have recently gained insight in how TET1 is involved in hypoxic responses in certain solid tumors. When MYCN-amplified neuroblastoma cell lines are exposed to hypoxia, 5-hmC levels increase (Mariani et al. 2014). This result was surprising given, as discussed earlier in this chapter, TETs rely on oxygen for activity. The increased 5-hmC was the result of hypoxia-induced expression of TET1. In hypoxia, 5-hmC was enriched along the gene bodies of direct targets of HIF-1, the master regulator of hypoxia. These HIF-1 targets are known as canonical hypoxic response genes. Upon loss of HIF-1α, TET1 expression no longer increased, and there was no induction of 5-hmC under hypoxic conditions. Loss of hypoxia induced-TET1 resulted in decreased expression from canonical hypoxic response genes in comparison to hypoxic controls, demonstrating TET1 facilitates expression from canonical hypoxic response genes in hypoxia (Mariani et al. 2014).
4.2 TET2 Mutations and Transcriptional Changes in Cancer
TET2 is the most commonly mutated gene among the three TET genes (Fig. 3). In hematologic malignancies, TET2 mutations are found in 20–30% of AML, 20–30% of MDS or myeloproliferative neoplasm, and up to 58% of CMML (Delhommeau et al. 2009; Jankowska et al. 2009; Langemeijer et al. 2009; Yamazaki et al. 2012, 2015; Itzykson et al. 2013). TET2 mutations are also found in other types of cancers at lower frequencies: 12% of T-cell ALL (Quivoron et al. 2011), 7% of clear cell renal cell carcinoma (clear cell RCC) (Sato et al. 2013), 12% of stage IV non-small cell lung cancer (Jin et al. 2016), and in metastatic prostate cancer (Nickerson et al. 2013). Copy number alterations of TET2 are found in about 5.6% of various hematologic malignancies, 70% of which also have cytogenetic alterations (Bacher et al. 2012). Notably, most in-frame deletions and missense mutations occur in exons that code for the C-terminal catalytic domain, whereas nonsense and frameshift deletions can occur along the entire gene (Fig. 3) (Langemeijer et al. 2009; Euba et al. 2012). Most of the known mutations result in truncated protein or impaired catalytic functions (Langemeijer et al. 2009; Euba et al. 2012). One study found that many of the leukemia associated missense mutations in the catalytic domain of TET2 reduced the affinity of the enzyme for Fe(II) and 2OG and greatly reduce its activity (Laukka et al. 2015).
The effect of TET2 mutation on 5-hmC and 5-mC has been studied, but not thoroughly. It is clear that TET2 mutated leukemia cells have globally decreased 5-hmC (Ko et al. 2010; Madzo et al. 2014), but the mechanism behind this loss is not understood. Early studies of TET2 mutated samples found no global difference in 5-mC, but showed site-specific hyper- and hypomethylation (Ko et al. 2010). In contrast, other studies have shown global hypermethylation in leukemic cells (Madzo et al. 2014). It should be noted that Ko et al. measured global methylation using the Illumina Infinium 27K array that probes 27,578 selected CpG sites, whereas Madzo et al. used mass spectrometry that measure all nucleotides in the genome, which may explain the discrepancy in these findings. Curiously, in this study, distribution of 5-hmC in TET2-mutated leukemia is dramatically different from normal, which is not explained solely by the global decrease of 5-hmC. Although most 5-hmC peaks decrease correspondingly with the global decrease of 5-hmC, certain sites gain high density of 5-hmC not seen in normal control. This redistribution process suggests that TET1 or TET3 may compensate for the loss of TET2, resulting in skewed 5-hmC distribution. Lastly, in CMML cells, TET2 mutation-induced cytosine modification changes, especially hyper-modifications, are enriched in enhancer regions and negatively correlate with gene expression changes (Meldi et al. 2015; Yamazaki et al. 2015). Note that these studies used bisulfite sequencing based methods to characterize genome-wide cytosine modification changes, which cannot distinguish 5-mC from 5-hmC. Using modification status from as few as 14 sites, a support vector machine model was able to achieve 79% accuracy in predicting patient response to decitabine, a hypomethylating agent commonly used to treat CMML (Meldi et al. 2015). This result suggests that the changes in cytosine modifications have prognostic value to risk-stratify patients at diagnosis.
In addition to TET2 mutations, it is becoming increasingly clear that TET2 expression is a target of microRNAs in blood cancers (Cheng et al. 2013; Song et al. 2013). TET2 has been found to be downregulated by miR-22, miR-29b, miR-125a, miR-26, miR-101, and miR-520d. 5-hmC levels were found to be decreased when TET2 was down-regulated by miR-22, miR-29b, or miR-125a. Some of these microRNAs were also shown to regulate TET1 and TET3, but an effect on 5-hmC level or distribution was not demonstrated (Cheng et al. 2013). Expression of miR-22, miR-29b, or miR-125a increases the oncogenic traits of cells, including increased replating ability of hematopoietic stem cells. In addition, mouse models with overexpression of these miRs featured development of MDS, and splenomegaly (Cheng et al. 2013; Song et al. 2013).
Although TET2 has been investigated mostly in the context of blood cancers, there have been some instances of abnormal TET2 expression in solid tumors. Low 5-hmC levels correlate with down-regulated TET2 expression in oral cell carcinoma and esophageal squamous cell carcinoma (ESCC) (Jäwert et al. 2013; Shi et al. 2016). In both tumors, low 5-hmC was associated with low TET2 expression. ESCC tumors have significantly decreased TET2 and TET3 expression compared to a normal tissue control (Shi et al. 2016). It is unknown how downregulation of TET2 and low 5-hmC impact tumor oncogenicity, but loss of 5-hmC is an unfavorable prognostic factor in ESCC (Shi et al. 2016).
4.3 TET3 Mutations and Transcriptional Changes in Cancer
TET3 mutations are rare compared to TET2 mutations in cancer. A previous study with a cohort of 408 leukemia patients did not find any TET3 mutations (Abdel-Wahab et al. 2009). More recently, two loss-of-function mutations were identified in TET3 in the bone marrow of 28 CMML patients, both coexisting with TET2 mutations (Merlevede et al. 2016) (Fig. 3). In addition, another study identified seven TET3 mutations in a study of 83 leukemia patients (Lasho et al. 2018) (Fig. 3). None of the newly identified mutations co-occurred with TET2 mutations. Six of these mutations were found in CMML patients (16% of all CMML patients), two of which were found in the same patient, and one of which co-occurred with a previously mentioned TET1 mutation (Lasho et al. 2018). These findings suggest that TET3 mutations preferentially occur in CMML, although the significance and consequences of TET3 mutations remain to be studied.
There are emerging data that TET3 expression is associated with an aggressive phenotype in RCC and head and neck cancer (Chen et al. 2017a; Misawa et al. 2018). High expression of TET3 in RCC is associated with poor survival, but the effect on the 5-hmC landscape is unknown (Chen et al. 2017a). TET3 is expressed at low levels in head and neck cancer, but perhaps more interestingly, the cytosine modification status of its promoter is negatively associated with disease survival (Misawa et al. 2018). The mechanism behind this finding is not yet known.
4.4 Deregulated Expression of Multiple TETs in Cancer
Many other cancers with low 5-hmC levels feature low expression or no change in expression of all three TET enzymes, indicating a more complex mechanism behind the aberrant 5-hmC landscape. Low levels of 5-hmC have been identified in brain, breast, liver, lung, pancreatic, prostate, and skin tumors (Jin et al. 2011; Lian et al. 2012; Yang et al. 2013). Breast, liver, and skin cancers all demonstrate low expression of all three TET enzymes (Lian et al. 2012; Yang et al. 2013; Chen et al. 2017b). The mechanism of low TET expression has only been explored in liver cancer. Low 5-hmC correlated with metastasis and poor prognosis of hepatocellular carcinoma (HCC) (Chen et al. 2017b). Expression from all three TET enzymes was downregulated by elevated expression of miR-29a, which in turn, resulted in decreased 5-hmC and increased 5-mC levels. The promoter of tumor suppressor gene SOCS1 was found to be hypermethylated, and cell motility was increased.
In contrast, there has been no reported change in expression in any of the TETs in lung and brain cancer (Jin et al. 2011). Tumors that arise from pancreatic and prostate tissue have featured low levels of 5-hmC, however TET expression level has yet to be investigated.
Correcting altered transcription from TET genes is an attractive goal because it has the potential to ‘reset’ the 5-hmC landscape to pre-cancerous status. Further work to identify mechanisms of TET transcription regulation is required to achieve this goal.
5 Post-Translational Modifications (PTMs) of TET Enzymes
TET proteins are subject to an assortment of PTMs, many of which are poorly understood functionally. However, TET PTMs might reconcile a major discrepancy in studies concerning the DNA binding properties of TETs. There is conflicting research into the binding of TET to DNA: some studies have found that TET does not bind DNA at all, and others have found TETs can bind sequences containing unmodified, methylated, and hydroxymethylated cytosine (Zhang et al. 2010; Frauer et al. 2011b; Xu et al. 2011b). Preliminary research indicates that PTMs play a role in stability, activity, and localization of the TET enzymes. Considering the role of TETs in the pathogenesis of diseases, it is likely the role of TET PTMs and their effects on any aberrant activity will become a target for more studies. Known PTMs of the TET proteins include phosphorylation, O-GlcNAcylation (O-GlcNAc), acetylation, and ubiquitination (Fig. 4) (Bauer et al. 2015; Nakagawa et al. 2015; Zhang et al. 2017).

General distribution of TET PTMs. (a) Schematic representation of the common TET structure featuring modifications that are consistent across all three TET enzymes. A phosphorylation and O-GlcNAcylation-rich region is comprised of a large number of phosphate and O-GlcNAc marks that are variably distributed. A monoubiquitination modification is also found in the cysteine-rich domain of all three TET enzymes. (b) General distribution of modifications unique to each TET protein. Colors: Red: Acetylation, Orange: Ubiquitination, Blue: Phosphorylation, Green: O-GlcNAcylation
5.1 Phosphorylation of TET Proteins
A study performed in mouse and human systems indicated phosphorylation of the three TET proteins largely occurs in the N-terminus (Fig. 4a) (Bauer et al. 2015). This modification can occur singly or in tandem with another modification. For example, in TET2, phospho-Ser-23 is found only with phosphor-Ser-15. The kinase(s) responsible for the phosphorylation modifications on the TET proteins are largely unknown at this time. Recent studies are starting to reveal the pathways regulating TET2 phosphorylation. For example, a specific site on TET2, Ser-99, was found to be phosphorylated by AMP-activated kinase (AMPK) (Wu et al. 2018). TET2 with this phosphorylation mark was found to have a longer half-life than TET2 without this mark. A Ser-99-Asp phospho-mimic TET2 was found to increase 5-hmC levels when expressed. However, distribution of 5-hmC was not investigated. Further research on TET2 phosphorylation will shed light on other kinases that modulate TET2 activity and stability. However, because modifications are enriched in the non-catalytic domains, it is hypothesized that TET phosphorylation may also play a role in interaction with TET binding partners (Bauer et al. 2015; Liu et al. 2018).
5.2 O-GlcNAc Modifications Facilitate TET Protein Stability, Activity, and Localization
O-GlcNAc and phosphorylation modifications are occasionally in competition with each other (Bauer et al. 2015). Specifically, Ser-97 and Ser-374 of TET2 and Ser-362 and Ser-557 of TET3 could be phosphorylated or O-GlcNAcylated. Overall, some TET phosphorylation marks are stable, such as Ser-950 of TET1, but some are lost when OGT is highly expressed, such as Ser-2016 of TET1. OGT encodes the enzyme O-linked N-Acetylglucosamine Transferase (OGT) which is responsible for the addition of an O-GlcNAc group to serine and threonine. Similar to phosphorylation modifications, O-GlcNAc modifications are generally localized to the N-terminus and the low complexity insert (Fig. 4a, b). However, there is some distinction of the O-GlcNAc modification patterns between the three TET proteins (Fig. 4b). TET1 features fewer O-GlcNAc modifications than TET2 or TET3, with most found in the N-terminus and hardly any in the C-terminal region. In addition, O-GlcNAc modifications of TET1 have been shown to be highly dynamic (Shi et al. 2013; Bauer et al. 2015). TET2 and TET3 display most O-GlcNAc in the low complexity insert region in one comprehensive study (Bauer et al. 2015). In addition, the first 350 amino acids of TET3 are unmodified. In mouse and human systems, there appears to be crosstalk between the phosphorylation and the O-GlcNAc modifications of the TET proteins. Although increase in OGT expression is associated with decreased phosphorylation in TETs, phosphate and O-GlcNAc groups are rarely found on the same residue. Instead of direct competition, it appears that neighboring residues are engaging in crosstalk as several modifications of the same type often occur in close proximity. There have been many proposed functions of TET O-GlcNAc modifications (Vella et al. 2013; Bauer et al. 2015; Hrit et al. 2018). Several studies have demonstrated that this modification increases Tet1 protein stability and activity (Vella et al. 2013; Hrit et al. 2018). Others have implicated O-GlcNAc groups in mediating binding of TET partners and localization (Bauer et al. 2015). Studies in mouse embryonic development have shown that loss of O-GlcNAc modifications on Tet3 resulted in nuclear rather than cytosolic subcellular localization of the protein and substantially increased global 5-hmC levels (Zhang et al. 2014). Interestingly, O-GlcNAcylation of Tet1 and Tet2 had no impact on their respective subcellular localizations (Zhang et al. 2014).
5.3 Acetylation Has Been Implicated in the Stability of TET2
Acetylation has been identified as a modification of only TET2 in human cell lines. One study found TET2 is acetylated by the epigenetic modifier p300 (Zhang et al. 2017). TET2 is acetylated at multiple lysines in the N-terminus, and the presence of two (K110 and K111) are necessary for p300-mediated acetylation (Fig. 4b). Although there is no accumulation of TET2 protein, global 5-hmC level increases in the presence of acetylated TET2, indicating that acetylation of TET2 enhances its enzymatic activity. The acetylation marks were also found to inhibit polyubiquitination of TET2, therefore promoting its stability. Lastly, TET2 acetylation has also been implicated in potential binding of DNMT1.
5.4 TETs Are Monoubiquitinated
Monoubiquitination of all three TET proteins occurs on a lysine, mediated by VprBP, in the highly conserved cysteine-rich domain (Fig. 4a) (Nakagawa et al. 2015). In vivo ubiquitination assays with TET proteins yielded a single band on a gel, indicating addition of a one ubiquitination group. The monoubiquitination occurs on K1589 in TET1, K1299 in TET2, and K859 in TET3. Loss of monoubiquitination prevents binding of TETs to DNA and therefore results in an overall loss of 5-hmC in in vitro and in vivo experiments. Several oncogenic mutations of TET2 disrupt its ubiquitination, indicating this modification is important for normal TET2 activity.
6 Proteins Interact Directly with Modified Cytosine and TET Binding Proteins
Stable epigenetic marks interact with epigenetic “reader” proteins that mediate their downstream effects. Although there is some overlap between binding partners, 5-mC, 5-hmC, and TETs each have their own unique set of interactors.
6.1 Proteins Recognize, Bind, and Read 5-mC and 5-hmC
Identified readers of 5-mC all contain the conserved methyl-binding domain, but not all proteins that feature a methyl-binding domain bind 5-mC (Hendrich and Tweedie 2003). Most proteins that read 5-mC are thought to reinforce the repression of genes with 5-mC rich promoters (Boyes and Bird 1991; Nan et al. 1997; Hendrich and Bird 1998). In fact, it is hypothesized that the ability to bind 5-mC may be cell type specific (Iurlaro et al. 2013). Therefore, this section will only discuss the most universal 5-mC readers. The very first proteins identified that read 5-mC were MeCP1 and MeCP2 (Meehan et al. 1989; Lewis et al. 1992). Although it was originally thought that 5-mC prevents transcription exclusively by directly blocking the binding of transcription factors, it is now known that MeCP1, MeCP2, and MBD1/2/4, are all indirect mediators of the biological function of the 5-mC mark by recruiting chromatin remodeling machinery such as NuRD complex to downregulate gene expression (Boyes and Bird 1991; Nan et al. 1997; Hendrich and Bird 1998; Hendrich and Tweedie 2003; Pan et al. 2017).
Researchers have tried to identify proteins that mediate the biological function of 5-hmC using a variety of screening methods (Iurlaro et al. 2013; Spruijt et al. 2013). Unfortunately, many studies have been unable to replicate results presented by other groups. This may be due to a lack of rigorous testing across multiple model systems, but also may indicate many 5-hmC readers are cell type specific, implying a more complex mechanism at work than simple hydroxymethylcytosine recognition (Iurlaro et al. 2013). Some proposed 5-hmC readers include RBM14, PRP8, RPL26, MSH6, PNKP, Wdr76, and Thy28. However, each of these has only been identified in a single study, and the findings remain to be validated (Hendrich and Bird 1998; Frauer et al. 2011a; Yildirim et al. 2011; Mellén et al. 2012; Iurlaro et al. 2013; Spruijt et al. 2013). Only three proteins, NP95/UHRF1, MeCP2, and MBD3, have been confirmed in more than one study (Sharif et al. 2007; Arita et al. 2008; Frauer et al. 2011a; Yildirim et al. 2011; Mellén et al. 2012; Baubec et al. 2013; Iurlaro et al. 2013; Spruijt et al. 2013). NP95/UHRF1 has been identified as a 5-hmC reader in three studies in two cell-free assays and in mouse ES cells (Frauer et al. 2011a; Iurlaro et al. 2013; Spruijt et al. 2013). Uhrf1 is traditionally recognized as a binder of hemimethylated DNA and functions in DNA methylation maintenance, but has lately been characterized as a reader of both 5-mC and 5-hmC (Sharif et al. 2007; Arita et al. 2008; Frauer et al. 2011a). The functional consequences of NP95/UHRF1 binding 5-hmC have yet to be examined in vivo (Frauer et al. 2011a). MeCP2, already established as a 5-mC binding protein, has also demonstrated 5-hmC binding in two studies in neural tissues and mouse ES cells (Mellén et al. 2012; Spruijt et al. 2013). It is unclear how the MeCP2:5-mC binding profile differs from MeCP2:5-hmC binding profile or whether the function of this protein depends on its ligand. However, a mutated version of MeCP2 is expressed in Rett Syndrome: R133C. Experiments with electrophoretic mobility shift assays indicated that this MeCP2 protein is incapable of binding 5-hmC (Mellén et al. 2012). This indicates that readers of 5-hmC may have an essential role in normal neuronal phenotype. Finally, MBD3 has been identified as a weak binder of 5-hmC in two studies (Yildirim et al. 2011; Mellén et al. 2012). One study that indicated MDB3 as a 5-hmC reader also indicated Tet1:Mbd3 complexes bind 5-hmC rich regions of the genome in ES cells, and identified Mbd3 as both a recruiter of Tet1 and an effector that mediates gene expression (Yildirim et al. 2011). However, other studies have found that MBD3 binds the same genomic regions independent of the presence of 5-mC and 5-hmC, contradicting its proposed function as a 5-hmC reader (Iurlaro et al. 2013; Spruijt et al. 2013). This also potentially complicates the findings of the relationship between Tet1 and Mdb3 (Baubec et al. 2013; Iurlaro et al. 2013; Spruijt et al. 2013). The significance of MBD3:5-hmC binding is under debate.
Overall, although many potential proteins that bind 5-mC and 5-hmC have been identified, the function of many of these interactions is mostly unknown. Future studies of the biological functions of these interactions are needed to better understand how cytosine modifications regulate the genome.
6.2 Some Binding Partners Interact with All Three TETs
Current research on TET binding proteins shows that these binding partners can modify TET activity, localization, and stability (Fig. 5). Many of the identified proteins are not yet fully characterized and may have functions outside of their relationship with TETs.

TET binding partners impact TET activity, localization, and stability. (a) Binding of OGT increases TET activity when bound to DNA. (b) Calpains bind and degrade TETs. (c) EBF1 brings TET2 to promoters of genes, where it hydroxylates 5-mC to 5-hmC
The interaction between Tet and Ogt has been studied extensively. Independent of the O-GlcNAcylation activity, stable Tet:Ogt interaction promotes Tet activity and regulates Tet genomic localization. OGT binds TET1 through a conserved C-terminal domain, and is thought to form a larger complex with at least SIN3A (Vella et al. 2013; Hrit et al. 2018). Ogt and Tet1 colocalized at unmethylated CpG-rich promoters (Vella et al. 2013). Additional studies have found that loss of Ogt protein resulted in abnormal Tet1 localization in ES cells, indicating Ogt is important for Tet1 binding of some genomic sites (Shi et al. 2013). Multiple groups have reported that the loss of Tet1:Ogt interaction also results in reduced 5-hmC level and expression from Tet1-activated genes (Shi et al. 2013; Hrit et al. 2018). Ultimately, these experiments indicate that the role of Tet1:Ogt binding is to increase its activity and recruit it to genomic sites (Fig. 5a) (Shi et al. 2013; Hrit et al. 2018). TET2/3:OGT binding does not appear to have a significant impact on the 5-hmC epigenetic landscape, instead playing a scaffolding role for OGT (Deplus et al. 2013; Ito et al. 2014).
Tet stability appears to be regulated, at least in part, by calpains in mouse ES cells (Wang and Zhang 2014). Calpains 1 and 2 regulate TET stability via direct binding and cleavage of TET enzymes (Fig. 5b) (Wang and Zhang 2014). This mechanism of TET protein degradation is predominantly found in certain stages of development when TET expression is high (Wang and Zhang 2014). Tet1 and Tet2 are degraded by calpain1 in mouse ES cells, whereas Tet3 is degraded by calpain2 during ES cell differentiation (Wang and Zhang 2014). Loss of calpains resulted in twofold higher 5-hmC levels, downregulation of Cdx2 and Eomes, and upregulation of Ngn2 and Pax6 in ES cells (Wang and Zhang 2014). Therefore, TET:calpain complexes are required for normal 5-hmC regulation and gene expression in ES cells.
TDG, MBD4, SMUG1, NEIL1, NEIL2, NEIL3, PARP1, LIG3, and XRCC1 are part of the BER pathway and interact with TET during this process (Müller et al. 2014). Their colocalization with TETs is thought to facilitate removal of 5-caC and repair of the site. However, a role for direct binding between TET and each specific BER protein has yet to be described (Müller et al. 2014).
Beyond the shared interactors, the TET proteins have been shown to have binding partners unique to only one or two of them. These unique partners may explain the non-overlapping functions of TETs and provide further insight to the regulation of 5-hmC.
6.3 TET1 Binding Proteins
The function of most described interactors is regulating the localization of TET1 within the genome. The binding partners of TET1 are chromatin-binding proteins: transcription factors and epigenetic modifiers. They are thought to function at different levels of TET1 localization, from promoting TET1 binding to DNA to recruiting TET1 to target regions in the genome.
Proteins that are thought to facilitate TET1 binding to DNA include MeCP2, HDAC1/6/7, EZH2, mSin3A, PCNA, and LSD1 (Cartron et al. 2013). All of these proteins were found to bind TET1 directly and each other, thus stabilizing the TET1-DNA interaction. Further experimentation revealed that, in addition to this function, TET1:EZH2 and TET1:SIN3A are thought to recruit TET1 to specific genomic elements (Chandru et al. 2018). It should be stated that many proteins labeled as ‘recruiters’ of TET1 have not been thoroughly tested on a genomic scale and are usually tested only at selected targets (Williams et al. 2011; Cartron et al. 2013). TET1:EZH2 and TET1:SIN3A have been implicated in the regulation of HOXD12 and NES1 respectively. Loss of the complexes resulted in increased methylation at each promoter and, therefore, decreased expression. Another study looked at the impact of Tet1:Sin3a complex on global genomic targets in mouse ES cells and showed that Sin3a recruited Tet1 to demethylate the promoter of Lefty1 (Zhu et al. 2018). Loss of Sin3a resulted in altered localization of Tet1 and deregulation of 111 genes. However, the level and distribution of 5-hmC was not assessed.
Other identified proteins that act to recruit TET1 to genomic sites include PCNA and the Mbd3/NuRD complex (Williams et al. 2011; Yildirim et al. 2011; Cartron et al. 2013; Costa et al. 2013). Complexes containing TET1:PCNA are thought to play a role in the regulation of MUC2 and RRMI, as experiments identified both proteins bound to these genes through chromatin-immunoprecipitation (ChIP) with α-PCNA antibody and sequential ChIP with α-TET1 antibody.
6.4 TET2 Binding Proteins
Identified binding partners of TET2 promote its localization and stabilization. Two factors that recruit TET2 to genomic sites are EBF1 and WT1 (Fig. 5c). EBF1 and TET2 are part of the same complex, which localize to hypermethylated promoters in the context of IDH mutations in chondrosarcoma (Fig. 5c) (Guilhamon et al. 2013). It is hypothesized that TET2:EBF1 complexes act to target tissue specific demethylation sites, but further research is required to identify changes in the 5-hmC landscape and impacts on gene expression. The relationship between WT1 and TET2 was discovered in hematopoietic cells, which commonly feature mutually exclusive mutations in WT1 and TET2 in AML (Rampal et al. 2014). The TET2:WT1 complexes are believed to play a role in maintaining normal 5-hmC levels and localization in hematopoietic cells. Another study showed that TET2 occupied the promoters of WT1 target genes, and this occupancy is lost when WT1 expression was depleted, indicating WT1 recruited TET2 to genomic sites (Wang et al. 2015). Further, mutation of either WT1 or TET2 resulted in lower total 5-hmC levels and aberrant distribution at enhancers, gene bodies, and distal regulatory elements. Conversely, when WT1 and TET2 were coexpressed in HEK293T cells, 5-hmC levels increased at promoters and transcription start sites of WT1 targets. When WT1 was depleted, 5-hmC enrichment at WT1 target genes was lost. When IDH, WT1, and TET2 were mutated separately, 5-hmC changes in the gene body and distal regulatory regions had a positive correlation with gene expression changes (Rampal et al. 2014). When TET2 was ectopically expressed in HEK293T cells, there was increased gene expression from WT1 targets, which was abrogated upon loss of TET2 catalytic activity (Wang et al. 2015). Finally, proliferation of leukemia cells was stimulated by the loss of either TET2 or WT1 but the loss of both was not additive, indicating that TET2 and WT1 function in the same pathway that controls cell proliferation. Taken together, evidence suggests that the TET2:WT1 complex acts to maintain 5-hmC and normal gene expression, and that loss of either protein results in an oncogenic phenotype (Rampal et al. 2014; Wang et al. 2015).
Idax and DNMT1 have been found to be involved in TET2 protein degradation and stability (Ko et al. 2010; Rampal et al. 2014; Wang et al. 2015; Zhang et al. 2017). A study shows that Idax and Tet2 bind to each other, and the coexpression of Idax and Tet2 in HEK293T cells led to a decrease of Tet2 protein and a decrease in 5-hmC, but only when the DNA binding domain of Idax was present (Ko et al. 2013). This evidence suggests that Idax binds DNA and Tet2 then recruits an unknown degradation complex to target Tet2, which results in decreased 5-hmC. Binding of TET2 to DNMT1 occurs after the acetylation of TET2, and appears to promote protein stability (Zhang et al. 2017). The impact of TET2:DNMT1 on 5-hmC has yet to be explored, but it appears to prevent abnormal methylation of DNA. The location of 5-hmC loss and any impact on gene expression have yet to be investigated.
6.5 TET3 Binding Proteins
The known binding partners of Tet3, Rest and WT1, are believed to be involved in TET3 genomic localization (Rampal et al. 2014). The Tet3:Rest complex forms during murine retina maturation, which requires the formation of a complex neural network (Perera et al. 2015). Rest binds a Tet3 isoform that lacks the CXXC domain, indicating it may serve as a DNA targeting factor for Tet3. Coexpression of Tet3 and Rest leads to a dramatic increase in 5-hmC levels, more than overexpression of Tet3 alone. Rest target genes were enriched in a subset of genes that gained 5-hmC, suggesting a synergistic interaction. Measurement of global protein levels indicated that the protein levels of many Rest gene targets were significantly increased when Tet3 was overexpressed. However, this increase was lost when Rest was not present, meaning both Tet3 and Rest are required for the observed phenotype. Finally, TET3:WT1 forms a complex following the depletion of TET2 in hematopoietic models, but this is considered a compensatory mechanism for the loss of TET2 (Rampal et al. 2014).
7 Conclusions
The 5-hmC epigenetic landscape is modulated by the regulation of TET enzymes at the genetic, transcription, and protein levels. As discussed above, the activity of TET enzymes is tightly controlled at each of these levels. However, much of our knowledge is incomplete. More research is needed to understand the mechanisms that control TET gene expression and TET PTMs. In addition, although many TET binding partners have been identified, more research is required to determine their functions and mechanisms of action. Loss or change of any of these regulation mechanisms causes TET deregulation, which can alter the 5-hmC landscape and play a pathogenic role in disease. For this reason, study of the TET enzymes, in particular how they are regulated and impact the 5-hmC landscape, will provide answers to questions about the role of the epigenetic state in health and disease.
References
Abdel-Wahab O, Mullally A, Hedvat C et al (2009) Genetic characterization of TET1, TET2, and TET3 alterations in myeloid malignancies. Blood 114:144–147
An J, Rao A, Ko M (2017) TET family dioxygenases and DNA demethylation in stem cells and cancers. Exp Mol Med 49:e323
Arita K, Ariyoshi M, Tochio H et al (2008) Recognition of hemi-methylated DNA by the SRA protein UHRF1 by a base-flipping mechanism. Nature 455:818–821
Bacher U, Weissmann S, Kohlmann A et al (2012) TET2 deletions are a recurrent but rare phenomenon in myeloid malignancies and are frequently accompanied by TET2 mutations on the remaining allele. Br J Haematol 156:67–75
Baubec T, Ivánek R, Lienert F et al (2013) Methylation-dependent and -independent genomic targeting principles of the MBD protein family. Cell 153:480–492
Bauer C, Göbel K, Nagaraj N et al (2015) Phosphorylation of TET proteins is regulated via O-GlcNAcylation by the O-linked N-acetylglucosamine transferase (OGT). J Biol Chem 290:4801–4812
Baylin SB, Höppener JW, de Bustros A et al (1986) DNA methylation patterns of the calcitonin gene in human lung cancers and lymphomas. Canc Res 46:2917–2922
Blaschke K, Ebata KT, Karimi MM et al (2013) Vitamin C induces Tet-dependent DNA demethylation and a blastocyst-like state in ES cells. Nature 500:222–226
Booth MJ, Ost TW, Beraldi D et al (2013) Oxidative bisulfite sequencing of 5-methylcytosine and 5-hydroxymethylcytosine. Nat Protocol 8:1841–1851
Boyes J, Bird A (1991) DNA methylation inhibits transcription indirectly via a methyl-CpG binding protein. Cell 64:1123–1134
Burmeister T, Meyer C, Schwartz S et al (2009) The MLL recombinome of adult CD10-negative B-cell precursor acute lymphoblastic leukemia: results from the GMALL study group. Blood 113:4011–4015
Cao X (2016) Self-regulation and cross-regulation of pattern-recognition receptor signalling in health and disease. Nat Rev Immunol 16:35–50
Cartron P-FF, Nadaradjane A, Lepape F et al (2013) Identification of TET1 partners that control its DNA-demethylating function. Gene Canc 4:235–241
Chandru A, Bate N, Vuister GW et al (2018) Sin3A recruits Tet1 to the PAH1 domain via a highly conserved Sin3-interaction domain. Sci Rep 8:14689
Chen C-CC, Wang K-YY, Shen C-KJK (2012) The mammalian de novo DNA methyltransferases DNMT3A and DNMT3B are also DNA 5-hydroxymethylcytosine dehydroxymethylases. J Biol Chem 287:33116–33121
Chen J, Guo L, Zhang L et al (2013) Vitamin C modulates TET1 function during somatic cell reprogramming. Nat Genet 45:1504–1509
Chen D, Maruschke M, Hakenberg O et al (2017a) TOP2A, HELLS, ATAD2, and TET3 are novel prognostic markers in renal cell carcinoma. Urology 102:e1-265–e1e267
Chen Q, Yin D, Zhang Y et al (2017b) MicroRNA-29a induces loss of 5-hydroxymethylcytosine and promotes metastasis of hepatocellular carcinoma through a TET-SOCS1-MMP9 signaling axis. Cell Death Dis 8:e2906
Cheng J, Guo S, Chen S et al (2013) An extensive network of TET2-targeting MicroRNAs regulates malignant hematopoiesis. Cell Rep 5:471–481
Chung TL, Brena RM, Kolle G et al (2010) Vitamin C promotes widespread yet specific DNA demethylation of the epigenome in human embryonic stem cells. Stem Cells 28:1848–1855
Coey CT, Malik SS, Pidugu LS et al (2016) Structural basis of damage recognition by thymine DNA glycosylase: key roles for N-terminal residues. Nucleic Acids Res 44:10248–10258
Cohen-Karni D, Xu D, Apone L et al (2011) The MspJI family of modification-dependent restriction endonucleases for epigenetic studies. Proc Natl Acad Sci U S A 108:11040–11045
Costa Y, Ding J, Theunissen TW et al (2013) NANOG-dependent function of TET1 and TET2 in establishment of pluripotency. Nature 495:370–374
Dang L, White DW, Gross S et al (2009) Cancer-associated IDH1 mutations produce 2-hydroxyglutarate. Nature 462:739–744
Dawlaty MM, Ganz K, Powell BE et al (2011) Tet1 is dispensable for maintaining pluripotency and its loss is compatible with embryonic and postnatal development. Cell Stem Cell 9:166–175
Dawlaty MM, Breiling A, Le T et al (2013) Combined deficiency of Tet1 and Tet2 causes epigenetic abnormalities but is compatible with postnatal development. Dev Cell 24:310–323
Dawlaty MM, Breiling A, Le T et al (2014) Loss of Tet enzymes compromises proper differentiation of embryonic stem cells. Dev Cell 29:102–111
Delhommeau F, Dupont S, Della Valle V et al (2009) Mutation in TET2 in myeloid cancers. N Engl J Med 360:2289–2301
Deplus R, Delatte B, Schwinn MK et al (2013) TET2 and TET3 regulate GlcNAcylation and H3K4 methylation through OGT and SET1/COMPASS. EMBO J 32:645–655
Euba B, Vizmanos JLL, García-Granero M et al (2012) A meta-analysis of TET2 mutations shows a distinct distribution pattern in de novo acute myeloid leukemia and chronic myelomonocytic leukemia. Leuk Lymphoma 53:1230–1233
Feinberg AP, Tycko B (2004) The history of cancer epigenetics. Nat Rev Canc 4:143–153
Feinberg AP, Vogelstein B (1983) Hypomethylation distinguishes genes of some human cancers from their normal counterparts. Nature 301:89–92
Feng J, Shao N, Szulwach KE et al (2015) Role of Tet1 and 5-hydroxymethylcytosine in cocaine action. Nat Neurosci 18:536–544
Fernandez AF, Valledor L, Vallejo F et al (2018) Quantification of global DNA methylation levels by mass spectrometry. Meth Mol Biol 1708:49–58
Ficz G, Branco MR, Seisenberger S et al (2011) Dynamic regulation of 5-hydroxymethylcytosine in mouse ES cells and during differentiation. Nature 473:398–402
Field SF, Beraldi D, Bachman M et al (2015) Accurate measurement of 5-methylcytosine and 5-hydroxymethylcytosine in human cerebellum DNA by oxidative bisulfite on an array (OxBS-array). PLoS One 10:e0118202
Figueroa ME, Abdel-Wahab O, Lu C et al (2010) Leukemic IDH1 and IDH2 mutations result in a hypermethylation phenotype, disrupt TET2 function, and impair hematopoietic differentiation. Canc Cell 18:553–567
Frauer C, Hoffmann T, Bultmann S et al (2011a) Recognition of 5-hydroxymethylcytosine by the Uhrf1 SRA domain. PLoS One 6:e21306
Frauer C, Rottach A, Meilinger D et al (2011b) Different binding properties and function of CXXC zinc finger domains in Dnmt1 and Tet1. PLoS One 6:e16627
Gaidzik VI, Paschka P, Späth D et al (2012) TET2 mutations in acute myeloid leukemia (AML): results from a comprehensive genetic and clinical analysis of the AML study group. J Clin Oncol 30:1350–1357
Gama-Sosa MA, Slagel VA, Trewyn RW et al (1983) The 5-methylcytosine content of DNA from human tumors. Nucleic Acids Res 11:6883–6894
Globisch D, Münzel M, Müller M et al (2010) Tissue distribution of 5-hydroxymethylcytosine and search for active demethylation intermediates. PLoS One 5:e15367
Good CR, Madzo J, Patel B et al (2017) A novel isoform of TET1 that lacks a CXXC domain is overexpressed in cancer. Nucleic Acids Res 45:8269–8281
Gorres KL, Raines RT (2010) Prolyl 4-hydroxylase. Crit Rev Biochem Mol Biol 45:106–124
Gu T-PP, Guo F, Yang H et al (2011) The role of Tet3 DNA dioxygenase in epigenetic reprogramming by oocytes. Nature 477:606–610
Guilhamon P, Eskandarpour M, Halai D et al (2013) Meta-analysis of IDH-mutant cancers identifies EBF1 as an interaction partner for TET2. Nat Comm 4:2166
Guo JU, Ma DK, Mo H et al (2011a) Neuronal activity modifies the DNA methylation landscape in the adult brain. Nat Neurosci 14:1345–1351
Guo JU, Su Y, Zhong C et al (2011b) Hydroxylation of 5-methylcytosine by TET1 promotes active DNA demethylation in the adult brain. Cell 145:423–434
Hackett JA, Sengupta R, Zylicz JJ et al (2013) Germline DNA demethylation dynamics and imprint erasure through 5-hydroxymethylcytosine. Science 339:448–452
Hajkova P, Erhardt S, Lane N et al (2002) Epigenetic reprogramming in mouse primordial germ cells. Mech Dev 117:15–23
Hashimoto H, Pais JE, Zhang X et al (2014) Structure of a Naegleria Tet-like dioxygenase in complex with 5-methylcytosine DNA. Nature 506:391–395
Hashimoto H, Pais JE, Dai N et al (2015) Structure of Naegleria Tet-like dioxygenase (NgTet1) in complexes with a reaction intermediate 5-hydroxymethylcytosine DNA. Nucleic Acids Res 43:10713–10721
He Y-FF, Li B-ZZ, Li Z et al (2011) Tet-mediated formation of 5-carboxylcytosine and its excision by TDG in mammalian DNA. Science 333:1303–1307
Hendrich B, Bird A (1998) Identification and characterization of a family of mammalian methyl-CpG binding proteins. Mol Cell Biol 18:6538–6547
Hendrich B, Tweedie S (2003) The methyl-CpG binding domain and the evolving role of DNA methylation in animals. Trends Genet 19:269–277
Hon GC, Song C-XX, Du T et al (2014) 5mC oxidation by Tet2 modulates enhancer activity and timing of transcriptome reprogramming during differentiation. Mol Cell 56:286–297
Hrit J, Goodrich L, Li C et al (2018) OGT binds a conserved C-terminal domain of TET1 to regulate TET1 activity and function in development. eLife 7:e34870
Hu L, Li Z, Cheng J et al (2013) Crystal structure of TET2-DNA complex: insight into TET-mediated 5mC oxidation. Cell 155:1545–1555
Hu X, Zhang L, Mao S-QQ et al (2014) Tet and TDG mediate DNA demethylation essential for mesenchymal-to-epithelial transition in somatic cell reprogramming. Cell Stem Cell 14:512–522
Hu L, Lu J, Cheng J et al (2015) Structural insight into substrate preference for TET-mediated oxidation. Nature 527:118–122
Huang H, Jiang X, Li Z et al (2013) TET1 plays an essential oncogenic role in MLL-rearranged leukemia. Proc Natl Acad Sci U S A 110:11994–11999
Huang Y, Chavez L, Chang X et al (2014) Distinct roles of the methylcytosine oxidases Tet1 and Tet2 in mouse embryonic stem cells. Proc Natl Acad Sci U S A 111:1361–1366
Huang H, Jiang X, Wang J et al (2016) Identification of MLL-fusion/MYC⊣miR-26⊣TET1 signaling circuit in MLL-rearranged leukemia. Canc Lett 372:157–165
Ito S, D’Alessio AC, Taranova OV et al (2010) Role of Tet proteins in 5mC to 5hmC conversion, ES-cell self-renewal and inner cell mass specification. Nature 466:1129–1133
Ito S, Shen L, Dai Q et al (2011) Tet proteins can convert 5-methylcytosine to 5-formylcytosine and 5-carboxylcytosine. Science 333:1300–1303
Ito R, Katsura S, Shimada H et al (2014) TET3-OGT interaction increases the stability and the presence of OGT in chromatin. Genes Cells 19:52–65
Ittel A, Jeandidier E, Helias C et al (2013) First description of the t(10;11)(q22;q23)/MLL-TET1 translocation in a T-cell lymphoblastic lymphoma, with subsequent lineage switch to acute myelomonocytic myeloid leukemia. Haematologica 98:e166–e168
Itzykson R, Kosmider O, Renneville A et al (2013) Prognostic score including gene mutations in chronic myelomonocytic leukemia. J Clin Oncol 31:2428–2436
Iurlaro M, Ficz G, Oxley D et al (2013) A screen for hydroxymethylcytosine and formylcytosine binding proteins suggests functions in transcription and chromatin regulation. Genome Biol 14:R119
Iyer LM, Tahiliani M, Rao A et al (2009) Prediction of novel families of enzymes involved in oxidative and other complex modifications of bases in nucleic acids. Cell Cycle 8:1698–1710
Jankowska AM, Szpurka H, Tiu RV et al (2009) Loss of heterozygosity 4q24 and TET2 mutations associated with myelodysplastic/myeloproliferative neoplasms. Blood 113:6403–6410
Jäwert F, Hasséus B, Kjeller G et al (2013) Loss of 5-hydroxymethylcytosine and TET2 in oral squamous cell carcinoma. Anticancer Res 33:4325–4328
Jiang D, Zhang Y, Hart RP et al (2015) Alteration in 5-hydroxymethylcytosine-mediated epigenetic regulation leads to Purkinje cell vulnerability in ATM deficiency. Brain 138:3520–3536
Jin S-GG, Jiang Y, Qiu R et al (2011) 5-Hydroxymethylcytosine is strongly depleted in human cancers but its levels do not correlate with IDH1 mutations. Canc Res 71:7360–7365
Jin Y, Shao Y, Shi X et al (2016) Mutational profiling of non-small-cell lung cancer patients resistant to first-generation EGFR tyrosine kinase inhibitors using next generation sequencing. Oncotarget 7:61755–61763
Jjingo D, Conley AB, Yi SV et al (2012) On the presence and role of human gene-body DNA methylation. Oncotarget 3:462–474
Jones PA (2012) Functions of DNA methylation: islands, start sites, gene bodies and beyond. Nat Rev Genet 13:484–492
Kafer GR, Li X, Horii T et al (2016) 5-Hydroxymethylcytosine marks sites of DNA damage and promotes genome stability. Cell Rep 14:1283–1292
Kang J, Lienhard M, Pastor WA et al (2015) Simultaneous deletion of the methylcytosine oxidases Tet1 and Tet3 increases transcriptome variability in early embryogenesis. Proc Natl Acad Sci U S A 112:45
Ko M, Huang Y, Jankowska AM et al (2010) Impaired hydroxylation of 5-methylcytosine in myeloid cancers with mutant TET2. Nature 468:839–843
Ko M, An J, Bandukwala HS et al (2013) Modulation of TET2 expression and 5-methylcytosine oxidation by the CXXC domain protein IDAX. Nature 497:122–126
Koh KP, Yabuuchi A, Rao S et al (2011) Tet1 and Tet2 regulate 5-hydroxymethylcytosine production and cell lineage specification in mouse embryonic stem cells. Cell Stem Cell 8:200–213
Kriaucionis S, Heintz N (2009) The nuclear DNA base 5-hydroxymethylcytosine is present in Purkinje neurons and the brain. Science 324:929–930
Kudo Y, Tateishi K, Yamamoto K et al (2012) Loss of 5-hydroxymethylcytosine is accompanied with malignant cellular transformation. Canc Sci 103:670–676
Kuiper C, Vissers MC (2014) Ascorbate as a co-factor for Fe- and 2-oxoglutarate dependent dioxygenases: physiological activity in tumor growth and progression. Front Oncol 4:359
Kumar D, Aggarwal M, Kaas GA et al (2015) Tet1 oxidase regulates neuronal gene transcription, active DNA hydroxy-methylation, object location memory, and threat recognition memory. Neuroepigenetics 4:12–27
Langemeijer SM, Kuiper RP, Berends M et al (2009) Acquired mutations in TET2 are common in myelodysplastic syndromes. Nat Genet 41:838–842
Langlois T, da Costa Reis Monte-Mor B, Lenglet G et al (2014) TET2 deficiency inhibits mesoderm and hematopoietic differentiation in human embryonic stem cells. Stem Cells 32:2084–2097
Lasho TL, Vallapureddy R, Finke CM et al (2018) Infrequent occurrence of TET1, TET3, and ASXL2 mutations in myelodysplastic/myeloproliferative neoplasms. Blood Canc J 8:32
Laukka T, Mariani CJ, Ihantola T et al (2015) Fumarate and succinate regulate expression of hypoxia-inducible genes via TET enzymes. J Biol Chem 291:4256–4265
Lee S-GG, Cho SY, Kim MJ et al (2013) Genomic breakpoints and clinical features of MLL-TET1 rearrangement in acute leukemias. Haematologica 98:7
Lee HJ, Hore TA, Reik W (2014) Reprogramming the methylome: erasing memory and creating diversity. Cell Stem Cell 14:710–719
Lewis JD, Meehan RR, Henzel WJ et al (1992) Purification, sequence, and cellular localization of a novel chromosomal protein that binds to methylated DNA. Cell 69:905–914
Li C, Lan Y, Schwartz-Orbach L et al (2015a) Overlapping requirements for Tet2 and Tet3 in normal development and hematopoietic stem cell emergence. Cell Rep 12:1133–1143
Li T, Yang D, Li J et al (2015b) Critical role of Tet3 in neural progenitor cell maintenance and terminal differentiation. Mol Neurobiol 51:142–154
Lian CG, Xu Y, Ceol C et al (2012) Loss of 5-hydroxymethylcytosine is an epigenetic hallmark of melanoma. Cell 150:1135–1146
Lin IHH, Chen Y-FF, Hsu M-TT (2017) Correlated 5-hydroxymethylcytosine (5hmC) and gene expression profiles underpin gene and organ-specific epigenetic regulation in adult mouse brain and liver. PLoS One 12:e0170779
Liu D, Li G, Zuo Y (2018) Function determinants of TET proteins: the arrangements of sequence motifs with specific codes. Brief Bioinform
Liutkevičiūtė Z, Kriukienė E, Ličytė J et al (2014) Direct decarboxylation of 5-carboxylcytosine by DNA C5-methyltransferases. J Am Chem Soc 136:5884–5887
Lorsbach RB, Moore J, Mathew S et al (2003) TET1, a member of a novel protein family, is fused to MLL in acute myeloid leukemia containing the t(10;11)(q22;q23). Leukemia 17:637–641
Losman J-AA, Kaelin WG (2013) What a difference a hydroxyl makes: mutant IDH, (R)-2-hydroxyglutarate, and cancer. Gene Dev 27:836–852
Lu F, Liu Y, Jiang L et al (2014) Role of Tet proteins in enhancer activity and telomere elongation. Gene Dev 28:2103–2119
Madzo J, Liu H, Rodriguez A et al (2014) Hydroxymethylation at gene regulatory regions directs stem/early progenitor cell commitment during erythropoiesis. Cell Rep 6:231–244
Maiti A, Drohat AC (2011) Thymine DNA glycosylase can rapidly excise 5-formylcytosine and 5-carboxylcytosine: potential implications for active demethylation of CpG sites. J Biol Chem 286:35334–35338
Mariani CJ, Vasanthakumar A, Madzo J et al (2014) TET1-mediated hydroxymethylation facilitates hypoxic gene induction in neuroblastoma. Cell Rep 7:1343–1352
Mayer W, Niveleau A, Walter J et al (2000) Demethylation of the zygotic paternal genome. Nature 403:501–502
Meehan RR, Lewis JD, McKay S et al (1989) Identification of a mammalian protein that binds specifically to DNA containing methylated CpGs. Cell 58:499–507
Meldi K, Qin T, Buchi F et al (2015) Specific molecular signatures predict decitabine response in chronic myelomonocytic leukemia. J Clin Investig 125:1857–1872
Mellén M, Ayata P, Dewell S et al (2012) MeCP2 binds to 5hmC enriched within active genes and accessible chromatin in the nervous system. Cell 151:1417–1430
Merlevede J, Droin N, Qin T et al (2016) Mutation allele burden remains unchanged in chronic myelomonocytic leukaemia responding to hypomethylating agents. Nat Commun 7:10767
Meyer C, Hofmann J, Burmeister T et al (2013) The MLL recombinome of acute leukemias in 2013. Leukemia 27:2165–2176
Minor EA, Young JI, Wang G (2013) Ascorbate induces ten-eleven translocation (Tet) methylcytosine dioxygenase-mediated generation of 5-hydroxymethylcytosine. J Biol Chem 288:13669–13674
Misawa K, Imai A, Mochizuki D et al (2018) Association of TET3 epigenetic inactivation with head and neck cancer. Oncotarget 9:24480–24493
Müller U, Bauer C, Siegl M et al (2014) TET-mediated oxidation of methylcytosine causes TDG or NEIL glycosylase dependent gene reactivation. Nucleic Acids Res 42:8592–8604
Nakagawa T, Lv L, Nakagawa M et al (2015) CRL4(VprBP) E3 ligase promotes monoubiquitylation and chromatin binding of TET dioxygenases. Mol Cell 57:247–260
Nan X, Campoy FJ, Bird A (1997) MeCP2 is a transcriptional repressor with abundant binding sites in genomic chromatin. Cell 88:471–481
Neri F, Incarnato D, Krepelova A et al (2015) TET1 is controlled by pluripotency-associated factors in ESCs and downmodulated by PRC2 in differentiated cells and tissues. Nucleic Acids Res 43:6814–6826
Nestor CE, Meehan RR (2014) Hydroxymethylated DNA immunoprecipitation (hmeDIP). Meth Mol Biol 1094:259–267
Nickerson ML, Im KM, Misner KJ et al (2013) Somatic alterations contributing to metastasis of a castration-resistant prostate cancer. Hum Mutat 34:1231–1241
Ono R, Taki T, Taketani T et al (2002) LCX, leukemia-associated protein with a CXXC domain, is fused to MLL in acute myeloid leukemia with trilineage dysplasia having t(10;11)(q22;q23). Canc Res 62:4075–4080
Oswald J, Engemann S, Lane N et al (2000) Active demethylation of the paternal genome in the mouse zygote. Curr Biol 10:475–478
Pan H, Bilinovich SM, Kaur P et al (2017) CpG and methylation-dependent DNA binding and dynamics of the methylcytosine binding domain 2 protein at the single-molecule level. Nucleic Acids Res 45:9164–9177
Pastor WA, Pape UJ, Huang Y et al (2011) Genome-wide mapping of 5-hydroxymethylcytosine in embryonic stem cells. Nature 473:394–397
Patel JP, Gönen M, Figueroa ME et al (2012) Prognostic relevance of integrated genetic profiling in acute myeloid leukemia. N Engl J Med 366:1079–1089
Perera A, Eisen D, Wagner M et al (2015) TET3 is recruited by REST for context-specific hydroxymethylation and induction of gene expression. Cell Rep 11:283–294
Pidugu LS, Flowers JW, Coey CT et al (2016) Structural basis for excision of 5-formylcytosine by thymine DNA glycosylase. Biochemistry 55:6205–6208
Pollard PJ, Brière JJ, Alam NA et al (2005) Accumulation of Krebs cycle intermediates and over-expression of HIF1alpha in tumours which result from germline FH and SDH mutations. Hum Mol Genet 14:2231–2239
Quivoron C, Couronné L, Valle DV et al (2011) TET2 inactivation results in pleiotropic hematopoietic abnormalities in mouse and is a recurrent event during human lymphomagenesis. Canc Cell 20:25–38
Rampal R, Alkalin A, Madzo J et al (2014) DNA hydroxymethylation profiling reveals that WT1 mutations result in loss of TET2 function in acute myeloid leukemia. Cell Rep 9:1841–1855
Reik W (2007) Stability and flexibility of epigenetic gene regulation in mammalian development. Nature 447:425–432
Robertson AB, Dahl JA, Ougland R et al (2012) Pull-down of 5-hydroxymethylcytosine DNA using JBP1-coated magnetic beads. Nat Protocol 7:340–350
Rudenko A, Dawlaty MM, Seo J et al (2013) Tet1 is critical for neuronal activity-regulated gene expression and memory extinction. Neuron 79:1109–1122
Sato Y, Yoshizato T, Shiraishi Y et al (2013) Integrated molecular analysis of clear-cell renal cell carcinoma. Nat Genet 45:860–867
Seiler CL, Fernandez J, Koerperich Z et al (2018) Maintenance DNA methyltransferase activity in the presence of oxidized forms of 5-methylcytosine: structural basis for ten eleven translocation-mediated DNA demethylation. Biochemistry 57:6061–6069
Seki Y, Hayashi K, Itoh K et al (2005) Extensive and orderly reprogramming of genome-wide chromatin modifications associated with specification and early development of germ cells in mice. Dev Biol 278:440–458
Sharif J, Muto M, Takebayashi S-i et al (2007) The SRA protein Np95 mediates epigenetic inheritance by recruiting Dnmt1 to methylated DNA. Nature 450:908–912
Shi F-TT, Kim H, Lu W et al (2013) Ten-eleven translocation 1 (Tet1) is regulated by O-linked N-acetylglucosamine transferase (Ogt) for target gene repression in mouse embryonic stem cells. J Biol Chem 288:20776–20784
Shi X, Yu Y, Luo M et al (2016) Loss of 5-hydroxymethylcytosine is an independent unfavorable prognostic factor for esophageal squamous cell carcinoma. PLoS One 11:e0153100
Slyvka A, Mierzejewska K, Bochtler M (2017) Nei-like 1 (NEIL1) excises 5-carboxylcytosine directly and stimulates TDG-mediated 5-formyl and 5-carboxylcytosine excision. Sci Rep 7:9001
Sohni A, Bartoccetti M, Khoueiry R et al (2015) Dynamic switching of active promoter and enhancer domains regulates Tet1 and Tet2 expression during cell state transitions between pluripotency and differentiation. Mol Cell Biol 35:1026–1042
Song C-XX, Szulwach KE, Fu Y et al (2011) Selective chemical labeling reveals the genome-wide distribution of 5-hydroxymethylcytosine. Nat Biotechnol 29:68–72
Song SJ, Ito K, Ala U et al (2013) The oncogenic microRNA miR-22 targets the TET2 tumor suppressor to promote hematopoietic stem cell self-renewal and transformation. Cell Stem Cell 13:87–101
Spruijt CG, Gnerlich F, Smits AH et al (2013) Dynamic readers for 5-(hydroxy)methylcytosine and its oxidized derivatives. Cell 152:1146–1159
Stroud H, Feng S, Morey Kinney S et al (2011) 5-Hydroxymethylcytosine is associated with enhancers and gene bodies in human embryonic stem cells. Genome Biol 12:R54
Szulwach KE, Li X, Li Y et al (2011) Integrating 5-hydroxymethylcytosine into the epigenomic landscape of human embryonic stem cells. PLoS Genet 7:e1002154
Tahiliani M, Koh KP, Shen Y et al (2009) Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in mammalian DNA by MLL partner TET1. Science 324:930–935
Tardy-Planechaud S, Fujimoto J, Lin SS et al (1997) Solid phase synthesis and restriction endonuclease cleavage of oligodeoxynucleotides containing 5-(hydroxymethyl)-cytosine. Nucleic Acids Res 25:553–559
Thienpont B, Steinbacher J, Zhao H et al (2016) Tumour hypoxia causes DNA hypermethylation by reducing TET activity. Nature 537:63–68
Turcan S, Rohle D, Goenka A et al (2012) IDH1 mutation is sufficient to establish the glioma hypermethylator phenotype. Nature 483:479–483
Valinluck V, Sowers LC (2007) Endogenous cytosine damage products alter the site selectivity of human DNA maintenance methyltransferase DNMT1. Canc Res 67:946–950
Varley KE, Gertz J, Bowling KM et al (2013) Dynamic DNA methylation across diverse human cell lines and tissues. Genome Res 23:555–567
Vasanthakumar A, Godley LA (2015) 5-Hydroxymethylcytosine in cancer: significance in diagnosis and therapy. Canc Genet 208:167–177
Vella P, Scelfo A, Jammula S et al (2013) Tet proteins connect the O-linked N-acetylglucosamine transferase Ogt to chromatin in embryonic stem cells. Mol Cell 49:645–656
Vincent JJ, Huang Y, Chen P-YY et al (2013) Stage-specific roles for tet1 and tet2 in DNA demethylation in primordial germ cells. Cell Stem Cell 12:470–478
Vogel Ciernia A, LaSalle J (2016) The landscape of DNA methylation amid a perfect storm of autism aetiologies. Nat Rev Neurosci 17:411–423
Wang Y, Zhang Y (2014) Regulation of TET protein stability by calpains. Cell Rep 6:278–284
Wang H, Guan S, Quimby A et al (2011) Comparative characterization of the PvuRts1I family of restriction enzymes and their application in mapping genomic 5-hydroxymethylcytosine. Nucleic Acids Res 39:9294–9305
Wang Y, Xiao M, Chen X et al (2015) WT1 recruits TET2 to regulate its target gene expression and suppress leukemia cell proliferation. Mol Cell 57:662–673
Wen L, Li X, Yan L et al (2014) Whole-genome analysis of 5-hydroxymethylcytosine and 5-methylcytosine at base resolution in the human brain. Genome Biol 15:R49
Williams K, Christensen J, Pedersen MT et al (2011) TET1 and hydroxymethylcytosine in transcription and DNA methylation fidelity. Nature 473:343–348
Wossidlo M, Nakamura T, Lepikhov K et al (2011) 5-Hydroxymethylcytosine in the mammalian zygote is linked with epigenetic reprogramming. Nat Commun 2:241
Wu H, Zhang Y (2011) Tet1 and 5-hydroxymethylation: a genome-wide view in mouse embryonic stem cells. Cell Cycle 10:2428–2436
Wu X, Zhang Y (2017) TET-mediated active DNA demethylation: mechanism, function and beyond. Nat Rev Genet 18:517–534
Wu D, Hu D, Chen H et al (2018) Glucose-regulated phosphorylation of TET2 by AMPK reveals a pathway linking diabetes to cancer. Nature 559:637–641
Wyatt GR, Cohen SS (1953) The bases of the nucleic acids of some bacterial and animal viruses: the occurrence of 5-hydroxymethylcytosine. Biochem J 55:774–782
Xu W, Yang H, Liu Y et al (2011a) Oncometabolite 2-hydroxyglutarate is a competitive inhibitor of α-ketoglutarate-dependent dioxygenases. Canc Cell 19:17–30
Xu Y, Wu F, Tan L et al (2011b) Genome-wide regulation of 5hmC, 5mC, and gene expression by Tet1 hydroxylase in mouse embryonic stem cells. Mol Cell 42:451–464
Xu Y, Xu C, Kato A et al (2012) Tet3 CXXC domain and dioxygenase activity cooperatively regulate key genes for Xenopus eye and neural development. Cell 151:1200–1213
Yamaguchi S, Hong K, Liu R et al (2012) Tet1 controls meiosis by regulating meiotic gene expression. Nature 492:443–447
Yamazaki J, Taby R, Vasanthakumar A et al (2012) Effects of TET2 mutations on DNA methylation in chronic myelomonocytic leukemia. Epigenetics 7:201–207
Yamazaki J, Jelinek J, Lu Y et al (2015) TET2 mutations affect non-CpG island DNA methylation at enhancers and transcription factor-binding sites in chronic myelomonocytic leukemia. Canc Res 75:2833–2843
Yang H, Liu Y, Bai F et al (2013) Tumor development is associated with decrease of TET gene expression and 5-methylcytosine hydroxylation. Oncogene 32:663–669
Yildirim O, Li R, Hung J-HH et al (2011) Mbd3/NURD complex regulates expression of 5-hydroxymethylcytosine marked genes in embryonic stem cells. Cell 147:1498–1510
Yin R, Mao SQ, Zhao B et al (2013) Ascorbic acid enhances Tet-mediated 5-methylcytosine oxidation and promotes DNA demethylation in mammals. J Am Chem Soc 135:10396–10403
Yu M, Hon GC, Szulwach KE et al (2012) Tet-assisted bisulfite sequencing of 5-hydroxymethylcytosine. Nat Protocol 7:2159–2170
Zhang H, Zhang X, Clark E et al (2010) TET1 is a DNA-binding protein that modulates DNA methylation and gene transcription via hydroxylation of 5-methylcytosine. Cell Res 20:1390–1393
Zhang R-RR, Cui Q-YY, Murai K et al (2013) Tet1 regulates adult hippocampal neurogenesis and cognition. Cell Stem Cell 13:237–245
Zhang Q, Liu X, Gao W et al (2014) Differential regulation of the ten-eleven translocation (TET) family of dioxygenases by O-linked β-N-acetylglucosamine transferase (OGT). J Biol Chem 289:5986–5996
Zhang W, Xia W, Wang Q et al (2016) Isoform switch of TET1 regulates DNA demethylation and mouse development. Mol Cell 64:1062–1073
Zhang YW, Wang Z, Xie W et al (2017) Acetylation enhances TET2 function in protecting against abnormal DNA methylation during oxidative stress. Mol Cell 65:323–335
Zhu F, Zhu Q, Ye D et al (2018) Sin3a-Tet1 interaction activates gene transcription and is required for embryonic stem cell pluripotency. Nucleic Acids Res 46:6026–6040
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2019 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Cao, J.Z., Hains, A.E., Godley, L.A. (2019). Regulation of 5-Hydroxymethylcytosine Distribution by the TET Enzymes. In: Jurga, S., Barciszewski, J. (eds) The DNA, RNA, and Histone Methylomes. RNA Technologies. Springer, Cham. https://doi.org/10.1007/978-3-030-14792-1_10
Download citation
DOI: https://doi.org/10.1007/978-3-030-14792-1_10
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-14791-4
Online ISBN: 978-3-030-14792-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)