
Abstract
Asthma and allergic disease result from complex environmental and genetic interactions. These factors have been identified by epidemiological studies, randomized prevention trials, and animal models. Vitamin E is one of the environmental factors that can regulate responses to challenge with allergens. Reports for vitamin E have seemingly varied outcomes regarding benefits of vitamin E, but these differences are consistent with the differential functions of the isoforms of vitamin E. Mechanistic studies have identified opposing functions of the vitamin E isoforms α-tocopherol and γ-tocopherol. Moreover, differences in global prevalence of asthma may be an outcome, at least in part, of differences in amount of isoforms of tocopherols. In order to interpret human and preclinical studies, reports need to include measurements of the tocopherol isoforms in the treatments, in vehicles for the treatments, and in the plasma and/or tissues before and after intervention. Studies of differential regulation of inflammation by tocopherol isoforms provide a foundation for development of interventions to improve lung function in disease and raise the possibility of early life dietary interventions to limit the development of lung disease.
Sources of Support: This study was supported by National Institutes of Health Grants R01AI127695, U01AI131337, and R01 AT004837.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
FormalPara Key Points-
Asthma and allergic disease result from complex environmental and genetic interactions.
-
Vitamin E is one of the environmental factors that can regulate responses to challenge with allergens.
-
However, outcomes of clinical studies on benefits of vitamin E regarding asthma and allergic inflammation vary.
-
The differential and partially opposing functions of the vitamin E isoforms α-tocopherol and γ-tocopherol as demonstrated in mechanistic studies appear to be an important variable in these inconsistent results reported in clinical studies.
-
A better understanding of the differential regulation of inflammation by isoforms of vitamin E is considered an important factor to improve quality of clinical studies when investigating the role of vitamin E to improve lung function in disease in adults and during development.
Asthma/Allergy Prevalence and Changes in Consumption of Forms of Vitamin E
Non-allergic asthma , allergic asthma, and allergy are heterogeneous diseases, resulting from complex interactions of environmental and genetic factors [1]. Allergic and asthmatic responses include bronchoconstriction, itch, pain, inflammation, and tissue remodeling. In humans, measurements of lung function for monitoring asthma include forced expiratory volume in 1 s (FEV1, forced volume blown out in 1 s) and forced vital capacity (FVC, forced volume when all air is blown out). Current therapies for asthma and allergies , including corticosteroids, have serious side effects. Therefore, it is critical to determine mechanisms for regulation of inflammation in allergy/asthma in order to identify novel approaches for interventions. Asthma is characterized by inflammatory processes with T-helper (Th) cell responses of the Th2 phenotype being considered crucial for the initiation and perpetuation of the inflammatory responses [2]. Important mediators of asthma and allergic inflammation are cytokines such as interleukin IL-4, IL-5, and IL-13. Asthmatic and allergic inflammation is characterized by elevated immunoglobulin E, mast cell degranulation, and eosinophilic inflammation [3]. Recruitment of eosinophils into the tissue is a consistent feature of allergic inflammation and allergic asthma [4,5,6]. A crucial component of recruitment of eosinophils during inflammation is leukocyte migration from the blood, across the endothelium, and into the tissue (transendothelial migration) [7,8,9]. Mechanisms for this eosinophil recruitment involve the coordinate actions of adhesion molecules, chemokines, and cytokines [10, 11]. Tocopherol isoform regulation of leukocyte recruitment and allergic inflammation has been studied in humans, animal models, and cell systems.
The World Health Organization reported a worldwide increase in asthma and allergies from 1950 to the present [12,13,14,15]. In 2012–2013, the US Centers for Disease Control and Prevention reported asthma and allergy prevalence as 10–20%, affecting about 26 million people, costing $56 million/year and 9 deaths/day [16,17,18]. The rapid rise in rates of asthma and the differences in rates among countries and in migrating populations indicate a role of the local environment, including the diet. In this same timespan, there has been an increase in the γ-tocopherol isoform in the diet, in infant formulas containing soybean oil that is rich in γ-tocopherol, and in vitamin supplements [19, 20]. The variation in global prevalence of asthma and allergies may be influenced, at least in part, by country-specific plasma γ-tocopherol concentrations. Moreover, it has been suggested that early life exposures to environmental factors increase risk of allergic disease [21]. Maternal exposure to environmental factors can alter neonatal hematopoietic or metabolic function [22,23,24,25,26,27,28,29]. In reports examining human maternal and paternal asthma associations with development of allergies in offspring, most associations are with maternal asthma [30]. Furthermore, there is higher prevalence of early-onset persistent asthma if the mother has uncontrolled asthma or moderate-to-severe controlled asthma as compared to mothers with mild controlled asthma [31].
In animal studies, offspring of allergic mothers have increased responses to suboptimal doses of allergens [30], and tocopherol isoforms regulate development of offspring cells of the immune system and immune responses [32,33,34,35,36,37,38,39,40]. Therefore, studies of the regulation of adult allergies and asthma as well as the development of allergic disease and asthma early in life are critical to generating approaches to limit these diseases.
Vitamin E Isoforms and Function During Asthma
During allergic responses and asthma, isoforms of vitamin E regulate cell signaling and scavenging of reactive oxygen species. The isoforms of vitamin E consumed and in human plasma vary among countries. The natural isoforms of vitamin E are synthesized by plants from tyrosine and chlorophyll [41]. Thus, although mammals do not synthesize vitamin E, mammals acquire vitamin E isoforms from food, cooking oils, and vitamin supplements. The natural isoforms of vitamin E are d-α-tocopherol, d-β-tocopherol, d-γ-tocopherol, and d-δ-tocopherol and the d-α-, d-β-, d-γ-, and d-δ-tocotrienol (Fig. 25.1). The level of these isoforms in cooking oils and supplements varies widely among products. Upon consumption of tocopherols, the tocopherols, which are lipids, are transported along with other lipids from the intestine by the lymph to the blood and then to the liver. These tocopherol isoforms are not interconverted by mammals. The isoform with the highest concentration in tissues is α-tocopherol. There are tenfold higher tissue concentrations of α-tocopherol than γ-tocopherol [42] because there is preferential loading of α-tocopherol on lipid particles in the liver by α-tocopherol transfer protein (αTTP) and because there is a higher rate of degradation of γ-tocopherol into its metabolites for excretion [43, 44]. The plasma concentrations of α-tocopherol are relatively similar across many countries [45]. αTTP is also expressed by trophoblasts, fetal endothelium, and amnion epithelium of the placenta, consistent with the critical role for α-tocopherol in placenta development [46, 47]. Tocopherols on plasma lipid particles are transferred to cells by plasma phospholipid transfer protein, scavenger receptors, or the lipoprotein lipase pathway [23]. In cells, tocopherols, which are lipids, are located in cell membranes and associated with lipophilic domains of proteins. On lipid particles and in cells, tocopherol isoforms have both antioxidant and non-antioxidant functions. The antioxidant activity of α-tocopherol and γ-tocopherol, at equal molar concentrations, is relatively similar with regard to scavenging reactive oxygen species (ROS) during lipid peroxidation [48, 49]. Because α-tocopherol is at tenfold higher concentrations in tissues than γ-tocopherol, α-tocopherol has tenfold more capacity for scavenging of ROS in vivo.

Natural vitamin E isoforms. The most abundant isoforms in human plasma and tissues are α-tocopherol and γ-tocopherol
In addition to antioxidant functions of tocopherols , non-antioxidant functions of tocopherol regulate allergic inflammation. Important for interpretation of tocopherol regulatory effects is that although γ-tocopherol is at tenfold lower concentrations in vivo, γ-tocopherol is very potent and at these tenfold lower concentrations, γ-tocopherol can block the benefit of α-tocopherol during allergic inflammation and asthma. This potent opposing effect of γ-tocopherol occurs because, at least, γ-tocopherol is an agonist of cell signals, whereas α-tocopherol is an antagonist of cell signals involved in the recruitment of inflammatory cells [50]. During allergic inflammation, leukocytes are recruited from the blood into the tissues by migrating across the vascular endothelial cells. The migration of leukocytes across endothelial cells is inhibited by pretreatment of the endothelial cells with α-tocopherol and elevated by pretreatment of the endothelial cells with γ-tocopherol [51]. Endothelial cells pretreated with α-tocopherol plus γ-tocopherol result in a phenotype that is not different from the vehicle-treated control endothelial cells [51]. Thus, α-tocopherol and γ-tocopherol have opposing regulatory functions during leukocyte recruitment. These opposing functions of tocopherol isoforms occur through direct regulation of mediators of signal transduction. Briefly, the recruitment of eosinophils to sites of allergic inflammation requires eosinophil binding to adhesion molecules on endothelial cells such as vascular cell adhesion molecule-1 (VCAM-1) and intercellular adhesion molecule-1 (ICAM-1) [6]. These adhesion molecules signal through protein kinase C-α (PKCα) for the recruitment of eosinophils, dendritic cells, lymphocytes, and mast cells [51,52,53]. Upon loading endothelial cells with physiological tocopherol concentrations, which are at the same concentrations of tocopherols as in lung tissues in mice, α-tocopherol inhibits and γ-tocopherol elevates VCAM-1 and ICAM-1 activation of PKCα in endothelial cells [51,52,53]. PKCα is regulated by the tocopherols because, upon binding to the C1A regulatory domain of PKCα, α-tocopherol is an antagonist and γ-tocopherol is an agonist of PKCα activity [50]. α-Tocopherol has been also reported to inhibit PKCα activation in other cell systems or cell extracts, but the mechanisms for inhibition in these systems were not demonstrated [54]. In summary, PKCα is differentially regulated by tocopherol isoforms in endothelial cells, which is critical for leukocyte recruitment in allergic lung inflammation and airway hyper-responsiveness. Thus, α-tocopherol and γ-tocopherol have opposing functions during regulation of allergic responses that utilize PKCα signaling.
Another difference in function of α-tocopherol and γ-tocopherol is that γ-tocopherol, but not α-tocopherol, scavenges reactive nitrogen species (RNS) with peroxynitrite forming 5-nitro-γ-tocopherol [55]. Although α-tocopherol has benefit in allergic eosinophilic inflammation, γ-tocopherol scavenging of RNS may be beneficial by short-term administration for acute inflammation in the lungs with increased RNS, such as neutrophil inflammation induced by ozone or endotoxin [56]. In sheep, nebulized γ-tocopherol reduced neutrophilia, IL-8, and IL-6 in burn and smoke inhalation injury [57]. In other studies, supplementation with mixed tocopherol isoforms containing γ-tocopherol blocks acute endotoxin-stimulated or ozone-stimulated neutrophil inflammation in the rat and human lung [58,59,60,61]. Although γ-tocopherol has been demonstrated to limit responses to ozone, a recent study also indicates that α-tocopherol can also inhibit ozone responses. Acute administration of α-tocopherol at the time of ozone/OVA challenge blocked ozone-induced exacerbation of OVA-stimulated lung neutrophil and eosinophil inflammation [62]. It is reported that γ-tocopherol supplementation reduces antigen induction of rat lung inflammation in which there was severalfold more neutrophils than eosinophils [63] and that high doses of another isoform γ-tocotrienol reduce house dust mite-induced asthma [64]. In asthmatic children exposed to ozone, vitamin E (isoforms not reported) and C supplementation reduced IL-6 in nasal lavages [65]. Therefore, γ-tocopherol may be of benefit for acute neutrophilic inflammation. In a study with ex vivo treatment of human macrophages, 300 μM γ-tocopherol decreased macrophage phagocytosis via modulation of surface receptor activity [57]. However, such highly elevated γ-tocopherol levels are not achievable in vivo. In contrast to reports of γ-tocopherol benefits in acute inflammation or neutrophilic inflammation , it is reported that plasma γ-tocopherol associates with lower lung function in humans [66] and with increased lung eosinophilia and airway hyper-responsiveness in mouse models of allergic asthma [66]. Therefore, chronic consumption of γ-tocopherol may be unfavorable during lung development or during chronic inflammatory diseases such as allergies and asthma. Further clinical studies for mechanisms of tocopherol isoform regulation of allergic lung inflammation are much needed.
Further substantiation of opposing functions of α-tocopherol and γ-tocopherol exists with other chronic inflammatory diseases, including cardiovascular disease and osteoarthritis. In osteoarthritis, plasma γ-tocopherol positively associates with osteoarthritis, whereas plasma α-tocopherol negatively associates with osteoarthritis [67]. In coronary heart disease and stroke, plasma γ-tocopherol is either not associated with heart disease or is associated with an increase in risk for myocardial infarction [68]. In contrast, for α-tocopherol, it is either not associated with heart disease or is associated with reduced death from heart disease [69,70,71,72]. Therefore, for those reports with an effect on heart disease, γ-tocopherol associates with an increase and α-tocopherol associates with a decrease in heart disease.
Asthma and Tocopherol Isoforms in Clinical Studies
Studies on vitamin E and disease outcomes often have focused on one isoform of tocopherol, but the subjects in these clinical studies in fact consume multiple isoforms of tocopherols in differing quantities in the diet, in over-the-counter multivitamins, and in vehicles used in studies with tocopherol supplements. Moreover, there are differences in outcomes of the clinical studies. These differences in clinical effects are consistent when taking into account regional diets, regional plasma tocopherol isoform concentration, multivitamin supplement use, and isoforms administered [20, 73, 74] and when taking into account the mechanistic studies of opposing functions of γ-tocopherol and α-tocopherol [50,51,52, 66, 73, 75]. Therefore, it is essential in clinical studies to determine the plasma or tissue concentration of tocopherol isoforms for adequate interpretation of study results. The tocopherol concentrations in plasma often correlate with those in the lung tissue in humans and in mice [51, 75, 76].
Intriguingly, countries with the highest prevalence rate of asthma tend to have the highest plasma levels of γ-tocopherol [45, 77]. In contrast, the average plasma levels of α-tocopherol are similar among all countries, likely because the liver regulates α-tocopherol uptake by the α-tocopherol transfer protein [45, 77]. The United States has one of the highest rates of asthma, and, in the United States, the average human plasma γ-tocopherol levels are 2–5 times higher than those of most European and Asian countries [45, 77]. These high plasma γ-tocopherol levels in the United States reflect the diet consumption of soybean oil, corn oil, and canola oil, all of which are high in γ-tocopherol per gram of oil (Table 25.1) [45, 77,78,79,80,81,82,83,84,85]. The administration of soybean oil does increase plasma γ-tocopherol levels two- to fivefold in humans and in hamsters [86, 87]. Similar fivefold changes in γ-tocopherol in mice elevate allergen-induced lung inflammation as well as suppress the anti-inflammatory functions of α-tocopherol [51]. In contrast to the United States, most European countries regularly use sunflower oil, safflower oil, and olive oil that are relatively low in γ-tocopherol per gram of oil (Table 25.1) [45, 51, 77, 80]. When administering dietary olive oil to preterm human infants starting 24 h after birth, there is a significant 1.5-fold increase in plasma α-tocopherol as compared to feeding with soybean oil [88]; unfortunately, in this study, plasma concentrations of γ-tocopherol were not reported.
Outcomes for clinical studies of α-tocopherol and asthma differ among countries that consume different dietary oils. In Italy and Finland which preferentially consume safflower oil or olive oil, α-tocopherol supplementation of asthmatic patients resulted in reduced incidence of physician-diagnosed asthma and better lung function as measured by forced expiratory volume in 1 s (FEV1) or wheeze [89,90,91,92,93]. Consistent with this, in two Scottish cohorts, it was reported that reduced maternal intake of vitamin E (likely referring to α-tocopherol) was associated with increased asthma and wheezing in children up to 5 years old [94, 95]. Unfortunately, in the United States or the Netherlands where there is preferential consumption of soybean oil, corn oil, and canola oil, α-tocopherol was not beneficial for adult asthmatic patients [89,90,91,92,93]. This is consistent with opposing functions of γ-tocopherol because the United States and the Netherlands have high plasma levels of γ-tocopherol and high intake of soybean oil (Table 25.1). However, in the United States, some benefit is shown with consumption of very high levels of the acetate form of α-tocopherol. Briefly, oral supplementation with acetate-conjugated α-tocopherol at a very high dose (1500 IU which is 1006 mg) to mild atopic asthmatics in the United States for 16 weeks resulted in increased plasma α-tocopherol, decreased plasma γ-tocopherol, and improved airway responsiveness to methacholine challenge [96]. The studies on vitamin E and lung function have been also examined by meta-analysis, but unfortunately, these analyses have not taken into account the opposing functions of tocopherol isoforms [97]. As would be expected, meta-analyses of data with multiple forms of tocopherols that have opposing functions indicate no association with lung function and wheeze [97]. Thus, we interpret the meta-analysis results [98] as a combination of data from studies with marked variation of vitamin E isoforms that have opposing functions and that were present in the diets, supplements, and supplement vehicles. In contrast to allergic asthma studies in the United States, a study of exercise-induced asthma demonstrated that α-tocopherol supplementation for 3 weeks blocks the exercise-induced transient drop in lung function in humans [99].
As countries adopt Western-like lifestyles and diets , there is increased consumption of soybean oil [100]. In a review by Devereaux et al. of increasing prevalence of asthma and changes in the environment in Scotland [94] from 1967 to 2004, the vegetable oil intake by Scottish significantly increased. We suggest that this would at least result in an increase in γ-tocopherol since vegetable oil (soybean oil) is rich in γ-tocopherol. In England, dietary supplementation with α-tocopherol in soybean oil vehicle to asthmatics had no impact on FEV1, asthma symptom scores, or bronchodilator use [101], which is consistent with γ-tocopherol in soybean oil opposing the function of α-tocopherol. In another study in the United Kingdom, α-tocopherol administration in soybean oil to asthmatics also did not have benefit [102]. Although a consideration is that the clinical studies have differences in asthma prevalence among racial and ethnic groups [103], the studies examining tocopherol association with clinical outcomes generally adjust for several known confounding factors such as gender, age, body mass index, race, and smoking. Even though other differences may occur regarding the environment and genetics of people in different countries, tocopherol isoforms and outcomes for asthma in clinical studies are consistent with the mechanistic studies demonstrating inhibitory functions of α-tocopherol and agonist functions of γ-tocopherol [51]. An interpretation is that differences in outcomes from clinical reports on the associations of vitamin E and asthma may, in part, reflect the opposing regulatory effects of α-tocopherol and γ-tocopherol in the supplements, in the vehicles for the supplements, and in the diets of the individuals.
Opposing regulatory functions of tocopherol isoforms in humans are demonstrated in studies of human lung function in the United States and in China. In these studies, since tocopherol isoforms have opposing functions, the analysis of associations of tocopherol isoforms in clinical studies includes quartiles of plasma tocopherols and the determination of whether there is an association of the tocopherol isoform with the clinical outcome when the concentration of the opposing tocopherol isoform is low and causing the least competing opposing effects. Taking the ratio of α-tocopherol to γ-tocopherol is an unsuitable approach because a high α-tocopherol to high γ-tocopherol ratio that is the same as a low α-tocopherol to low γ-tocopherol ratio is functionally quite different. In a study of individuals in China, high plasma γ-T with low α-T resulted in a fourfold increase in odds ratio for adult asthma in 8 years [104]. In 4526 adults in the United States in the Coronary Artery Risk Development in Young Adults (CARDIA) multicenter cohort , α-tocopherol and γ-tocopherol had opposing associations with lung spirometry [66]. In this study, there were equal numbers of blacks and whites and equal numbers of females and males by study design. Interestingly, increasing serum levels of γ-tocopherol associated with lower FEV1 or FVC, whereas increasing serum levels of α-tocopherol associated with higher FEV1 or FVC [66], with adjustments for several known confounding factors such as gender, age, body mass index, race, and smoking. In the CARDIA cohort, fivefold higher human plasma γ-tocopherol (>10 μM γ-tocopherol) associated with reduced FEV1 and FVC in all participants (asthmatics and non-asthmatics) by age 21–27 years old. This association with decreased FEV1 and FVC before age 21 may occur during development and lung responses to environmental pollutants, allergens, or infections because tocopherol isoforms can directly regulate PKCα during leukocyte recruitment and cell activation [20, 51, 73, 75]. For asthmatics with plasma γ-tocopherol >10 μM, there was 350–570 mL lower FEV1 or FVC as compared to the low to moderate γ-tocopherol concentrations (<10 μM γ-tocopherol) at ages 21–27 [66]. This 10–17% decrease in FEV1 with >10 μM plasma γ-tocopherol in asthmatics is comparable to the 5–10% reduction in FEV1 reported for other environmental factors. In illustration, in asthmatics with occupational allergen exposure, there is a 5–8% decrease in FEV1 compared to non-asthmatics, and this decrease is associated with dyspnea, chest tightness, chronic bronchitis, and chronic cough [105]. Responders to particulate matter have a 2–6% decrease in FEV1 [106], responders to cold or exercise have a 5–11% decrease in FEV1 [107], and responders to house dust mite or dog/cat dander have a 2–8% decrease in FEV1 [108]. With a 2% prevalence of serum γ-tocopherol >10 μM in adults in CARDIA and the adult US population in the 2011 census, 4.5 million adults in the US population may have >10 μM serum γ-tocopherol and have lower FEV1 and FVC. These results are consistent with the preclinical studies demonstrating opposing functions for α-tocopherol and γ-tocopherol in cell signaling, in cell recruitment in allergic responses, and in lung function of allergic mice [32, 33, 50,51,52, 75, 109]. Also in a study in Finland, lower serum α-tocopherol is associated with self-reported asthma [110], and the highest quartile of γ-tocopherol in early childhood (age 1–4) increased the risk of developing asthma [111].
In tissues of asthmatics , tocopherols are reduced. Low plasma α-tocopherol levels are present in adults or children with asthma [90, 91, 112,113,114,115]. It is reported that patients with asthma have reduced α-tocopherol and ascorbic acid in airway fluid but the average plasma concentration of α-tocopherol and ascorbic acid in these patients is normal [112, 113], despite reports in normal individuals that plasma and tissue tocopherols correlate [51, 75, 76]. As in humans, in allergic guinea pigs, α-tocopherol and ascorbic acid levels are decreased in bronchoalveolar lavage [116], and in allergic mice, tocopherol isoforms are reduced in the plasma and lungs [32, 33]. Therefore, since α-tocopherol levels are low in asthmatics and since α-tocopherol can reduce allergic inflammation , supplementation with natural α-tocopherol and maintenance of low dietary levels of γ-tocopherol in combination with other treatments may be an ideal strategy to either prevent or improve control of allergic disease/asthma. A potential target for balance of tocopherol isoforms during allergic disease and asthma may be about 1–1.4 μM plasma γ-tocopherol and 22–30 μM plasma α-tocopherol (Fig. 25.2) as based on average human plasma tocopherol isoforms in countries [45], prevalence of asthma in countries [45], and low α-tocopherol in asthma [90, 91, 112,113,114,115]. Additional intervention studies in humans with analysis of the tocopherol isoforms in plasma and tissues are necessary to assess tocopherol isoform regulation of allergic inflammation and asthma.

A potential balance in human plasma for α-tocopherol (α-T) and γ-tocopherol (γ-T) concentrations during lung development and allergic disease. Additional clinical and mechanistic preclinical studies are needed
Tocopherol Doses in Humans Versus Preclinical Mouse Studies
The tocopherol levels in preclinical animal models of human disease need to be relevant to tocopherol levels in humans. Direct dose comparisons per kg body weight are not sufficient because of large differences in rates of metabolism between mice and humans. Thus, calculations for mouse tocopherol doses that are relevant to humans need to include adjustments for the large differences in lipid metabolic rates and in body weight. Standard mouse basal diet contains about 45 mg α-tocopherol/kg of the diet. The calculation for humans is translated as follows: [(45 mg α-tocopherol/kg of diet) × (1 kg/1000 g) × (6 g diet eaten/mouse/day)/(28 g body weight for an adult mouse)] × 65,000 g human adult = 627 mg α-tocopherol/day for human adult. However, mouse metabolism is about eightfold less efficient, and mice have a higher metabolic turnover rate/kg of body weight than humans [117, 118]. Thus, mice require about eightfold higher intake/g body weight. Furthermore, mice eat 1/6 their body weight in food/day [119] which is considerably higher than the average amount of food/day for adult humans. Thus, to adjust for metabolic rate: (627 mg/day for adult human)/(8 for metabolic rate difference) = 78 mg α-tocopherol/day for human adults. For supplementation levels during disease, a three- to fivefold increase in α-tocopherol for supplementation of mice during studies of lung inflammation (150 or 250 mg α-T/kg of diet for mice) is then 235–392 mg α-tocopherol/day for human adults (calculation: 78 mg/day × (3 or 5)). The 235–392 mg α-tocopherol supplemental doses are well below upper limits suggested for human safety of 1000 mg α-tocopherol/day in pregnancy. Moreover, the supplemental level is near clinical levels in preeclampsia pregnancy trials of 268 mg (400 IU) α-tocopherol [120,121,122,123,124,125]. Also, a supplemented mouse diet with 150 or 250 mg α-tocopherol/kg of diet is reasonable in mice as it is 30–60 times lower than the rodent maternal α-tocopherol diet dose that reduces rodent hippocampus function [126]. Importantly, the doses of 150 mg or 250 mg α-tocopherol/kg of diet for mice achieve two- to threefold increases in tissue concentrations of α-tocopherol, which is similar to fold tissue changes achievable in humans [20, 50,51,52, 73, 75].
Physiological, nontoxic doses of tocopherol isoforms for studies in mice are doses that achieve fold changes in mouse tissues similar to fold changes in human tissues. For healthy adult humans, the recommended daily allowance of α-tocopherol is 15 mg/day. However, because during asthma the levels of tocopherol isoforms are decreased in tissues in humans and mice, it is imperative that studies be done to determine recommended daily doses of tocopherol isoforms during disease. Recommended daily doses of γ-tocopherol have not been addressed. For mice, the standard basal mouse chow diet contains about 45 mg α-tocopherol/kg of diet and 45 mg γ-tocopherol/kg of diet [32, 33]. With equal levels of α-tocopherol and γ-tocopherol in the diet, it results in a tenfold higher tissue α-tocopherol concentration than γ-tocopherol concentration [32, 33] because of the preferential transfer of α-tocopherol by α-TTP in the liver. This is also relevant to studies in humans with mixed tocopherol isoforms. It is important to be aware that the α-tocopherol isoform is preferentially transferred in the liver but nevertheless, γ-tocopherol is potent and, at tenfold lower levels than α-tocopherol in tissues, γ-tocopherol can ablate the benefit of α-tocopherol in allergic responses. Moreover, in humans and mice, a basal rather than deficient α-tocopherol dose is relevant as a baseline, especially in studies of development because α-tocopherol is necessary for mouse and human placental development [46, 47].
In Preclinical Mechanistic Studies in Adult Mice, α-Tocopherol and γ-Tocopherol Have Opposing Functions in the Regulation of Allergic Inflammation and Airway Hyper-responsiveness
As in the design of human studies, in preclinical animal studies, there are differences in the intake of tocopherol isoforms and doses of tocopherols present in the supplementation and in the oil vehicles used for supplementation. For example, Suchankova et al. reported that the administration of purified α-tocopherol in soybean oil by gavage had no major effect on immune parameters or lung airway responsiveness in mice challenged with OVA [127], but tissue levels of tocopherols were not reported. Moreover, an interpretation of this study is that high γ-tocopherol in the soybean oil vehicle opposed the effect of the α-tocopherol. Okamoto et al. [128] found that feeding mice α-tocopherol starting 2 weeks before antigen sensitization did not affect IgE levels but did reduce the number of eosinophils in the bronchoalveolar lavage, but the form and purity of α-tocopherol were not indicated. Mabalirajan et al. [129] reported that oral administration of α-tocopherol in ethanol after antigen sensitization blocked OVA-induced lung inflammation and airway hyper-responsiveness. In this report, α-tocopherol treatment reduced airway hyper-responsiveness and mediators of inflammation including IL-4, IL-5, IL-13, OVA-specific IgE, eotaxin, TGFβ, 12/15-LOX, lipid peroxidation, and lung nitric oxide metabolites [130]. In summary, α-tocopherol supplementation without γ-tocopherol supplementation reduced allergic inflammation.
As in clinical studies, preclinical studies indicate that α-tocopherol supplementation and γ-tocopherol supplementation have opposing effects on allergic inflammation. In a mouse model of allergic lung responses to house dust mite, mouse diet supplemented with 250 mg α-tocopherol/kg of diet during house dust mite challenges reduces eosinophilia in the lung [104]. In contrast, mouse diet supplemented with 250 mg γ-tocopherol/kg of diet elevated house dust mite-induced eosinophilia in the lung [104]. Another mouse model of allergic lung inflammation is induced by intraperitoneal sensitization with chicken egg ovalbumin (OVA) in adjuvant and followed by challenge of the lung with inhaled OVA. Using this model, to determine whether tocopherols regulate the response to the OVA challenge to the lung, tocopherols were administered after OVA sensitization [51]; this is particularly relevant because patients are already sensitized. In this study, tocopherols were administered by daily subcutaneous injections after sensitization but before allergen challenge [51]. Importantly, tocopherols administered subcutaneously or in the diet have the same route of trafficking through the body. By either of these methods of administration, the tocopherols enter the lymph, then the thoracic duct, and then the liver where the tocopherols are loaded on lipoproteins that then enter circulation. The subcutaneous administration of tocopherols reaches plateaus in tissue levels of tocopherols in only a few days, whereas dietary supplementation of tocopherols takes a couple of weeks to achieve a plateau in tissue tocopherol levels [86, 131]. The subcutaneous administration of α-tocopherol or γ-tocopherol raises lung and plasma concentrations of the tocopherol isoform four- to fivefold without affecting body or lung weight [51], and this fold change is clinically relevant because it is achievable in humans. In this study, subcutaneous administration of γ-tocopherol elevates lung eosinophil recruitment by 175%, and α-tocopherol reduces lung eosinophil recruitment by 65% [49]. Furthermore, in these mice, α-tocopherol blocks and γ-tocopherol increases airway hyper-responsiveness [51]. The levels of tocopherols in these studies did not alter numbers of blood eosinophils, indicating that sufficient numbers of eosinophils were available for recruitment [51]. Tocopherol supplementation does not alter the expression of adhesion molecules, cytokines, and chemokines required for the leukocyte recruitment [51]. This modulation of leukocyte infiltration in allergic inflammation, without alteration of adhesion molecules, cytokines, or chemokines, is similar to several other reports of in vivo inhibition of lung inflammation by inhibition of intracellular signals in endothelial cells during leukocyte recruitment [132,133,134].
These pro-inflammatory allergic effects of γ-tocopherol in mice are partially reversible by switching supplements from γ-tocopherol to α-tocopherol at these doses for 4 weeks [75]. There is full reversibility of the elevated inflammatory effects of γ-tocopherol but only with highly elevated levels (10 × supplemental levels) of α-tocopherol [75] that may be potentially risky in humans because very high levels of α-tocopherol may increase the incidence of hemorrhagic stroke and elevate blood pressure [63, 65, 135]. A possibly safer alternative to reverse pro-inflammatory effects of γ-tocopherol on allergic inflammation may be longer supplementation with modest levels of α-tocopherol supplementation.
When α-tocopherol and γ-tocopherol are administered at the same time, γ-tocopherol opposes the anti-inflammatory benefit of α-tocopherol [51, 75]. Supplementation of α-tocopherol plus γ-tocopherol during challenge with OVA does not alter the level of allergic inflammation such that the numbers of lung eosinophils and airway responses are similar to that of the allergic mice with the vehicle control [49]. This suggests that these two tocopherols have competing opposing functions. Moreover, γ-tocopherol is very potent because this opposing function of γ-tocopherol occurs even though γ-tocopherol was about 5–10 times lower in concentration in vivo than α-tocopherol.
In summary, in adult mice, raising tissue concentrations of α-tocopherol fivefold is anti-inflammatory and blocks airway hyper-reactivity, and raising tissue concentrations of γ-tocopherol fivefold is pro-inflammatory and increases airway hyper-reactivity during eosinophilic allergic lung inflammation [20, 50,51,52, 75]. These studies are consistent with the clinical studies, demonstrating that a fivefold increase in human plasma γ-tocopherol associates with a reduction in lung function in adult humans [66]. To relate this to the prevalence of disease, a fivefold difference in plasma γ-tocopherol concentrations is consistent with fivefold higher γ-tocopherol in Americans versus most Western Europeans and Asians (Table 25.1) and higher prevalence of asthma in Americans [45, 77]. Mechanistically, during allergic inflammation in the lung, tocopherol isoforms regulate eosinophil migration on VCAM-1 [5, 6], VCAM-1 signals through PKCα [136], and tocopherols regulate PKCα directly by binding to the C1A regulatory domain of PKCα [50]. Upon binding to PKCα, α-tocopherol is an antagonist of PKCα, and γ-tocopherol is an agonist of PKCα. Thus, a mechanism for the opposing regulatory functions for α-tocopherol and γ-tocopherol on allergic inflammation in adult mice is, at least in part, a result of tocopherol regulation of signals for leukocyte transendothelial migration from the blood into the tissue.
Maternal Tocopherol and Offspring Development of Allergy
Clinical Studies of Maternal Tocopherols and Allergy/Asthma
There has been an increase in the d-γ-tocopherol isoform of vitamin E in the diet and in infant formulas that contain soybean oil [19, 20, 51, 137, 138]. Thus, tocopherol isoforms that regulate allergy and asthma in mothers may affect the risk of development of allergy and asthma in offspring. Some studies suggest that development of allergen responsiveness may occur prenatally [139,140,141] and it is suggested that in utero and early exposures to environmental factors are critical for increased risk of allergic disease [21]. Higher intake of vitamin E is associated with lower odds of wheeze in childhood, but in this analysis, the isoforms of vitamin E are unclear [142]. Increasing maternal α-tocopherol during pregnancy in humans negatively associates with production of inflammatory mediators in vitro in endotoxin-stimulated nasal airway epithelial cells isolated from neonates shortly after birth [143]. Also, maternal plasma α-tocopherol at 11-week gestation associates with reduced asthma treatments in children in the United Kingdom [144]. In studies of human maternal and paternal asthma and development of allergies in offspring, most associations are with maternal allergy/asthma [24, 30, 145,146,147,148,149,150,151], suggesting that sensitization can occur prenatally or early postnatally. There is an association of higher risk of eczema, wheezing, and lower respiratory tract infections in early life with increases in human maternal and cord blood C-reactive protein, which is an acute-phase protein produced during inflammation [152]. It is reported that, by age 21, human plasma α-tocopherol associates with better lung spirometry and human plasma γ-tocopherol associates with worse lung spirometry [66], suggesting that in human development, tocopherols may have early life regulatory functions on responsiveness to allergen and perhaps to other environmental challenges to the lung. It has been demonstrated that maternal α-tocopherol dietary intake is inversely associated with cord blood mononuclear cell proliferative responses to allergen challenge in vitro [140, 153]. Also, from ultrasound studies of the fetus, maternal α-tocopherol levels are reported to associate with fetal growth [154]. In rats, maternal α-tocopherol supplementation during pregnancy results in larger lungs with normal structure in offspring [155]. In several clinical studies, α-tocopherol did not associate with asthma [156,157,158], but these studies did not measure tissue or plasma tocopherol isoforms or include analysis of potential opposing functions of γ-tocopherol. Moreover clinical reports demonstrate that the risk for allergy in children has been associated with mothers with existing allergic disease before conception [24, 30, 145,146,147,148,149,150,151], but whether maternal tocopherol isoforms regulate development of asthma and allergic diseases in offspring needs further study. Specifically, additional clinical studies are needed to address the balance of α-tocopherol and γ-tocopherol in healthy and asthmatic/allergic pregnant women and in infants that may influence the development of risk of allergies and asthma in children.
Preclinical Animal Studies Demonstrating a Maternal Contribution to Offspring Allergy and Asthma
Before discussing tocopherol isoform regulation of maternal influence over development of offspring allergies, it is imperative to first discuss mechanisms of the maternal effect on offspring of allergic mothers. In pregnant mice, tocopherol supplementation of the mother alters the risk of development of allergies in the offspring. The mouse model of maternal transfer of risk of allergy to offspring reflects many of the parameters of development of allergic disease [24, 30, 34, 35, 145,146,147,148,149,150,151, 159, 160], including the fact that the allergic responses of the offspring are not specific to the allergen of the mother [34]. In this mouse model, allergy is induced in adult female mice by sensitizing with intraperitoneal injection of OVA with the adjuvant alum on weeks 1 and 2 and then challenging with aerosolized OVA on weeks 4, 8, and 12 [30, 34,35,36,37,38,39,40, 160]. On the day of the last OVA challenge on week 12, these allergic female mice are mated [30, 33,34,35,36,37,38,39]. Another OVA challenge during mouse pregnancy does not increase risk of allergy in the offspring [34], likely because it takes about 2 weeks for resolution of allergic lung inflammation during the 3-week gestation. The OVA challenge just prior to mating is required for the offspring responsiveness (unpublished observations). The allergic response during pregnancy in mice is consistent with what occurs in humans, as it is anticipated that allergic mothers would have an allergen challenge during their 9-month pregnancy. In the neonatal mouse model, after birth, all of the offspring from allergic mothers and non-allergic mothers are treated with a suboptimal OVA protocol. This suboptimal protocol is comprised of neonates receiving only one instead of two OVA/alum treatments at postnatal days 3–5, and then starting 7 days later, the neonates are challenged with aerosolized OVA for 3 consecutive days [30, 33,34,35,36,37,38,39]. The offspring from allergic mothers develop allergic lung inflammation and airway responsiveness, whereas pups from non-allergic mothers do not develop allergic inflammation (Fig. 25.3). This allergen responsiveness of the offspring of allergic mothers is sustained for up to 8 weeks of age, which is adulthood in the mouse, but then declines after 8 weeks old [35]. The response of the offspring can be abrogated by blocking the allergic response of the mothers at preconception with anti-IL-4 antibody administration [34] or by antibody depletion of T cells in allergic mothers [36]. Further evidence indicates that maternal T cells are sufficient for the maternal effect on offspring because adoptive transfer of allergen-specific T cells from OVA TCR transgenic mice DO11.10 mice to females prior to mating results in offspring with responsiveness to suboptimal challenge of antigen [37]. However, the maternal factors are not entirely clear. IL-4 and IgE which are elevated in the mother do not pass to the fetus [141, 159, 161]. Th2 cytokines (IL-4, IL-5, and IL-13) are elevated in the placenta, but transplacental crossing of these cytokines has not been demonstrated [162,163,164]. Only 2% of maternal GM-CSF crosses the human placenta in ex vivo perfusate studies [165], but whether maternal GM-CSF increases the risk of offspring for allergic responses is not known. Allergens have been reported to perhaps cross the placenta, but it has been demonstrated that offspring are able to respond to BLG, whereas mothers were stimulated with OVA, suggesting that the process is antigen-independent [141]. In another report demonstrating antigen independence of the maternal effect, offspring are responsive to casein, whereas the mothers were sensitized and challenged at preconception with OVA [34]. This antigen-independent effect of maternal allergy on allergen responsiveness in pups is also demonstrated in canines [166]. Similarly to animals, human offspring respond to different allergens than the allergic mother [30]. Thus, the offspring responses are not specific to the allergen that the mother responds to, but instead, the offspring have an increased ability to respond to allergen sensitization. Therefore, female mice that are allergic before conception and develop a Th2 response during pregnancy produce offspring that have augmented responsiveness to suboptimal allergen.

Dendritic cells in maternal transmission of risk for allergy in offspring
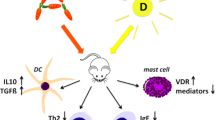
The antigen-independent effect of maternal allergy on allergen responsiveness in pups is a result of changes in dendritic cells of the offspring (Fig. 25.3). The increased responsiveness of the offspring is not through changes in pup macrophages [160]. In adoptive transfer studies, transfer of splenic dendritic cells from non-challenged neonates of allergic mothers into neonates from non-allergic mothers confers increased allergic susceptibility in recipient neonates (Fig. 25.3) [160]. In contrast, the transfer of macrophages from non-challenged neonates of allergic mothers into neonates from non-allergic mothers does not confer increased allergic susceptibility in recipient neonates [160]. The dendritic cell changes include changes in numbers of discrete subsets of dendritic cells [32, 33] and in responses by dendritic cells [160, 167, 168].
There are an increased number of discrete subsets of dendritic cells in offspring of allergic mothers. In the fetal livers from allergic mothers and in the OVA-challenged pup lungs from offspring of allergic mothers, there are increased numbers of CD11b + subsets of CD11c + dendritic cells [169], a dendritic cell subset that is critical for generation of allergic responses [168]. In contrast, in these tissues, there are no changes in CD11b- regulatory dendritic cell subsets, including plasmacytoid dendritic cells and CD103+ dendritic cells [169]. Furthermore, before antigen challenge of the pups, the dendritic cells of pups from allergic mothers had little transcriptional changes but extensive DNA methylation changes [170]. Then, after allergen challenge, there were many transcriptional changes in the dendritic cells of pups of allergic mothers as compared to pups of non-allergic mothers [170]. These studies suggest that maternal mediators, which do not direct allergen specificity, may be transferred from the mother to the offspring and these mediators regulate offspring dendritic cells and heighten the responsiveness of offspring to challenge with suboptimal doses of allergens.
A maternal effect on offspring allergic responses has also been demonstrated for maternal exposure to environmental irritants. Maternal inhalation of titanium oxide or diesel exhaust particles during pregnancy increases responses of offspring to allergen challenge [22]. Also, skin sensitization to toluene diisocyanate (TDI) induces a Th2 response in the mother, and when the mother is mated after a second dose of TDI, the offspring have increased allergic responses to suboptimal OVA [171]. In contrast, a Th1 response in the mother may protect the offspring from developing allergic responses. When females are sensitized to dinitrochlorobenzene (DNCB) which induces a Th1 response and then mated, the offspring do not develop an allergic response to suboptimal OVA [171]. Also, offspring are protected from development of asthma by prenatal challenge of the mother with LPS, which induces a Th1 inflammation, an increase in the Th1 cytokine IFNγ, and a decrease in the Th2 cytokines IL-5 and IL-13 [172,173,174,175]. Injection of non-allergic mothers with IFNγ on gestational day 6.5 protects against the development of allergic responses in offspring [176]. Fedulov et al. [177] demonstrated that treatment of the offspring from allergic mothers on postnatal day 4 with CpG oligonucleotides, a TLR9 agonist and Th1-type stimulant [178], protected the offspring from development of allergic responses to suboptimal OVA challenge. Therefore, exposure of allergic mothers or offspring from allergic mothers to a Th1 stimuli inhibited offspring responses to allergen challenge.
After birth, early in life, breast milk of allergic mother mice may influence development of allergic responses in offspring. However, milk of allergic mothers is not necessary for the offspring allergic responses, because the in utero maternal effects are sufficient for allergic responses by offspring of allergic mothers. Briefly, pups from allergic mothers that are nursed by non-allergic mothers still have an allergic response to suboptimal challenge with OVA [159]. Therefore, maternal effects in utero mediate development of allergen responsiveness in offspring of allergic mothers [159]. However, breast milk is sufficient, but not necessary, for maternal transmission of asthma risk in the offspring because when pups from non-allergic mothers are nursed by allergic mothers, the pups exhibit a response to suboptimal allergen challenge [159]. In that study, the breast milk from allergic and non-allergic mothers contained no detectable IFNγ, IL-2, IL-4, IL-5, IL-13, or TNFα, suggesting that other mediators increase the risk of offspring allergy through breast milk [159]. In clinical studies of other nutrients in breast milk, the mediators, omega-3 and omega-6 polyunsaturated fatty acids, in human milk associate with asthma and atopy, but the mechanism is not known [179, 180]. It is also reported that omega-3 fatty acids during pregnancy associate with lower infantile wheeze [181].
In contrast to studies of allergic mothers, the function of breast milk has been studied for mothers that were not allergic at preconception . In these models, the non-allergic mother mice were exposed during pregnancy or lactation to allergen or to an antigen tolerance protocol. In mouse models where the mother was not allergic at preconception but then exposed during pregnancy or lactation to allergen or an antigen tolerance protocol, there was protection of offspring responses to allergen sensitization/challenge [182]. In these studies, exposure of normal female mice during lactation to OVA results in the transfer of antigen and TGFβ in milk, and this inhibited the development of inflammation of offspring treated later in life as adults (6–8 weeks old) by two sensitizations with OVA/alum and five OVA challenges [182]. These adult offspring had elevated regulatory CD4+ T cells, and the increase in regulatory T cells was dependent on milk TGFβ but not milk immunoglobulins [182], consistent with inhibition of allergic responses as adults. In another approach, it was demonstrated that sensitization of females before mating and then extensive antigen challenges (ten OVA challenges) during lactation resulted in the transfer of IgG immune complexes in the milk and induction of regulatory T cells and tolerance in the offspring when offspring were challenged with OVA at 6–8 weeks old; in this model, immune complexes but not TGFβ in the milk were required for tolerance [183]. In summary, depending on timing, doses, and number of antigen challenges, factors in breast milk can contribute mediators that either increase or decrease offspring responses to allergen . However, consistently, if the mother is allergic before mating, the offspring have elevated responses to allergen.
An endogenous transplacental maternal mediator in allergic mice has been suggested to contribute to the increased responsiveness in the offspring to suboptimal OVA challenge [184]. In adult mice and rats, OVA sensitization and challenge increase stress [185,186,187,188,189,190] and increase endogenous serum corticosterone [191, 192]. Symptoms of stress/anxiety are commonly associated with allergy/asthma in adult mice and in humans [193,194,195,196,197], but whether this contributes to allergic responses in children is not known. During pregnancy, maternal corticosterone is elevated, can cross the placenta, can affect fetal cortisol levels [198, 199], and is a strong inducer of Th2 responses [200, 201]. Cortisol is also present in human breast milk [202] and has the potential to affect allergic responses in neonates. Consistent with the mechanistic studies in mouse models, in pregnant asthmatic women without treatment for asthma, a deficiency in the placenta of a cortisol-metabolizing enzyme 11beta-hydroxysteroid dehydrogenase 2 leads to increased fetal cortisol and low birth weight which is predictive of lower lung function later in life [203, 204]. In adult 4-week-old mice, stress exacerbates OVA-induced allergic responses, and this is blocked by pretreatment with a glucocorticoid receptor antagonist [192]. Subjecting pregnant female mice to stress increases endogenous corticosterone, increases offspring allergic responses to suboptimal allergen , and increases offspring airway responsiveness after suboptimal allergen challenge [184, 198]. Glucocorticoid during pregnancy may be sufficient for allergic responses in offspring because administration of a low dose of glucocorticoid to non-allergic mothers on day 15 of gestation increases offspring allergic responsiveness to suboptimal allergen challenge [184]. When the mother mice are subjected to stress and treated during pregnancy with an inhibitor of endogenous corticosterone synthesis, there is a reduction in the allergic response by the offspring [184]. Therefore, elevated corticosterone in allergic pregnant mice might contribute as a mediator that is transferred from the mother to the fetus or in the breast milk to the neonate, resulting in enhanced responses of offspring to suboptimal allergen challenge. However, the studies with corticosterone were done with BALB/c mice, and in our hands, administration of corticosterone to C57BL/6 does not result in offspring with allergic responses (unpublished observations), even though offspring of allergic C57BL/6 mice consistently exhibit responsiveness to allergen [32, 33]. This suggests that there are mouse strain-specific effects of corticosterone. Thus, perhaps maternal corticosterone contributes, but there are likely additional mechanisms as well. Understanding mechanisms of maternal transfer of risk for allergy to offspring and mechanisms for tocopherol isoform regulation of this risk will have an impact on limiting the development of allergic disease early in life.
In Preclinical Studies, α-Tocopherol Supplementation of the Mother Reduces Allergic Responses in Offspring
The allergic responsiveness of offspring of allergic mothers is inhibited when allergic female mice are bred and then receive α-tocopherol supplementation during the pregnancy/lactation (Fig. 25.4) [169]. In this study, mothers received 250 mg α-tocopherol/kg diet or a basal α-tocopherol diet (45 mg α-tocopherol/kg diet) [169]. A basal α-tocopherol diet is used as the control rather than a tocopherol-deficient diet because adequate α-tocopherol levels are required for placental development and thus the fetus [46, 47]. There are no effects on pup weight, pup numbers, or pup gender distribution by tocopherol supplementation or OVA treatments [169]. The α-tocopherol-supplemented diet increases liver α-tocopherol in the saline-treated mothers threefold as compared to basal diet controls [169], which is consistent with previous reports for this diet in adult female mice [51, 75]. The α-tocopherol tissue concentrations are lower in allergic mothers than non-allergic mothers after α-tocopherol supplementation, which is consistent with reduced α-tocopherol levels in human asthmatics [112,113,114, 205]. In the future, studies of levels for sufficient α-tocopherol supplementation with disease are needed in nonpregnant and pregnant humans, especially since prenatal vitamins often contain multiple tocopherol isoforms. The maternal α-tocopherol supplementation increases pup liver α-tocopherol 2.5-fold [169]. Regarding allergic inflammation in the pups, the maternal α-tocopherol supplementation during pregnancy/lactation results in a dose-dependent inhibition of lung eosinophils [169] in the OVA-challenged pups from allergic mothers as compared to OVA-challenged pups from non-allergic mothers [169]. Maternal α-tocopherol supplementation of allergic female mothers reduces OVA-induced pup lung mRNA expression of cytokines that regulate allergic inflammation (IL-33 and IL-4) and chemokines for eosinophil recruitment (CCL11 and CCL24) [169]. Therefore, α-tocopherol supplementation of allergic mothers inhibits allergic inflammation in OVA-challenged pups from the allergic mothers.

Maternal tocopherol supplementation regulates development of allergic lung inflammation in offspring of allergic mothers
There is a regulatory effect of α-tocopherol in utero and in the milk as determined by cross-fostering pups at birth. Cross-fostering pups from allergic mothers with 250 mg α-tocopherol/kg diet to allergic mothers with basal diet (45 mg α-tocopherol/kg diet) indicated that α-tocopherol supplementation of the allergic mother during pregnancy was sufficient to inhibit the OVA-induced increase in neonate lung eosinophils [169]. In addition, α-tocopherol supplementation during lactation reduces the allergic responses in the neonates [169], suggesting a contribution of α-tocopherol after birth [169]. In summary, α-tocopherol supplementation of allergic mothers during pregnancy is sufficient to reduce development of allergic responses in the offspring.
α-Tocopherol supplementation starting at conception of a second pregnancy of allergic female mice also inhibits development of allergic lung inflammation in offspring. The offspring from allergic mothers that were supplemented with α-tocopherol at the time of a second mating had a > 90% inhibition of lung lavage eosinophils in the OVA-challenged pups [169]. Moreover, in OVA-challenged pups from allergic mothers, α-tocopherol reduces pup lung mRNA expression of several mediators of allergic inflammation: the cytokines IL-4, IL-33, and TSLP and the chemokines CCL11 and CCL24 [169]. There are no effects of maternal α-tocopherol supplementation on pup low levels of the Th1 cytokine IFNγ or the regulatory cytokine IL-10 [169], indicating that α-tocopherol does not switch the response to OVA to a Th1 response.
In Preclinical Studies, γ-Tocopherol Supplementation of Allergic Mothers Highly Elevates Offspring Allergic Responses
Maternal diets supplemented with 250 mg γ-tocopherol/kg diet during pregnancy/lactation increase the maternal liver γ-tocopherol level twofold and the pup liver γ-tocopherol fivefold, consistent with the fold tocopherol changes in human and mouse tissues after supplementation [20, 50,51,52, 73, 75]. This γ-tocopherol supplementation of allergic female mice increased offspring lung eosinophils in response to suboptimal allergen (Fig. 25.4) [33]. It is important to note that γ-tocopherol does not induce allergic inflammation in the OVA-challenged pups from non-allergic mothers [33], indicating that endogenous maternal factors of allergic mothers are required for offspring inflammation. In pups from allergic mothers, maternal d-γ-tocopherol supplementation increases inflammatory mediators including the Th2 mediator amphiregulin, IL-5, CCL11, CCL24, activin A, and GM-CSF. Of concern and potential relevance in humans, maternal γ-tocopherol supplementation also decreases the percentage of female mice that had pups. However, for those females that had pups, there is no affect numbers of pups per litter or pup body weight. It is not known whether γ-tocopherol influences placentation, placental development of fetal development. The data suggest that reduced numbers of mothers with pups and increased pup allergic responses with γ-tocopherol supplementation have potential important implications for children of allergic mothers that consumed γ-tocopherol in the diet or prenatal vitamins as well as for infant formulas supplemented with γ-tocopherol. To better define implications of early life tocopherol regulation in humans, further clinical studies are needed.
In Preclinical Studies, Offspring Dendritic Cells Are Regulated by Maternal γ-Tocopherol and α-Tocopherol Supplementation
The increase in the numbers of dendritic cell subsets in offspring of allergic mothers is regulated by tocopherol isoforms. Maternal supplementation with α-tocopherol reduces the OVA-stimulated pup lung and fetal liver numbers of CD11b + subsets of CD11c + dendritic cells, including resident dendritic cells, myeloid dendritic cells, and CD11b + alveolar dendritic cells, without altering CD11b- subsets of CD11c + dendritic cells, including plasmacytoid dendritic cells, CD103+ dendritic cells, CD11b- alveolar dendritic cells, and alveolar macrophages [169].
Most remarkably, α-tocopherol supplementation does not completely deplete CD11b + dendritic cells, but instead, α-tocopherol supplementation of allergic mothers reduces the numbers of pup CD11b + dendritic cells to the numbers of these dendritic cells in pups from non-allergic mothers [169]. This suggests that α-tocopherol does not block the baseline dendritic cell hematopoiesis but instead may block the signals from allergic mothers that specifically induce the increase in differentiation of CD11c + CD11b + dendritic cell subsets in offspring. Consistent with this, the offspring CD11b- subsets of CD11c + dendritic cells are not altered by maternal supplementation of α-tocopherol [169]. It is also reported that in the pup lungs and fetus of allergic mothers, the changes in CD11c + CD11b + dendritic cells occur without altering the expression of the antigen-presenting molecule MHCII and the costimulatory molecules CD80 and CD86 [169].
In contrast to α-tocopherol supplementation, maternal supplementation with γ-tocopherol increases fetal liver and pup development of CD11c + CD11b + dendritic cells but not numbers of offspring regulatory CD11b- dendritic cell subsets [33]. In addition, with γ-tocopherol supplementation, there is an increase in pup cytokines, chemokines, and lung IRF4 + CD11c + CD11b + dendritic cell subsets which are critical to development of allergic responses. γ-Tocopherol supplementation of the allergic mothers also increases generation of IRF4 + CD11c + CD11b + dendritic cells in the fetal liver [33]. In the fetal livers of γ-tocopherol-supplemented mothers, there are fewer regulatory CD11b-CD11c + pDCs [33], suggesting that with γ-tocopherol supplementation, there may be a reduced control of magnitude of responses to allergen challenge early in life. However, in the OVA-challenged pup lung, the number of pDCs was not altered with γ-tocopherol supplementation [33]. For the fetal liver and pup lung, there was no effect of d-γ-tocopherol on the level of expression per dendritic cell of MHCII, CD80, or IRF4 [33].
Maternal d-γ-tocopherol supplementation also partially increases numbers of resident DCs in the fetus and OVA-challenged pup lung of offspring of non-allergic mothers but not as much as the increase in resident dendritic cells of the OVA-challenged pups from d-γ-tocopherol-supplemented allergic mothers [33]. This is consistent with the increased GM-CSF with γ-tocopherol supplementation in pup lungs from allergic mothers [33]. γ-Tocopherol also increased activin A in pups from non-allergic and allergic mothers [33]. Activin A is a member of the TGFβ superfamily of cytokines and regulates allergic inflammation [206]. Activin A can induce differentiation of monocytes to mDCs, and recruitment of DCs and activin A is produced by several cell types including epithelium, endothelium, mast cells, fibroblasts, and dendritic cells [207]. Therefore, with maternal γ-tocopherol, activin A may function in concert with other mediators to increase numbers of DCs and allergic inflammation. Consistent with γ-tocopherol increasing inflammation, it increases the chemokine CCL11 in pups from allergic and non-allergic mothers, and it increases the chemokine CCL24 and the cytokine IL-5 in the pups from non-allergic mothers [33]. OVA challenge in pups from allergic mothers with d-γ-tocopherol does not result in further increases in CCL24 or IL-5 [33], which may indicate that a maximum response was achieved with OVA challenge. Nevertheless, the pups from the allergic mothers with d-γ-tocopherol have elevated CCL11 and amphiregulin suggesting that in combination, these signals as well as the presence of GM-CSF, CCL24, IL-5, and activin A may function to amplify recruitment of eosinophils in pups from γ-tocopherol-supplemented allergic mothers.
The tocopherol isoforms can directly regulate bone marrow dendritic cell differentiation. α-Tocopherol supplementation during 8-day cultures of GM-CSF-stimulated bone marrow cells reduces the generation of CD45+ CD11b + CD11c + dendritic cells and the number of cells with resident dendritic cell phenotype (CD45 + CD11b + CD11c + Ly6c-MHCII- dendritic cells) without affecting the percentage of viable cells in the culture [169]. γ-Tocopherol also directly regulates hematopoietic development of dendritic cells because d-γ-tocopherol increased the generation of IRF4 + CD11c + CD11b + bone marrow-derived dendritic cells in vitro [33].
In summary, maternal supplementation with γ-tocopherol increases and maternal supplementation with α-tocopherol decreases generation of CD11c + CD11b + DCs and signals for allergic inflammation during development. Studies of tocopherol isoform-specific regulation of inflammation provide a basis toward designing drugs, supplements, and diets that more effectively modulate these pathways in allergic disease. The function of tocopherol isoforms on allergic inflammation and asthma has implications for tocopherol isoforms in prenatal vitamins, in infant formula, and in the diet which may impact risk for allergic disease in future generations. More studies are needed in humans to examine short-term versus long-term outcomes of a range of plasma concentrations of tocopherol isoforms.
Conclusion
The differences in outcomes of clinical studies with tocopherol isoforms are consistent with the mechanistic studies of opposing regulatory functions of these tocopherol isoforms in animal asthma models and in cell cultures with physiological doses of the tocopherol isoforms. The anti-inflammatory function of α-tocopherol and potent pro-inflammatory function γ-tocopherol, at least, reflects the opposing function of these in the regulation of signaling pathways essential to the inflammatory process. Understanding of the differential regulation of inflammation by isoforms of vitamin E provides a basis toward designing drugs and diets that more effectively modulate inflammation and improve lung function in disease in adults and during development.
The rapid increase in rates of asthma implies that the environment influences the generation of asthma and allergic inflammation. Therefore, changes in diet and/or lifestyle could modify disease. Forthcoming studies in preclinical models and clinical studies need to include measurements of tocopherol isoforms in the supplements, vehicles for the supplements, and, most importantly, the plasma and/or tissues before and after intervention. The measurement of plasma or tissue levels is necessary for adequate interpretation of study outcomes. Further studies are necessary to define and provide a basis for recommendations for doses for tocopherol isoforms in healthy individuals and particularly in inflammatory disease states in adult human females and males as well as ethnic groups that differ in prevalence of asthma [208, 209]. The potential of dietary manipulation and supplementation in allergic pregnant mothers and children with asthma requires further work. Interventions in diet early in life, in relation to childhood asthma, raise the possibility of limiting development of allergic disease.
References
Martinez FD. Genes, environments, development and asthma: a reappraisal. Eur Respir J. 2007;29(1):179–84.
Palli D, Masala G, Vineis P, Garte S, Saieva C, Krogh V, et al. Biomarkers of dietary intake of micronutrients modulate DNA adduct levels in healthy adults. Carcinogenesis. 2003;24(4):739–46.
Fiscus LC, Van Herpen J, Steeber DA, Tedder TF, Tang ML. L-Selectin is required for the development of airway hyperresponsiveness but not airway inflammation in a murine model of asthma. J Allergy Clin Immunol. 2001;107(6):1019–24.
Hakugawa J, Bae SJ, Tanaka Y, Katayama I. The inhibitory effect of anti-adhesion molecule antibodies on eosinophil infiltration in cutaneous late phase response in Balb/c mice sensitized with ovalbumin (OVA). J Dermatol. 1997;24:73–9.
Sagara H, Matsuda H, Wada N, Yagita H, Fukuda T, Okumura K, et al. A monoclonal antibody against very late activation antigen-4 inhibits eosinophil accumulation and late asthmatic response in a guinea pig model of asthma. Int Arch Allergy Immunol. 1997;112:287–94.
Chin JE, Hatfield CA, Winterrowd GE, Brashler JR, Vonderfecht SL, Fidler SF, et al. Airway recruitment of leukocytes in mice is dependent on alpha4-integrins and vascular cell adhesion molecule-1. Am J Physiol. 1997;272:L219–L29.
Cook-Mills JM, Deem TL. Active participation of endothelial cells in inflammation. J Leukoc Biol. 2005;77(4):487–95.
Vestweber D. Novel insights into leukocyte extravasation. Curr Opin Hematol. 2012;19(3):212–7.
Muller WA. Mechanisms of leukocyte transendothelial migration. Annu Rev Pathol. 2011;6:323–44.
Yang L, Cohn L, Zhang DH, Homer R, Ray A, Ray P. Essential role of nuclear factor kappaB in the induction of eosinophilia in allergic airway inflammation. J Exp Med. 1998;188:1739–50.
Mould AW, Ramsay AJ, Matthaei KI, Young IG, Rothenberg ME, Foster PS. The effect of IL-5 and eotaxin expression in the lung on eosinophil trafficking and degranulation and the induction of bronchial hyperreactivity. J Immunol. 2000;164:2142–50.
Bousquet J, Bousquet PJ, Godard P, Daures JP. The public health implications of asthma. Bull World Health Organ. 2005;83(7):548–54.
Vollmer WM, Osborne ML, Buist AS. 20-year trends in the prevalence of asthma and chronic airflow obstruction in an HMO. Am J Respir Crit Care Med. 1998;157(4 Pt 1):1079–84.
Friebele E. The attack of asthma. Environ Health Perspect. 1996;104(1):22–5.
van Schayck CP, Smit HA. The prevalence of asthma in children: a reversing trend. Eur Respir J. 2005;26(4):647–50.
CDC National Asthma Control Program. http://www.cdc.gov/asthma/impacts_nation/asthmafactsheet.pdf (2012).
Akinbami LJ, Moorman JE, Bailey C, Zahran HS, King M, Johnson CA, et al. Trends in asthma prevalence, health care use, and mortality in the United States, 2001–2010. NCHS Data Brief. 2012;(94):1–8.
Center For Disease Control Report. http://www.cdc.gov/nchs/data/databriefs/db121.pdf (2013).
Uauy R, Hoffman DR, Birch EE, Birch DG, Jameson DM, Tyson J. Safety and efficacy of omega-3 fatty acids in the nutrition of very low birth weight infants: soy oil and marine oil supplementation of formula. J Pediatr. 1994;124(4):612–20.
Cook-Mills JM, McCary CA. Isoforms of vitamin E differentially regulate inflammation. Endocr Metab Immune Disord Drug Targets. 2010;10:348–66.
Bousquet J, Anto J, Auffray C, Akdis M, Cambon-Thomsen A, Keil T, et al. MeDALL (Mechanisms of the development of ALLergy): an integrated approach from phenotypes to systems medicine. Allergy. 2011;66(5):596–604.
Fedulov AV, Leme A, Yang Z, Dahl M, Lim R, Mariani TJ, et al. Pulmonary exposure to particles during pregnancy causes increased neonatal asthma susceptibility. Am J Respir Cell Mol Biol. 2008;38(1):57–67.
Odaka Y, Nakano M, Tanaka T, Kaburagi T, Yoshino H, Sato-Mito N, et al. The influence of a high-fat dietary environment in the fetal period on postnatal metabolic and immune function. Obesity (Silver Spring). 2010;18(9):1688–94.
Lim RH, Kobzik L, Dahl M. Risk for asthma in offspring of asthmatic mothers versus fathers: a meta-analysis. PLoS One. 2010;5(4):e10134.
Izzotti A, Balansky RM, Cartiglia C, Camoirano A, Longobardi M, De Flora S. Genomic and transcriptional alterations in mouse fetus liver after transplacental exposure to cigarette smoke. FASEB J. 2003;17(9):1127–9.
Vanhees K, Coort S, Ruijters EJ, Godschalk RW, van Schooten FJ. Barjesteh van Waalwijk van Doorn-Khosrovani S. Epigenetics: prenatal exposure to genistein leaves a permanent signature on the hematopoietic lineage. FASEB J. 2011;25(2):797–807.
Rebholz SL, Jones T, Burke KT, Jaeschke A, Tso P, D’Alessio DA, et al. Multiparity leads to obesity and inflammation in mothers and obesity in male offspring. Am J Physiol Endocrinol Metab. 2012;302:E449–57.
Burke KT, Colvin PL, Myatt L, Graf GA, Schroeder F, Woollett LA. Transport of maternal cholesterol to the fetus is affected by maternal plasma cholesterol concentrations in the golden Syrian hamster. J Lipid Res. 2009;50(6):1146–55.
Woollett LA. Maternal cholesterol in fetal development: transport of cholesterol from the maternal to the fetal circulation. Am J Clin Nutr. 2005;82(6):1155–61.
Lim RH, Kobzik L. Maternal transmission of asthma risk. Am J Reprod Immunol. 2009;61(1):1–10.
Liu X, Agerbo E, Schlunssen V, Wright RJ, Li J, Munk-Olsen T. Maternal asthma severity and control during pregnancy and risk of offspring asthma. J Allergy Clin Immunol. 2017;27(17):30854–0.
Abdala-Valencia H, Berdnikovs S, Soveg F, Cook-Mills JM. Alpha-Tocopherol supplementation of allergic female mice inhibits development of CD11c+CD11b+ dendritic cells in utero and allergic inflammation in neonates. Am J Physiol Lung Cell Mol Physiol. 2014;307(00132):L482–96.
Abdala-Valencia H, Soveg F, Cook-Mills JM. γ-Tocopherol supplementation of allergic female mice augments development of CD11c+CD11b+ dendritic cells in utero and allergic inflammation in neonates. Am J Physiol Lung Cell Mol Physiol. 2016;310(8):L759–L71.
Hamada K, Suzaki Y, Goldman A, Ning YY, Goldsmith C, Palecanda A, et al. Allergen-independent maternal transmission of asthma susceptibility. J Immunol. 2003;170(4):1683–9.
Fedulov AV, Leme AS, Kobzik L. Duration of allergic susceptibility in maternal transmission of asthma risk. Am J Reprod Immunol. 2007;58(2):120–8.
Hubeau C, Apostolou I, Kobzik L. Targeting of CD25 and glucocorticoid-induced TNF receptor family-related gene-expressing T cells differentially modulates asthma risk in offspring of asthmatic and normal mother mice. J Immunol. 2007;178(3):1477–87.
Hubeau C, Apostolou I, Kobzik L. Adoptively transferred allergen-specific T cells cause maternal transmission of asthma risk. Am J Pathol. 2006;168(6):1931–9.
Herz U, Joachim R, Ahrens B, Scheffold A, Radbruch A, Renz H. Allergic sensitization and allergen exposure during pregnancy favor the development of atopy in the neonate. Int Arch Allergy Immunol. 2001;124(1-3):193–6.
Herz U, Joachim R, Ahrens B, Scheffold A, Radbruch A, Renz H. Prenatal sensitization in a mouse model. Am J Respir Crit Care Med. 2000;162(3 Pt 2):S62–5.
Jarrett E, Hall E. Selective suppression of IgE antibody responsiveness by maternal influence. Nature. 1979;280(5718):145–7.
Hunter SC, Cahoon EB. Enhancing vitamin E in oilseeds: unraveling tocopherol and tocotrienol biosynthesis. Lipids. 2007;42(2):97–108.
Wolf G. How an increased intake of alpha-tocopherol can suppress the bioavailability of gamma-tocopherol. Nutr Rev. 2006;64(6):295–9.
Leonard SW, Paterson E, Atkinson JK, Ramakrishnan R, Cross CE, Traber MG. Studies in humans using deuterium-labeled alpha- and gamma-tocopherols demonstrate faster plasma gamma-tocopherol disappearance and greater gamma-metabolite production. Free Radic Biol Med. 2005;38(7):857–66.
Bella DL, Schock BC, Lim Y, Leonard SW, Berry C, Cross CE, et al. Regulation of the alpha-tocopherol transfer protein in mice: lack of response to dietary vitamin E or oxidative stress. Lipids. 2006;41(2):105–12.
Cook-Mills JM, Abdala-Valencia H, Hartert T. Two faces of vitamin e in the lung. Am J Respir Crit Care Med. 2013;188(3):279–84.
Muller-Schmehl K, Beninde J, Finckh B, Florian S, Dudenhausen JW, Brigelius-Flohe R, et al. Localization of alpha-tocopherol transfer protein in trophoblast, fetal capillaries’ endothelium and amnion epithelium of human term placenta. Free Radic Res. 2004;38(4):413–20.
Jishage K, Tachibe T, Ito T, Shibata N, Suzuki S, Mori T, et al. Vitamin E is essential for mouse placentation but not for embryonic development itself. Biol Reprod. 2005;73(5):983–7.
Yoshida Y, Saito Y, Jones LS, Shigeri Y. Chemical reactivities and physical effects in comparison between tocopherols and tocotrienols: physiological significance and prospects as antioxidants. J Biosci Bioeng. 2007;104(6):439–45.
Keiko Nishio MH, Akazawa Y, Shichiri M, Iwahashi H, Yoshihisa Hagihara YY, Niki E. Attenuation of lipopolysaccharide (LPS)-induced cytotoxicity by tocopherols and tocotrienols. Redox Biol. 2013;1:97–103.
McCary CA, Yoon Y, Panagabko C, Cho W, Atkinson J, Cook-Mills JM. Vitamin E isoforms directly bind PKCalpha and differentially regulate activation of PKCalpha. Biochem J. 2012;441:189–98.
Berdnikovs S, Abdala-Valencia H, McCary C, Somand M, Cole R, Garcia A, et al. Isoforms of Vitamin E have Opposing Immunoregulatory Functions during Inflammation by Regulating Leukocyte Recruitment. J Immunol. 2009;182:4395–405.
Abdala-Valencia H, Berdnikovs S, Cook-Mills JM. Vitamin E isoforms differentially regulate intercellular adhesion molecule-1 activation of PKCalpha in human microvascular endothelial cells. PLoS One. 2012;7(7):e41054.
Jourd’heuil D, Morise Z, Conner EM, Kurose I, Grisham MB. Oxidant-regulation of gene expression in the chronically inflamed intestine. Keio J Med. 1997;46(1):10–5.
de Luis DA, Armentia A, Aller R, Asensio A, Sedano E, Izaola O, et al. Dietary intake in patients with asthma: a case control study. Nutrition. 2005;21(3):320–4.
Patel A, Liebner F, Netscher T, Mereiter K, Rosenau T. Vitamin E chemistry. Nitration of non-alpha-tocopherols: products and mechanistic considerations. J Org Chem. 2007;72(17):6504–12.
Fakhrzadeh L, Laskin JD, Laskin DL. Ozone-induced production of nitric oxide and TNF-alpha and tissue injury are dependent on NF-kappaB p50. Am J Physiol Lung Cell Mol Physiol. 2004;287(2):L279–85.. Epub 2004 Apr 2.
Wardlaw A. Eosinophil trafficking: new answers to old questions. Clin Exp Allergy. 2004;34(5):676–9.
Hernandez ML, Wagner JG, Aline Kala R, Mills K, Wells HB, Alexis NE, et al. Vitamin E gamma-tocopherol reduces airway neutrophil recruitment after inhaled endotoxin challenge in rats and in healthy volunteers. Free Radic Biol Med. 2013;8(13):00054–3.
Wiser J, Alexis NE, Jiang Q, Wu W, Robinette C, Roubey R, et al. In vivo gamma-tocopherol supplementation decreases systemic oxidative stress and cytokine responses of human monocytes in normal and asthmatic subjects. Free Radic Biol Med. 2008;45(1):40–9.
Wagner JG, Harkema JR, Jiang Q, Illek B, Ames BN, Peden DB. Gamma-tocopherol attenuates ozone-induced exacerbation of allergic rhinosinusitis in rats. Toxicol Pathol. 2009;37(4):481–91.
Burbank AJ, Duran CG, Pan Y, Burns P, Jones S, Jiang Q, et al. Gamma tocopherol-enriched supplement reduces sputum eosinophilia and endotoxin-induced sputum neutrophilia in volunteers with asthma. J Allergy Clin Immunol. 2017;20(17):31110–7.
Duan L, Li J, Ma P, Yang X, Xu S. Vitamin E antagonizes ozone-induced asthma exacerbation in Balb/c mice through the Nrf2 pathway. Food Chem Toxicol. 2017;107(Pt A):47–56.
Wagner JG, Jiang Q, Harkema JR, Ames BN, Illek B, Roubey RA, et al. Gamma-tocopherol prevents airway eosinophilia and mucous cell hyperplasia in experimentally induced allergic rhinitis and asthma. Clin Exp Allergy. 2008;38(3):501–11.
Peh HY, Ho WE, Cheng C, Chan TK, Seow AC, Lim AY, et al. Vitamin E isoform gamma-tocotrienol downregulates house dust mite-induced asthma. J Immunol. 2015;195(2):437–44.
Barthel SR, Johansson MW, McNamee DM, Mosher DF. Roles of integrin activation in eosinophil function and the eosinophilic inflammation of asthma. J Leukoc Biol. 2008;83(1):1–12.
Marchese ME, Kumar R, Colangelo LA, Avila PC, Jacobs DR Jr, Gross M, et al. The vitamin E isoforms alpha-tocopherol and gamma-tocopherol have opposite associations with spirometric parameters: the CARDIA study. Respir Res. 2014;15(1):31.
Jordan JM, De Roos AJ, Renner JB, Luta G, Cohen A, Craft N, et al. A case-control study of serum tocopherol levels and the alpha- to gamma-tocopherol ratio in radiographic knee osteoarthritis: the Johnston County Osteoarthritis Project. Am J Epidemiol. 2004;159(10):968–77.
Dietrich M, Traber MG, Jacques PF, Cross CE, Hu Y, Block G. Does gamma-tocopherol play a role in the primary prevention of heart disease and cancer? A review. J Am Coll Nutr. 2006;25(4):292–9.
Munteanu A, Zingg JM. Cellular, molecular and clinical aspects of vitamin E on atherosclerosis prevention. Mol Asp Med. 2007;28(5-6):538–90.
Siekmeier R, Steffen C, Marz W. Role of oxidants and antioxidants in atherosclerosis: results of in vitro and in vivo investigations. J Cardiovasc Pharmacol Ther. 2007;12(4):265–82.
Meydani M. Vitamin E modulation of cardiovascular disease. Ann N Y Acad Sci. 2004;1031:271–9.
Dutta A, Dutta SK. Vitamin E and its role in the prevention of atherosclerosis and carcinogenesis: a review. J Am Coll Nutr. 2003;22(4):258–68.
Cook-Mills JM, Marchese ME, Abdala-Valencia H. Vascular cell adhesion molecule-1 expression and signaling during disease: regulation by reactive oxygen species and antioxidants. Antioxid Redox Signal. 2011;15(6):1607–38.
Cook-Mills JM. Eosinophil-endothelial cell interactions. Eosinophils in health and disease. In: Lee J, Rosenberg HF, editors. Eosinophils in health and disease. Atlanta: Elsevier; 2012. p. 139–53.
McCary CA, Abdala-Valencia H, Berdnikovs S, Cook-Mills JM. Supplemental and highly elevated tocopherol doses differentially regulate allergic inflammation: reversibility of alpha-tocopherol and gamma-tocopherol’s effects. J Immunol. 2011;186(6):3674–85.
Redlich CA, Grauer JN, Van Bennekum AM, Clever SL, Ponn RB, Blaner WS. Characterization of carotenoid, vitamin A, and alpha-tocopherol levels in human lung tissue and pulmonary macrophages. Am J Respir Crit Care Med. 1996;154:1436–43.
Cook-Mills JM, Avila PC. Vitamin E and D regulation of allergic asthma immunopathogenesis. Int Immunopharmacol. 2014;29(14):007.
Jiang Q, Christen S, Shigenaga MK, Ames BN. gamma-tocopherol, the major form of vitamin E in the US diet, deserves more attention. Am J Clin Nutr. 2001;74(6):714–22.
Talegawkar SA, Johnson EJ, Carithers T, Taylor HA Jr, Bogle ML, Tucker KL. Total alpha-tocopherol intakes are associated with serum alpha-tocopherol concentrations in African American adults. J Nutr. 2007;137(10):2297–303.
Abdala-Valencia H, Berdnikovs S, Cook-Mills JM. Vitamin E isoforms as modulators of lung inflammation. Nutrients. 2013;5(11):4347–63.
Bieri JG, Evarts RP. Tocopherols and fatty acids in American diets. The recommended allowance for vitamin E. J Am Diet Assoc. 1973;62(2):147–51.
Bieri JG, Evarts RP. Vitamin E adequacy of vegetable oils. J Am Diet Assoc. 1975;66(2):134–9.
Choo JH, Nagata M, Sutani A, Kikuchi I, Sakamoto Y. Theophylline attenuates the adhesion of eosinophils to endothelial cells. Int Arch Allergy Immunol. 2003;131(Suppl 1):40–5.
Miller M, Sung KL, Muller WA, Cho JY, Roman M, Castaneda D, et al. Eosinophil tissue recruitment to sites of allergic inflammation in the lung is platelet endothelial cell adhesion molecule independent. J Immunol. 2001;167(4):2292–7.
Davenpeck KL, Berens KL, Dixon RA, Dupre B, Bochner BS. Inhibition of adhesion of human neutrophils and eosinophils to P-selectin by the sialyl Lewis antagonist TBC1269: preferential activity against neutrophil adhesion in vitro. J Allergy Clin Immunol. 2000;105(4):769–75.
Meydani SN, Shapiro AC, Meydani M, Macauley JB, Blumberg JB. Effect of age and dietary fat (fish, corn and coconut oils) on tocopherol status of C57BL/6Nia mice. Lipids. 1987;22:345–50.
Myou S, Zhu X, Boetticher E, Myo S, Meliton A, Lambertino A, et al. Blockade of focal clustering and active conformation in beta 2-integrin-mediated adhesion of eosinophils to intercellular adhesion molecule-1 caused by transduction of HIV TAT-dominant negative Ras. J Immunol. 2002;169(5):2670–6.
Gobel Y, Koletzko B, Bohles HJ, Engelsberger I, Forget D, Le Brun A, et al. Parenteral fat emulsions based on olive and soybean oils: a randomized clinical trial in preterm infants. J Pediatr Gastroenterol Nutr. 2003;37(2):161–7.
Weiss ST. Diet as a risk factor for asthma. Ciba Found Symp. 1997;206:244–57.
Troisi RJ, Willett WC, Weiss ST, Trichopoulos D, Rosner B, Speizer FE. A prospective study of diet and adult-onset asthma [see comments]. Am J Respir Crit Care Med. 1995;151:1401–8.
Dow L, Tracey M, Villar A, Coggon D, Margetts BM, Campbell MJ, et al. Does dietary intake of vitamins C and E influence lung function in older people? Am J Respir Crit Care Med. 1996;154:1401–4.
Smit HA, Grievink L, Tabak C. Dietary influences on chronic obstructive lung disease and asthma: a review of the epidemiological evidence. Proc Nutr Soc. 1999;58:309–19.
Tabak C, Smit HA, Rasanen L, Fidanza F, Menotti A, Nissinen A, et al. Dietary factors and pulmonary function: a cross sectional study in middle aged men from three European countries. Thorax. 1999;54(11):1021–6.
Devereux G. Early life events in asthma--diet. Pediatr Pulmonol. 2007;42(8):663–73.
Martindale S, McNeill G, Devereux G, Campbell D, Russell G, Seaton A. Antioxidant intake in pregnancy in relation to wheeze and eczema in the first two years of life. Am J Respir Crit Care Med. 2005;171(2):121–8.
Hoskins A, Roberts JL 2nd, Milne G, Choi L, Dworski R. Natural-source d-alpha-tocopheryl acetate inhibits oxidant stress and modulates atopic asthma in humans in vivo. Allergy. 2012;67(5):676–82.
Devaraj S, Tang R, Adams-Huet B, Harris A, Seenivasan T, de Lemos JA, et al. Effect of high-dose alpha-tocopherol supplementation on biomarkers of oxidative stress and inflammation and carotid atherosclerosis in patients with coronary artery disease. Am J Clin Nutr. 2007;86(5):1392–8.
Allen S, Britton JR, Leonardi-Bee JA. Association between antioxidant vitamins and asthma outcome measures: systematic review and meta-analysis. Thorax. 2009;64(7):610–9.
Kurti SP, Murphy JD, Ferguson CS, Brown KR, Smith JR, Harms CA. Improved lung function following dietary antioxidant supplementation in exercise-induced asthmatics. Respir Physiol Neurobiol. 2016;220:95–101.
Devereux G, Seaton A. Diet as a risk factor for atopy and asthma. J Allergy Clin Immunol. 2005;115(6):1109–17.
Pearson PJ, Lewis SA, Britton J, Fogarty A. Vitamin E supplements in asthma: a parallel group randomised placebo controlled trial. Thorax. 2004;59(8):652–6.
Hernandez M, Zhou H, Zhou B, Robinette C, Crissman K, Hatch G, et al. Combination treatment with high-dose vitamin C and alpha-tocopherol does not enhance respiratory-tract lining fluid vitamin C levels in asthmatics. Inhal Toxicol. 2009;21(3):173–81.
Zahran HS, Bailey C. Factors associated with asthma prevalence among racial and ethnic groups-United States, 2009-2010 Behavioral Risk Factor Surveillance System. J Asthma. 2013;11:11.
Cook-Mills J, Gebretsadik T, Abdala-Valencia H, Green J, Larkin EK, Dupont WD, et al. Interaction of vitamin E isoforms on asthma and allergic airway disease. Thorax. 2016;71(10):954–6.
Jacobs RR, Boehlecke B, van Hage-Hamsten M, Rylander R. Bronchial reactivity, atopy, and airway response to cotton dust. Am Rev Respir Dis. 1993;148(1):19–24.
Delfino RJ, Quintana PJ, Floro J, Gastanaga VM, Samimi BS, Kleinman MT, et al. Association of FEV1 in asthmatic children with personal and microenvironmental exposure to airborne particulate matter. Environ Health Perspect. 2004;112(8):932–41.
Koskela H, Tukiainen H, Kononoff A, Pekkarinen H. Effect of whole-body exposure to cold and wind on lung function in asthmatic patients. Chest. 1994;105(6):1728–31.
Blanc PD, Eisner MD, Katz PP, Yen IH, Archea C, Earnest G, et al. Impact of the home indoor environment on adult asthma and rhinitis. J Occup Environ Med. 2005;47(4):362–72.
Hess KL, Babcock GF, Askew DS, Cook-Mills JM. A novel flow cytometric method for quantifying phagocytosis of apoptotic cells. Cytometry. 1997;27:145–52.
Elenius V, Palomares O, Waris M, Turunen R, Puhakka T, Ruckert B, et al. The relationship of serum vitamins A, D, E and LL-37 levels with allergic status, tonsillar virus detection and immune response. PLoS One. 2017;12(2):e0172350.
Hamalainen N, Nwaru BI, Erlund I, Takkinen HM, Ahonen S, Toppari J, et al. Serum carotenoid and tocopherol concentrations and risk of asthma in childhood: a nested case-control study. Clin Exp Allergy. 2017;47(3):401–9.
Kalayci O, Besler T, Kilinc K, Sekerel BE, Saraclar Y. Serum levels of antioxidant vitamins (alpha tocopherol, beta carotene, and ascorbic acid) in children with bronchial asthma. Turk J Peds. 2000;42:17–21.
Kelly FJ, Mudway I, Blomberg A, Frew A, Sandstrom T. Altered lung antioxidant status in patients with mild asthma. Lancet. 1999;354:482–3.
Schunemann HJ, Grant BJ, Freudenheim JL, Muti P, Browne RW, Drake JA, et al. The relation of serum levels of antioxidant vitamins C and E, retinol and carotenoids with pulmonary function in the general population. Am J Respir Crit Care Med. 2001;163:1246–55.
Al-Abdulla NO, Al Naama LM, Hassan MK. Antioxidant status in acute asthmatic attack in children. J Pak Med Assoc. 2010;60(12):1023–7.
Ratnasinghe D, Tangrea JA, Forman MR, Hartman T, Gunter EW, Qiao YL, et al. Serum tocopherols, selenium and lung cancer risk among tin miners in China. Cancer Causes Control. 2000;11(2):129–35.
Kleiber M. Metabolic turnover rate: a physiological meaning of the metabolic rate per unit body weight. J Theor Biol. 1975;53(1):199–204.
Terpstra AH. Differences between humans and mice in efficacy of the body fat lowering effect of conjugated linoleic acid: role of metabolic rate. J Nutr. 2001;131(7):2067–8.
Bachmanov AA, Reed DR, Beauchamp GK, Tordoff MG. Food intake, water intake, and drinking spout side preference of 28 mouse strains. Behav Genet. 2002;32(6):435–43.
Villar J, Purwar M, Merialdi M, Zavaleta N, Ngoc TNN, Anthony J, et al. World Health Organisation multicentre randomised trial of supplementation with vitamins C and E among pregnant women at high risk for pre-eclampsia in populations of low nutritional status from developing countries. BJOG. 2009;116(6):780–8.
McCance DR, Holmes VA, Maresh MJ, Patterson CC, Walker JD, Pearson DW, et al. Vitamins C and E for prevention of pre-eclampsia in women with type 1 diabetes (DAPIT): a randomised placebo-controlled trial. Lancet. 2010;376(9737):259–66.
Hauth JC, Clifton RG, Roberts JM, Spong CY, Myatt L, Leveno KJ, et al. Vitamin C and E supplementation to prevent spontaneous preterm birth: a randomized controlled trial. Obstet Gynecol. 2010;116(3):653–8.
Greenough A, Shaheen SO, Shennan A, Seed PT, Poston L. Respiratory outcomes in early childhood following antenatal vitamin C and E supplementation. Thorax. 2010;65(11):998–1003.
Kalpdev A, Saha SC, Dhawan V. Vitamin C and E supplementation does not reduce the risk of superimposed PE in pregnancy. Hypertens Pregnancy. 2011;30(4):447–56.
Gungorduk K, Asicioglu O, Gungorduk OC, Yildirim G, Besimoglu B, Ark C. Does vitamin C and vitamin E supplementation prolong the latency period before delivery following the preterm premature rupture of membranes? A randomized controlled study. Am J Perinatol. 2014;31(3):195–202.
Betti M, Ambrogini P, Minelli A, Floridi A, Lattanzi D, Ciuffoli S, et al. Maternal dietary loads of alpha-tocopherol depress protein kinase C signaling and synaptic plasticity in rat postnatal developing hippocampus and promote permanent deficits in adult offspring. J Nutr Biochem. 2011;22(1):60–70.
Suchankova J, Voprsalova M, Kottova M, Semecky V, Visnovsky P. Effects of oral alpha-tocopherol on lung response in rat model of allergic asthma. Respirology. 2006;11(4):414–21.
Okamoto N, Murata T, Tamai H, Tanaka H, Nagai H. Effects of alpha tocopherol and probucol supplements on allergen-induced airway inflammation and hyperresponsiveness in a mouse model of allergic asthma. Int Arch Allergy Immunol. 2006;141(2):172–80.
Mabalirajan U, Aich J, Leishangthem GD, Sharma SK, Dinda AK, Ghosh B. Effects of vitamin E on mitochondrial dysfunction and asthma features in an experimental allergic murine model. J Appl Physiol. 2009;107(4):1285–92.
Mamdouh Z, Mikhailov A, Muller WA. Transcellular migration of leukocytes is mediated by the endothelial lateral border recycling compartment. J Exp Med. 2009;206(12):2795–808.
Mustacich DJ, Leonard SW, Devereaux MW, Sokol RJ, Traber MG. Alpha-tocopherol regulation of hepatic cytochrome P450s and ABC transporters in rats. Free Radic Biol Med. 2006;41(7):1069–78.
Abdala-Valencia H, Berdnikovs S, McCary CA, Urick D, Mahadevia R, Marchese ME, et al. Inhibition of allergic inflammation by supplementation with 5-hydroxytryptophan. Am J Physiol Lung Cell Mol Physiol. 2012;303:L642–L60.
Abdala-Valencia H, Earwood J, Bansal S, Jansen M, Babcock G, Garvy B, et al. Nonhematopoietic NADPH oxidase regulation of lung eosinophilia and airway hyperresponsiveness in experimentally induced asthma. Am J Physiol Lung Cell Mol Physiol. 2007;292(5):L1111–25.
Keshavan P, Deem TL, Schwemberger SJ, Babcock GF, Cook-Mills JM, Zucker SD. Unconjugated bilirubin inhibits VCAM-1-mediated transendothelial leukocyte migration. J Immunol. 2005;174:3709–18.
Wagner KH, Kamal-Eldin A, Elmadfa I. Gamma-tocopherol – an underestimated vitamin? Ann Nutr Metab. 2004;48(3):169–88.
Abdala-Valencia H, Cook-Mills JM. VCAM-1 signals activate endothelial cell protein kinase Ca via oxidation. J Immunol. 2006;177:6379–87.
Boyle FG, Yuhas RJ, Lien EL. Red blood cell and tissue phospholipid fatty acid profiles of weanling rats fed infant formula fat blends containing soy and/or corn oil. Ann Nutr Metab. 1996;40(4):234–42.
Nelson SE, Rogers RR, Frantz JA, Ziegler EE. Palm olein in infant formula: absorption of fat and minerals by normal infants. Am J Clin Nutr. 1996;64(3):291–6.
Blumer N, Herz U, Wegmann M, Renz H. Prenatal lipopolysaccharide-exposure prevents allergic sensitization and airway inflammation, but not airway responsiveness in a murine model of experimental asthma. Clin Exp Allergy. 2005;35(3):397–402.
Devereux G, Barker RN, Seaton A. Antenatal determinants of neonatal immune responses to allergens. Clin Exp Allergy. 2002;32(1):43–50.
Uthoff H, Spenner A, Reckelkamm W, Ahrens B, Wolk G, Hackler R, et al. Critical role of preconceptional immunization for protective and nonpathological specific immunity in murine neonates. J Immunol. 2003;171(7):3485–92.
Beckhaus AA, Garcia-Marcos L, Forno E, Pacheco-Gonzalez RM, Celedon JC, Castro-Rodriguez JA. Maternal nutrition during pregnancy and risk of asthma, wheeze, and atopic diseases during childhood: a systematic review and meta-analysis. Allergy. 2015;70(12):1588–604.
Miller DR, Turner SW, Spiteri-Cornish D, Scaife AR, Danielian PJ, Devereux GS, et al. Maternal vitamin D and E intakes during early pregnancy are associated with airway epithelial cell responses in neonates. Clin Exp Allergy. 2015;45(5):920–7.
Allan KM, Prabhu N, Craig LC, McNeill G, Kirby B, McLay J, et al. Maternal vitamin D and E intakes during pregnancy are associated with asthma in children. Eur Respir J. 2015;45(4):1027–36.
Kurukulaaratchy RJ, Waterhouse L, Matthews SM, Arshad SH. Are influences during pregnancy associated with wheezing phenotypes during the first decade of life? Acta Paediatr. 2005;94(5):553–8.
Celedon JC, Litonjua AA, Ryan L, Platts-Mills T, Weiss ST, Gold DR. Exposure to cat allergen, maternal history of asthma, and wheezing in first 5 years of life. Lancet. 2002;360(9335):781–2.
Kurukulaaratchy RJ, Matthews S, Waterhouse L, Arshad SH. Factors influencing symptom expression in children with bronchial hyperresponsiveness at 10 years of age. J Allergy Clin Immunol. 2003;112(2):311–6.
Latzin P, Frey U, Roiha HL, Baldwin DN, Regamey N, Strippoli MP, et al. Prospectively assessed incidence, severity, and determinants of respiratory symptoms in the first year of life. Pediatr Pulmonol. 2007;42(1):41–50.
Martinez FD, Wright AL, Taussig LM, Holberg CJ, Halonen M, Morgan WJ. Asthma and wheezing in the first six years of life. The Group Health Medical Associates. N Engl J Med. 1995;332(3):133–8.
Litonjua AA, Carey VJ, Burge HA, Weiss ST, Gold DR. Parental history and the risk for childhood asthma. Does mother confer more risk than father? Am J Respir Crit Care Med. 1998;158(1):176–81.
Folsgaard NV, Chawes BL, Rasmussen MA, Bischoff AL, Carson CG, Stokholm J, et al. Neonatal cytokine profile in the airway mucosal lining fluid is skewed by maternal atopy. Am J Respir Crit Care Med. 2012;185(3):275–80.
Sonnenschein-van der Voort AM, Jaddoe VW, Moll HA, Hofman A, van der Valk RJ, de Jongste JC, et al. Influence of maternal and cord blood C-reactive protein on childhood respiratory symptoms and eczema. Pediatr Allergy Immunol. 2013;24(5):469–75.
Wassall HJ, Devereux G, Seaton A, Barker RN. Complex effects of vitamin E and vitamin C supplementation on in vitro neonatal mononuclear cell responses to allergens. Nutrients. 2013;5(9):3337–51.
Turner SW, Campbell D, Smith N, Craig LC, McNeill G, Forbes SH, et al. Associations between fetal size, maternal {alpha}-tocopherol and childhood asthma. Thorax. 2010;65(5):391–7.
Islam S, Narra V, Cote GM, Manganaro TF, Donahoe PK, Schnitzer JJ. Prenatal vitamin E treatment improves lung growth in fetal rats with congenital diaphragmatic hernia. J Pediatr Surg. 1999;34(1):172–6. discussion 6-7.
Erkkola M, Karppinen M, Javanainen J, Rasanen L, Knip M, Virtanen SM. Validity and reproducibility of a food frequency questionnaire for pregnant Finnish women. Am J Epidemiol. 2001;154(5):466–76.
West CE, Dunstan J, McCarthy S, Metcalfe J, D’Vaz N, Meldrum S, et al. Associations between maternal antioxidant intakes in pregnancy and infant allergic outcomes. Nutrients. 2012;4(11):1747–58.
Maslova E, Hansen S, Strom M, Halldorsson TI, Olsen SF. Maternal intake of vitamins A, E and K in pregnancy and child allergic disease: a longitudinal study from the Danish National Birth Cohort. Br J Nutr. 2014;111(6):1096–108.
Leme AS, Hubeau C, Xiang Y, Goldman A, Hamada K, Suzaki Y, et al. Role of breast milk in a mouse model of maternal transmission of asthma susceptibility. J Immunol. 2006;176(2):762–9.
Fedulov AV, Kobzik L. Allergy risk is mediated by dendritic cells with congenital epigenetic changes. Am J Respir Cell Mol Biol. 2011;44:285–92.
Lim RH, Kobzik L. Transplacental passage of interleukins 4 and 13? PLoS One. 2009;4(3):e4660.
Zourbas S, Dubanchet S, Martal J, Chaouat G. Localization of pro-inflammatory (IL-12, IL-15) and anti-inflammatory (IL-11, IL-13) cytokines at the foetomaternal interface during murine pregnancy. Clin Exp Immunol. 2001;126(3):519–28.
Ostojic S, Dubanchet S, Chaouat G, Abdelkarim M, Truyens C, Capron F. Demonstration of the presence of IL-16, IL-17 and IL-18 at the murine fetomaternal interface during murine pregnancy. Am J Reprod Immunol. 2003;49(2):101–12.
Bowen JM, Chamley L, Mitchell MD, Keelan JA. Cytokines of the placenta and extra-placental membranes: biosynthesis, secretion and roles in establishment of pregnancy in women. Placenta. 2002;23(4):239–56.
Gregor H, Egarter C, Levin D, Sternberger B, Heinze G, Leitich H, et al. The passage of granulocyte-macrophage colony-stimulating factor across the human placenta perfused in vitro. J Soc Gynecol Investig. 1999;6(6):307–10.
Barrett EG, Rudolph K, Bowen LE, Bice DE. Parental allergic status influences the risk of developing allergic sensitization and an asthmatic-like phenotype in canine offspring. Immunology. 2003;110(4):493–500.
van Rijt LS, Lambrecht BN. Dendritic cells in asthma: a function beyond sensitization. Clin Exp Allergy. 2005;35(9):1125–34.
Williams JW, Tjota MY, Clay BS, Vander Lugt B, Bandukwala HS, Hrusch CL, et al. Transcription factor IRF4 drives dendritic cells to promote Th2 differentiation. Nat Commun. 2013;4:2990.
Abdala-Valencia H, Berdnikovs S, Soveg F, Cook-Mills JM. Alpha-tocopherol supplementation of allergic female mice inhibits development of CD11c+CD11b+ dendritic cells in utero and allergic inflammation in neonates. Am J Physiol Lung Cell Mol Physiol. 2014;307(6):L482–96.
Mikhaylova L, Zhang Y, Kobzik L, Fedulov AV. Link between epigenomic alterations and genome-wide aberrant transcriptional response to allergen in dendritic cells conveying maternal asthma risk. PLoS One. 2013;8(8):e70387.
Lim RH, Arredouani MS, Fedulov A, Kobzik L, Hubeau C. Maternal allergic contact dermatitis causes increased asthma risk in offspring. Respir Res. 2007;8:56.
Gerhold K, Avagyan A, Seib C, Frei R, Steinle J, Ahrens B, et al. Prenatal initiation of endotoxin airway exposure prevents subsequent allergen-induced sensitization and airway inflammation in mice. J Allergy Clin Immunol. 2006;118(3):666–73.
Gerhold K, Bluemchen K, Franke A, Stock P, Hamelmann E. Exposure to endotoxin and allergen in early life and its effect on allergen sensitization in mice. J Allergy Clin Immunol. 2003;112(2):389–96.
Tulic MK, Knight DA, Holt PG, Sly PD. Lipopolysaccharide inhibits the late-phase response to allergen by altering nitric oxide synthase activity and interleukin-10. Am J Respir Cell Mol Biol. 2001;24(5):640–6.
Gerhold K, Blumchen K, Bock A, Seib C, Stock P, Kallinich T, et al. Endotoxins prevent murine IgE production, T(H)2 immune responses, and development of airway eosinophilia but not airway hyperreactivity. J Allergy Clin Immunol. 2002;110:110–6.
Lima C, Souza VM, Faquim-Mauro EL, Hoshida MS, Bevilacqua E, Macedo MS, et al. Modulation of the induction of lung and airway allergy in the offspring of IFN-gamma-treated mother mice. J Immunol. 2005;175(6):3554–9.
Fedulov A, Silverman E, Xiang Y, Leme A, Kobzik L. Immunostimulatory CpG oligonucleotides abrogate allergic susceptibility in a murine model of maternal asthma transmission. J Immunol. 2005;175(7):4292–300.
de Brito CA, Fusaro AE, Victor JR, Rigato PO, Goldoni AL, Muniz BP, et al. CpG-induced Th1-type response in the downmodulation of early development of allergy and inhibition of B7 expression on T cells of newborn mice. J Clin Immunol. 2010;30(2):280–91.
Stoney RM, Woods RK, Hosking CS, Hill DJ, Abramson MJ, Thien FC. Maternal breast milk long-chain n-3 fatty acids are associated with increased risk of atopy in breastfed infants. Clin Exp Allergy. 2004;34(2):194–200.
Reichardt P, Muller D, Posselt U, Vorberg B, Diez U, Schlink U, et al. Fatty acids in colostrum from mothers of children at high risk of atopy in relation to clinical and laboratory signs of allergy in the first year of life. Allergy. 2004;59(4):394–400.
Miyake Y, Okubo H, Sasaki S, Tanaka K, Hirota Y. Maternal dietary patterns during pregnancy and risk of wheeze and eczema in Japanese infants aged 16–24 months: the Osaka Maternal and Child Health Study. Pediatr Allergy Immunol. 2011;22(7):734–41.
Verhasselt V, Milcent V, Cazareth J, Kanda A, Fleury S, Dombrowicz D, et al. Breast milk-mediated transfer of an antigen induces tolerance and protection from allergic asthma. Nat Med. 2008;14(2):170–5.
Mosconi E, Rekima A, Seitz-Polski B, Kanda A, Fleury S, Tissandie E, et al. Breast milk immune complexes are potent inducers of oral tolerance in neonates and prevent asthma development. Mucosal Immunol. 2010;3(5):461–74.
Lim R, Fedulov AV, Kobzik L. Maternal stress during pregnancy increases neonatal allergy susceptibility: role of glucocorticoids. Am J Physiol Lung Cell Mol Physiol. 2014;307(2):L141–8.
Costa-Pinto FA, Basso AS, Britto LR, Malucelli BE, Russo M. Avoidance behavior and neural correlates of allergen exposure in a murine model of asthma. Brain Behav Immun. 2005;19(1):52–60.
Costa-Pinto FA, Basso AS, De Sa-Rocha LC, Britto LR, Russo M, Palermo-Neto J. Neural correlates of IgE-mediated allergy. Ann N Y Acad Sci. 2006;1088:116–31.
Portela Cde P, Massoco Cde O, de Lima WT, Palermo-Neto J. Stress-induced increment on total bronchoalveolar cell count in OVA-sensitized rats. Physiol Behav. 2001;72(3):415–20.
Portela CP, Leick-Maldonado EA, Kasahara DI, Prado CM, Calvo-Tiberio IF, Martins MA, et al. Effects of stress and neuropeptides on airway responses in ovalbumin-sensitized rats. Neuroimmunomodulation. 2007;14(2):105–11.
Portela Cde P, Tiberio Ide F, Leick-Maldonado EA, Martins MA, Palermo-Neto J. Effects of diazepam and stress on lung inflammatory response in OVA-sensitized rats. Am J Physiol Lung Cell Mol Physiol. 2002;282(6):L1289–95.
Tonelli LH, Katz M, Kovacsics CE, Gould TD, Joppy B, Hoshino A, et al. Allergic rhinitis induces anxiety-like behavior and altered social interaction in rodents. Brain Behav Immun. 2009;23(6):784–93.
Lu Y, Liu M, Shi S, Jiang H, Yang L, Liu X, et al. Effects of stress in early life on immune functions in rats with asthma and the effects of music therapy. J Asthma. 2010;47(5):526–31.
Chida Y, Sudo N, Sonoda J, Hiramoto T, Kubo C. Early-life psychological stress exacerbates adult mouse asthma via the hypothalamus-pituitary-adrenal axis. Am J Respir Crit Care Med. 2007;175(4):316–22.
Strine TW, Mokdad AH, Balluz LS, Gonzalez O, Crider R, Berry JT, et al. Depression and anxiety in the United States: findings from the 2006 Behavioral Risk Factor Surveillance System. Psychiatr Serv. 2008;59(12):1383–90.
Cheung TK, Lam B, Lam KF, Ip M, Ng C, Kung R, et al. Gastroesophageal reflux disease is associated with poor asthma control, quality of life, and psychological status in Chinese asthma patients. Chest. 2009;135(5):1181–5.
Sansone RA, Sansone LA. Asthma: wheezing, woes, and worries. Psychiatry (Edgmont). 2008;5(10):48–52.
Di Marco F, Verga M, Santus P, Giovannelli F, Busatto P, Neri M, et al. Close correlation between anxiety, depression, and asthma control. Respir Med. 2010;104(1):22–8.
Cordina M, Fenech AG, Vassallo J, Cacciottolo JM. Anxiety and the management of asthma in an adult outpatient population. Ther Adv Respir Dis. 2009;3(5):227–33.
von Hertzen LC. Maternal stress and T-cell differentiation of the developing immune system: possible implications for the development of asthma and atopy. J Allergy Clin Immunol. 2002;109(6):923–8.
Huang CC, Shih MC, Hsu NC, Chien Y, Chung BC. Fetal glucocorticoid synthesis is required for development of fetal adrenal medulla and hypothalamus feedback suppression. Endocrinology. 2012;153(10):4749–56.
Norbiato G, Bevilacqua M, Vago T, Clerici M. Glucocorticoids and Th-1, Th-2 type cytokines in rheumatoid arthritis, osteoarthritis, asthma, atopic dermatitis and AIDS. Clin Exp Rheumatol. 1997;15(3):315–23.
Ramirez F, Fowell DJ, Puklavec M, Simmonds S, Mason D. Glucocorticoids promote a TH2 cytokine response by CD4+ T cells in vitro. J Immunol. 1996;156(7):2406–12.
Groer MW, Humenick S, Hill PD. Characterizations and psychoneuroimmunologic implications of secretory immunoglobulin A and cortisol in preterm and term breast milk. J Perinat Neonatal Nurs. 1994;7(4):42–51.
Murphy VE, Zakar T, Smith R, Giles WB, Gibson PG, Clifton VL. Reduced 11beta-hydroxysteroid dehydrogenase type 2 activity is associated with decreased birth weight centile in pregnancies complicated by asthma. J Clin Endocrinol Metab. 2002;87(4):1660–8.
Murphy VE, Gibson PG, Giles WB, Zakar T, Smith R, Bisits AM, et al. Maternal asthma is associated with reduced female fetal growth. Am J Respir Crit Care Med. 2003;168(11):1317–23.
Shvedova AA, Kisin ER, Kagan VE, Karol MH. Increased lipid peroxidation and decreased antioxidants in lungs of guinea pigs following an allergic pulmonary response. Toxicol Appl Pharmacol. 1995;132:72–81.
Hardy CL, Rolland JM, O’Hehir RE. The immunoregulatory and fibrotic roles of activin A in allergic asthma. Clin Exp Allergy. 2015;45(10):1510–22.
Hedger MP, Winnall WR, Phillips DJ, de Kretser DM. The regulation and functions of activin and follistatin in inflammation and immunity. Vitam Horm. 2011;85:255–97.
Sheikh A, Steiner MF, Cezard G, Bansal N, Fischbacher C, Simpson CR, et al. Ethnic variations in asthma hospital admission, readmission and death: a retrospective, national cohort study of 4.62 million people in Scotland. BMC Med. 2016;14(1):3.
Kim HB, Zhou H, Kim JH, Habre R, Bastain TM, Gilliland FD. Lifetime prevalence of childhood eczema and the effect of indoor environmental factors: analysis in Hispanic and non-Hispanic white children. Allergy Asthma Proc. 2016;37(1):64–71.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2019 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Cook-Mills, J.M. (2019). The Impact of Vitamin E Isoforms on Asthma and Allergy. In: Weber, P., Birringer, M., Blumberg, J., Eggersdorfer, M., Frank, J. (eds) Vitamin E in Human Health. Nutrition and Health. Humana Press, Cham. https://doi.org/10.1007/978-3-030-05315-4_25
Download citation
DOI: https://doi.org/10.1007/978-3-030-05315-4_25
Published:
Publisher Name: Humana Press, Cham
Print ISBN: 978-3-030-05314-7
Online ISBN: 978-3-030-05315-4
eBook Packages: MedicineMedicine (R0)