
Abstract
Tamarisk (Tamarix chinensis), a pioneer native halophyte from the coastal region of North China, has a high potential to establish in high salinity soil. The ecological benefits from planting tamarisk include changes to the structure and function of the saline land ecosystem. However, field experiments on the ecological benefit of using tamarisk in the restoration of saline lands along the coastal areas of China are lacking. In the current study, tamarisk was planted in 2005, 2007, and 2009 to assess the effectiveness of the plant in vegetation and saline soil restoration. The individual biomass, carbon sequestration, species diversity, and soil characteristics of tamarisk plantations in the different planting years were investigated. The results showed that tamarisk grew well with increasing individual biomass during the experimentation period. Carbon density increased from 0.98 t(C) hm−2 (land without tamarisk) to 4.78 t(C) hm−2 in the 3-year plots, 5.56 t(C) hm−2 in the 5-year plots, and 6.89 t(C) hm−2 in the 7-year plots. Significant soil quality improvement was indicated by lower soil salinity and higher organic matter. Salt content in the 0- to 10-cm soil layer of barren land was 10.53 g kg−1, while it was only 3.35 g kg−1, 1.86 g kg−1, and 5.54 g kg−1 in the 3-, 5-, and 7-year plots with tamarisk, respectively. Potassium (K) availability in the soil was increased. In addition, tamarisk played a significant “eco-engine” role in terms of increasing the species of surrounding grasses. Where only 8 grass species existed in this area before tamarisk planting, a total of 15 grass species existed in the 7-year plot. However, the importance value of salt-tolerant species decreased in tamarisk plots. Clonal plants such as Aeluropus sinensis and Sonchus arvensis became more dominant than seed propagation plants such as Suaeda salsa. This suggested that tamarisk plants progressively alleviated salt stress in the coastal environment. The study demonstrated that tamarisk was effective in vegetation regeneration in saline lands, improving soil quality and increasing carbon sequestration.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
Introduction
Saline land in the west bank of Bo Sea Bay of North China is characterized by a shallow groundwater table and high soil salt content, which is usually more than 5 g kg−1. Therefore, a large area of the high salinity land in the area was abandoned (Liu et al. 2010) due to low land productivity . How to improve the land productivity of coastal saline land has become an urgent problem that needs to be addressed.
The planting of native flora in such areas was reported to combat salinity- related issues in these types of soil (Shan et al. 2011, 2012). Tamarisk (Tamarix chinensis) is one of the native halophytes from the coastal area of China. It has a strong potential to utilize phytoremediation to improve soil fertility, which not only helps in reducing soil salinity but also improves biodiversity of plants and microbes. Soil organic matter, nitrogen (N), and phosphorus (P) increased and soil pH decreased (Guan et al. 2009; Hou et al. 2008; Lei et al. 2011; Wang et al. 2006). Hence, tamarisk is widely used in the vegetation restoration of saline land (Zhao et al. 2011).
At present, most studies focus on the application of tamarisk in coastal saline land greening projects. It has been proved that tamarisk is a good greening species because it is a fast-growing shrub and has a high carbon sequestration ability. However, it is difficult to form a tamarisk-dominant community only using natural diffusion, in which case the effect of carbon sequestration and saline soil recovery is not obvious. It is therefore necessary to manually plant tamarisk in saline land to study the changes in structure and function of the saline land ecosystem.
The interaction between plants and the environment is the driving force for plant community succession. After tamarisk planting, the species composition and structure of the plant community changes. Therefore, it is important to study the diversity of herbaceous plant species after planting tamarisk to understand the process of vegetation restoration in high salinity land. There has been little to no research on coastal saline tamarisk planting and its biomass , carbon reserve dynamics, and influence on plant community diversity . The alterations in community biodiversity directly affect the productivity of the saline land and the soil fertility.
In this study, we investigated the properties of the 3 different tamarisk plantations (3-year, 5-year, and 7-year plantations). We studied the biomass dynamic of individual tamarisk plants and tamarisk communities and their distribution pattern. These data were used to evaluate the carbon sequestration potential of planted tamarisks and their influence on the vegetation diversity of the entire community and on the salt soil content.
Study Site
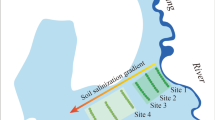
The study site was in the high salinity coastal land in Haixing County of Hebei Province, North China (E117.85°, N38.17°), as shown in Fig. 9.1. The region is a coastal low-lying plain in the west bank of Bo Sea Bay. The soil salt content is composed mainly of chloride (Cl-,45–50% of the total ion content), with sodium (Na+) as the major cation. The groundwater level is 0.3–1.0 m below the surface with groundwater salinity 7–27 g L−1. The climate is a warm, temperate, semi-humid, continental, monsoon one, with an annual average temperature of 12.1 °C. January is the coldest month with an average temperature of −4.5 °C. Annual average precipitation is 582.3 mm, with the precipitation mainly concentrated in July and August. There is little precipitation in winter (November, December, and January).

Study site of the tamarisk plantation in Haixing County, Hebei Province, China, in the west bank of Bo Sea Bay
Due to the distinctive wet and dry seasons, the soil salt content also has a distinct seasonal pattern. High evaporation in spring and autumn cause high salinization while high precipitation washes/dilutes the soil salt in the summer. The winter is cold and dry and salt movement through the soil stops at this time. For this study, the tamarisks were planted in 2005, 2007, and 2009.
Planting method: In March, cuttings of annual tamarisk branches of 0.8~1.2 cm diameter and 12 cm long were planted in plastic bags in a nursery. When the seedlings were approximately 40 cm tall, they were transplanted into the research field and with a spacing of 0.5 m between each seedling. The seedlings were planted at the end of June or before the rainy season.
Methods
In October 2011, a survey was conducted on the different tamarisk plantations (i.e., 3-year, 5-year, and 7-year plantations) and the abandoned land (CK), (Fig. 9.2). Following a random pattern in the planting lattice, the height and ground diameter (the measure point was 5 cm from the ground at the base trunk) of each tamarisk was measured. These data were used to calculate the average height and ground diameter. 6 individual plants were cut and separated into leaves, stems, and roots of the 1 m depth were digged out and washed. The fresh weight of each part was immediately measured. In each tamarisk plot and the abandoned land , 10 quadrats (1 × 1 m sampling square frames) were set randomly to investigate community attributes. The number, height, and coverage of each plant species was determined in the quadrats. The fresh weight of these herbs was also recorded.

The abandoned land and the tamarisk plantation in high salinity soil
Soil samples were taken from 4 soil layers (0–10 cm, 10–20 cm, 20–30 cm, 30–50 cm). The salt content, total nitrogen, available phosphorus, available potassium, and organic matter were determined from the soil samples. The tamarisk and herb samples were dried at 85 °C in an oven until a constant weight was obtained. According to Xu et al.’s (2003) study, the carbon content of different tamarisk organs was used. The carbon content of the above-ground parts was 44.3% and the carbon content of the root was 42.6%. The carbon content of the reference herb was 35.0% (Zheng et al. 2007). The species importance value was used to compare the superiority of different species in the community: importance value = (relative coverage + relative frequency + relative density)/3 × 100 (Song 2001). The redundancy analysis (RDA) of the importance value, the soil salt content, and the planting time of different species was used to explain the relationship between the species and the environment.
Results and Analysis
Growth Characteristics of Tamarisk
Tamarisk height and stem diameter increased with age. The mean height was 150 ± 29 cm, 176 ± 25 cm, and 201 ± 33 cm in the 3-, 5-, and 7-year plantations, respectively. The annual increase in height was approximately 12.5 cm per year. The mean stem diameter was 18.09 ± 4.70 mm, 21.95 ± 4.29 mm, and 26.96 ± 7.75 mm in the 3-, 5-, and 7-year plantations, respectively. The plant diameters in the 5-year to 7-year plantations showed rapid growth. With the increase of age, the relationship between the stem diameter and the height of the tamarisk plant changed from a linear to a logarithmic relationship. The 3-year plants presented a linear relationship; y = 5.62x + 48.74 (R2 = 0.79, n = 53). The 5-year plants had a relatively poor linear relationship; y = 4.51 + 77.59x (R2 = 0.58, n = 43), while the 7-year tamarisk plants showed a logarithmic relationship; y = 99.83 ln(x) – 122.75 (R2 = 0.71, n = 48). This showed that with increasing age, the height growth speed slowed down gradually, and the radial growth of the stem diameter accelerated. This illustrated that the growth of tamarisk shifted activity from the apical meristem to the radial meristem (Figs. 9.3 and 9.4).

Height and diameter of tamarisk plants in the different age plantations

The relationship between the diameter and the height in different age plantations
Changes in Biomass and Carbon Storage of Individual Tamarisk Plants
Biomass
With the increase in age, the biomass of different organs increased gradually. The allocation of biomass to different organs was observed to occur at different rates. Tamarisk plants aged 3 years, 5 years, and 7 years had biomass measurements of 640.24 g, 758.97 g, and 970.63 g, respectively. The shoot to root biomass ratio generally remained unchanged. When comparing the data between the 3-year and 7-year plantations, the biomass of the branches gradually increased (from 33.64% to 48.23%) out of the total plant’s biomass, while the proportion of the root biomass reduced from 52.80% to 42.50%. The ratio between the above-ground and root biomass increased from 0.87 to 1.44. Tamarisk growth in the first 3 years was mainly seen in root elongation and plant height increase, but the radial growth (i.e., stem diameter) accelerated significantly after 3 years and biomass accumulation was mainly focused on the branches. The biomass differences between individual plants gradually increased with age. The biomass of different organs and their proportion in the total biomass is shown in Table 9.1.
Community Carbon Storage
The plants coverage was higher in areas where tamarisk was planted in comparison to land where other herbs were growing. The herb biomass gradually declined over time (Table 9.2). In the bare land of the control plot, the herb biomass had a larger standard deviation but the coverage in the contral plot was higher than the biomass of the herb quadrat taken from the tamarisk stands. This was because tamarisk coverage caused a decrease in the number of herbs.
The mean carbon storage of the abandoned land (CK) was 0.98 t(C) hm−2, but the carbon storage of the 3-year, 5-year, and 7-year tamarisk plantations was 4.78 t(C) hm−2, 5.56 t(C) hm−2, and 6.89 t(C) hm−2, respectively. The trend changed with the change in biomass seen within the individual tamarisk plant. The carbon storage of leaves and branches increased significantly with plant age. The carbon storage increased more slowly in the later period than in the early stage, but its rate of increase was lower than that of the total tamarisk plant. This was because as the canopy density increased gradually, competition between individual plants grew fiercer and some individual plants in a dominant position grew rapidly. Some individual plants grew under the shade of superior plants, and thus their growth was restrained, and they might have even died, so that the density of the tamarisk community declined slightly. Thus, the individual biomass increased at a higher rate than the increase rate of the community’s carbon storage. When viewed from the dynamic process of the tamarisk’s growth, the fastest growth was in the first 3 years, when the carbon accumulation rate was also high, but 3 years later and as a result of increased community canopy density and competition between individual plants, the growth rate and carbon accumulation rate both slowed down.
Soil Salt Content
Tamarisk planting brought about a reduction in the surface evaporation, which slowed the upward movement of soil salt and significantly reduced the soil salt content (Fig. 9.5). In the contrast plot, the soil salt content in the 0- to 10-cm depth of soil was 10.53 g kg−1, which was much higher than that of the land covered by other vegetation. The soil of the 5-year plantation had the lowest soil salinity of 1.86 g kg−1, but it did not decrease further with an increase in time. The 7-year plantation had higher soil salinity than the soil of the 3-year and 5-year plantations. The soil salt content was slightly higher in the 0- to 10-cm layer than in the 10- to 20-cm layer of soil.

Soil water and salt content of the different layers
With increasing plant age, soil organic matter, total nitrogen, and available phosphorus content increased slightly, but no significant difference was observed between the communities. Available potassium content in the soil was significantly increased, as shown in Fig. 9.6.

Organic matter, total N, available P, and available K soil in the 0- to 5-cm and 5- to 10-cm layers in different age plantations. a,b indicate signiifcant differences among different treatments, p<0.05
Herb Species
In the community, there were 17 herbaceous species, belonging to 6 families and 17 genera. There were 7 species of compositae, belonging to 7 different genera, and 6 species of gramineae, belonging to 7 genera. The proportion of the 2 families makes up two-thirds of all species; among them 9 are perennial , 4 are biennial, and 4 are annual . The importance value of each species is shown in Table 9.3. There were only 8 species in the herb community of the contrast plot. The dominant species were halophyte Suaeda salsa and Imperata cylindrica, whose importance values were 34.8 and 36.1 respectively. In the 3-year tamarisk plantation, there were 10 species, with the dominant species being I. cylindrical, Sonchus arvensis, and Aeluropus sinensis. Their importance values were 21.1, 19.0, and 18.8 respectively. In the 5-year tamarisk plantation, there were 12 species, with the dominant species being A. sinensis, S. arvensis, and I. cylindrical. Their importance values were 24.6, 20.0, and 17.7 respectively. In the 7-year tamarisk plantation, there were 15 species. The dominant species were A. sinensis and S. arvensis, whose importance values were 31.3 and 21.3, respectively. It can be seen that tamarisk planting changed the undergrowth environment, promoted the settlement of plants that were not tolerant to saline environments, and increased the species abundance of the entire community. With the increase of tamarisk age, the importance value of A. sinensis, S. arvensis, and Setaria viridis increased. The importance value of saline S. salsa and I. cylindrical gradually reduced, and the importance value of Scorzonera mongolica generally remained unchanged. Plants with a poor salt tolerance, such as Plantago asiatica, Cirsium setosum, Taraxacum mongolicum, and Artemisia capillaris, appeared in the 7-year tamarisk plantation. With regard to the reproduction method of herbaceous plants, with an increase in tamarisk age, the perennial herbaceous plants that reproduced with both seeds and rhizomes were the best suited. This was because the different stages of the plant growth could occupy the undergrowth and use rhizome propagation to become dominant in the community.
RDA analysis on the importance values of species, the soil salinity of each soil layer, and the planting time showed that the importance values of species were mainly affected by the planting time and the soil salinity . As shown in Fig. 9.7, with the horizontal axis as the first axis, the variance was 0.883. With the vertical axis as the second axis, the variance was 0.090 and accumulation was 0.973, and this explains the salt content. According to the species position and direction in the ordination axes, the species can be divided into three types: those with a positive correlation with salt content, those with a positive correlation with time, and those that were not sensitive, which corresponds to salt-tolerant species, species without salt tolerant ability, and neutral species.

The RDA analysis of the importance values and the soil salinity
Discussion
Tamarisk Growth, Biomass, and Community Carbon Storage
The 3-year tamarisk plantation exhibited a rapid growth phase that reached up to 150 cm in height with an average increment of 50 cm per year. Tamarisk cuttings with a height of 40 cm can grow to 80 cm 100 days after cutting during spring (Guo and Wang 2010), while after stumping the new branch, it can grow more than 150 cm with a stem diameter of >8 mm (Fu et al. 2010). This suggests that in their first year, tamarisks grow quickly after cutting. This is similar to the growing process of Caragana microphylla seedlings in arid zones (Zhu et al. 2011).
The 5-year tamarisk plantation presents a relatively poor relationship between stem diameter and plant height. This indicates that after 3 years, the height growth rate slowed down. For the 7-year tamarisk plantation, the relationship between its height and diameter was no longer linear. This indicates that with an increase in age, the growth of tamarisk slows down. Because tamarisk is drought-tolerant, the growth of its seedling root takes precedence over its above-ground part, which is a type of adaptive growth strategy in drought environments (Wei et al. 2007). Tamarisk biomass changes with the growth of the individual plant in the first 3 years, and biomass is mainly allocated in the root. With time, the proportion of stem biomass increases.
Tamarisk planting decreased the biomass of surrounding herbs. This was mainly due to the gradually increased canopy cover from the tamarisk. According to our results, if one harvests the above-ground part of the tamarisk, the highest productivity age should be 3 years. Compared with other vegetation, tamarisk has a biomass equivalent to the planted Hippophae rhamnoides in the Loess Plateau (Liu and Liu 1994), and it is higher than the biomass (780 g m−2) of the natural tamarisk community at the lower reaches of the Heihe River (Peng et al. 2010). This is due to the scattered distribution in nature versus the evenly distributed planting pattern in this study. The biomass of the tamarisk in this study was higher than the theoretical production rate (310 g m−2) of global saline land (Wicke et al. 2011). This illustrates that tamarisk planting in coastal saline land is an effective method of carbon sequestration.
Changes in Tamarisk Community Species Diversity
The change of the salt content in the saline soil was the limiting factor that determined the community succession (Karlin et al. 2011). The increased abundance of herbaceous plants in the community is an important sign of progress succession. This has been previously observed in the succession from saline to non-saline areas in the muddy beach near our study site (Zhao et al. 2008). Two years after the tamarisk planting, herbaceous species had increased in the community and there was an interaction between saline soil vegetation and the saline soil. In northeast China, the reverse succession of saline land was a result of the destruction of vegetation (Wang et al. 2003). With the passing of time, the importance values of salt-tolerant plants such as S. salsa declined. Similarly, after planting Caragana Fabr in Datong, Shanxi Province, the amount of salt-tolerant plants fell sharply in the community (Zhang et al. 2009). S. salsa is a strongly salt-tolerant plant, but it is not as competitive as A. sinensis in soil with low salinity, thus the coverage and number of A. sinensis increased. This result also indicates the sharp decline of competitiveness of saline plants in environments with lower salinity (Ungar 1998). An increase in vegetation coverage inhibits soil salinization in the winter. Thus, low salt-resistant species enhance the diversity of the community, which further decreases the soil salinity . In the 5-year tamarisk plantation, the importance value of A. sinensis was higher than in the 3-year plantation, and species such as Setaria viridis, a low salt-resistant plant, appeared in the community. In the 7-year tamarisk plantation, the number of species was even greater. RDA analysis shows that of the 3 types of plants, salt-tolerant plants mainly included S. salsa and Phragmites australis ; non-salt-tolerant plants were S. viridis and Asiatic plantain; plants that were not sensitive to salt included Scorzonera mongolica. As the age of tamarisk increased in the community, the advantages of root-breeding perennial plants increased gradually. This was because of the niche genetic effect of the root, which gave those plants an advantage in the competition. In the progressive succession of the saline land in the Songnen Plain of northeast China, vegetative propagation occupies a dominant status (Yang and Zheng 1998). Tamarisk planting promotes the progress of coastal saline land succession, gradually reduces the soil salinity , and increases the vegetation community’s richness.
The Effect of Tamarisk on Soil Salinity
Soil salt content changes with the season in coastal saline land . Due to the shallow underground saline water level, surface evaporation forces the salt to move up and accumulate in the soil surface in spring and early summer. Rainfall in the summer causes salt leaching. Increased vegetation coverage and salt leaching during the rainy season are two important ways to reduce the soil salt. Tamarisk planting increases the surface coverage and decreases the surface soil salt in spring by reducing evaporation, which allows more herbaceous plants to grow under the canopy, thereby further increasing the vegetation coverage and reducing the surface water losses and the salinity. This is the cycle of decreasing soil salinity . Therefore, after tamarisk planting, soil salt content was significantly lower than in the contrasting land, which was the result of the interaction between plants and the environment. In different communities, soil salinity increased with the depth of soil, and this was because the sampling time was in early October, which was after the rain and consequent leaching, and the salt content in the soil had leached from the surface to the deeper layers, increasing soil salinity with soil depth. According to a study in Ningxia, tamarisk reduced the soil pH (Lei et al. 2010), but soil pH in this study did not change. The soil in Ningxia was saline-alkaline and plantation tamarisk reduced the pH, while the coastal saline soil already had a low pH, therefore there were no significant changes in the pH in the current study. The study in the desert region of Xinjiang showed an increase in soil salinity below the tamarisk canopy due to the salt-secreting function of the soil between the tamarisk plants (Yin et al. 2008). In addition, multiple factors increase soil salinity in deserts, such as a naturally low salt soil with low rainfall but high evaporation, which accumulates higher salt on the surface. This study demonstrated a significant decrease of soil salt in the 5-year plantation but a slight increase in the 7-year plantation. This may be associated with the background salt content before planting as well as the salt-secreting properties of leaves, although this needs to be investigated further. Without a doubt, tamarisk planting significantly reduced the soil salinity compared with the saline land without tamarisk planting.
The returning of plant branches and leaves to the soil increased the soil nutrient content. In this study, the potassium content of the surface soil significantly increased with plantation age. Although planting tamarisk could promote nutrient accumulation in the surface soil, this saline soil still lacks organic matter, nitrogen, and phosphorus.
Conclusions
Planting tamarisk in the saline soil created a shrub layer and promoted the progressive succession of the plant community. The biomass of tamarisk increased with time. Carbon storage of the 7-year plantation was 6.89 t(C) hm−2, while the carbon storage of bare saline land was 0.98 t(C) hm−2. Salt content of the surface soil significantly decreased after planting tamarisk and the available potassium significantly increased. Herbaceous plant abundance increased and the dominance of salt-tolerant species in the community gradually decreased with time, while perennial herbaceous plants that reproduced with both seeds and rhizomes gradually dominated. T. chinensis is the “ecosystem engine species” in high salinity coastal land, and planting tamarisk is an effective way to improve the utilization of high salinity coastal land.
References
Fu RS, Guan CZ, Yin LR et al (2010) Primary research on growth and development of tow species of Tamarix and application. Tianjin Agric Sci 16(2):112–114
Guan HB, Wang XL, Ju D et al (2009) Soiled modification and application of Tamarix Chinensis on the saline soil. Resour Dev Market 25(10):918–921
Guo L, Wang QL (2010) Cutting experiment of Tarnarix chinensis Lour. on alkaline soil. East China For Manag 24(1):25–27
Hou BD, Ma FY, Wu HY et al (2008) Characteristics of soil nematodes communities at different succession stages of wetland in the Yellow River Delta. Chin J Appl Environ Biol 14(2):202–206
Karlin MS, Bachmeier OA, Dalmasso A et al (2011) Environmental dynamics in salinas grandes, Catamarca, Argentina. Arid Land Res Manag 25(4):328–350
Lei JY, Ban NR, Zhang YH (2011) Effects and partition characteristics of Tamarix ramosissima on nutrients and salt of saline-alkali soils. Bull Soil Water Conserv 31(4):73–78
Liu ZD, Liu ZW (1994) Study on biomass of Hippophae rhamoides and Caragana microphyllal. on loess plateau. Acta Agriculturae Boreali-Occidentalis Sinica 3(2):92–96
Liu XJ, Li XJ, Chen LN et al (2010) Study on the adaptive farming system in saline soils—a case study in saline area of strand plain in Hebei Province. Chin J Eco-Agric 18(4):911–913
Peng SZ, Zhao CY, Peng HH et al (2010) Spatial distribution of Tamarix ramosissima aboveground biomass and water consumption in the lower reaches of Heihe River, Northwest China. Chin J Appl Ecol 20(9):1940–1946
Shan QH, Zhang JF, Ruan WJ et al (2011) Response of soil quality indicators to comprehensive amelioration measures in coastal salt-affected land. Acta Ecol Sin 31(3):6072–6079
Shan QH, Zhang JF, Shen LM et al (2012) Effects of forestry ecological engineering on herb community in coastal saline-alkali land. Chin J Ecol 31(6):1411–1418
Song YC (2001) Vegetation ecology. East China Normal University Press, Shanghai, pp 45–46
Ungar IA (1998) Are biotic factors significant in influencing the distribution of halophytes in saline habitats. Bot Rev 64(2):176–199
Wang HJ, Sheng LX, Chen P et al (2003) Study on influencing factors and control system of reversal evolvement in Western Songnen plain. J Northeast Norm Univ (Natural Science Edition) 35(3):60–65
Wang YZ, Liu YX, Wei CL et al (2006) Improvement of salt-affected soils with six halophytes. J Anhui Agric Sci 34(2):951–957
Wei J, Zhang XM, Maw D et al (2007) Seedling growth dynamics of Tamarix austromongolica and its acclimation strategy in hinterland of desert. Arid Land Geogr 30(2):666–673
Wicke B, Smeets E, Dornburg V et al (2011) The global technical and economic potential of bioenergy from salt-affected soils. Energy Environ Sci 4(8):2669–2681
Xu YR, Zhang WJ, Feng ZW et al (2003) Caloric values, elemental contents and correlations between them of some plants on sea-beach salinity soil in Tianjin. Acta Ecol Sin 23(3):450–455
Yang YF, Zheng HY (1998) Comparison analysis on the experimental communities during progressive succession on alkaline patches in the Songnen plain of China. J Plant Ecol 22(1):214–221
Yin CH, Feng G, Tian CY et al (2008) Enrichment effects of soil organic matter and salinity under the tamarisk shrubs in arid area. Chin J Eco-Agric 16(1):263–265
Zhang LZ, Niu W, Niu Y et al (2009) Impact of Caragana Fabr. plantation on plant community and soil properities of saline-alkali wasteland. Acta Ecol Sin 29(9):4693–4699
Zhao LY, Huang MY, Yuan JJ et al (2008) Research on floral succession in plants protecting engineer of muddy sea wall. Tianjin Agric Sci 14(3):56–60
Zhao KF, Zhang WJ, Fan H et al (2011) Biological measures for utilization and development of salinized soil. Chin J Soil Sci 32(S1):115–119
Zheng WJ, Bao WK, Gu B et al (2007) Carbon concentration and its characteristics in terrestrial higher plants. Chin J Ecol 26(1):307–313
Zhu YL, Wang S, Lin YG et al (2011) Development of Caragana microphylla seedling root system in hilly regions of loess plateau. Bull Soil Water Conserv 31(2):232–237
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2019 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Feng, X., Liu, X., Zhang, X., Li, J.S. (2019). Growth Dynamic of Tamarix chinensis Plantations in High Salinity Coastal Land and Its Ecological Effect. In: Gul, B., Böer, B., Khan, M., Clüsener-Godt, M., Hameed, A. (eds) Sabkha Ecosystems. Tasks for Vegetation Science, vol 49. Springer, Cham. https://doi.org/10.1007/978-3-030-04417-6_9
Download citation
DOI: https://doi.org/10.1007/978-3-030-04417-6_9
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-04416-9
Online ISBN: 978-3-030-04417-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)