
Abstract
The HIV-1-associated neurologic disorders were described earlier in HIV-1 pandemia and are now recognized as a complex of pathologic changes related to the HIV-1 infection of brain perivascular and resident cells, such as microglia. At the same time, the significant contribution to the neurocognitive deficit in humans could be related to the secondary events, such as chronic emotional stress, chronic immune activation due to mucosal barriers deficiency, opportunistic viral and bacterial infections, malnutrition, and other factors that could not be excluded or controlled like on experimental laboratory animals. Animals carrying human hemato-lymphoid tissue and chronically infected with HIV-1 could provide valuable information about the pathogenesis of neurocognitive dysfunction and serve as a model for therapeutic development.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
The HIV-1-associated damage of central and peripheral nervous systems became evident very early in the HIV pandemic [1]. The three clinical forms, now considered as HIV-1-associated neurologic disorders (HAND): asymptomatic, which do not affect human daily life, but could be found by rigorous neuropsychological evaluation; mild cognitive and motor deficit; and dementia. In the era of highly active antiretroviral therapy (HAART), the incidence of dementia reduced and the character of mild neurocognitive impairment changed to the memory/learning and executive control rather than the motor and cognitive speed deficits seen pre-HAART [2]. However, up to now several questions remain unanswered. Is the damage of nervous system occurring primarily due to persistent productive infection within the brain? Is HAND a combination of several connected pathological processes, such as peripheral lymphoid tissue infection or immune suppression? Is it a combination of both events and therefore related to total levels of viral replication? Are neurological symptoms directly related to the neurotoxicity of viral proteins or indirect-metabolic effects of inflammation ? Does chronic immune activation associated with increased permeability of gut contributes to neuronal dysfunction? Does HIV-1 tropism to different cell types (lymphocytes or macrophages) or viral envelope protein coreceptor usage (CCR5 or CXCR4) and ability to bind different types of G-protein-coupled chemokine receptors important for neuronal dysfunction?
Humanized mice on NSG/NOG backgrounds with hemato-lymphoid reconstitution produce human host cells for HIV-1 infection and the infected-lymphocytes and macrophages, are able to enter mouse brain.
Thus it was very attractive to investigate if it is possible to elicit HIV-1-associated neuropathology in humanized animals.
2 Human HIV-1 Infected Cells in the Mouse Brain
HIV-1 virus belongs to Lentiviradae [3, 4].Encephalopathy as well as encephalitis driven by persistent infection of brain resident microglia/macrophages are the cause of neurocognitive deficits, neurodegeneration and dementia. For two other members of the family that induce encephalitis without immune suppression, like the visnamaedi virus and the caprine arthritis encephalitis virus, the major target in the brain are macrophages/microglia [5]. The suggested common features of the pathogenesis of Lentiviruses are their ability to integrate DNA into monocytes/dendritic cells. When these cells reach their final tissue destinations, including brain, they mature to macrophages in the presence of CSF-1 (monocyte-colony stimulating factor, MCSF) and disseminate viral particles and spread infection to microglia/macrophages. Short-term modeling of HIV-1 encephalitis in C.B-17/scid and NOD/scid mice was achieved by transplantation of HIV-1-infected bone marrow-derived human macrophages, with consequential neuropathologies characterized, and this model was applied for antiretroviral and adjunctive therapeutic development [6, 7]. However, this model was lacking important components of the peripheral HIV-1 infection of lymphoid tissues, such as loss of CD4+ cells that contribute to disease progression. Moreover, infection within the brain is secondary to systemic infection and appropriate animal modeling should utilize blood-brain transmission of cell-free virus or infected cells. Perivascular distribution of infected macrophages within the highly vascularized brain are uniquely found in simian immunodeficiency viruses (SIV) infected animals, but for human encephalitic brains up to two third of infected cells are parenchymal microglia [8] . For HIV-1, not all laboratory variants and primary isolates of different viral clades are able to productively infect monocyte/macrophages, and not all viral strains will be able to induce brain pathology [9]. The selection of viruses for humanized mice infection is important, as well as the time of analysis. We were able to observe scattering HIV-1p24+ human lymphocytes through the white-matter tracks, striatum, olfactory bulbs, and their interaction with macrophages in the meninges mostly in mice sacrificed at peak of viremia at 5–7 weeks [10]. Depletion of human CD8+ cells in HIV-1-infected humanized mice also significantly increased the entry of infected cells into the brain and incidence of meningitis [11]. In a rare case of encephalitis, developed in HIV-1-infected humanized mouse after CD8+ cell depletion, HIV-1p24 positive cells with microglial morphology were found. The origin of these cells remains unknown leaving with two predictions: mouse microglia phagocytized human HIV-1 infected cells, or human microglia-like cells became infected (Fig. 25.1). As of today there is no evidence that HIV-1 is productively infecting neurons, astrocytes, oligodendrocytes, epindemal cells of choroid plexus, and brain microvasculature of murine origin. Animals sacrificed at 16–17 weeks after infection occasionally had infected meningeal or perivascular cells. Productive infection of mouse microglial cells is possible, if virus will be delivered by different mechanisms (not human CD4, CCR5, or CXCR4), such as vesicular stomatitis virus (VSV)-pseudotyped [12, 13]. HIV-1 infection of human brain endothelial cells, oligodendrocytes, astrocytes, immature neuroblasts, and mature neurons was suggested and discussed in review [8].

HIV-1p24-positive cells in humanized mouse encephalitic brain. a Representative 5 μ thick horizontal sections of paraffin-embedded mouse brain tissue stained with antibodies to human HLA-DR antigen. Human cells ( brown) are distributed around the vessels, along the meninges and fissures. Magnification 1×. Red squares are areas magnified in panels and b and e, stained for HIV-1p24 antigen. Positive cells are shown in perivascular spaces and white-matter and white-matter tracks. Magnification 10×. c-d, and f-g are corresponding further magnified views of two adjacent sections confirming the microglia-like morphology of HIV-1p24-positive cells indicated with red arrows. Magnification 40×
For the humanized-mouse model for HAND, several questions are unanswered. Are the human cell products associated with chronic HIV-1 infection affecting the mouse endothelial cells and blood-brain-barrier? On the other hand, do we need to transplant human neurons and astrocytes into the mouse brain to confirm the effects induced by virus and human-specific cytokines/chemokines? The transplantation of human neuronal progenitors or fetal brain cells in combination with chronic peripheral HIV-1 infection could be a model to evaluate the infection of neural cell types other than macrophages/microglia cells during human CNS disease. Do we need reconstitution of mouse brain with human microglial cells to mimic HAND and HIV-1-associated encephalitis? Or is the infection of perivascular macrophages and scattering of activated-human T cells enough to induce neuronal dysfunction? Could the compartmentalization of HIV-1 in mouse brain be created and used for evaluation of the special therapeutic strategies for viral eradication [14]?
3 Repopulation of Mouse Brain with Human Microglia
Establishment of a stable viral brain-reservoir is an unmet challenge due to the requirement of a substantial population of human microglial cells within the mouse brain. The nature of mouse microglia as a yolk-sac-born cell and a unique lineage of macrophages was confirmed by several recent studies [15–18]. These cells are developmentally dependent on the transcription factor PU.1 and interferon regulatory factor 8 (IRF8), as well independent of Myb (myeloblastosis) family of transcription factors, DNA-binding protein inhibitor Id2, basic leucine zipper transcription factor, ATF-like 3 (Batf3), and Kruppel-like factor 4 (Klf4) that delineate bone marrowderived monocytic lineage. Microglia also differentiate independently of CSF-1 and are present in CSF-1 KO mice as a result of the IL-34 action. The human microglia seed brain during the embryonic stage. Limited studies about microglia about microglia during human embryonic development detect microglial cells as early as in the fifth week of gestation [19]. Microglia are expected to accumulate parallel to neurovasculogenesis, but the precise markers to distinguish human microglial cells from the later hematopoietic monocytes/macrophages remain unknown.
In mice, fate-mapping studies showed that by one year of age ~ 30 % of microglia-like cells in the brain will be bone-marrow-derived cells [15]. This observation is in concordance with data obtained earlier on NOD/scid-humanized mice when stable and significant human myelopoiesis was established [20]. Accordingly, humanized mice should have someextent of human origin microglia-like cells in the brain. Animals carrying human hematopoiesis survive over a year with maintained meylopoiesis.
Analysis of human myelopoiesis in a one-year-old animal engrafted neonatally with cord-blood-isolated CD34+ cells is shown in Fig. 25.2. The lifelong development of monocytes by retaining lineage-negative CD34+ HSCs in the BM of humanized animals ensure that human monocytes/macrophages are generated and able to enter mouse brain.

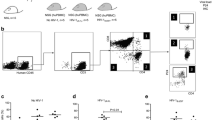
Morphology of human microglia and perivascular macrophages in mouse brain and development of human monocytes in mouse bone marrow. a and b, Representative 5 µ thick sections of paraffin-embedded mouse brain tissue stained for CD14 showing microglia-like cells with different morphologies found in humanized NSG mice at 6–12 months of age. Cells are close to meninges They are ramified and expressed low levels of CD14(a), or had elongated shape and located around the vessel (b). Images were captured under objective 20× and insert under objective 100×. c FACS analysis of the bone marrow cells obtained from a 1 year old humanized mouse created by transplantating cord-blood-derived CD34+ cells at birth. Bone marrow contained 69 % of human CD45+ cells. Several sets of antibodies were used to characterize progenitors and monocyte development. Upper line panels: left panel shows human CD45+, middle panel -CD34+, and right panel -CD34+/CD117+ progenitors and CD117b+ mast cells. Low line panels: left represents myeloid progenitors CD33/CD34+, middle -CD14+ cells from CD45+ gated cells, and right panel highlights the proportion of CD45/CD14/CD4+ promonocytes. (Data were collected using a Beckman Coulter FC500 flow cytometer (Beckman Coulter, Miami, FL) and analyzed using Beckman Coulter Cytomics CXP software (Applied Cytometry Systems, Dinnington, United Kingdom). Results are expressed as percentages of total human CD45+ cells. Courtesy of Professor Samuel Pirrucello, Pathology/Microbiology, UNMC)
These may not be “true” microglial cells and have different properties. Development of these cells requires, in addition to CSF-1, a species-specific cytokine IL-34 that also reacts via CSF-1R [17]. Taken together and resolving around these uncertainties, populating the mouse brain with human microglia requires human microglial precursors and a niche for them in mouse brain.
We confirmed the feasibility of generating human brain-resident microglia and macrophages by transplantation of human fetal brain cells into the brain of newborn irradiated NSG mouse without peripheral transplantation of human CD34+ cells. As shown in Fig. 25.3, human cells with microglial (CD45low) or macrophage (CD163+) morphology are present and distributed outside the clusters of human cells and distinguished by nuclei stained with antibodies to human DNA-PKcs, a molecule that has truncated and reduced expression in scid/scid animals. This observation suggests that appropriate cytokine support (like IL-34), appropriate selection of microglial precursors, and development of strategies to oblate mouse microglia at birth followed by human cell transplantation, are needed to create a stable pool of human cells resident in the mouse brain. Such achievement will provide a novel animal model system susceptible to HIV-1 infection to study viral compartmentalization and CNS penetrating antiretroviral drugs. Although the introduction of HAART has been able to reverse some of the clinical manifestations, pathological alterations persist within the CNS of infected patients, current drug regimens hardly penetrate the CNS. Therefore limited in controlling HIV replication within the brain.

Pattern of the distribution of human microglia and macrophages derived from fetal brain cells. NSG mice were injected at birth with human fetal brain derived mixed cell population cultured for 1 week under serum-free conditions. At 3 months of age, human cells of macrophage/microglia lineage were found without peripheral transplantation of human CD34+ HSC. a and c, Representative distribution of human DNA-PKcs-positive cells around lateral ventricle and along the aqueduct. b Human microglia inside and around the cluster of human immature neurons/astrocytes stained for CD45 with characteristic ramified morphology. d Human CD163-positive cells with round macrophages morphology. Inserts represent magnified views of a single or group of cells. Black and white schematic representation of sagittal brain section at the levels of the captured images depicted as blue rectangles. a magnification 4×, b, c , and d magnification 20×, inserts–—magnification 100×. Sections counterstained with hematoxylin
4 Conclusion and Future Directions
Humanized mice are life-long carriers of human hematopoiesis and human-cell-populated lymphoid tissues susceptible to HIV-1 infection. Limited in the spectrum of brain-resident populations of human cells (macrophages/microglia), they are nonetheless attractive and important tools for HAND research. Introduction of strategies to evaluate in vivo longitudinal changes of HIV-1-infected mouse brain by clinically translatable tools such as magnetic resonance imaging (MRI), magnetic resonance spectroscopy (MRS) [21], diffusion-tensor MR imaging [22, 23], and new experimental imaging methods such as manganese-enhanced MRI (MEMRI) [24] will open new horizons in our understanding of brain pathology and facilitate the development of a new prevention and treatment strategies.
Abbreviations
- CSF-1:
-
Monocyte-colony stimulating factor
- DNA-PKcs:
-
Catalytic subunit of the DNA-dependent protein kinase
- HAND:
-
HIV-1-associated neurologic disorders
- HSC:
-
Hematopoietic stem cells
- IL-2Rγc :
-
Interleukin-2 receptor common gamma chain
- NSG/NOG:
-
NOD/Shi LtJ-scid/IL2Rγnull mice
- scid :
-
Severe combined immune deficiency
- SIRP- α:
-
Signal regulatory protein-α
References
Epstein LG, Sharer LR, Cho ES, Myenhofer M, Navia B, Price RW. HTLV-III/LAV-like retrovirus particles in the brains of patients with AIDS encephalopathy. AIDS Res. 1984;1(6):447–54.
Heaton RK, Franklin DR, Ellis RJ, McCutchan JA, Letendre SL, Leblanc S, et al. HIV-associated neurocognitive disorders before and during the era of combination antiretroviral therapy: differences in rates, nature, and predictors. J Neurovirol. 2010;17(1):3–16.
Koenig S, Gendelman HE, Orenstein JM, Dal Canto MC, Pezeshkpour GH, Yungbluth M, et al. Detection of AIDS virus in macrophages in brain tissue from AIDS patients with encephalopathy. Science. 1986;233(4768):1089–93.
Epstein LG, Sharer LR, Gajdusek DC. Hypothesis: AIDS encephalopathy is due to primary and persistent infection of the brain with a human retrovirus of the lentivirus subfamily. Med Hypotheses. 1986;21(1):87–96.
Blacklaws BA. Small ruminant lentiviruses: immunopathogenesis of visna-maedi and caprine arthritis and encephalitis virus. Comp Immunol Microbiol Infect Dis. 2012;35(3):259–69.
Tyor WR, Power C, Gendelman HE, Markham RB. A model of human immunodeficiency virus encephalitis in scid mice. Proc Natl Acad Sci U S A. 1993;90(18):8658–62.
Persidsky Y, Limoges J, McComb R, Bock P, Baldwin T, Tyor W, et al. Human immunodeficiency virus encephalitis in SCID mice. Am J Pathol 1996;149(3):1027–53.
Bissel SJ, Wiley CA. Human immunodeficiency virus infection of the brain: pitfalls in evaluating infected/affected cell populations. Brain Pathol. 2004;14(1):97–108.
Koppensteiner H, Brack-Werner R, Schindler M. Macrophages and their relevance in human immunodeficiency virus type I infection. Retrovirology. 2012;9:82.
Gorantla S, Makarov E, Finke-Dwyer J, Castanedo A, Holguin A, Gebhart CL, et al. Links between progressive HIV-1 infection of humanized mice and viral neuropathogenesis. Am J Pathol. 2010;177(6):2938–49.
Gorantla S, Makarov E, Finke-Dwyer J, Gebhart CL, Domm W, Dewhurst S, et al. CD8 + cell depletion accelerates HIV-1 immunopathology in humanized mice. J Immunol. 2010;184(12):7082–91.
Dou H, Morehead J, Bradley J, Gorantla S, Ellison B, Kingsley J, et al. Neuropathologic and neuroinflammatory activities of HIV-1-infected human astrocytes in murine brain. Glia. 2006;54(2):81–93.
Gorantla S, Liu J, Sneller H, Dou H, Holguin A, Smith L, et al. Copolymer-1 induces adaptive immune anti-inflammatory glial and neuroprotective responses in a murine model of HIV-1 encephalitis. J Immunol. 2007;179(7):4345–56.
Forsman A, Weiss RA. Why is HIV a pathogen? Trends Microbiol. 2008;16(12):555–60.
Ginhoux F, Greter M, Leboeuf M, Nandi S, See P, Gokhan S, et al. Fate mapping analysis reveals that adult microglia derive from primitive macrophages. Science. 2010;330(6005):841–5.
Horiuchi M, Wakayama K, Itoh A, Kawai K, Pleasure D, Ozato K, et al. Interferon regulatory factor 8/interferon consensus sequence binding protein is a critical transcription factor for the physiological phenotype of microglia. J Neuroinflammation. 2012;9:227.
Wang Y, Szretter KJ, Vermi W, Gilfillan S, Rossini C, Cella M, et al. IL-34 is a tissuerestricted ligand of CSF1R required for the development of Langerhans cells and microglia. Nat Immunol. 2012;13(8):753–60.
Kierdorf K, Erny D, Goldmann T, Sander V, Schulz C, Perdiguero EG, et al. Microglia emerge from erythromyeloid precursors via Pu.1- and Irf8-dependent pathways. Nat Neurosci. 2013;16(3):273–80.
Verney C, Monier A, Fallet-Bianco C, Gressens P. Early microglial colonization of the human forebrain and possible involvement in periventricular white-matter injury of preterm infants. J Anat. 2010;217(4):436–48.
Asheuer M, Pflumio F, Benhamida S, Dubart-Kupperschmitt A, Fouquet F, Imai Y, et al. Human CD34 + cells differentiate into microglia and express recombinant therapeutic protein. Proc Natl Acad Sci U S A. 2004;101(10):3557–62.
Dash PK, Gorantla S, Gendelman HE, Knibbe J, Casale GP, Makarov E, et al. Loss of neuronal integrity during progressive HIV-1 infection of humanized mice. J Neurosci. 2011;31(9):3148–57.
Towgood KJ, Pitkanen M, Kulasegaram R, Fradera A, Kumar A, Soni S, et al. Mapping the brain in younger and older asymptomatic HIV-1 men: frontal volume changes in the absence of other cortical or diffusion tensor abnormalities. Cortex. 2012;48(2):230–41.
Muller-Oehring EM, Schulte T, Rosenbloom MJ, Pfefferbaum A, Sullivan EV. Callosal degradation in HIV-1 infection predicts hierarchical perception: a DTI study. Neuropsychologia. 2010;48(4):1133–43.
Liu Y, Sajja BR, Gendelman HE, Boska MD. Mouse brain fixation to preserve In vivo manganese enhancement for ex vivo manganese-enhanced MRI. J Magn Reson Imaging. 24 Jan 2013. doi:10.1002/jmri.24005.
Acknowledgments
This contribution was supported by the Chair of the Department of Pharmacology and Experimental Neuroscience Dr. Howard E. Gendelman and grants P01 NS043985 DHHS/NIH/NINDS, P01 DA028555 DHHS/NIH/NIDA, R01AG043540. The authors would like to thank Edward Makarov, Jaclyn Knibbe-Hollinger, Tanuja Gutti, Prashanta Dash, Aditya Bade, and Sidra Akhter for ongoing contributions in humanized mice for NeuroAIDS projects.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer Science+Business Media New York
About this chapter
Cite this chapter
Poluektova, L., Epstein, A., Gorantla, S. (2014). Brain HIV-1 Infection Modeling in Humanized Mice. In: Poluektova, L., Garcia, J., Koyanagi, Y., Manz, M., Tager, A. (eds) Humanized Mice for HIV Research. Springer, New York, NY. https://doi.org/10.1007/978-1-4939-1655-9_25
Download citation
DOI: https://doi.org/10.1007/978-1-4939-1655-9_25
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4939-1654-2
Online ISBN: 978-1-4939-1655-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)