
Abstract
Articular cartilage defects are among the most common disabling conditions of humans in the western world. The most prominent risk factors are age, weight, and trauma. Osteoarthritis is the typical outcome of chronic articular cartilage defects and up to date cartilage regeneration remains elusive. The discovery of amniotic fluid-derived stem cells has opened a multitude of new therapeutic options, one of them being their use for novel stem cell-based tissue regeneration techniques in order to treat cartilage defects. Here, we summarize developmental stages and growth factors as well as extracellular molecules involved in articular cartilage formation as well as degeneration. Furthermore, we discuss recent advances in the use of amniotic fluid stem cells as novel cell sources for the generation of cartilage tissue and how the endogenous cartilage formation process could be recapitulated during artificial tissue engineering.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Mesenchymal Stem Cell
- Articular Cartilage
- Cartilage Defect
- Chondrogenic Differentiation
- Autologous Chondrocyte Implantation
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
Articular cartilage starts to form during fetal joint development and subsequently covers the surface at the bony ends of articular joints, where it is maintained throughout life. Its main function is to reduce friction and therefore to facilitate movement of the joint. Defects within articular cartilage are the reason for osteoarthritis (OA), which is also described as a primarily noninflammatory, degenerative joint disease. The main histologic features of OA include disruption of articular cartilage, degradation of extracellular matrix, and reduced cellularity within cartilage [1]. The incidence of osteoarthritis is still increasing and it is the single most common cause of disability in the elderly population [2]. Cartilage is an avascular and aneural tissue and hence regeneration especially after partial injury, which does not penetrate to the subchondral bone, is limited due to poor recruitment of extrinsic cells. Today, marrow stimulation, autograft, and allograft-based techniques are applied to treat severe cartilage defects [3–5], but all of these procedures suffer from several drawbacks. Bone marrow stimulation like microfracture-based treatment results in the production of fibrocartilage with inferior mechanical stability compared to native cartilage. For autograft, transplantation-based treatment the patient has to undergo multiple surgeries, once to isolate chondrocytes from less weight-bearing areas of the joint and another time for re-transplantation of in vitro expanded cells. In addition, in vitro expansion of chondrocytes is associated with dedifferentiation processes, which reduce the ability of cells to produce extracellular matrix [6, 7]. Stem cells isolated from various adult tissues have been tested for their use as cell sources to regenerate chondrogenic tissue [8]. Transplantation of differentiated mesenchymal stem cells remains a promising approach for regenerative therapies, but often the tissue generated in vivo shows lower matrix deposition and, consequently, less mechanical stability as compared to chondrocyte-based transplantations [9, 10]. Amniotic fluid stem (AFS) cells possess properties that make them ideal candidates for cell-based therapy and in particular for the treatment of cartilage defects. AFS cells are genetically stable, immune-privileged, non-tumorigenic and they can be bio-banked [11, 12]. Here we describe the current knowledge on the differentiation of AFS cells towards a chondrogenic phenotype. We focus on AFS cell marker expression, use of chondrogenic differentiation factors, and briefly discuss scaffolds used for tissue generation. In vitro differentiation of AFS cells is supposed to work best if developmental processes that give rise to permanent native cartilage are recapitulated. Therefore we also review developmental processes known to be involved in articular cartilage formation, but also in degeneration.
2 Development of Articular Cartilage
Synovial joint formation is initiated with the establishment of the interzone at the prospective joint site consisting of densely packed, flattened mesenchymal cells interconnected by gap junctions [13]. The interzone plays a dual role by providing progenitor cells for all tissues constituting joints including articular cartilage, ligaments, and the synovial membrane as well as the signaling center by preventing chondrocyte differentiation at the prospective joint site and by controlling maturation of chondrocytes within adjacent future skeletal elements [14–16]. How interzone cells or subsets of them contribute to different joint tissues is not clear yet since the interzone is species-specifically organized which impedes extracting a common mechanism concerning the developmental fate of interzone cells. For example, the interzone of chicks, a model organism for developmental studies, consists of three layers [17] whereas that of humans represents a non-structured thin flattened layer [13, 18, 19]. Moreover, the developmental origin of these cells is still a matter of debate and is associated with a crucial and early event in skeletogenesis by endochondral ossification, the mesenchymal condensation, which subsequently gives rise to the cartilaginous anlagen [20]. Two concepts concerning the time of patterning of the future skeletal elements and hence the appearance of the interzone have been proposed by different authors (reviewed in [20] and citations therein). Those imply that interzone cells might either originate from the cartilaginous anlagen requiring localized cellular re-differentiation and matrix removal or from non-condensed mesenchymal cells in the area of the adjacent ends of preformed bone templates. The latter is supported by Pacifici et al. who showed that peri-articular cells actively migrate into the incipient joint and suggested that they contribute to or entirely constitute the interzone [21].
Joint formation can be divided into interzone formation, cavitation, and establishment of permanent articular cartilage (see also Table 8.1). Interzone development is controlled by BMP, TGF-β, and WNT signaling pathways [22]. GDF5 is a member of the TGF-β superfamily and is an early marker expressed by cells of the interzone and in the immediate proximity of the prospective joint [16, 23]. GDF5 is pivotal for joint maintenance rather than for induction of joint formation since a dominant-negative mutation in GDF5 indeed impairs but its ectopic expression is not sufficient to induce joint formation [24, 25]. Sox11 might contribute to the regulation of GDF5 expression in joint development although its ectopic expression cannot induce GDF5 [26]. GDF5 binds to and oligomerizes type I and type II serine-threonine kinase receptors and signals through phosphorylation of transcription factors of the SMAD family [27]. Two type I receptors are expressed in the interzone and perichondrium, BMPRIA and BMPRIB, but GDF5 specifically binds to and signals through BMPRIB [28, 29]. The importance of GDF5-BMPRIB-mediated signaling is underlined by the fact that mutations in either one or the other lead to similar skeletal malformations (e.g., brachydactyly, symphalangism) and that many mutations of GDF5 are in its receptor-binding loop [29]. Notably, BMPRIA expression seems to be important for cartilage homeostasis and repair since conditional knockout leads to postnatal erosion of articular cartilage of normally developed joints in mice [30]. In addition, GDF6 and GDF7 are expressed at joint sites distinct from that of GDF5 and inactivation of GDF6 causes defects in the formation of joints, ligaments, and cartilage distinct from that observed due to GDF5 mutation indicating a joint-specific role of the different members of the BMP family [31]. Noggin is a BMP antagonist and binds to and thereby limits the activity of GDF5 [32]. This interaction is important for joint development since certain mutations in NOG and GDF5 are associated with symphalangism (SYM1) and multiple synostosis syndrome (SYNS1) [32, 33].
WNT9a (former WNT14) is pivotal for joint formation. In developing chick limbs, it is expressed at the prospective joint site appearing as a transverse stripe at stage 27, which separates as joint development progresses to form two stripes possibly setting the interzone boundaries. Eventually, WNT9a is detected in the fibrous part and the synovial lining of the joint capsule [34]. However, the notion that WNT9a is sufficient to induce joint formation as proposed by some authors has recently been challenged by Später et al. [35, 36]. The latter showed that Wnt9a acts cooperatively with WNT4 to suppress chondrogenic differentiation of interzone cells and that the fusion of the three carpal elements observed in Wnt9a:Wnt4 double mutant mice was not due to the failing of joint induction [37].
A critical step in joint formation is cavitation, the formation of the joint space between opposing skeletal elements. It precedes the morphogenesis of synovial joint structures. The importance of mechanical stimuli for cavitation by movement of the developing joint has early been appreciated. In ovo application of neuromuscular blocking agents leading to either flaccid or rigid paralysis revealed the differential impact of dynamic and static stimuli for initiation of cavitation and maintenance of the joint cavity [38]. The molecular link to mechanostimulation is the expression of hyaluronic acid (HA). Indeed, the lack of static and dynamic stimuli prevents cavity formation and leads to reduced cartilage volume occurring concomitantly with reduced amounts of HA in the interzone and the lack of menisci and ligaments [38]. HA emerges at the interzone concomitant with cavitation [39]. Moreover, the activity of uridine diphosphoglucose dehydrogenase (UDPGD) which synthesizes precursors of HA precedes cavitation and is maintained within articular surfaces and synovium during cavitation [40], and CD44, an HA receptor, is expressed by interzone cells. The sequestering of HA-binding proteins (HABPs) to high-molecular HA is suggested to be crucial for cavitation since HA oligosaccharides applied in ovo exert an effect similar to the lack of mechanical stimulation by preventing cavitation and being hallmarked by decreased CD44 expression and UDPGD activity [41].
3 Native Articular Cartilage Organization
Articular cartilage is avascular and nutrient supply occurs by diffusion. It has been postulated that nutrition is mediated by synovial fluid as well as diffusion through calcified cartilage and the subchondral bone [42]. Especially synovial fluid plays a critical role in maintaining adult normal cartilage homeostasis [43].
The importance of mechanical stimulation in the regeneration has been shown through zone-specific gene expression after stimulatory effects of compression [44–46]. Mature articular cartilage consists of three zones, which constitute also different subpopulations of chondrocytes: the superficial zone (SZ), the transitional or middle zone (MZ), and the radial zone or deep zone (DZ). The superficial or tangential zone (10–20 %) harbors small and flat chondrocytes and collagen fibrils are orientated parallel to the cartilage surface. Cells within this zone produce and secret lubricin and proteoglycan 4, which are the most important joint lubricant superficial zone proteins [47, 48]. Human articular cartilage was predicted to not contain any source of stem cells because of the avascular architecture. However, recent discovered studies show phenotypic and functional properties of mesenchymal stem or progenitor cell populations in the SZ of mature articular cartilage [49–51]. These cells are characterized by expression of mesenchymal markers CD105, CD166, Notch-1, STRO-1, and VCAM-1 [52]. Superficial zone cells are positive for alpha-smooth muscle actin, which is also present in progenitor cells [53, 54].
The middle zone represents the thickest part (40–60 %) of the cartilage where collagen fibers are less organized and the pericellular matrix (PCM) is located. The PCM is a thin layer of extracellular matrix that immediately surrounds the chondrocyte. It plays a major role in the communication between chondrocytes and extracellular matrix [55]. The PCM is best studied in the form of chondrons which are chondrocytes clustered in one unit and form a functional and metabolic microenvironment. The PCM is different from the rest of the extracellular matrix in cartilage concerning vicinity, matrix density, and matrix composition. Collagen type VI proteins built up a fibrillar network in PCM [56]. Further identified PCM proteins are triosephosphate isomerase, transforming growth factor-ß induced protein, peroxiredoxin-4, ADAM (A disintegrin and metalloproteinases) 28, and latent-transforming growth factor ß-binding protein-2 [57].
The deep zone is characterized by ellipsoid cells and fibers, orientated perpendicular to the surface and oxygen tension is below 1 % [58]. Underneath the DZ is the tidemark to calcified cartilage and subchondral bone.
4 Articular Cartilage Degeneration
Age, trauma, and disease are the factors that facilitate joint cartilage damage, which subsequently can lead to osteoarthritis (OA) formation. The so-called primary OA develops without any known cause, but genetic, biomechanical, or biochemical factors often play a role [59, 60]. OA is a big burden of society since the majority of individuals over the age of 65 show evidence for OA on hands, knees, hips, and spine which cause symptoms like inflammation, pain, and loss of mobility [61]. All joint structures are affected by OA, but articular cartilage is affected most by degeneration. In the initial phase of the disease chondrocytes become clustered and show increased cell proliferation. When OA persits, proteinases like MMPs and aggrecanases are expressed and type II collagen and proteoglycans undergo changes in composition and are eventually degraded [62]. The excessive reorganization of hyaline cartilage finally leads to fibrillation of the surface and localized production of fibrocartilage which contributes to the observed joint stiffness during OA. Progressive OA is accompanied by hypertrophy of articular chondrocytes as evidenced by a thickening of the zone of calcified cartilage [63]. Hypertrophy is hallmarked by release of pro-angiogenic as well as osteogenic factors with subsequent apoptosis of chondrocytes [64].
Hypertrophy in articular chondrocytes is thought to recapitulate hypertrophy during endochondral ossification. During bone formation chondrocytes undergo several developmental stages which end with post-mitotic hypertrophy [65]. Upon hypertrophy, growth plate chondrocytes downregulate SOX9 and type II collagen expression, whereas RUNX2, type X collagen, alkaline phosphatase, and VEGF-A are up-regulated [66–68]. Research on articular cartilage regeneration has profited from studies on endochondral ossification because the mechanisms of hypertrophy induction are now better understood and it has become clear that for permanent cartilage formation hypertrophy induction has to be avoided.
5 Novel Approaches for the Treatment of Degenerative Joint Disorders
Articular cartilage defects can be divided into full thickness and partial thickness defects. In the first group not only cartilaginous tissue is lost but also part of the subchondral bone mass. This kind of defect will trigger an intrinsic repair response, activating mesenchymal stem cells residing in the bone marrow and leading to the formation of fibrocartilaginous repair tissue [69]. Fibrocartilage is structurally and biomechanically inferior to articular cartilage, still this endogenous repair program is the basis for so-called marrow-stimulating technique [70]. These include arthroscopic procedures where the subchondral bone is partially abrased, penetrated by drilling or microfractured. On the contrary, partial thickness defects that do not lead to stimulation of cells residing within the subchondral bone do not heal spontaneously. One reason for the persistence of partial cartilage loss is the inert nature of adult cartilage, which basically only consists of chondrocytes. In a healthy state it neither contains blood vessels, lymph vessels, nor does it contain nerve cells. Therefore a promising approach to treat partial, but also full thickness defects are cell-based transplantation approaches. The most widely applied method is autologous chondrocyte implantation (ACI), where chondrocytes are isolated from less weight-bearing areas of the joint, expanded in vitro and retransplanted into the defective area. Since this method relies on two separate surgical procedures it poses a burden for patients. Additionally, ACI leads to increased donor site morbidity, loss of chondrocytic phenotype in the course of in vitro expansion, and also fibrocartilage formation at the site of the defect [6, 7]. To reduce the risk as well as the discomfort associated with two surgical procedures potential future treatment options will rely on the application of stem cells for the treatment of degenerative joint disorders. Embryonic stem cells as well as induced pluripotent stem cells could be used, but on the one hand they potentially give rise to tumors [71–73] and on the other hand for high yield chondrogenic differentiation embryonic stem cells will first need to be differentiated towards mesenchymal precursor cells and then further on to chondrocytes, which makes the whole differentiation process more complex [74]. Mesenchymal stem cells are nowadays directly available from patient-derived tissues like bone marrow, muscle, and adipose tissue [8, 75, 76]. These cells are an attractive cell source for cartilage tissue engineering since they show strong proliferation and are easy to differentiate into chondrocytes, but their proliferation capacity can be exhausted and, when isolated from older patients, a mutation free status cannot be guaranteed. For the isolation of fetal cells the amniotic fluid is the best accessible compartment and it has been shown that cells within the amniotic fluid can be used to generate 3D fetal constructs able to be used for fetal surgery [77, 78]. With the discovery of AFS cells novel approaches in regenerative medicine have been enabled [79]. AFS cells are termed broadly pluripotent and are hallmarked by the expression of the stem cell-associated transcription factor OCT-4 [80–82]. AFS cells can be grown in large quantities and show a higher differentiation potential compared to adult stem cells [83]. AFS cells are devoid of a risk for tumor development and do not raise the ethical issues of embryonic stem cells. Furthermore AFS cells do not need exogenic treatment to induce pluripotency, are chromosomally stable and do not harbor the epigenetic memory and accumulated somatic mutations frequently found in adult source cells. Accordingly, AFS cells are now a much appreciated cell source for regenerative therapy approaches [84–87].
6 Use of Undifferentiated Amniotic Fluid Stem Cells for the Treatment of Degenerated Articular Cartilage
It is not always necessary to differentiate stem cells before they are used for cell-based regenerative approaches. For AFS cells for example it has been shown that they can contribute to renal tissue when co-cultured with embryonic kidney cells [85], or they can contribute to heart muscle after induction of heart failure [88]. The downside of this approach is that the number of differentiated cells is expected to be drastically lowered compared to the use of already pre-differentiated cells (see next paragraph), but transplantation of stem cells is known to elicit a plethora of beneficial effects not directly related to novel tissue formation. These effects include specific homing to the site of injury, attraction of cells necessary to stimulate the healing process and modulation of the host’s immune response. Interestingly, these effects can be of a transient nature since most transplanted stem cells do not integrate into the host tissue and will therefore be abandoned. For example, recently it was shown that amniotic fluid-derived cells with mesenchymal stem cell characteristics have the potential to accelerate the wound-healing process by stimulating proliferation and migration of dermal fibroblasts [89]. This phenomenon was attributed to the secretion of growth factors and cytokines including IL-8, IL-6, TGF-beta, TNFRI, VEGF, and EGF by the stem cells, which are important in normal wound healing. Up to date there a numerous clinical trials, which study the beneficial effects of mesenchymal stem cells directly transplanted to the region of articular cartilage degeneration, but to our knowledge AFS cells have not been used for this purpose so far.
7 Differentiation of Stem Cells from the Amniotic Fluid as a Novel Cell Source for Regenerative Therapy
Research on fetal tissue engineering lead to the discovery that amniotic fluid-derived cells harbor chondrogenic potential [90]. Ovine amniotic fluid cells were isolated and grown in culture medium without serum, but containing dexamethasone, ascorbic acid, sodium pyruvate, insulin, transferrin, and selenium. Additionally cells were treated with TGF-β2 and kept in culture for a total of 12 weeks. Compared to fetal hyaline cartilage the engineered constructs displayed less amounts of type II collagen, but similar amounts of glycosaminoglycans. Basically the culture conditions used in this first study reassembled the differentiation protocol described for mesenchymal stem cells derived from adult bone marrow aspirates [75]. First reports on human AFS cells used c-kit magnetically sorted cell populations and demonstrated increased production of type II collagen and sGAG after treatment with TGF-β1 and IGF-I [91]. The use of c-kit selected cells started a new field of research based upon the finding that c-kit selected single cell clones are broadly pluripotent [84]. Comparing c-kit positive and negative populations for their chondrogenic potential revealed that the c-kit positive stem cell population, after differentiation, displayed significant higher alcian blue staining and therefore stronger chondrogenic differentiation [92]. Therefore for chondrogenic differentiation either c-kit selected amniotic fluid cells or stem cell marker positive cells as depicted in Table 8.2 are used. The culture medium used is basically identical and Table 8.2 also shows that chondrogenesis can be triggered by different TGF-beta family members and by IGF-1, which has been shown in humans and also in other species. What can be concluded from chondrogenic differentiation of AFS cell up to now is that supplementation of chondrogenesis promoting factors is necessary for successful reprogramming of AFS cells and that the isolation of c-kit positive cells also enhances the differentiation process.
8 Spatial and Temporal Regulations of Chondrocytic Differentiation of Amniotic Fluid Stem Cells
The aim for the future is to improve in vitro protocols for the generation of artificial cartilage that can be transplanted to previously degenerated joints and will furthermore permit a permanent lubrication and full function of the joint (see Fig. 8.1). As already discussed human AFS cells represent a valuable cell source for ex vivo tissue regeneration, and ongoing work now focuses on the correct choice of differentiation factors, scaffolds and mechanical force. Optimal differentiation conditions are achieved when developmental processes regulating articular joint formation are recapitulated during in vitro culturing. This requires control of spatial and temporal changes, which on the one hand lead to efficient induction of chondrogenesis and on the other hand prevents the so-called hypertrophic state of chondrocytes. Adult chondrocytes reside in a hypoxic environment with very little nutrient supply. They are hallmarked by the expression of the chondrogenic transcription factors Sox5, Sox6, Sox9, and by formation of extracellular matrix components like Collagen type II, Collagen type IX, Collagen type XI and proteoglycans. As also shown in Table 8.2 generally TGF-β family members are sufficient for chondrogenesis in amnion fluid-derived cells, but recent data has also shown that in vitro expanded autologous chondrocytes, when treated with TGF-β1, show onset of hypertrophy [93]. Chondrocyte hypertrophy is evidenced, among others, by morphologic changes, expression of transcription factor Runx2 and production of Collagen type X and endothelial growth factor VEGFA. Hypertrophy generally leads to calcification of the tissue and subsequently to the loss of functional chondrocytes, an undesirable process within the joint region. Here we propose that developmental programs able to suppress onset of hypertrophy can also be used during cartilage engineering. For example Noggin, an antagonist of BMP signaling, has been identified to play an essential role in joint formation, since its deletion results in lack of GDF5 expression, a prominent member of the TGF-β family, and affected mice do not form joints [94]. Also WNT-4, WNT-14, WNT-16, and the WNT signaling mediator β-catenin show anti-chondrogenic activity during joint formation [35, 95]. The concept of exchanging factors during differentiation has already been used in a recent study were mesenchymal stem cells have been treated with TGF-β3 combined with β-catenin activation, which resulted in the prevention of hypertrophy and gave rise to a more stable chondrogenic phenotype than the induction with TGF-β3 alone [96]. Another way of achieving full differentiation without signs of hypertrophy is by forcing continuous expression of the master regulator SOX9. Studies in growth plate chondrocytes have shown that permanent induction of SOX9 inhibits terminal differentiation and hypertrophy by repression of the transcription factor RUNX2 [97, 98]. RUNX2 and RUNX3 are the pivotal inducers of type X and type I collagen as well as osteocalcin and MMP13 [99–101]. Therefore, it is tempting to speculate that treatment with other anti-chondrogenic stimuli during chondrogenic differentiation will result in the generation of fully functional and non-hypertrophic chondrocytes.

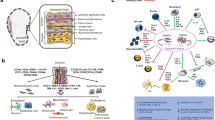
AFS cells as a cell source for articular cartilage regeneration. Cartilage is solely composed of chondrocytes and the extracellular matrix they produce. Articular cartilage can be divided into different zones, starting from the top with the superficial zone, where collagen fibers are arranged horizontally to the surface. Underneath resides the transitional zone with spherical chondrocytes. The thickest part of cartilage consists of the radial zone with collagen fibers and chondrocytes orientated vertically to the subchondral bone. The figure shows a partial cartilage defect which does not elicit a bone marrow response. This type of defect is best healed by transplantation with artificial generated tissue. As shown AFS cells can be used to generate artificial tissue finally consisting of cells and extracellular matrix similar to native cartilage. This can be achieved by the use of growth factors, scaffolds and environmental conditions reflecting developmental processes during cartilage formation
As shown in Fig. 8.1, partial cartilage defects need to be filled with regenerated tissue consisting of appropriate extra cellular matrix. AFS cells have mostly been induced for chondrogenic differentiation in so-called pellet culture systems which is a scaffoldless approach were approximately 250,000 cells are used to self assemble into a 3D structure. These culture conditions are reminiscent of in vivo joint formation where mesenchymal cell condensation precedes the induction of chondrogenesis and extracellular matrix molecules like type I collagen and type II collagen are produced. Since pellets are difficult to attach at sites of injury within cartilage, for transplantation approaches cells are frequently grown and differentiated on various scaffolds, which are supposed to exhibit biomechanical stability and aid in the construction of artificial tissue. Commonly used biomaterials are either biodegradable like agarose, alginate, hyaluronic acid, and collagen, or of synthetic origin like polyethylene glycol or poly-lactic-co-glycolic acid. Human amniotic fluid cells have so far been differentiated in three-dimensional pellets [92, 102] and in hyaluronan-based hydrogels [103]. Ovine amniotic fluid cells have been differentiated in biodegradable polyglycolic acid scaffolds [90] and in synthetic polyglycolic acid [104].
Articular cartilage is devoid of blood vessels, hence oxygen levels from the superficial zone to the calcified zone gradually decrease. Low oxygen tension termed hypoxia enhances the differentiation process of human mesenchymal stem cells and of human articular cartilage cells [105, 106], whereas in AFS cells, as far as we know, the impact of hypoxia on the chondrogenic potential has not been evaluated. Interestingly, chondrocyte hypertrophy, which is also a hallmark of osteoarthritis, is marked by extensive VEGF-A production. Secretion of this growth factor leads to increased angiogenesis in the affected organ area, leading to a resupply with oxygen and a subsequent mineralization process. This generation of osteophytes during progression of OA has been shown to occur in the knee joint of rabbits induced to form OA by anterior cruciate ligament transection [107]. On the contrary, hypoxia has the potential to inhibit upregulation of RUNX2 and thereby leads to a block of osteoblastic differentiation as demonstrated in mesenchymal stem cells [108].
Another physiologic process, which can be observed in native cartilage, is autophagy, since it protects chondrocytes from age-related cell death and preserves homeostasis within the tissue [109]. This process can protect cells under starvation conditions by turnover of intracellular organelles and molecules. Especially during aging-related cellular changes as well as biochemical stimuli and mechanical stress, all this can lead to osteoarthritis and during this process the competence to undergo autophagy is lost [110]. Therefore autophagy protects from aging-related cell death and probably it is also an important process during chondrogenic differentiation, even though its regulation during the differentiation process of amniotic fluid-derived stem cells has not been thoroughly studied.
9 Future Perspectives
Stem cells derived from the amniotic fluid represent a novel and promising source for cell-based therapy approaches in regenerative medicine. Cells can either be applied directly to the site of injury in order to stimulate repair processes or cells are used for chondrogenic differentiation in order to form artificial cartilage constructs. Differentiation of AFS cells has already been shown many times, most often via induction with TGF-β family members either embedded within matrix or in a scaffoldless approach. Still the molecular differentiation process is incompletely understood and up to date the regeneration of cartilage defects is elusive because novel generated tissue does not show the required toughness and hardness required for long term stability. Therefore differentiation protocols need to be improved and this is best achieved via recapitulation of the spacio-temporal aspects during chondrocyte differentiation. The hope is that future tissue regeneration will be based on stem cells stimulated to undergo self organized cartilage formation with almost native extracellular matrix composition and without signs of chondrocyte hypertrophy which will finally lead to durable repair tissue. Ways to achieve this includes the treatment with growth factors only for specific time periods and in combinations of pro and anti-chondrogenic treatment regiments in a defined ratio. This will yield the beneficial effects of chondrogenic stimuli, while preventing the end stage maturation effects of hypertrophy. Scaffolds are important three-dimensional environments, which regulate cell–cell interaction as well as oxygen supply, but usually scaffolds show too little mechanic stability and a negative impact on cell survival upon degradation. Recent efforts promote a scaffoldless approach where differentiated cells provide their own extra cellular matrix, like it is achieved when AFS cells are differentiated in a pellet culture approach. In conclusion, the demand for novel strategies to heal articular cartilage defects is high in our society and AFS cells have proven to harbor therapeutic potential. Especially the modification of existing chondrogenic differentiation approaches holds the promise to yield durable articular cartilage.
Abbreviations
- ACI:
-
Autologous chondrocyte implantation
- ADAM:
-
A disintegrin and metalloproteinases
- AFS cells:
-
Amniotic fluid stem cells
- BMP:
-
Bone morphogenic protein
- DZ:
-
Deep zone
- HA:
-
Hyaluronic acid
- HABP:
-
Hyaluronic acid-binding protein
- MMP:
-
Matrix-metalloprotease
- MZ:
-
Middle zone
- OA:
-
Osteoarthritis
- PCM:
-
Pericellular matrix
- PGA:
-
Polyglycolic acid
- sGAG:
-
Glycosaminoglycan
- SYM1:
-
Symphalangism
- SYNS1:
-
Synostosis syndrome
- SZ:
-
Superficial zone
References
Nehrer S, Minas T. Treatment of articular cartilage defects. Invest Radiol. 2000;35:639–46.
Jiang L, Tian W, Wang Y, Rong J, Bao C, Liu Y, Zhao Y, Wang C. Body mass index and susceptibility to knee osteoarthritis: a systematic review and meta-analysis. Joint Bone Spine. 2012; 79(3):291–7.
Jiang YZ, Zhang SF, Qi YY, Wang LL, Ouyang HW. Cell transplantation for articular cartilage defects: principles of past, present, and future practice. Cell Transplant. 2011;20:593–607.
Bedi A, Feeley BT, Williams 3rd RJ. Management of articular cartilage defects of the knee. J Bone Joint Surg Am. 2010;92: 994–1009.
Nehrer S, Dorotka R, Domayer S, Stelzeneder D, Kotz R. Treatment of full-thickness chondral defects with hyalograft C in the knee: a prospective clinical case series with 2 to 7 years’ follow-up. Am J Sports Med. 2009;37 Suppl 1:81S–7.
Salzmann GM, Sauerschnig M, Berninger MT, Kaltenhauser T, Schonfelder M, Vogt S, Wexel G, Tischer T, Sudkamp N, Niemeyer P, Imhoff AB, Schottle PB. The dependence of autologous chondrocyte transplantation on varying cellular passage, yield and culture duration. Biomaterials. 2011;32:5810–8.
Schnabel M, Marlovits S, Eckhoff G, Fichtel I, Gotzen L, Vecsei V, Schlegel J. Dedifferentiation-associated changes in morphology and gene expression in primary human articular chondrocytes in cell culture. Osteoarthritis Cartilage. 2002;10:62–70.
Sakaguchi Y, Sekiya I, Yagishita K, Muneta T. Comparison of human stem cells derived from various mesenchymal tissues: superiority of synovium as a cell source. Arthritis Rheum. 2005;52:2521–9.
Vinardell T, Buckley CT, Thorpe SD, Kelly DJ. Composition-function relations of cartilaginous tissues engineered from chondrocytes and mesenchymal stem cells isolated from bone marrow and infrapatellar fat pad. J Tissue Eng Regen Med. 2011;5:673–83.
Erickson IE, Huang AH, Chung C, Li RT, Burdick JA, Mauck RL. Differential maturation and structure-function relationships in mesenchymal stem cell- and chondrocyte-seeded hydrogels. Tissue Eng Part A. 2009;15:1041–52.
Rosner M, Schipany K, Gundacker C, Shanmugasundaram B, Li K, Fuchs C, Lube G, Hengstschlager M. Renal differentiation of amniotic fluid stem cells: perspectives for clinical application and for studies on specific human genetic diseases. Eur J Clin Invest. 2012;42(6):677–84.
Rosner M, Mikula M, Preitschopf A, Feichtinger M, Schipany K, Hengstschlager M. Neurogenic differentiation of amniotic fluid stem cells. Amino Acids. 2011;42(5):1591–6.
Archer CW, Dowthwaite GP, Francis-West P. Development of synovial joints. Birth Defects Res C Embryo Today. 2003;69:144–55.
Holder N. An experimental investigation into the early development of the chick elbow joint. J Embryol Exp Morphol. 1977;39:115–27.
Koyama E, Shibukawa Y, Nagayama M, Sugito H, Young B, Yuasa T, Okabe T, Ochiai T, Kamiya N, Rountree RB, Kingsley DM, Iwamoto M, Enomoto-Iwamoto M, Pacifici M. A distinct cohort of progenitor cells participates in synovial joint and articular cartilage formation during mouse limb skeletogenesis. Dev Biol. 2008;316:62–73.
Francis-West PH, Parish J, Lee K, Archer CW. BMP/GDF-signalling interactions during synovial joint development. Cell Tissue Res. 1999;296:111–9.
Mitrovic DR. Development of the metatarsophalangeal joint of the chick embryo: morphological, ultrastructural and histochemical studies. Am J Anat. 1977;150:333–47.
Edwards JC, Wilkinson LS, Jones HM, Soothill P, Henderson KJ, Worrall JG, Pitsillides AA. The formation of human synovial joint cavities: a possible role for hyaluronan and CD44 in altered interzone cohesion. J Anat. 1994;185(Pt 2):355–67.
Li T, Longobardi L, Myers TJ, Temple JD, Chandler RL, Ozkan H, Contaldo C, Spagnoli A. Joint TGF-beta type II receptor-expressing cells: ontogeny and characterization as joint progenitors. Stem Cells Dev. 2013;22(9):1342–59.
Pitsillides AA, Ashhurst DE. A critical evaluation of specific aspects of joint development. Dev Dyn. 2008;237:2284–94.
Pacifici M, Koyama E, Shibukawa Y, Wu C, Tamamura Y, Enomoto-Iwamoto M, Iwamoto M. Cellular and molecular mechanisms of synovial joint and articular cartilage formation. Ann N Y Acad Sci. 2006;1068:74–86.
Pazin DE, Gamer LW, Cox KA, Rosen V. Molecular profiling of synovial joints: use of microarray analysis to identify factors that direct the development of the knee and elbow. Dev Dyn. 2012;241:1816–26.
Storm EE, Kingsley DM. GDF5 coordinates bone and joint formation during digit development. Dev Biol. 1999;209:11–27.
Storm EE, Huynh TV, Copeland NG, Jenkins NA, Kingsley DM, Lee SJ. Limb alterations in brachypodism mice due to mutations in a new member of the TGF beta-superfamily. Nature. 1994;368:639–43.
Masuya H, Nishida K, Furuichi T, Toki H, Nishimura G, Kawabata H, Yokoyama H, Yoshida A, Tominaga S, Nagano J, Shimizu A, Wakana S, Gondo Y, Noda T, Shiroishi T, Ikegawa S. A novel dominant-negative mutation in Gdf5 generated by ENU mutagenesis impairs joint formation and causes osteoarthritis in mice. Hum Mol Genet. 2007;16:2366–75.
Kan A, Ikeda T, Fukai A, Nakagawa T, Nakamura K, Chung UI, Kawaguchi H, Tabin CJ. SOX11 contributes to the regulation of GDF5 in joint maintenance. BMC Dev Biol. 2013;13:4.
Massague J. TGFbeta signaling: receptors, transducers, and Mad proteins. Cell. 1996;85:947–50.
Baur ST, Mai JJ, Dymecki SM. Combinatorial signaling through BMP receptor IB and GDF5: shaping of the distal mouse limb and the genetics of distal limb diversity. Development. 2000;127: 605–19.
Kotzsch A, Nickel J, Seher A, Sebald W, Muller TD. Crystal structure analysis reveals a spring-loaded latch as molecular mechanism for GDF-5-type I receptor specificity. EMBO J. 2009;28:937–47.
Rountree RB, Schoor M, Chen H, Marks ME, Harley V, Mishina Y, Kingsley DM. BMP receptor signaling is required for postnatal maintenance of articular cartilage. PLoS Biol. 2004;2:e355.
Settle Jr SH, Rountree RB, Sinha A, Thacker A, Higgins K, Kingsley DM. Multiple joint and skeletal patterning defects caused by single and double mutations in the mouse Gdf6 and Gdf5 genes. Dev Biol. 2003;254:116–30.
Seemann P, Brehm A, Konig J, Reissner C, Stricker S, Kuss P, Haupt J, Renninger S, Nickel J, Sebald W, Groppe JC, Ploger F, Pohl J, Schmidt-von Kegler M, Walther M, Gassner I, Rusu C, Janecke AR, Dathe K, Mundlos S. Mutations in GDF5 reveal a key residue mediating BMP inhibition by NOGGIN. PLoS Genet. 2009;5:e1000747.
Gong Y, Krakow D, Marcelino J, Wilkin D, Chitayat D, Babul-Hirji R, Hudgins L, Cremers CW, Cremers FP, Brunner HG, Reinker K, Rimoin DL, Cohn DH, Goodman FR, Reardon W, Patton M, Francomano CA, Warman ML. Heterozygous mutations in the gene encoding noggin affect human joint morphogenesis. Nat Genet. 1999;21:302–4.
Khan IM, Redman SN, Williams R, Dowthwaite GP, Oldfield SF, Archer CW. The development of synovial joints. Curr Top Dev Biol. 2007;79:1–36.
Guo X, Day TF, Jiang X, Garrett-Beal L, Topol L, Yang Y. Wnt/beta-catenin signaling is sufficient and necessary for synovial joint formation. Genes Dev. 2004;18:2404–17.
Hartmann C, Tabin CJ. Wnt-14 plays a pivotal role in inducing synovial joint formation in the developing appendicular skeleton. Cell. 2001;104:341–51.
Spater D, Hill TP, O'Sullivan JR, Gruber M, Conner DA, Hartmann C. Wnt9a signaling is required for joint integrity and regulation of Ihh during chondrogenesis. Development. 2006;133:3039–49.
Osborne AC, Lamb KJ, Lewthwaite JC, Dowthwaite GP, Pitsillides AA. Short-term rigid and flaccid paralyses diminish growth of embryonic chick limbs and abrogate joint cavity formation but differentially preserve pre-cavitated joints. J Musculoskelet Neuronal Interact. 2002;2:448–56.
Craig FM, Bayliss MT, Bentley G, Archer CW. A role for hyaluronan in joint development. J Anat. 1990;171:17–23.
Pitsillides AA, Archer CW, Prehm P, Bayliss MT, Edwards JC. Alterations in hyaluronan synthesis during developing joint cavitation. J Histochem Cytochem. 1995;43:263–73.
Dowthwaite GP, Edwards JC, Pitsillides AA. An essential role for the interaction between hyaluronan and hyaluronan binding proteins during joint development. J Histochem Cytochem. 1998;46: 641–51.
Pan J, Zhou X, Li W, Novotny JE, Doty SB, Wang L. In situ measurement of transport between subchondral bone and articular cartilage. J Orthop Res. 2009;27:1347–52.
Wang Y, Wei L, Zeng L, He D, Wei X. Nutrition and degeneration of articular cartilage. Knee Surg Sports Traumatol Arthrosc. 2012;21(8):1751–62.
Grad S, Lee CR, Wimmer MA, Alini M. Chondrocyte gene expression under applied surface motion. Biorheology. 2006;43: 259–69.
Tuan RS, Chen AF, Klatt BA. Cartilage regeneration. J Am Acad Orthop Surg. 2013;21:303–11.
Spitters TW, Leijten JC, Deus FD, Costa IB, van Apeldoorn AA, van Blitterswijk CA, Karperien M. A dual flow bioreactor with controlled mechanical stimulation for cartilage tissue engineering. Tissue Eng Part C Methods. 2013;19(10):774–83.
Flannery CR, Hughes CE, Schumacher BL, Tudor D, Aydelotte MB, Kuettner KE, Caterson B. Articular cartilage superficial zone protein (SZP) is homologous to megakaryocyte stimulating factor precursor and Is a multifunctional proteoglycan with potential growth-promoting, cytoprotective, and lubricating properties in cartilage metabolism. Biochem Biophys Res Commun. 1999;254:535–41.
Schumacher BL, Block JA, Schmid TM, Aydelotte MB, Kuettner KE. A novel proteoglycan synthesized and secreted by chondrocytes of the superficial zone of articular cartilage. Arch Biochem Biophys. 1994;311:144–52.
Alsalameh S, Amin R, Gemba T, Lotz M. Identification of mesenchymal progenitor cells in normal and osteoarthritic human articular cartilage. Arthritis Rheum. 2004;50:1522–32.
Hattori S, Oxford C, Reddi AH. Identification of superficial zone articular chondrocyte stem/progenitor cells. Biochem Biophys Res Commun. 2007;358:99–103.
Dowthwaite GP, Bishop JC, Redman SN, Khan IM, Rooney P, Evans DJ, Haughton L, Bayram Z, Boyer S, Thomson B, Wolfe MS, Archer CW. The surface of articular cartilage contains a progenitor cell population. J Cell Sci. 2004;117:889–97.
Grogan SP, Miyaki S, Asahara H, D'Lima DD, Lotz MK. Mesenchymal progenitor cell markers in human articular cartilage: normal distribution and changes in osteoarthritis. Arthritis Res Ther. 2009;11:R85.
Kim AC, Spector M. Distribution of chondrocytes containing alpha-smooth muscle actin in human articular cartilage. J Orthop Res. 2000;18:749–55.
Hung SC, Kuo PY, Chang CF, Chen TH, Ho LL. Alpha-smooth muscle actin expression and structure integrity in chondrogenesis of human mesenchymal stem cells. Cell Tissue Res. 2006;324:457–66.
Guilak F, Alexopoulos LG, Upton ML, Youn I, Choi JB, Cao L, Setton LA, Haider MA. The pericellular matrix as a transducer of biomechanical and biochemical signals in articular cartilage. Ann N Y Acad Sci. 2006;1068:498–512.
Poole CA. Articular cartilage chondrons: form, function and failure. J Anat. 1997;191(Pt 1):1–13.
Zhang Z, Jin W, Beckett J, Otto T, Moed B. A proteomic approach for identification and localization of the pericellular components of chondrocytes. Histochem Cell Biol. 2011;136:153–62.
Grimshaw MJ, Mason RM. Modulation of bovine articular chondrocyte gene expression in vitro by oxygen tension. Osteoarthritis Cartilage. 2001;9:357–64.
Buckwalter JA. Articular cartilage injuries. Clin Orthop Relat Res. 2002;(402):21-37.
Goldring MB, Goldring SR. Articular cartilage and subchondral bone in the pathogenesis of osteoarthritis. Ann N Y Acad Sci. 2010;1192:230–7.
Goldring MB, Goldring SR. Osteoarthritis. J Cell Physiol. 2007;213:626–34.
Murphy G, Nagase H. Reappraising metalloproteinases in rheumatoid arthritis and osteoarthritis: destruction or repair? Nat Clin Pract Rheumatol. 2008;4:128–35.
Goldring SR. Role of bone in osteoarthritis pathogenesis. Med Clin North Am. 2009;93:25–35, xv.
Schroeppel JP, Crist JD, Anderson HC, Wang J. Molecular regulation of articular chondrocyte function and its significance in osteoarthritis. Histol Histopathol. 2011;26:377–94.
Gadjanski I, Spiller K, Vunjak-Novakovic G. Time-dependent processes in stem cell-based tissue engineering of articular cartilage. Stem Cell Rev. 2012;8(3):863–81.
Zhao Q, Eberspaecher H, Lefebvre V, De Crombrugghe B. Parallel expression of Sox9 and Col2a1 in cells undergoing chondrogenesis. Dev Dyn. 1997;209:377–86.
Ng LJ, Wheatley S, Muscat GE, Conway-Campbell J, Bowles J, Wright E, Bell DM, Tam PP, Cheah KS, Koopman P. SOX9 binds DNA, activates transcription, and coexpresses with type II collagen during chondrogenesis in the mouse. Dev Biol. 1997;183: 108–21.
Leung VY, Gao B, Leung KK, Melhado IG, Wynn SL, Au TY, Dung NW, Lau JY, Mak AC, Chan D, Cheah KS. SOX9 governs differentiation stage-specific gene expression in growth plate chondrocytes via direct concomitant transactivation and repression. PLoS Genet. 2011;7:e1002356.
Redman SN, Oldfield SF, Archer CW. Current strategies for articular cartilage repair. Eur Cell Mater. 2005;9:23–32. discussion 23–32.
Madry H, Grun UW, Knutsen G. Cartilage repair and joint preservation: medical and surgical treatment options. Dtsch Arztebl Int. 2011;108:669–77.
Roberts S, Genever P, McCaskie A, De Bari C. Prospects of stem cell therapy in osteoarthritis. Regen Med. 2011;6:351–66.
Toh WS, Lee EH, Cao T. Potential of human embryonic stem cells in cartilage tissue engineering and regenerative medicine. Stem Cell Rev. 2011;7:544–59.
Kawaguchi J, Mee PJ, Smith AG. Osteogenic and chondrogenic differentiation of embryonic stem cells in response to specific growth factors. Bone. 2005;36:758–69.
Barberi T, Willis LM, Socci ND, Studer L. Derivation of multipotent mesenchymal precursors from human embryonic stem cells. PLoS Med. 2005;2:e161.
Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S, Marshak DR. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284:143–7.
Liu TM, Martina M, Hutmacher DW, Hui JH, Lee EH, Lim B. Identification of common pathways mediating differentiation of bone marrow- and adipose tissue-derived human mesenchymal stem cells into three mesenchymal lineages. Stem Cells. 2007;25: 750–60.
Fauza DO, Fishman SJ, Mehegan K, Atala A. Videofetoscopically assisted fetal tissue engineering: bladder augmentation. J Pediatr Surg. 1998;33:7–12.
Fauza DO, Marler JJ, Koka R, Forse RA, Mayer JE, Vacanti JP. Fetal tissue engineering: diaphragmatic replacement. J Pediatr Surg. 2001;36:146–51.
Prusa AR, Marton E, Rosner M, Bernaschek G, Hengstschlager M. Oct-4-expressing cells in human amniotic fluid: a new source for stem cell research? Hum Reprod. 2003;18:1489–93.
Kim J, Lee Y, Kim H, Hwang KJ, Kwon HC, Kim SK, Cho DJ, Kang SG, You J. Human amniotic fluid-derived stem cells have characteristics of multipotent stem cells. Cell Prolif. 2007;40:75–90.
Tsai MS, Lee JL, Chang YJ, Hwang SM. Isolation of human multipotent mesenchymal stem cells from second-trimester amniotic fluid using a novel two-stage culture protocol. Hum Reprod. 2004;19:1450–6.
Bossolasco P, Montemurro T, Cova L, Zangrossi S, Calzarossa C, Buiatiotis S, Soligo D, Bosari S, Silani V, Deliliers GL, Rebulla P, Lazzari L. Molecular and phenotypic characterization of human amniotic fluid cells and their differentiation potential. Cell Res. 2006;16:329–36.
Miranda-Sayago JM, Fernandez-Arcas N, Benito C, Reyes-Engel A, Carrera J, Alonso A. Lifespan of human amniotic fluid-derived multipotent mesenchymal stromal cells. Cytotherapy. 2011;13: 572–81.
De Coppi P, Bartsch Jr G, Siddiqui MM, Xu T, Santos CC, Perin L, Mostoslavsky G, Serre AC, Snyder EY, Yoo JJ, Furth ME, Soker S, Atala A. Isolation of amniotic stem cell lines with potential for therapy. Nat Biotechnol. 2007;25:100–6.
Siegel N, Rosner M, Unbekandt M, Fuchs C, Slabina N, Dolznig H, Davies JA, Lubec G, Hengstschlager M. Contribution of human amniotic fluid stem cells to renal tissue formation depends on mTOR. Hum Mol Genet. 2010;19:3320–31.
Valli A, Rosner M, Fuchs C, Siegel N, Bishop CE, Dolznig H, Madel U, Feichtinger W, Atala A, Hengstschlager M. Embryoid body formation of human amniotic fluid stem cells depends on mTOR. Oncogene. 2010;29:966–77.
Rosner M, Siegel N, Fuchs C, Slabina N, Dolznig H, Hengstschlager M. Efficient siRNA-mediated prolonged gene silencing in human amniotic fluid stem cells. Nat Protoc. 2010;5:1081–95.
Castellani C, Vescovo G, Ravara B, Franzin C, Pozzobon M, Tavano R, Gorza L, Papini E, Vettor R, De Coppi P, Thiene G, Angelini A. The contribution of stem cell therapy to skeletal muscle remodeling in heart failure. Int J Cardiol. 2013;168(3): 2014–21.
Yoon BS, Moon JH, Jun EK, Kim J, Maeng I, Kim JS, Lee JH, Baik CS, Kim A, Cho KS, Lee HH, Whang KY, You S. Secretory profiles and wound healing effects of human amniotic fluid-derived mesenchymal stem cells. Stem Cells Dev. 2010;19: 887–902.
Kunisaki SM, Jennings RW, Fauza DO. Fetal cartilage engineering from amniotic mesenchymal progenitor cells. Stem Cells Dev. 2006;15:245–53.
Kolambkar YM, Peister A, Soker S, Atala A, Guldberg RE. Chondrogenic differentiation of amniotic fluid-derived stem cells. J Mol Histol. 2007;38:405–13.
Arnhold S, Gluer S, Hartmann K, Raabe O, Addicks K, Wenisch S, Hoopmann M. Amniotic-fluid stem cells: growth dynamics and differentiation potential after a CD-117-based selection procedure. Stem Cells Int. 2011;2011:715341.
Narcisi R, Quarto R, Ulivi V, Muraglia A, Molfetta L, Giannoni P. TGF beta-1 administration during ex-vivo expansion of human articular chondrocytes in a serum-free medium redirects the cell phenotype toward hypertrophy. J Cell Physiol. 2012;227(9): 3282–90.
Brunet LJ, McMahon JA, McMahon AP, Harland RM. Noggin, cartilage morphogenesis, and joint formation in the mammalian skeleton. Science. 1998;280:1455–7.
Day TF, Guo X, Garrett-Beal L, Yang Y. Wnt/beta-catenin signaling in mesenchymal progenitors controls osteoblast and chondrocyte differentiation during vertebrate skeletogenesis. Dev Cell. 2005;8:739–50.
Yang Z, Zou Y, Guo XM, Tan HS, Denslin V, Yeow CH, Ren XF, Liu TM, Hui JH, Lee EH. Temporal activation of β-catenin signaling in the chondrogenic process of mesenchymal stem cells affects the phenotype of the cartilage generated. Stem Cells Dev. 2012;21(11):1966–76.
Saito T, Ikeda T, Nakamura K, Chung UI, Kawaguchi H. S100A1 and S100B, transcriptional targets of SOX trio, inhibit terminal differentiation of chondrocytes. EMBO Rep. 2007;8:504–9.
Yamashita S, Andoh M, Ueno-Kudoh H, Sato T, Miyaki S, Asahara H. Sox9 directly promotes Bapx1 gene expression to repress Runx2 in chondrocytes. Exp Cell Res. 2009;315:2231–40.
Higashikawa A, Saito T, Ikeda T, Kamekura S, Kawamura N, Kan A, Oshima Y, Ohba S, Ogata N, Takeshita K, Nakamura K, Chung UI, Kawaguchi H. Identification of the core element responsive to runt-related transcription factor 2 in the promoter of human type X collagen gene. Arthritis Rheum. 2009;60:166–78.
Soung do Y, Dong Y, Wang Y, Zuscik MJ, Schwarz EM, O’Keefe RJ, Drissi H. Runx3/AML2/Cbfa3 regulates early and late chondrocyte differentiation. J Bone Miner Res. 2007;22:1260–70.
Mengshol JA, Vincenti MP, Brinckerhoff CE. IL-1 induces collagenase-3 (MMP-13) promoter activity in stably transfected chondrocytic cells: requirement for Runx-2 and activation by p38 MAPK and JNK pathways. Nucleic Acids Res. 2001;29:4361–72.
Kolambkar YM, Peister A, Soker S, Atala A, Guldberg RE. Chondrogenic differentiation of amniotic fluid-derived stem cells. J Mol Histol. 2007;38:405–13.
Park JS, Shim MS, Shim SH, Yang HN, Jeon SY, Woo DG, Lee DR, Yoon TK, Park KH. Chondrogenic potential of stem cells derived from amniotic fluid, adipose tissue, or bone marrow encapsulated in fibrin gels containing TGF-beta3. Biomaterials. 2011;32:8139–49.
Kunisaki SM, Fuchs JR, Steigman SA, Fauza DO. A comparative analysis of cartilage engineered from different perinatal mesenchymal progenitor cells. Tissue Eng. 2007;13:2633–44.
Khan WS, Adesida AB, Tew SR, Lowe ET, Hardingham TE. Bone marrow-derived mesenchymal stem cells express the pericyte marker 3G5 in culture and show enhanced chondrogenesis in hypoxic conditions. J Orthop Res. 2010;28:834–40.
Kay A, Richardson J, Forsyth NR. Physiological normoxia and chondrogenic potential of chondrocytes. Front Biosci (Elite Ed). 2011;3:1365–74.
Hashimoto S, Creighton-Achermann L, Takahashi K, Amiel D, Coutts RD, Lotz M. Development and regulation of osteophyte formation during experimental osteoarthritis. Osteoarthritis Cartilage. 2002;10:180–7.
Yang DC, Yang MH, Tsai CC, Huang TF, Chen YH, Hung SC. Hypoxia inhibits osteogenesis in human mesenchymal stem cells through direct regulation of RUNX2 by TWIST. PLoS One. 2011;6:e23965.
Lotz MK, Carames B. Autophagy and cartilage homeostasis mechanisms in joint health, aging and OA. Nat Rev Rheumatol. 2011;7:579–87.
Carames B, Taniguchi N, Otsuki S, Blanco FJ, Lotz M. Autophagy is a protective mechanism in normal cartilage, and its aging-related loss is linked with cell death and osteoarthritis. Arthritis Rheum. 2010;62:791–801.
Bell DM, Leung KK, Wheatley SC, Ng LJ, Zhou S, Ling KW, Sham MH, Koopman P, Tam PP, Cheah KS. SOX9 directly regulates the type-II collagen gene. Nat Genet. 1997;16:174–8.
Shepard JB, Krug HA, LaFoon BA, Hoffman S, Capehart AA. Versican expression during synovial joint morphogenesis. Int J Biol Sci. 2007;3:380–4.
Niswander L. Interplay between the molecular signals that control vertebrate limb development. Int J Dev Biol. 2002;46:877–81.
Minina E, Kreschel C, Naski MC, Ornitz DM, Vortkamp A. Interaction of FGF, Ihh/Pthlh, and BMP signaling integrates chondrocyte proliferation and hypertrophic differentiation. Dev Cell. 2002;3:439–49.
Ikeda T, Kamekura S, Mabuchi A, Kou I, Seki S, Takato T, Nakamura K, Kawaguchi H, Ikegawa S, Chung UI. The combination of SOX5, SOX6, and SOX9 (the SOX trio) provides signals sufficient for induction of permanent cartilage. Arthritis Rheum. 2004;50:3561–73.
Nagy A, Kenesi E, Rentsendorj O, Molnar A, Szenasi T, Sinko I, Zvara A, Oommen ST, Barta E, Puskas LG, Lefebvre V, Kiss I. Evolutionarily conserved, growth plate zone-specific regulation of the matrilin-1 promoter: L-Sox5/Sox6 and Nfi factors bound near TATA finely tune activation by Sox9. Mol Cell Biol. 2011;31:686–99.
Dowthwaite GP, Edwards JC, Pitsillides AA. An essential role for the interaction between hyaluronan and hyaluronan binding proteins during joint development. J Histochem Cytochem. 1998;46: 641–51.
Chen WH, Lai MT, Wu AT, Wu CC, Gelovani JG, Lin CT, Hung SC, Chiu WT, Deng WP. In vitro stage-specific chondrogenesis of mesenchymal stem cells committed to chondrocytes. Arthritis Rheum. 2009;60:450–9.
Kou I, Ikegawa S. SOX9-dependent and -independent transcriptional regulation of human cartilage link protein. J Biol Chem. 2004;279:50942–8.
Liu CJ, Zhang Y, Xu K, Parsons D, Alfonso D, Di Cesare PE. Transcriptional activation of cartilage oligomeric matrix protein by Sox9, Sox5, and Sox6 transcription factors and CBP/p300 coactivators. Front Biosci. 2007;12:3899–910.
Sekiya I, Tsuji K, Koopman P, Watanabe H, Yamada Y, Shinomiya K, Nifuji A, Noda M. SOX9 enhances aggrecan gene promoter/enhancer activity and is up-regulated by retinoic acid in a cartilage-derived cell line, TC6. J Biol Chem. 2000;275:10738–44.
Zhang P, Jimenez SA, Stokes DG. Regulation of human COL9A1 gene expression. Activation of the proximal promoter region by SOX9. J Biol Chem. 2003;278:117–23.
DeLise AM, Fischer L, Tuan RS. Cellular interactions and signaling in cartilage development. Osteoarthritis Cartilage. 2000;8:309–34.
Quintana L, zur Nieden NI, Semino CE. Morphogenetic and regulatory mechanisms during developmental chondrogenesis: new paradigms for cartilage tissue engineering. Tissue Eng Part B Rev. 2009;15:29–41.
Rodrigues MT, Lee SJ, Gomes ME, Reis RL, Atala A, Yoo JJ. Bilayered constructs aimed at osteochondral strategies: the influence of medium supplements in the osteogenic and chondrogenic differentiation of amniotic fluid-derived stem cells. Acta Biomater. 2012;8:2795–806.
Baghaban Eslaminejad M, Jahangir S, Aghdami N. Mesenchymal stem cells from murine amniotic fluid as a model for preclinical investigation. Arch Iran Med. 2011;14:96–103.
Roubelakis MG, Bitsika V, Zagoura D, Trohatou O, Pappa KI, Makridakis M, Antsaklis A, Vlahou A, Anagnou NP. In vitro and in vivo properties of distinct populations of amniotic fluid mesenchymal progenitor cells. J Cell Mol Med. 2011;15:1896–913.
Disclosure Statement
No competing financial interests exist.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer Science+Business Media New York
About this chapter
Cite this chapter
Preitschopf, A., Busch, J., Zwickl, H., Nehrer, S., Hengstschläger, M., Mikula, M. (2014). Amniotic Fluid Stem Cells for the Treatment of Articular Cartilage Defects. In: Atala, A., Murphy, S. (eds) Perinatal Stem Cells. Springer, New York, NY. https://doi.org/10.1007/978-1-4939-1118-9_8
Download citation
DOI: https://doi.org/10.1007/978-1-4939-1118-9_8
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4939-1117-2
Online ISBN: 978-1-4939-1118-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)