
Abstract
In this chapter, we examine, from an ultimate perspective, how and why various bodily traits—leg length, foot size, breast size, body shape (waist-to-hip ratio), and body size (body mass index)—affect judgments of female attractiveness. Bodily trait values associated with optimal health and reproductive profiles are generally considered attractive across cultures and time periods, yet some traits may showcase certain classes of information better than others. Namely, bodily traits appear to better convey a woman’s immediate fertility, while facial traits may better convey her reproductive value. Consistent with this premise, we review research demonstrating a perceptual tendency for men to preferentially attend to women’s bodies in short-term mating contexts but not long-term mating contexts. Finally, we suggest future directions that promise to spur research on the asymmetrical abilities of facial and bodily traits to reveal key information pertaining to women’s reproductive condition.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
Intellectual debate regarding the universality of standards of beauty has waned in recent years. An unprecedented number of researchers now embrace the evolutionary perspective that components of physical attractiveness reflect individuals’ health and reproductive condition, rather than arbitrary cultural norms (e.g., Singh & Singh, 2011; Sugiyama, 2005) (cf. Wolf, 1991). An increasing amount of attention has instead been placed on the relative strength with which morphological traits predict perceptions of overall attractiveness. For instance, variations in body mass index (BMI) are frequently compared to those in waist-to-hip ratio (WHR) to determine whether BMI or WHR is the more salient index of a woman’s bodily attractiveness (Singh, 1993; Tovée, Maisey, Emery, & Cornelissen, 1999). Studies consistently find that both measures affect perceptions of attractiveness (e.g., Furnham, Petrides, & Constantinides, 2005; Furnham, Swami, & Shah, 2006); however, the specific values associated with maximum levels of attractiveness appear to depend on the availability of local resources. For example, in subsistence-based societies, greater priority is given to body weight (BMI) than body shape (WHR), as the former indicates the availability of sufficient resources to support the metabolic costs of reproduction (Marlowe & Wetsman, 2001; Sugiyama, 2004; Wetsman & Marlowe, 1999). Nearly every morphological trait imaginable—from head (e.g., hair color; Swami, Furnham, & Joshi, 2008) to toe (e.g., foot size; Fessler, Haley, & Lal, 2005)—has received some empirical attention. In order to make the process of drawing inferences from this data more tractable, a line is often drawn at the neck, separating facial and bodily components of attractiveness (e.g., Confer, Perilloux, & Buss, 2010). The primary focus of the current chapter is the argument that bodily components of attractiveness convey certain information about a woman’s reproductive profile that cannot be gleaned as easily from facial components of attractiveness.
We begin this chapter by integrating evidence that various bodily traits predict a woman’s health, hormonal profile, and reproductive status with empirical findings that demonstrate systematic preferences for optimal levels within those traits. We then consider the plasticity of attractiveness judgments across cultures and time periods. In this part, we present new evidence challenging the popular belief that Baroque ideals of attractiveness (e.g., high BMI) are vastly different from modern ideals. We conclude with evidence showing that men preferentially attend to women’s bodies in short-term mating contexts (e.g., one-night stand). These results are discussed in light of the hypothesis that fertility cues may be better gleaned from a woman’s body than her face.
Components of Bodily Attractiveness
Aspects of physical attractiveness are experienced as “attractive” because they have been reliably associated with individuals’ health, hormonal profile, and reproductive status throughout human evolutionary history (Symons, 1979; Williams, 1975). These traits are said to be honest, meaning the integrity of their signaling value is maintained by the inability of individuals with decreased fitness to imitate such cues (Zahavi, 1975). Bilateral symmetry, for example, reflects an individual’s ability to withstand environmental (e.g., parasitic) and genetic perturbations during development (Thornhill & Gangestad, 1994). Individuals with lower-quality immune systems are more susceptible to developmental insults and are therefore less likely to maintain symmetrical features. Consequently, individuals with symmetrical faces and bodies are preferentially sought as mates (Gangestad & Thornhill, 1997; Thornhill & Gangestad, 1994).
Although attended to by individuals of both sexes, men place relatively greater priority than women on a potential mate’s physical attractiveness (Buss, 1989; Li, Bailey, Kenrick, & Linsenmeier, 2002). This is because men’s reproductive success, more so than women’s, was primarily limited by access to healthy, fertile mates over human evolutionary history (Sugiyama, 2005; Symons, 1979). Consequently, men’s mating psychology is designed to attend to cues of a woman’s reproductive value (a measure of future reproductive potential that is strongly correlated with a woman’s age) and fertility (i.e., fecundability, a measure of a woman’s current ability to become pregnant) and find women who possess high levels of both especially attractive. Although these two dimensions are partially dissociable (e.g., a young pregnant woman is likely to have high reproductive value despite a current fertility of zero), many bodily traits simultaneously convey information pertaining to both. In this first part, we review various bodily features, detailing their health and reproductive correlates, as well as empirical evidence showing systematic preferences for specific variations in each trait.
Leg Length
One of the more recent empirical developments over the past decade has been the identification of leg length as a determinant of attractiveness. This trait is frequently operationalized as a leg-to-body ratio (LBR), representing the proportion of an individual’s height (usually including the head) that is accounted for by the legs (Swami, Einon, & Furnham, 2006; Swami, Gray, & Furnham, 2006). Before its application in evolutionary psychology, LBR was primarily used as a measure of childhood nutritional status, with lower LBRs representing periods of interrupted growth (Davey Smith et al., 2001). LBR has also been associated with various indices of health, including lower BMI, blood pressure, and cholesterol, as well as a reduced risk of coronary heart disease, diabetes, and cancer (Davey Smith et al., 2001; Gunnell, May, Ben-Shlomo, Yarnell, & Davey Smith, 2003; Gunnell, Whitley, et al., 2003). Importantly, childhood environmental conditions have been shown to influence leg length more strongly than any other component of stature (e.g., trunk length; Gunnell, May, et al., 2003), rendering LBR an especially powerful marker of resource availability and health during development.
In addition to functioning as an honest signal of health, Swami, Einon, et al. (2006) propose that women have a slightly higher LBR than men, and thus, LBR is used to differentiate masculine from feminine body types. To test this possibility, Swami et al. presented men and women with line drawings of male and female figures that varied in LBR. They hypothesized that if high LBRs are attractive because they indicate genetic quality, high LBRs should be considered more attractive than low LBRs regardless of sex. If, on the other hand, high LBRs signal femininity, high LBRs should be considered more attractive only within female figures. The results supported the latter relationship between LBR and femininity: as LBR increased, attractiveness ratings increased for female figures, but decreased for male figures. This pattern of results also replicated in a cross-cultural sample of British and Malaysian participants (Swami, Einon, & Furnham, 2007). Other studies eschew the explanation provided by Swami, Gray, et al. (2006), given research that shows no sexual dimorphism in LBR (for relevant citations, see Sorokowski & Pawlowski, 2008). These studies instead find a curvilinear, rather than linear, preference for LBR, with attractiveness assessments peaking at average (approximately 0.50) to slightly above-average (elongated by 5–10 %) LBRs for both male and female stimuli (Frederick, Hadji-Michael, Furnham, & Swami, 2010; Sorokowski, Sorokowska, & Mberira, 2012; Sorokowski & Pawlowski, 2008). Most notably, Sorokowski et al. (2011) surveyed the LBR preferences of men and women from 27 nations and found average LBRs to be maximally attractive across figures of both sexes. Clearly more research is needed to resolve the discrepancy between these findings and those of Swami, Einon, et al. (2006, 2007); however, hypotheses that LBR signals femininity (Swami, Einon, et al., 2006) or health (Frederick et al., 2010; Sorokowski et al., 2012; Sorokowski & Pawlowski, 2008) need not be mutually exclusive. Higher than average LBRs may be selected for up until the point at which long legs become biomechanically inefficient (e.g., in running or jumping; Sorokowski et al., 2012). Thus, leg length may serve as a cue of health and possibly, femininity.
Foot Size
Foot size is another bodily trait hypothesized to act as a sexually dimorphic signal. Women have smaller feet proportionate to their stature compared to men (Fessler et al., 2005; Voracek, Fisher, Rupp, Lucas, & Fessler, 2007), thereby posing a biomechanical challenge of maintaining body stability, particularly during pregnancy (Fessler et al., 2005). If women have been selected toward smaller foot size, counteracting evolutionary pressures (e.g., an increase in perceived attractiveness) must have outweighed costs associated with small feet. In women, foot size gradually increases with age and parity (for relevant citations, see Fessler et al., 2005); thus, small feet may be considered attractive because they indicate high reproductive value. Indeed, women with relatively large feet are judged to be older (Fessler et al., 2012) and less attractive (Fessler et al., 2005, 2012; Voracek et al., 2007) than those with relatively small feet. As would be expected in light of biomechanical efficiencies, women prefer men with average-to-large-sized feet (Fessler et al., 2012; Voracek et al., 2007), although, consistent with unidirectional sexual selection for foot size, women are less interested in men’s feet than men are in women’s feet (Fessler et al., 2012; Voracek et al., 2007). Foot size may therefore indicate a woman’s reproductive value and has been recognized (albeit not for the ultimate reasons indicated here) as an important component of female attractiveness as evidenced by practices such as historical Chinese foot binding (Fessler et al., 2005).
Breast Size
Human females are unique among primates because their breasts are perennially enlarged even when they are not pregnant or lactating (Marlowe, 1998). This, bolstered by the lack of evidence to suggest that breast size is directly related to reproductive capabilities (e.g., milk production; Anderson, 1983), suggests that prominent breasts are maintained by a process of sexual selection (Symons, 1979). The majority of research on the issue of preferred breast size shows medium- to large-sized breasts to be maximally attractive (Dixson, Grimshaw, Linklater, & Dixson, 2011; Zelazniewicz & Pawlowski, 2011; but see Furnham & Swami, 2007), especially in combination with other desirable bodily traits (e.g., low WHR; Furnham, Dias, & McClelland, 1998; Furnham et al., 2006; Singh & Young, 1995). Members of both sexes generally agree that women with large breasts are particularly attractive (e.g., Furnham et al., 1998; cf. Gitter, Lomranz, Saxe, & Bar-Tal, 1983); however, a preference for large breasts is exaggerated in men with a stronger proclivity toward casual sex (i.e., men with an unrestricted sociosexual orientation; Penke & Asendorpf, 2008; Zelazniewicz & Pawlowski, 2011). Guéguen (2007) showed that men were more likely than women to offer assistance to large-breasted women, despite the sexes’ general agreement in attractiveness ratings, indicating an instrumental motivation behind assistance patterns.
A number of hypotheses have been proposed to explain the function of perpetually enlarged breasts in women. It was originally thought that large breasts signaled the availability of fat reserves necessary for reproduction (Cant, 1981; Gallop, 1982). This conjecture is supported by research showing that ovarian function ceases in women with too little body fat (amenorrhea; Ellison, 1990; Frisch, 1987). Still this hypothesis only explains why a certain amount of body fat is attractive; it does not explain why fat reserves would be localized around the mammary glands. Other research suggests that large breasts signal fecundity. Jasieńska, Ziomkiewicz, Ellison, Lipson, and Thune (2004) found that women with large breasts have higher estrogen levels then women with small breasts. This finding is given practical significance in combination with research showing women to be more likely to conceive during cycles with higher estrogen concentrations (Lipson & Ellison, 1996; Venners et al., 2006). Thus, an attraction to large-breasted women might function to preferentially direct mating effort toward women who are especially fecund.
Some researchers have also proposed a “good genes” hypothesis, in which breasts are purported to function as honest signals of phenotypic quality. Manning, Scutt, Whitehouse, and Leinster (1997), for instance, showed that large breasts were less asymmetrical than expected allometrically, in spite of the associated increase in estrogen (Jasieńska et al., 2004), which Manning et al. argue suppresses the immune system. Other research has shown breast symmetry to positively predict number of offspring, indicating that women with symmetrical breasts may be preferentially sought after as mates because they offer direct (i.e., high fecundity) and indirect (i.e., highly fecund daughters) fitness benefits (Møller, Soler, & Thornhill, 1995). Combined, these pieces of evidence suggest that symmetrical breasts indicate high phenotypic quality because they signal the ability to withstand the associated immunosuppressing costs of estrogen (Manning et al., 1997) while larger breasts allow for easier detection of asymmetry, increasing their signaling value.
The nubility hypothesis, proposed by Marlowe (1998), provides a complementary explanation. In this explanation, large breasts provide two key pieces of information. First, large breasts are only present in women who have passed puberty and are thus of reproductive age; prepubescent girls have small, non-protruding breasts (Marlowe, 1998). Second, large breasts serve as a more honest indication of a woman’s age than small breasts because their greater weight stretches and slackens fibrous breast tissue over time, leading to more obvious age-related changes in firmness. Thus, large breasts—a signal of sexual maturity (Sugiyama, 2005)—begin to sag and signal declining reproductive value (Barber, 1995). For large breasts, relative to small breasts, the difference between firm breasts and sagging breasts is more pronounced. Hence, men’s preference for ample breasts (and women’s desire for breasts larger than their current size; Thompson & Tantleff, 1992) can be accounted for by the ability of large breasts to better signal fertility (i.e., sexual maturity) via their size and reproductive value via their firmness.
Body Shape and Size
A woman’s body is subjected to what Singh (1993) refers to as a “wide first-pass filter,” identifying women who exhibit cues of poor reproductive condition. WHR, for example, is affected by three factors directly relating to a woman’s ability to conceive: (1) her hormonal profile, (2) her pregnancy status, and possibly, (3) her ovulatory status. First, a woman’s WHR provides information regarding her hormonal profile, indicating whether she is within the reproductive window of her life-span and, if she is, the ease with which she can conceive. Supporting this point, the WHR of young boys and girls is remarkably similar until puberty, at which time a bimodal distribution emerges with minimal overlap (Marti et al., 1991). In women, the increase in estrogen that accompanies puberty inhibits fat deposition in the abdominal region and stimulates fat deposition in the gluteofemoral region (hips, buttocks, and thighs), generating WHR values that typically fall between 0.67 and 0.80. In men, the increase in testosterone causes the reverse pattern of fat deposition, generating significantly higher WHR values that range from 0.85 to 0.95. At menopause, decreased estrogen levels increase WHR, thereby reducing the disparity between men and women’s WHRs. Because sex differences in WHR are manifested most prominently during the reproductive window of a woman’s life-span, a low WHR probabilistically indicates that a woman is both post-pubescent and premenopausal (Singh, 1993, 2006; Singh & Singh, 2011).
Furthermore, women with low WHRs have more optimal hormone profiles (i.e., higher estrogen levels; Jasieńska et al., 2004), have fewer irregular menstrual cycles, ovulate more frequently, and have less difficulty conceiving than women with abnormally high WHRs (for relevant citations, see Singh & Singh, 2011). This is further evidenced in women with polycystic ovarian syndrome, a condition marked by impaired estrogen production, who have higher WHRs and experience greater difficulty conceiving and an increased risk of miscarriage (Jakubowicz, Iuorno, Jakubowicz, Roberts, & Nestler, 2002; Singh & Singh, 2011). Lower WHRs also advertise reproductive quality beyond conception: gluteofemoral fat appears to be a special store of neurodevelopmental resources such that women with lower WHRs (greater gluteofemoral fat storage) produce children who score higher on cognitive tests (Lassek & Gaulin, 2008).
Second, as a woman progresses through pregnancy, her WHR increases dramatically (vastly exceeding 1.0), a clear indication that she is currently incapable of conceiving. As copulations with a pregnant woman cannot increase a man’s reproductive success, it would be adaptive to find women with very high WHRs to be unattractive. Finally, preliminary evidence suggests that WHR decreases at ovulation (Kirchengast & Gartner, 2002). Thus, a woman’s figure might also reveal whether she is at peak cycle fertility; however, these results should be interpreted with caution, as Bleske-Rechek et al. (2011) failed to replicate this effect. Clearly, a bounty of information pertaining to a woman’s fertility and reproductive value can be estimated from just a brief glimpse of her WHR.
Research over the past two decades has supported these ultimate explanations for why WHR is associated with women’s attractiveness. Women with WHR values that fall at the low end of the typical female range (0.68–0.72) are considered more attractive than women with masculine WHRs (>0.80). In the study that pioneered the investigation of WHR and physical attractiveness, Singh (1993) provided participants with line drawings of women that varied only by weight and WHR. Within each body weight category—underweight (90 lbs.), normal weight (120 lbs.), and overweight (150 lbs.)—were four values of WHR: 0.70, 0.80, 0.90, and 1.0 (see Fig. 7.1). Results indicated that, within each weight class, attractiveness ratings increased as WHR decreased in a linear fashion. N7, a normal weight figure with a WHR of 0.70, was rated as most attractive and was associated with sexiness and good health more so than any other figure.

The stimuli used in Singh (1993) represent three body weights—underweight (I), normal weight (II), and overweight (III)—and four levels of WHR. From Singh, D. (1993), “Adaptive significance of female physical attractiveness: Role of waist-to-hip ratio,” Journal of Personality and Social Psychology, 65, 293–307. Copyright 1993 by the American Psychological Association. Reproduced with permission
Singh’s (1993) results have been systematically replicated using various methodologies, including line drawings (e.g., Furnham, McClelland, & Omer, 2003; Schmalt, 2006; Singh, 1994a, 1994b, 2004), actual photographs (Henss, 2000; Singh, 1994b; Wilson, Tripp, & Boland, 2005), online advertisements of female escorts (Saad, 2008), archival data from the sixteenth- to eighteenth-century British literature (Singh, Renn, & Singh, 2007), and ancient Indian, Egyptian, Greco-Roman, and African sculptures (Singh, 2002) (for a review, see Singh, 2006). A preference for low WHR has also been demonstrated by evaluating modifications made by plastic surgeons to Singh’s original line drawings with the goal of making them more attractive. As expected, the normal weight figure with a WHR of 0.70 (N7) was altered the least (Singh, 2006).
Several studies have shown cross-cultural agreement in the preference for low WHR with participants representing ethnicities from the Azore Islands, Cameroon, Guinea-Bissau, Greece, Indonesia, Kenya, Samoa, Uganda, the United Kingdom, the United States, and New Zealand (Furnham, Moutafi, & Baguma, 2002; Furnham et al., 2003; Singh, 2004; Singh, Dixson, Jessop, Morgan, & Dixson, 2010; Singh & Luis, 1995). Other studies, particularly those involving inhabitants of subsistence-based societies, show a less consistent preference for low WHR (Marlowe & Wetsman, 2001; Wetsman & Marlowe, 1999; Yu & Shepard, 1998) (cf. Sugiyama, 2004). For instance, Wetsman and Marlowe (1999) found that men from a foraging population (Hadza) in Tanzania were not differentially attracted to women with feminine (e.g., 0.70) or masculine (e.g., 0.90) WHRs. These researchers later showed a preference among Hadza men for women with WHRs at or above 0.80 (Marlowe & Wetsman, 2001); however, this result was subsequently shown to be a mere artifact of frontal-view stimuli (Marlowe, Apicella, & Reed, 2005). Unlike stimuli presented in profile, frontal-view stimuli do not account for the protrusion of the buttocks. Marlowe et al. (2005) found that Hadza men prefer a relatively low-profile WHR (more protruding buttocks) and relatively high frontal WHR (thicker waist), whereas the opposite is true for American men. Thus, cross-cultural disparity may be less extreme than originally thought for preferences of actual WHRs, where both the width of the waist and the protrusion of the buttocks are taken into account (Marlowe et al., 2005). This cross-cultural evidence indicates that men’s preferences for hourglass shapes were not invented by Western media, a point made even clearer by the finding that congenitally blind individuals—who cannot have been inundated with media images of models and celebrities—show a similar preference for women with low WHR when assessing female body shapes through touch (Karremans, Frankenhuis, & Arons, 2010).
Some researchers allege that the preference for low WHR is actually driven by a preference for women with low body weight (Tassinary & Hansen, 1998; Tovée & Cornelissen, 1999). By narrowing the waist, critics argue, abdominal fat is eliminated, decreasing the perceived overall weight of the target stimuli (Tovée & Cornelissen, 1999). The goal of recent research has been to adjudicate between adaptationist (Singh, 1993) and by-product (Tassinary & Hansen, 1998) explanations of WHR preferences by controlling for the effects of BMI. Some researchers have done so statistically (Streeter & McBurney, 2003), others by increasing the thickness of arms and legs in line drawings to compensate for thinner waists (Furnham et al., 2005). In all cases, WHR remains a significant predictor of attractiveness, with assessments peaking at approximately 0.70. A particularly compelling set of studies utilized pre- and post-operational photographs of women who underwent micro-fat grafting surgery (Singh et al., 2010; Singh & Randall, 2007). In this procedure, fat cells are removed from the circumference of the waist and transplanted into the buttocks. Body size thus remains unchanged: Only the distribution of fat is altered. For every pair of photographs, men and women judged the postoperative photographs (lower WHR) to be more attractive than the preoperative photographs (higher WHR), a pattern of results that replicated across diverse racial groups (Singh et al., 2010).
Much of the research that tests body shape preferences (WHR) also tests body weight preferences (BMI) (Faries & Bartholomew, 2012; Furnham et al., 2003; Henss, 1995; Singh, 1993; Sugiyama, 2004). Fairly consistently, WHR and BMI both show robust effects on perceptions of attractiveness; however, debate continues over which is the more influential factor (e.g., Singh, 1993, 2006; Tovée & Cornelissen, 2001; Tovée, Hancock, Mahmoodi, Singleton, & Cornelissen, 2002; Tovée et al., 1999; Tovée, Reinhardt, Emery, & Cornelissen, 1998). Effect sizes produced by BMI are frequently larger than those produced by WHR, leading researchers to conclude that BMI is a greater determinant of female physical attractiveness (e.g., Tovée et al., 1998).
We caution against such an inference. For one, there is little practical value in determining whether BMI or WHR is the more influential determinant of attractiveness when it is already known that both body size and body shape affect attractiveness judgments. There are also several methodological problems in this line of research that limit the ability to draw conclusions from a comparison of effect sizes. First, greater variance accounted for by one factor (e.g., BMI) might simply be the result of stimuli varying more widely on that factor than the other (Singh, personal communication; Streeter & McBurney, 2003). Indeed, the figures used in several studies vary widely in weight (emaciated to obese) but very little in WHR (0.68–0.98; Tovée & Cornelissen, 2001; Tovée et al., 1998). When the effect of WHR is evaluated within an ancestrally valid range of body weights (which eliminates obesity as a category because resources were likely never in surplus), WHR more strongly influences perceptions of attractiveness (Furnham et al., 2002, 2005; Furnham, Tan, & McManus, 1997; Singh, 1993). Likewise, WHR would affect attractiveness judgments to a greater extent if the range of WHR values more accurately represented ancestral conditions, where young women were often pregnant and thus possessing WHRs greater than 1.00 (Strassmann, 1997). Finally, clothing might obscure the view of female targets’ WHR more so than their BMI in studies that use images of real women as stimuli (Perilloux, Cloud, & Buss, 2013; Tovée & Cornelissen, 2001; Tovée et al., 1998). Needless to say, if neither WHR nor BMI can be readily assessed, these variables can exert little effect on attractiveness judgments.
Plasticity of Attractiveness Judgments
In spite of mounting evidence that components of physical attractiveness are in large part defined by their health and fertility correlates (for a review, see Sugiyama, 2005), some scholars maintain that standards of beauty are guided by arbitrary dictates of culture (e.g., Wolf, 1991). For instance, in their discussion of changing trends in body size, Voracek and Fisher (2002) diagrammed three women who epitomized female beauty in their respective time periods, each notably thinner than the previous: Hélène Fourment (1636–1638, the wife of Peter Paul Rubens), Marilyn Monroe (1926–1962), and finally supermodel, Eva Herzigova (1973–present). Other examples include research showing a trend toward slenderness for Miss America pageant winners and Playboy centerfolds between the 1960s and 1980s (Garner, Garfinkel, Schwartz, & Thompson, 1980; Mazur, 1986). Fluctuations in ideal body weight (such as those in the examples above) are often used as evidence against evolutionary explanations for attractiveness preferences. These researchers reason that if specific body morphologies are associated with greater reproductive potential, evolution should have shaped attractiveness judgments to be stable across cultures and time periods (e.g., Swami, Gray, et al., 2006).
Such an argument misconstrues the nature of humans’ evolved psychology. Universality is expected at the level of the evolved mechanism, not at the level of its output (Tooby & Cosmides, 1992). In other words, while the algorithms that underlie attractiveness judgments are predicted to be cross-culturally and cross-generationally stable, the output of those algorithms can vary as a function of environmental input. This point is illustrated by the decision rules that underlie trade-offs. When making decisions to optimize one trait over another—as a given woman rarely possesses indicators of good genes, high fertility, and high reproductive value simultaneously—men do not downregulate their preferences for all of the relevant traits. Rather, they systematically adjust certain preferences based on environmental conditions.
For example, in societies where food is scarce or the energetic costs of work are high, a preference for heavy women would direct mating effort toward those who have sufficient fat stores to maintain pregnancy and lactation during times of resource scarcity (Marlowe & Wetsman, 2001). There would be little benefit to mating with women who do not have the fat reserves to support pregnancy, no matter how fit they otherwise appear to be. Consistent with this, studies have demonstrated a preference for overweight women in non-Western samples (Furnham et al., 2002; Sugiyama, 2004; Wetsman & Marlowe, 1999; Yu & Shepard, 1998; but see Singh, 2004). This pattern differs dramatically from the preference for underweight to normal weight women in Western samples (Faries & Bartholomew, 2012; Furnham et al., 2003; Henss, 1995; Schmalt, 2006; Singh, 1993; Singh & Young, 1995; Wilson et al., 2005), where resource streams are so reliable that women can “afford” a lower average body weight. When resources are plenty, women do not need to store excess fat on their bodies to support future pregnancies. As a result, they are able to avoid the negative health outcomes associated with being overweight (e.g., cardiovascular disease; Must et al., 1999). These results demonstrate how environmental circumstances can serve as input to algorithms that generate attractiveness judgments, recalibrating the desired values of various traits based on adaptive trade-offs.
Secondly, environmental input can factor into the attractiveness assessment algorithms of some traits more heavily than others. In particular, preferences for traits that are distally related to fitness consequences might be more permeable to environmental influences than those that are more strongly associated with health and fertility. In the case of BMI and WHR, the results of several studies suggest that the former are more culturally malleable than the latter. In his original study, Singh (1993) reanalyzed the dimensions of Miss America pageant winners and Playboy centerfolds to assess whether preferences for WHR changed over time as they did for BMI (Garner et al., 1980; Mazur, 1986). Replicating the findings of Garner et al. (1980) and Mazur (1986), Singh found a trend for increased thinness in both groups, but despite this reduction in body size, WHR values remained consistent ranging between 0.68 and 0.72. Freese and Meland (2002) further replicated this finding, showing Miss America pageant winners and Playboy centerfolds to have WHR values that consistently fell within the feminine range over a multi-decade span (despite finding a wider range of WHR values than those calculated by Singh). These results, showing BMI preferences to be less cross-culturally and cross-generationally stable than WHR preferences, do not necessarily imply that the former is less a product of evolution than the latter. Algorithms that underlie judgments of optimal body size (i.e., BMI) may simply be designed to incorporate more environmental input than those that underlie judgments of optimal body shape (i.e., WHR). It is likely that the fitness payoffs associated with various body sizes vary more as a function of environment than those associated with various body shapes, which may be more constant across environments.
A common argument used to support the claim that standards of beauty vary across cultures and time periods is the assertion that Europeans considered plump women to be attractive in the sixteenth and seventeenth centuries. Almost exclusively, the evidence presented to justify this widely held belief contrasts Peter Paul Rubens’ (1577–1640) paintings of fat women with present-day idealization of thin women. Swami, Gray, et al. (2006), for example, challenge previous research showing a WHR of 0.70 to be universally attractive, citing as evidence a mean WHR of 0.77 across 30 nude women depicted in paintings by Rubens. We argue that this conclusion is unwarranted for two reasons: (1) An analysis comparing the fatness of women depicted by Rubens and other Baroque artists suggests that Rubens was unusual in his predilection for heavy women (detailed below) and (2) there is nothing special about a WHR of exactly 0.70. Other than the fact that 0.70 happens to fall at the low end of the distribution of feminine WHR values in many modern cultures, there is no systematic reason to expect this WHR to be more attractive than other feminine WHRs. The key point is simply that because WHR distributions overlap very little between the sexes, WHRs that are more clearly in the female distribution should be perceived as optimally attractive (Singh, personal communication).
One of us (J.M.C.), in collaboration with Singh, empirically tested the validity of the claim that plump women were considered attractive in the Baroque era by assessing the proportion of Baroque artists who shared Rubens’ penchant for fat women (Confer & Singh, 2009). If Rubens’ paintings represent a sixteenth- and seventeenth-century European ideal of beauty, a significant proportion of Baroque artists should have also portrayed women as heavyset. If, instead, Rubens portrayals of women were atypical for that era, his paintings may simply reflect his personal taste rather than an overall societal trend. To examine this issue, independent judges (23 men, 29 women) compared 30 European paintings from 1500 to 1650 with a classic Rubens painting (Die drei Grazien; 1639) to determine whether his contemporaries painted women as fat as or fatter than Rubens did. The WHR of the women in each painting was also measured to assess whether Baroque artists preferred a body shape different from an hourglass figure (Singh, 1993).
Figure 7.2 presents the percentages of paintings depicting women with varying degrees of fatness relative to the women depicted in Die drei Grazien (ranging from definitely less fat to definitely more fat). For each 50-year interval between 1500 and 1650, the majority of artists depicted women as less fat than those in Die drei Grazien. These findings indicate that like Picasso’s (1881–1973) unusual depictions of the human form, Rubens portrayed atypical characterizations of women for the Baroque era. The fact that the preponderance of Baroque artists did not idealize a female figure as considerably different from the figure preferred today calls into question the most prevalent example for the argument that standards of beauty are culturally defined.

The percentage of paintings by the sixteenth- and seventeenth-century European artists who depicted women with varying degrees of fatness relative to the women depicted in Rubens’s classic Die drei Grazien (1639)
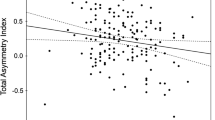
In addition, this analysis corroborates the research described above documenting a preference for women with low WHR. Every portrait selected, including the women depicted in Rubens’ paintings, exhibited WHR values within the feminine range (<0.80; see Fig. 7.3). Thus, despite idiosyncrasies with regard to a woman’s body size (weight), women were never depicted as possessing a masculine body shape (WHR). The results of this study provide further evidence that preferences for some traits (i.e., BMI) may be more culturally malleable than preferences for others (i.e., WHR). Yet even for BMI, a trait that shows relatively high levels of cultural dependency, the disparity between Baroque ideals of body weight and those of modern day appears to be less extreme than originally thought.

The mean WHR of women depicted in the sixteenth- and seventeenth-century European paintings by Rubens (right column) and other contemporary artists (leftmost three columns)
One final point regarding the plasticity of attractiveness judgments is simply that minor fluctuations in the optimum value of a trait (e.g., 0.68 vs. 0.70 WHR; Freese & Meland, 2002) do not provide prima facie evidence against evolutionary explanations of attractiveness. As stated earlier, there is nothing “magical” about a 0.70 WHR (Singh, personal communication). Indeed, there is no evidence to suggest that WHR values of 0.68 or 0.72 are any more or less strongly associated with health and reproductive outcomes than a WHR of 0.70. A much more relevant comparison is between two starkly different WHRs, one from a female distribution and the other from a male distribution. Women with WHRs closer to the male range should be predicted to experience more adverse health and fertility effects than women with WHRs more solidly in the female range. After all, many variations in a woman’s health and reproductive status cause dramatic (not minor) fluctuations from a feminine baseline (except for the possibility that WHR slightly decreases at ovulation; Kirchengast & Gartner, 2002). For example, soon after a woman becomes pregnant, her WHR increases not from 0.70 to 0.72, but from 0.70 to well above 1.00. A similar change in WHR occurs after a woman enters menopause (Singh, 1993, 2006; Singh & Singh, 2011). It is no surprise then that extreme fluctuations in WHR influence judgments of attractiveness more strongly than minor fluctuations, and thus small differences in preferred WHRs across time and space should not be considered incompatible with an evolutionary explanation.
Relative Importance of Facial and Bodily Attractiveness
An enormous amount of research has been devoted to identifying the specific features that make some individuals more physically attractive than others (for a review, see Sugiyama, 2005). Some of these features pertain exclusively to facial attractiveness (e.g., averageness; Langlois & Roggman, 1990), others to bodily attractiveness (e.g., WHR; Singh, 1993), while still others pertain to both facial and bodily attractiveness (e.g., symmetry; Perrett et al., 1999; Thornhill & Gangestad, 1994). Thus far, we have exclusively discussed bodily components of attractiveness; Chaps 4 and 14 of this volume discuss facial components of attractiveness in detail. Recent research has shifted focus away from identifying subcomponents of facial and bodily attractiveness toward evaluating the face and body as whole units of attractiveness (Confer et al., 2010; Currie & Little, 2009; Jonason, Raulston, & Rotolo, 2012; Lu & Chang, 2012). Of particular interest is the relative importance of the face and body in judgments of overall attractiveness, and whether the prioritization of facial or bodily attractiveness is dependent upon mating context (short-term vs. long-term mating; Buss & Schmitt, 1993). Confer et al. (2010) argue that the differential efficacy with which the face and body can convey cues of fertility and reproductive value is of key importance in addressing this issue.
As reviewed above both dimensions of a woman’s reproductive profile—fertility and reproductive value—can be assessed through a number of her bodily features. The same is also true of a woman’s facial features. Fluctuations in facial asymmetry, for example, might indicate whether a woman is ovulating (Scutt & Manning, 1996), an event associated with increased fertility. Other facial features, especially those that are age dependent (e.g., wrinkles and sagginess; Fink, Grammar, & Thornhill, 2001), better indicate a woman’s reproductive value. The face and body, therefore, convey cues of fertility and reproductive value with substantial overlap (Thornhill & Grammer, 1999), as evidenced by a high correlation between facial and bodily attractiveness ratings in real women (Peters, Rhodes, & Simmons, 2007). Even with this high degree of overlap, one component—the face or the body—may convey relatively richer information about a woman’s reproductive condition than the other (Confer et al., 2010). A woman’s body, for example, may better convey information regarding fertility because WHR advertises pregnancy status to a degree that facial features cannot (Singh, 1993). In contrast, information regarding a woman’s reproductive value might be gleaned more effectively from her face where age-dependent features (e.g., full lips; Cunningham, 1986) are most densely concentrated and can be easily observed (e.g., wrinkles, Fink et al., 2001).
Although a man’s reproductive success seems best served by selecting a maximally fertile mate with maximum residual reproductive value, men typically prioritize cues associated with one dimension over the other. This is because the two dimensions peak at different ages—reproductive value at approximately age 17 and fertility at approximately age 24 (Symons, 1979; Williams, 1975)—necessitating a trade-off between women who are at the pinnacle of fertility and others who are at the pinnacle of reproductive value. One factor that influences the priority men place on each dimension is the intended duration of the mateship (short term vs. long term; Buss & Schmitt, 1993). The reproductive success of men pursuing casual sexual dalliances is more directly affected by a potential mate’s current fertility than her future reproductive potential (Buss & Schmitt, 1993). Theoretically then, men should prioritize cues of fertility over cues of reproductive value when evaluating a woman as a short-term mate. The opposite should be true for men evaluating a woman as a long-term mate because transient fluctuations in fertility are less consequential to men’s reproductive success, given the probability of future reproductive opportunities. This is indeed what research has shown (Confer et al., 2010; Currie & Little, 2009; Jonason et al., 2012; Lu & Chang, 2012). Confer et al. (2010) presented men with a picture of a woman whose face was occluded by a “face box” and whose body was occluded by a “body box” (see Fig. 7.4). Men were instructed to evaluate the woman behind the boxes as either a short-term mate or long-term mate; however, they could only remove one box—the face box or the body box—to inform their decision about whether they would engage in the designated relationship with the occluded individual. As predicted, significantly more men assigned to the short-term condition than the long-term condition chose to remove the body box.Footnote 1 In a follow-up analysis, men in this study who were dispositionally oriented more toward short-term mating showed an even stronger preference in the predicted direction, providing additional evidence for the overall pattern. These results are consistent with the hypothesis that indices of fertility, which are of particular importance to men pursuing a short-term relationship, are better assessed through a woman’s body than her face.

Box choice procedure used in Confer et al. (2010). Column A represents the image that is first presented to participants: an opposite-sex individual occluded by a “face box” and a “body box.” Column B represents the image that is presented to participants upon removal of the “face box” (B1) or “body box” (B2). Copyright 2010 by Elsevier
Similar context-dependent shifts in the prioritization of a woman’s bodily attractiveness have been demonstrated through a variety of methodologies. Currie and Little (2009) showed ratings of a woman’s bodily attractiveness to better predict ratings of her overall attractiveness when she was evaluated as a short-term mate than as a long-term mate. Likewise, Perilloux et al. (2013) found that bodily traits (i.e., BMI) better predicted self-perceptions of overall attractiveness among women who pursue casual sexual opportunities (compared to women who pursue long-term, committed relationships), suggesting that women are, at some level, aware of men’s greater prioritization of bodily attractiveness in short-term mating contexts.
Further replicating these results, Jonason et al. (2012) showed men to be more desirous of bodily attractiveness than facial attractiveness in a potential mate, particularly within the context of a short-term relationship. Jonason et al. also conducted a budget allocation study in which participants designed a short-term and long-term mate by distributing a finite number of “mate dollars” across various traits. Men allocated more “mate dollars” to a potential mate’s bodily attractiveness than facial attractiveness, an effect that again was particularly strong within the context of a short-term relationship. Because this procedure requires participants to make trade-offs in their mate preferences—each mate dollar spent on one trait reduces the amount left to spend on other traits—it more accurately represents real-life decision-making processes.
Most recently, Lu and Chang (2012) explored how the prioritization of a woman’s bodily attractiveness in short-term mating contexts affects lower-level attentional processes. In their first experiment, the authors used a visual dot-probe methodology and found that men attended to the waist/hip region of a woman more frequently than her facial region after a short-term mating prime but attended to both regions with equal frequency after a long-term mating prime. A similar pattern of results was found using a change blindness paradigm. Participants were instructed to indicate whether a feature (e.g., clothing accessories) in the waist/hip region or the facial region differed across two otherwise identical images. Preferential attention to one region was inferred from how quickly participants were able to identify the difference. Results indicated that men noticed a change to a woman’s waist/hip region more quickly than a change to her facial region following a short-term mating prime, whereas the opposite was true following a long-term mating prime. Finally, the authors presented participants with an image of a woman’s waist/hip region or an image of a woman’s facial region. Participants were instructed to identify the letter that appeared alongside either image, with response latency indicating participants’ degree of distraction by the image. Men assigned to the short-term mating condition identified the letter more slowly when it was presented alongside a woman’s waist/hip region than alongside her facial region. Men assigned to the long-term mating condition showed no difference in response latency across conditions. Taken together, these results show robust evidence of an adaptive perceptual shift in men to preferentially attend to women’s bodies in short-term mating contexts. Remarkably, this increase in the importance of a woman’s bodily attractiveness occurs in spite of research showing the face to be a better predictor of overall attractiveness than the body generally (i.e., when no differentiation is made between short-term and long-term mating contexts; Furnham & Reeves, 2006; Peters et al., 2007; Riggio, Widaman, Tucker, & Salinas, 1991).
Even as studies that investigate faces and bodies as whole units of attractiveness grow in popularity, much remains to be explored. For instance, does women’s assessment of intrasexual (same-sex) competitors coincide with men’s greater prioritization of bodily attractiveness in short-term mating contexts? Perhaps a mated woman would preferentially attend to the body of an intrasexual competitor, relative to her face, when the competitor in question approaches her partner with a short-term mating opportunity. Doing so would allow women to better simulate the decision-making processes that influence their partners’ desire to take advantage of the mating opportunity and react accordingly. Other research might explore how cross-cultural differences in the availability of short-term mating opportunities predict attitudes regarding the relative importance of a woman’s facial and bodily attractiveness. For example, in populations with a female-biased sex ratio, there are more opportunities for men to engage in short-term relationships (Pedersen, 1991; Schmitt, 2005). Consequently, women may experience greater pressure (e.g., in the media, from peers) to enhance their bodily attractiveness in female-biased populations than in male-biased populations. The approach of examining faces and bodies as whole units of attractiveness does not detract from the study of individual traits, but rather complements it by introducing testable hypotheses to fine-tune our understanding of how attractiveness is assessed.
Conclusions
The bodily traits reviewed in this chapter influence judgments of attractiveness precisely because they communicate fitness-relevant information. (If other bodily traits—such as the elbow—functioned as honest signals of a woman’s health and reproductive status, they too would be sexually arousing.) A high degree of informational overlap is predicted and documented both between and within many bodily traits (Thornhill & Grammer, 1999). For example, fertility can be assessed through the size of a woman’s breasts as well as her WHR because both traits are estrogen-dependent (Jasieńska et al., 2004). This redundancy has been argued to increase the reliability of fitness assessments (Johnstone, 1996). By cross-referencing information conveyed by individual traits—each only probabilistically associated with relevant fitness outcomes—one can triangulate on a more accurate fitness assessment.
Some classes of fitness-relevant information (i.e., indices of fertility vs. reproductive value) may be especially pertinent to the reproductive goals of a short-term or long-term relationship. In such cases, we expect men’s perceptual systems to bias attention toward traits that most effectively convey that information. A growing body of research supports this basic premise. Men attend to a woman’s bodily attractiveness with particular frequency in short-term mating contexts (Confer et al., 2010), where cues of fertility are of greater importance than cues of reproductive value (Buss & Schmitt, 1993). This effect is unlikely to be the result of arbitrary cultural norms or media effects, as even lower-level attentional processes have been shown to manifest the same systematic bias toward bodily attractiveness in short-term relationships (Lu & Chang, 2012). The algorithms that underlie judgments of attractiveness, and the prioritization of various morphological traits, are instead products of evolution. The adaptationist perspective applied throughout this chapter is a powerful theoretical framework that provides functional explanations for why standards of beauty exist in the form that they do. Through a careful consideration of adaptive problems, specialized mechanisms that constitute human mating psychology have been, and will continue to be, discovered.
Notes
- 1.
Women’s box choice was also evaluated. Significantly more women chose to remove the face box than the body box, and this did not differ based on mating context.
References
Anderson, P. (1983). The reproductive role of the human breast. Current Anthropology, 24, 25–45.
Barber, N. (1995). The evolutionary psychology of physical attractiveness: Sexual selection and human morphology. Ethology and Sociobiology, 16, 395–424.
Bleske-Rechek, A., Harris, H. D., Denkinge, K., Webb, R. M., Erickson, L., & Nelson, L. A. (2011). Physical cues of ovulatory status: A failure to replicate enhanced facial attractiveness and reduced waist-to-hip ratio at high fertility. Evolutionary Psychology, 9, 336–353.
Buss, D. M. (1989). Sex differences in human mate preferences: Evolutionary hypotheses tested in 37 cultures. Behavioral & Brain Sciences, 12, 1–49.
Buss, D. M., & Schmitt, D. P. (1993). Sexual strategies theory: A contextual evolutionary analysis of human mating. Psychological Review, 100, 204–232.
Cant, J. (1981). Hypotheses for the evolution of human breasts and buttocks. American Naturalist, 117, 199–204.
Confer, J. C., & Singh, D. (2009). Did Europeans in the sixteenth and seventeenth centuries really idealize fat (Rubenesque) women? Examining the widely held belief. Poster presented at the 10th annual meeting of the Society for Personality and Social Psychology, Tampa, FL.
Confer, J. C., Perilloux, C., & Buss, D. M. (2010). More than just a pretty face: Men’s priority shifts toward bodily attractiveness in short-term versus long-term mating contexts. Evolution and Human Behavior, 31, 348–353.
Cunningham, M. (1986). Measuring the physical in physical attractiveness: Quasi-experiments on the sociobiology of female facial beauty. Journal of Personality and Social Psychology, 50, 925–935.
Currie, T. E., & Little, A. C. (2009). The relative importance of the face and body in judgments of human physical attractiveness. Evolution and Human Behavior, 30, 409–416.
Davey Smith, G., Greenwood, R., Gunnell, D., Sweetnam, P., Yarnell, J., & Elwood, P. (2001). Leg length, insulin resistance, and coronary heart disease risk: The Caerphilly Study. Journal of Epidemiology and Community Health, 55, 867–872.
Dixson, B. J., Grimshaw, G. M., Linklater, W. L., & Dixson, A. F. (2011). Eye tracking of men’s preferences for female breast size and areola pigmentation. Archives of Sexual Behavior, 40, 51–58.
Ellison, P. (1990). Human ovarian function and reproductive ecology: New hypotheses. American Anthropologist, 92, 933–952.
Faries, M. D., & Bartholomew, J. B. (2012). The role of body fat in female attractiveness. Evolution and Human Behavior, 33, 672–681.
Fessler, D. M. T., Haley, K. J., & Lal, R. D. (2005). Sexual dimorphism in foot length proportionate to stature. Annals of Human Biology, 32, 44–59.
Fessler, D. M. T., Stieger, S., Asaridou, S. S., Bahia, U., Cravalho, M., de Barros, P., et al. (2012). Testing a postulated case of intersexual selection in humans: The role of foot size in judgments of physical attractiveness and age. Evolution and Human Behavior, 33, 147–164.
Fink, B., Grammar, K., & Thornhill, R. (2001). Human (Homo sapiens) facial attractiveness in relation to skin texture and color. Journal of Comparative Psychology, 115, 92–99.
Frederick, D. A., Hadji-Michael, M., Furnham, A., & Swami, V. (2010). The influence of leg-to-body ratio (LBR) on judgments of female physical attractiveness: Assessments of computer-generated images varying in LBR. Body Image, 7, 51–55.
Freese, J., & Meland, S. (2002). Seven tenths incorrect: Heterogeneity and change in the waist-to-hip ratio of Playboy centerfold models and Miss America pageant winners. The Journal of Sex Research, 39, 133–138.
Frisch, R. E. (1987). Body fat, menarche, fitness and fertility. Human Reproduction, 2, 521–533.
Furnham, A., Dias, M., & McClelland, A. (1998). The role of body weight, waist-to-hip ratio, and breast size in judgments of female attractiveness. Sex Roles, 39, 311–326.
Furnham, A., McClelland, A., & Omer, L. (2003). A cross-cultural comparison of ratings of perceived fecundity and sexual attractiveness as a function of body weight and waist-to-hip ratio. Psychology, Health, and Medicine, 8, 219–230.
Furnham, A., Moutafi, J., & Baguma, P. (2002). A cross-cultural study on the role of weight and waist-to-hip ratio on female attractiveness. Personality and Individual Differences, 32, 729–745.
Furnham, A., Petrides, K. V., & Constantinides, A. (2005). The effects of body mass index and waist-to-hip ratio of ratings of female attractiveness, fecundity, and health. Personality and Individual Differences, 38, 1823–1834.
Furnham, A., & Reeves, E. (2006). The relative influence of facial neoteny and waist-to-hip ratio on judgments of female attractiveness and fecundity. Psychology, Health & Medicine, 11, 129–141.
Furnham, A., & Swami, V. (2007). Perception of female buttocks and breast size in profile. Social Behavior and Personality, 35, 1–8.
Furnham, A., Swami, V., & Shah, K. (2006). Body weight, waist-to-hip ratio and breast size correlates of ratings of attractiveness and health. Personality and Individual Differences, 41, 443–454.
Furnham, A., Tan, T., & McManus, C. (1997). Waist-to-hip ratio and preferences for body shape: A replication and extension. Personality and Individual Differences, 22, 539–549.
Gallop, G. G. (1982). Permanent breast enlargement in human females: A sociobiological analysis. Journal of Human Evolution, 11, 597–601.
Gangestad, S. W., & Thornhill, R. (1997). The evolutionary psychology of extra-pair sex: The role of fluctuating asymmetry. Evolution and Human Behavior, 18, 69–88.
Garner, D. M., Garfinkel, P. E., Schwartz, D., & Thompson, M. (1980). Cultural expectations of thinness in women. Psychological Reports, 47, 183–191.
Gitter, G., Lomranz, J., Saxe, L., & Bar-Tal, Y. (1983). Perceptions of female physique characteristics by American and Israeli students. The Journal of Social Psychology, 121, 7–13.
Guéguen, N. (2007). Bust size and hitchhiking: A field study. Perceptual and Motor Skills, 105, 1294–1298.
Gunnell, D., May, M., Ben-Shlomo, Y., Yarnell, J., & Davey Smith, G. (2003). Height, leg length, and cancer: The Caerphilly Study. Nutrition and Cancer, 47, 34–39.
Gunnell, D., Whitley, E., Upton, M. N., McConnachie, A., Davey Smith, G., & Watt, G. C. M. (2003). Associations of height, leg length and lung function with cardiovascular risk factors in the Midspan Family Study. Journal of Epidemiology and Community Health, 57, 141–146.
Henss, R. (1995). Waist-to-hip ratio and attractiveness: Replication and extension. Personality and Individual Differences, 19, 479–488.
Henss, R. (2000). Waist-to-hip ratio and female attractiveness. Evidence from photographic stimuli and methodological considerations. Personality and Individual Differences, 28, 501–513.
Jakubowicz, D. J., Iuorno, M. J., Jakubowicz, S., Roberts, K. A., & Nestler, J. E. (2002). Effects of Metformin on early pregnancy loss in the polycystic ovary syndrome. The Journal of Clinical Endocrinology and Metabolism, 87, 524–529.
Jasieńska, G., Ziomkiewicz, A., Ellison, P. T., Lipson, S. F., & Thune, I. (2004). Large breasts and narrow waists indicate high reproductive potential in women. Proceedings of the Royal Society of London B, 271, 1213–1217.
Johnstone, R. A. (1996). Multiple displays in animal communication: ‘Backup signals’ and ‘multiple messages’. Philosophical Transactions of the Royal Society of London B, 351, 329–338.
Jonason, P. K., Raulston, T., & Rotolo, A. (2012). More than just a pretty face and a hot body: Multiple cues in mate choice. The Journal of Social Psychology, 152, 174–184.
Karremans, J. C., Frankenhuis, W. E., & Arons, S. (2010). Blind men prefer low waist-to-hip ratio. Evolution and Human Behavior, 31, 182–186.
Kirchengast, S., & Gartner, M. (2002). Changes in fat distribution (WHR) and body weight across the menstrual cycle. Collegium Antropologicum, 26, 47–57.
Langlois, J. H., & Roggman, L. A. (1990). Attractive faces are only average. Psychological Science, 1, 115–121.
Lassek, W. D., & Gaulin, S. J. C. (2008). Waist-hip ratio and cognitive ability: Is gluteofemoral fat a privileged store of neurodevelopmental resources? Evolution and Human Behavior, 29, 26–34.
Li, N. P., Bailey, J. M., Kenrick, D. T., & Linsenmeier, J. A. W. (2002). The necessities and luxuries of mate preferences: Testing the tradeoffs. Journal of Personality and Social Psychology, 82, 947–955.
Lipson, S. F., & Ellison, P. T. (1996). Comparison of salivary steroid profiles in naturally occurring conception and non-conception cycles. Human Reproduction, 11, 2090–2096.
Lu, H. J., & Chang, L. (2012). Automatic attention towards the face or body as a function of mating motivation. Evolutionary Psychology, 10, 120–135.
Manning, J. T., Scutt, D., Whitehouse, G. H., & Leinster, S. J. (1997). Breast asymmetry and phenotypic quality in women. Evolution and Human Behavior, 18, 223–236.
Marlowe, F. (1998). The nubility hypothesis: The human breast as an honest signal of residual reproductive value. Human Nature, 9, 263–271.
Marlowe, F., Apicella, C., & Reed, D. (2005). Men’s preferences for women’s profile waist-to-hip ratio in two societies. Evolution and Human Behavior, 26, 458–468.
Marlowe, F., & Wetsman, A. (2001). Preferred waist-to-hip ratio and ecology. Personality and Individual Differences, 30, 481–489.
Marti, B., Tuomilehto, J., Soloman, V., Kartovaara, L., Korhonen, H. J., & Pietinen, P. (1991). Body fat distribution in the Finnish population: Environmental determinants and predictive power for cardiovascular risk factor levels. Journal of Epidemiology and Community Health, 45, 131–137.
Mazur, A. (1986). U.S. trends in feminine beauty and overadaptation. Journal of Sex Research, 22, 281–303.
Møller, A. P., Soler, M., & Thornhill, R. (1995). Breast asymmetry, sexual selection, and human reproductive success. Ethology and Sociobiology, 16, 207–219.
Must, A., Spadano, J., Coakley, E. H., Field, A. E., Colditz, G., & Dietz, W. H. (1999). The disease burden associated with overweight and obesity. The Journal of the American Medical Association, 282, 1523–1529.
Pedersen, F. A. (1991). Secular trends in human sex ratios: Their influence on individual and family behavior. Human Nature, 2, 271–291.
Penke, L., & Asendorpf, J. B. (2008). Beyond global sociosexual orientations: A more differentiated look at sociosexuality and its effects on courtship and romantic relationships. Journal of Personality and Social Psychology, 95, 1113–1135.
Perilloux, C., Cloud, J. M., & Buss, D. M. (2013). Women’s physical attractiveness and short-term mating strategies. Personality and Individual Differences, 54, 490–495.
Perrett, D. I., Burt, M., Penton-Voak, I. S., Lee, K. J., Rowland, D. A., & Edwards, R. (1999). Symmetry and human facial attractiveness. Evolution and Human Behavior, 20, 295–307.
Peters, M., Rhodes, G., & Simmons, L. W. (2007). Contributions of the face and body to overall attractiveness. Animal Behavior, 73, 937–942.
Riggio, R. E., Widaman, K. F., Tucker, J. S., & Salinas, C. (1991). Beauty is more than skin deep: Components of attractiveness. Basic and Applied Social Psychology, 12, 423–439.
Saad, G. (2008). Advertised waist-to-hip ratios of online female escorts: An evolutionary perspective. International Journal of e-Collaboration, 4, 40–50.
Schmalt, H. D. (2006). Waist-to-hip ratio and female physical attractiveness: The moderating role of power motivation and the mating context. Personality and Individual Differences, 41, 455–465.
Schmitt, D. P. (2005). Sociosexuality from Argentina to Zimbabwe: A 48-nation study of sex, culture, and strategies of human mating. Behavioral and Brain Sciences, 28, 247–311.
Scutt, D., & Manning, J. T. (1996). Symmetry and ovulation in women. Human Reproduction, 11, 2477–2480.
Singh, D. (1993). Adaptive significance of female physical attractiveness: Role of waist-to-hip ratio. Journal of Personality and Social Psychology, 65, 293–307.
Singh, D. (1994a). Ideal female body shape: Role of body weight and waist-to-hip ratio. International Journal of Eating Disorders, 16, 283–288.
Singh, D. (1994b). Is thin really beautiful and good? Relationship between waist-to-hip ratio (WHR) and female attractiveness. Personality and Individual Differences, 16, 123–132.
Singh, D. (2002). Female mate value at a glance: Relationship of waist-to-hip ratio to health, fecundity, and attractiveness. Neuroendocrinology Letters, 23, 81–91.
Singh, D. (2004). Mating strategies of young women: Role of physical attractiveness. The Journal of Sex Research, 41, 43–54.
Singh, D. (2006). Universal allure of the hourglass figure: An evolutionary theory of female physical attractiveness. Clinics in Plastic Surgery, 33, 359–370.
Singh, D., Dixson, B. J., Jessop, T. S., Morgan, B., & Dixson, A. F. (2010). Cross-cultural consensus for waist-hip ratio and women’s attractiveness. Evolution and Human Behavior, 31, 176–181.
Singh, D., & Luis, S. (1995). Ethnic and gender consensus for the effect of waist-to-hip ratio on judgment of women’s attractiveness. Human Nature, 6, 51–65.
Singh, D., & Randall, P. (2007). Beauty is in the eye of the plastic surgeon: Waist-to-hip ratio (WHR) and women’s attractiveness. Personality and Individual Differences, 43, 329–340.
Singh, D., Renn, P., & Singh, A. (2007). Did the perils of abdominal obesity affect depiction of feminine beauty in the sixteenth to eighteenth century British literature? Exploring the health and beauty link. Proceedings of the Royal Society B, 274, 891–894.
Singh, D., & Singh, D. (2011). Shape and significance of feminine beauty: An evolutionary perspective. Sex Roles, 64, 723–731.
Singh, D., & Young, R. K. (1995). Body weight, waist-to-hip ratio, breasts, and hips: Role in judgments of female attractiveness and desirability for relationships. Ethology and Sociobiology, 16, 483–507.
Sorokowski, P., & Pawlowski, B. (2008). Adaptive preferences for leg length in a potential partner. Evolution and Human Behavior, 29, 86–91.
Sorokowski, P., Sorokowska, A., & Mberira, M. (2012). Are preferences for legs length universal? Data from a semi-nomadic Himba population from Namibia. The Journal of Social Psychology, 152, 370–378.
Sorokowski, P., Szmajke, A., Sorokowska, A., Cunen, M. B., Fabrykant, M., Zarafshani, K., et al. (2011). Attractiveness of leg length: Report from 27 nations. Journal of Cross-Cultural Psychology, 42, 131–139.
Strassmann, B. I. (1997). The biology of menstruation in Homo sapiens: Total lifetime menses, fecundity, and nonsynchrony in a natural-fertility population. Current Anthropology, 38, 123–129.
Streeter, S. A., & McBurney, D. H. (2003). Waist-to-hip ratio and attractiveness: New evidence and a critique of “a critical test”. Evolution and Human Behavior, 24, 88–98.
Sugiyama, L. S. (2004). Is beauty in the context-sensitive adaptations of the beholder? Shiwiar use of waist-to-hip ratio in assessments of female mate value. Evolution and Human Behavior, 25, 51–62.
Sugiyama, L. S. (2005). Physical attractiveness in adaptationist perspective. In D. M. Buss (Ed.), The handbook of evolutionary psychology (pp. 292–343). Hoboken, NJ: Wiley.
Swami, V., Einon, D., & Furnham, A. (2006). The leg-to-body ratio as a human aesthetic criterion. Body Image, 3, 317–323.
Swami, V., Einon, D., & Furnham, A. (2007). Cultural significance of leg-to-body ratio preferences? Evidence from Britain and rural Malaysia. Asian Journal of Social Psychology, 10, 265–269.
Swami, V., Furnham, A., & Joshi, K. (2008). The influence of skin tone, hair length, and hair colour on ratings of women’s physical attractiveness, health, and fertility. Scandinavian Journal of Psychology, 49, 429–437.
Swami, V., Gray, M., & Furnham, A. (2006). The female nude in Rubens: Disconfirmatory evidence of the waist-to-hip ratio hypothesis of female physical attractiveness. Imagination, Cognition, and Personality, 26, 139–147.
Symons, D. (1979). The evolution of human sexuality. New York: Oxford University Press.
Tassinary, L. G., & Hansen, K. A. (1998). A critical test of the waist-to-hip ratio hypothesis of female physical attractiveness. Psychological Science, 9, 150–155.
Thompson, J. K., & Tantleff, S. (1992). Female and male ratings of upper torso: Actual, ideal, and stereotypical conceptions. Journal of Social Behavior and Personality, 7, 345–354.
Thornhill, R., & Gangestad, S. W. (1994). Human fluctuating asymmetry and sexual behavior. Psychological Science, 5, 297–302.
Thornhill, R., & Grammer, K. (1999). The body and face of woman: One ornament that signals quality? Evolution and Human Behavior, 20, 105–120.
Tooby, J., & Cosmides, L. (1992). The psychological foundations of culture. In J. Barkow, L. Cosmides, & J. Tooby (Eds.), The adapted mind: Evolutionary psychology and the generation of culture. New York: Oxford University Press.
Tovée, M. J., & Cornelissen, P. L. (1999). The mystery of female beauty. Nature, 399, 215–216.
Tovée, M. J., & Cornelissen, P. L. (2001). Female and male perceptions of female physical attractiveness in front-view and profile. British Journal of Psychology, 92, 391–402.
Tovée, M. J., Hancock, P. J. B., Mahmoodi, S., Singleton, B. R. R., & Cornelissen, P. L. (2002). Human female attractiveness: Waveform analysis of body shape. Proceedings of the Royal Society of London B, 269, 2205–2213.
Tovée, M. J., Maisey, D. S., Emery, J. L., & Cornelissen, P. L. (1999). Visual cues to female sexual attractiveness. Proceedings of the Royal Society of London B, 266, 211–218.
Tovée, M. J., Reinhardt, S., Emery, J. L., & Cornelissen, P. L. (1998). Optimum body-mass index and maximum sexual attractiveness. The Lancet, 352, 548.
Venners, S. A., Liu, X., Perry, M. J., Korrick, S. A., Li, Z., Yang, F., et al. (2006). Urinary estrogen and progesterone metabolite concentrations in menstrual cycles of fertile women with non-conception, early pregnancy loss or clinical pregnancy. Human Reproduction, 21, 2272–2280.
Voracek, M., & Fisher, M. L. (2002). Shapely centerfolds? Temporal change in body measures: Trend analysis. British Medical Journal, 325, 1447–1448.
Voracek, M., Fisher, M. L., Rupp, B., Lucas, D., & Fessler, D. M. T. (2007). Sex differences in relative foot length and perceived attractiveness of female feet: Relationships among anthropometry, physique, and preference ratings. Perceptual and Motor Skills, 104, 1123–1138.
Wetsman, A., & Marlowe, F. (1999). How universal are preferences for female waist-to-hip ratios? Evidence from the Hadza of Tanzania. Evolution and Human Behavior, 20, 219–228.
Williams, G. C. (1975). Sex and evolution. Princeton, NJ: Princeton University Press.
Wilson, J. M. B., Tripp, D. A., & Boland, F. J. (2005). The relative contributions of waist-to-hip ratio and body mass index to judgments of attractiveness. Sexualities, Evolution, and Gender, 7, 245–267.
Wolf, N. (1991). The beauty myth: How images of female beauty are used against women. New York: William Morrow.
Yu, D. W., & Shepard, G. H. (1998). Is beauty in the eye of the beholder? Nature, 396, 321–322.
Zahavi, A. (1975). Mate selection: A selection for a handicap. Journal of Theoretical Biology, 53, 205–214.
Zelazniewicz, A. M., & Pawlowski, B. (2011). Female breast size attractiveness for men as a function of sociosexual orientation (restricted vs. unrestricted). Archives of Sexual Behavior, 40, 1129–1135.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Additional information
Dedicated to the memory of Devendra Singh (1938–2010)
Rights and permissions
Copyright information
© 2014 Springer Science+Business Media New York
About this chapter
Cite this chapter
Cloud, J.M., Perilloux, C. (2014). Bodily Attractiveness as a Window to Women’s Fertility and Reproductive Value. In: Weekes-Shackelford, V., Shackelford, T. (eds) Evolutionary Perspectives on Human Sexual Psychology and Behavior. Evolutionary Psychology. Springer, New York, NY. https://doi.org/10.1007/978-1-4939-0314-6_7
Download citation
DOI: https://doi.org/10.1007/978-1-4939-0314-6_7
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4939-0313-9
Online ISBN: 978-1-4939-0314-6
eBook Packages: Behavioral ScienceBehavioral Science and Psychology (R0)