
Abstract
Cancer is a multistep process involving genetic and epigenetic changes in the somatic genome. Genetic mutations as well as environmental factors lead to the initiation, promotion, and progression of cancer. Cancer progression ends in tumor metastasis to distant sites, and metastasis is the major reason for cancer patient deaths. Recent experimental evidence suggests the pivotal role of cancer stem cells. A tumor is heterogeneous and composed of different cell types. The cancer stem cells in the tumor have the capacity both to self-renew and differentiate to sustain the tumor. Features of cancer stem cells are described in this review, with an emphasis on the role that dietary phytochemicals may play in cancer chemoprevention. Ingredients in the diet can inhibit cancer cells and cancer stem cells. These compounds include curcumin from curry, epigallocatechin gallate from green tea, resveratrol from red wine, genistein from soy, sulforaphane from broccoli, and many others. Current research findings advocate the beneficial effects towards cancer chemoprevention via uptake of a combination of different dietary phytochemicals.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
- Phytochemicals
- Cancer stem cells
- Chemoprevention
- Curcumin
- Epigallocatechin gallate
- Resvertrol
- Genistein
- Sulforaphane
- Combination therapy
Cancer and Cancer Stem Cells
Biology of Cancer
A somatic cell undergoes changes in its genome and epigenome during its lifetime. If the accumulated changes involve regulation of cell growth and death, the resulting uncharted growth may alter the normal somatic cell to turn into a cancer cell. During the carcinogenesis process, there are malfunctions of genes, including gain of function of proto-oncogenes and loss of function of tumor suppressor genes .
Cancer cells exhibit hallmark alterations in their physiology, namely: (1) self-sufficiency of growth signals, (2) insensitivity to growth inhibitory signals, (3) evasion of programmed cell death , (4) limitless replicative potential, (5) reprogrammed cellular energetics, (6) induction of angiogenesis, (7) presence of tumor-promoting inflammation, (8) avoidance of immune destruction, (9) maintenance of genome instability and mutation, and (10) activation of tissue invasion and metastasis (Hanahan and Weinberg 2000, 2011) .
Because cancer is a multistep process, it may take decades to develop. Carcinogenesis can be traced from its initiation, followed by promotion, progression, and metastasis. Accumulation of a handful of mutations (10–20) is adequate for cancer development. The cancer cell will then self-renew, migrate, and invade, as a clonal expansion in its complex tissue ecosystem (Greaves and Maley 2012) . The niche or microenvironment surrounding the cancer cell can shape the path of carcinogenesis; as an example, the host immune system will attempt to destroy the developing cancer, although results may vary, as seen in the outcomes of cancer immunoediting (Schreiber et al. 2011) . The tumor evolves from being benign to being malignant, and finally leads to the death of its host, the cancer patient. Investigators are interested in details of this process of carcinogenesis, and ways to halt its progression. Dietary phytochemicals have been shown to modulate the carcinogenetic procession.
Cancer Stem Cells
History and Definition
By histological examination , the tumor is heterogeneous, composed of tumor cells at various stages of cell differentiation, plus nontumor cells such as fibroblasts and immune cells. Thus, not all cells within a tumor are equal. Clonal evolution and competition within the tumor result in the dominance of the cell that is resistant to therapy and can both self-renew and differentiate its progeny cells. This observed tumor heterogeneity has been explained by the cancer stem cell (CSC) hypothesis . A workshop of the American Association for Cancer Research in 2006 settled with a consensus definition: CSC “is a cell within a tumor that possesses the capacity to self-renew and to cause the heterogeneous lineages of cancer cells that comprise the tumor” (Clarke et al. 2006) . The putative CSC is also known as a tumor-initiating cell , a term preferred by some investigators, as seen in this example: “tumor-initiating cells (popularly known as cancer stem cells)” (Zhou et al. 2009) . In this chapter, we use the popular term, CSC.
In a review published in 2001, the abstract starts with “stem cell biology has come of age” but ends with “cancer cells may include ‘cancer stem cells’—rare cells with indefinite potential for self-renewal that drive tumorigenesis” (Reya et al. 2001) . Research on various aspects of CSCs has been reported in the intervening years. In addition to research and review publications, CSCs are the topic of edited monographs. The first book dedicated to CSCs came out in 2009 (Bapat 2009) ; its editor was a scientist from India who first isolated human ovarian CSCs (Bapat et al. 2005) . Other volumes subsequently appeared (Farrar 2010; Allan 2011; Scatena et al. 2012) . One addressed CSC methods (Yu 2009) . There are even Web-published volumes (Shostak 2011a, 2011b) and a series dedicated to cancer cells and CSCs (Hayat 2012) . So, has CSC biology come of age?
In reality, the CSC hypothesis and the CSC concept have a long history. The subject has been billed as “old concepts, new insights” (Vermeulen et al. 2008) and “an evolving concept” (Nguyen et al. 2012) . Linking stem cells and cancer, histological similarities were noted between embryonic and tumor tissues in the nineteenth century. This became the “embryonic rest” theory: cancers arise from cells with properties similar to those of early embryos. Later investigations concluded that cancer can be viewed as a “caricature” of normal development. The uncharted cell proliferation seen in cancer is the result of distortion of normal development, and a lack of coordination between growth and differentiation (Nguyen et al. 2012) . In addition, knowledge on stem cell biology in normal tissues has been translated to the concept of CSCs in cancerous tissues.
Perhaps the best known normal stem cell is the hematopoietic stem cell (HSC). HSC has a hierarchy that permits an organized process of self-renewal and differentiation, giving rise to progeny of differentiated cell types including erythrocytes, macrophages, lymphocytes and many others. Immunologists have identified cluster of differentiation (CD) markers for the different cell types, and these cell surface antigens allow for the isolation of cells by methods such as fluorescence-activated cell sorting (FACS) . The rare HSC exhibits the surface marker CD34, the member of a family of single-pass transmembrane sialomucin proteins. It should be noted that although CD34+ is seen in human HSC, the mouse equivalent is CD34− or CD34low. Thus, there are species differences in CD distribution.
In studying hematopoietic malignancies , in 1937, Jacob Furth and Morton Kahn provided the first quantitative assay for the assessment of the frequency of cancer cells maintaining the hematopoietic tumor. They showed that a single mouse leukemic cell was capable of transmitting the systemic disease when transplanted into a recipient mouse (as quoted by Clevers 2011) . By definition, this has to be the CSC of murine leukemia.
Like its murine counterpart, the human CSC was first demonstrated in leukemia and then extended to solid tumors. Bonnet and Dick (1997) reported the presence of CD34+/CD38− CSCs in human acute myeloid leukemia. Isolated patient CSCs will reconstitute the leukemia when transplanted in immunodeficient mice. Al-Hajj et al. (2003) reported the presence of CD44+/CD24−/lowCSCs in human breast cancer. They wrote: “As few as 100 cells with this phenotype were able to form tumors in mice, whereas tens of thousands of cells with alternate phenotypes failed to form tumors.” Singh et al. (2003) reported the presence of CD133+ CSCs in human brain cancer. They commented:
The increased self-renewal capacity of the brain tumor stem cell (BTSC) was highest from the most aggressive clinical samples of medulloblastoma compared with low-grade gliomas. The BTSC was exclusively isolated with the cell fraction expressing the neural stem cell surface marker CD133. These CD133+ cells could differentiate in culture into tumor cells that phenotypically resembled the tumor from the patient.
These initial studies on CSCs were expanded to many cancers in the following decade.
Biomarkers and Experimental Studies
Cellular antigens of CSCs can serve as biomarkers. The more common ones are CD24, CD34, CD44, and CD133 (Woodward and Sulman 2008; Keysar and Jimeno 2010) . They are used in the isolation of CSCs from leukemia, breast cancer, and many other cancers. CD24 is a heat-stable antigen, a sialoprotein that acts as a ligand for P–selectin, thus enabling the cell to bind to platelets and protecting tumor cells in the blood stream. Although CD24 is negative in breast CSCs, it is present in ovarian CSCs (Gao et al. 2010) . CD44 is a transmembrane glycoprotein which is a hyaluronic acid receptor. Besides breast cancer, CD44 is found in CSCs from pancreatic, gastric, head and neck, ovarian, and colon cancer. CD133, also named prominin-1, is a glycoprotein consisting of five transmembrane domains with a restricted expression within the plasma membrane protrusion sites. In addition to brain cancer, CD133 is found in CSCs from colon, liver, pancreatic, and prostate cancer (Alison et al. 2011; Zobalova et al. 2011; Hu and Fu 2012) . It should be noted that these antigens are also present in the respective normal stem cells, although they have been used as biomarkers for their cancerous counterparts. Hence, the significance of CD24 and CD44 as CSC markers has been seen as “an enduring ambiguity” (Jaggupilli and Elkord 2012) . Another important point is the lack of a single CD antigen common to all CSCs. However, a universal marker may be close at hand. In a recent drug screening study specifically targeting CSCs, dopamine receptors have been discovered as a biomarker for CSCs but not normal stem cells. Because only leukemic and breast CSCs have been tested so far, the novel biomarker requires confirmation and validation (Sachlos et al. 2012) .
In addition to identification through biomarkers, CSCs can also be characterized by functional assays, namely, the detection of side population (SP) activity and the assessment of aldehyde dehydrogenase (ALDH) activity (Keysar and Jimeno 2010; Tirino et al. 2013) . The SP assay measures the ability of cells to expel the fluorescent dye, Hoechst 33342, caused by the activity of ATP-dependent drug transporters on the plasma membrane, especially the ABCG2 transporter (Mo and Zhang 2012) . As analyzed by flow cytometry, cells with fast drug exit form a side population, as opposed to the main population composed of the majority of cells. Initially identified in HSCs, SP has been found in many CSCs. Similarly, ALDH is a detoxifying enzyme and acts by the oxidation of aldehydes to carboxylic acids for further metabolism or liver exit. A fluorescent substrate assay using ALDEFLUOR, biodipy-aminoacetaldehyde (BAAA), allows for the isolation of ALDH+ cells. The ALDH assay was initially used to isolate HSCs and normal breast stem cells, and then extended to CSCs (Ginestier et al. 2007) . Inasmuch as both SP and ALDH assays involve drug extrusion and metabolism, exhibiting these capacities indicates drug resistance of normal and CSCs. For example, SP and ALDH assays, in addition to specific biomarker (such as CD44 and CD133) screening, have been utilized for the identification of human ovarian CSCs (as summarized by Bapat 2010) . However, it should be emphasized that these assays only enrich CSCs. The biomarker and functional assays may not characterize all the CSCs in the analyzed samples; therefore, additional CSCs not exhibiting the attribute being tested may still be present. For a list of biomarkers currently being used in the isolation of cancer stem cells, please see Table 1.
In addition to biomarkers and functional assays, another in vitro method to study CSCs is the use of serum-free spheres or spheroid cultures. Tissue culture cells are usually grown as a monolayer in a nutrient-rich medium containing fetal bovine serum as a source of the necessary growth factors and other components. However, in the absence of serum but in the presence of growth factors such as fibroblast growth factor (FGF) and epidermal growth factor (EGF), cells will grow as spheres in suspension in unattached/untreated tissue culture plastic dishes, flasks, or plates. Spheroid cultures mimic the three-dimensional nature of a tissue. Furthermore, it should be noted that oxygen will be less available to the cells located at the interior of a sphere. The hypoxic condition may modulate their differentiated state: towards stemness. Originally developed for neurobiological studies, the spheroid culture is used to “identify stem cells based on their reported capacity to evaluate self-renewal and differentiation at the single-cell level in vitro” (Pastrana et al. 2011) . Spheroid cultures have been developed for normal tissues and tumors (Chen et al. 2012a) . When isolated, individual sphere-forming cells from a tumor will form secondary and tertiary spheres upon sequential subcultures; by operational definition these will be CSCs. However, there have been critiques on this spheroid assay. Spheres are prone to aggregate. Hence, cell density will influence clonality. The quiescent stem cell may be missed by this method (Pastrana et al. 2011) . In fact, the quiescent nature of putative normal and CSCs has been utilized for their characterization. These cells are known as label-retaining cells, because they can retain labels such as the lipophilic dye PKH26, which is diluted in subsequent cell divisions but not so with slow-dividing cells (Pece et al. 2010; Xin et al. 2012) .
From a tumor sample, putative CSCs may be characterized by the in vitro techniques discussed above. In addition to isolating CSCs from tumor samples, it is also possible to isolate them from established cancer cell lines (Drewa et al. 2011; Mather 2012) . The advantage of the latter is the absence of nontumor cells as contaminants; the disadvantage is the additional accumulated changes during the long period of in vitro culture. As an example, our interest in CSCs and SP analysis led us to isolate SPs from the rat C6 glioma cell line, deriving putative CSCs from an established cell line (Fong et al. 2010) . However, the C6 stemness state is dynamic: although SPs give rise to both SP and non-SP progenies, as expected, non-SPs can do the same (Fong et al. 2010; Fong and Chan 2012) .
Complementary to in vitro studies of CSCs , in vivo models are available. The isolated cells are tested by their ability to initiate new tumor growth after xenotransplantation into immunocompromised mice (Cheng et al. 2010) . Limiting dilution xenotransplantation yields an estimate of CSC abundance in the tumor sample; sequential transplantation yielding the original tumor will confirm the presence of CSCs. Different murine models with varying degrees of immunodeficiency have been applied to CSC studies. Examples include athymic nude mice, nonobese diabetic/severe combined immunodeficiency (NOD/SCID) mice, and NOD/SCID interleukin-2 receptor gamma chain null (Il2rg −/−) mice. For example, using the severely immunodeficient NOD/SCID Il2rg −/− mice in single-cell transplants (in an appropriate extracellular matrix, Matrigel) from human melanoma samples, 27% of unselected melanoma cells develop tumors in mice (Quintana et al. 2008) . Does this finding detect the abundance of human skin CSCs, whereas CSCs from other cancers are rare? There have been critiques on this xenotransplantation assay. Placing human cancer cells in immunodeficient mice is artificial because the human cancer cell will not encounter its normal host immune response. Therefore, results from this assay cannot reflect the physiological fate of the CSC in its native environment. Despite these objections, the limiting dilution murine in vivo xenotransplantation assay has been held as the “gold standard” for CSC identification (Ghiaur et al. 2012) . However, other in vivo assays have been attempted for identifying CSCs. One example is the zebrafish model (Blackburn et al. 2011) . Nonetheless, we found this quote: “despite being considered the gold standard assay for CSCs by many in the field, there is no reason to assume that growth in immunocompromised mice is in fact a relevant assay for CSC activity” (Ghiaur et al. 2012) .
Controversy and Evidence
Even now, the CSC hypothesis is controversial. In 2009, one review asked: “Cancer stem cells: mirage or reality?” (Gupta et al. 2009a) ; another discussed: “Cancer stem cells and cancer nonstem cells: from adult stem cells or from reprogramming of differentiated somatic cells” (Trosko 2009) ; a third commented: “The investigation and study of cancer stem cells (CSCs) have received enormous attention over the past 5–10 years but remain topics of considerable controversy” (Rosen and Jordan 2009) . Similar statements were made in 2013: “The investigation and study of CSCs have received enormous attention only over the past 5–10 years and remain topics of considerable controversy. Opinions about the validity of the CSC hypothesis, the biological properties of CSCs, and the relevance of CSCs to cancer therapy differ widely” (Tirino et al. 2013) .
However, there is accumulating evidence favoring the CSC hypothesis. The CSCs may explain concepts of cancer and therefore are relevant to cancer therapy. The major supporting data are: (1) the presence of CSCs in minimal residual disease (MRD) and (2) the demonstration of CSCs in cell lineage tracing studies of murine tumors.
MRD is a term first used in leukemia, to denote the small numbers of leukemic cells that remain in the patient during or after treatment, when the patient is in remission (no symptoms/signs of disease). It is the major cause of relapse in cancer. MRD has been applied to solid tumors. If CSCs are resistant to therapy, they should be enriched after chemo- and radiotherapy and found in MRD. This has been shown to be the case. For example, resident breast CSC populations surviving conventional treatment (such as docetaxel) have been found to be enriched for CD44+/CD24−/low breast CSCs that express epithelial-mesenchymal-transition (EMT)–associated genes (Creighton et al. 2009) . Thus, the presence of CSCs after therapy predicts recurrence (Ghiaur et al. 2012) .
Lineage tracing is a common technique for studying cell origins in developmental biology. Tracking cells with fluorescent proteins such as green fluorescent protein (GFP) in three different murine solid tumors, as reported in 2012, has been cheered as “resolving the stem-cell debate” (Gilbertson 2012; Graham 2012) . In murine intestinal adenoma, 5–10% of the cells were CSCs, exhibiting the marker leucine-rich repeat-containing heterotrimeric guanine nucleotide-binding protein-coupled receptor 5 (Lgr5). The cell lineage tracing was made possible through the use of Cre-reporter at the Rosa26-Confetti locus, showing fluorescence (with four colors: green, red, yellow, blue) and even color conversion during tumor development. Lgr5 cells generated additional Lgr5 cells, as well as other adenoma cell types (Schepers et al. 2012) . In murine papilloma, a benign skin tumor, 20% of cells were stem cells (tracked by yellow fluorescent protein) that divided twice a day whereas the others became terminally differentiated tumor cells (Driessens et al. 2012) . In murine glioblastoma, a transgene was created to label both the quiescent adult neural stem cells and a subset of the endogenous glioma tumor cells (expressing GFP). The transgene also contained a viral thymidine kinase gene that could be targeted by the drug ganciclovir. Gliomas were treated with the drug temozolomide (TMZ) ; but TMZ treatment alone led to the regrowth of a subpopulation of CSCs, that were then controlled by ganciclovir. TMZ-ganciclovir cotreatment impeded tumor development, by destroying both cancer cells and CSCs. Hence, this last study demonstrated the existence of murine glioma CSCs and their selective targeting (Chen et al. 2012b) .
Resistance to Therapy and Stem Cell Pathways
With the assumption that findings in mice are extrapolatable to humans, the demonstration of CSCs in murine glioma and TMZ-ganciclovir cotreatment shows clinical relevance of CSCs. CSCs are resistant to therapy; they are or become chemo- and radio-resistant during or after therapeutic treatments (Donnenberg and Donnenberg 2005; Krause et al. 2011) . These characteristics are due to the activity of drug transporters and metabolism enzymes, and a DNA repair system activated by genomic instability. CSCs may possess less reactive oxygen species (ROS) , and thus are less susceptible to radiation therapy (Diehn et al. 2009) . Depending on individual cases of cancer, CSCs may arise from either mutated normal stem cells, or dedifferentiated cancer cells exhibiting stem cell features. They display pathways of gene expression in common with those of normal stem cells. Therefore, thinking along therapeutic approaches, compounds targeting CSCs must be capable of differentiating them from the normal stem cells and sparing the latter, otherwise unforeseen problems with normal tissue homeostasis can occur.
Several signal transduction pathways are active in CSCs and may be amenable for intervention. The self-renewal pathways seen in CSCs relate to the expression of proteins involved in Hedgehog, Wnt, and Notch signaling. Additional pathways include PI3K and NFκB pathways (Garvalov and Acker 2011; Alison et al. 2011, 2012; Hu and Fu 2012) .
The Hedgehog (Hh) signaling pathway starts with a secreted morphogenetic factor. The term Hh comes from the fruit fly genetic mutation Hh that leads to spiny-looking larva; the gene is essential for arthropod segmentation and mammalian development. The mammalian Hh morphogen, as a ligand, binds to its receptor, Patched 1. This binding activates another plasma membrane protein, Smoothened, which eventually leads to activation of the transcription factor known as Gli (glioma).
The Wnt signaling pathway also starts with a secreted morphogenetic factor. The term Wnt comes from the fruit fly genetic mutation Wingless (Wg), which is important for arthropod polarity and segmentation, and the murine gene Integration 1 (Int1), a gene activated in breast cancer of mice infected with mouse mammary tumor virus. Wnt morphogen binds to its receptor, and after a series of intermediate steps, results in the mobilization of a cytoskeletal protein, beta-catenin, from the cytoplasm to the nucleus to activate its specific transcription factor known as lymphoid enhancer binding factor/T-cell factor (LEF/TCF).
Instead of secreted factors, the Notch signaling pathway starts with the membrane-associated Notch protein. The term Notch also comes from a fruit fly genetic mutation, which results in a notch in the fly wing. The binding of Notch by its ligand, such as the membrane protein Delta from a neighboring cell, initiates the pathway. Notch protein undergoes limited proteolysis by the proteinase named gamma-secretase to yield the Notch intracellular domain (NICD) , which mobilizes from the cytoplasm to the nucleus to activate its specific transcription factor named recombination signal binding protein for immunoglobulin kappa J (RBPJ), alias CSL (after the mammalian centromere promoter binding factor 1, CBF-1, the fruit fly suppressor of hairless mutation, and the nematode lag-2 gene). Notch gene mutations/polymorphisms have been found in cancer patients, and may be involved in CSC chemoresistance (Crea et al. 2011) .
The three signaling pathways initiated by Hedgehog , Wnt, and Notch are functional in embryonic stem cell development and may be dysregulated in CSCs. Activation of stem cell signaling pathways results in the expression of stemness genes (pluripotency) in CSCs. Examples are Oct4 (octamer-binding transcription factor 4, a homeodomain transcription factor), Nanog (a homeobox protein, another transcription factor), and Sox2 (sex determining region Y-box 2, a transcription factor with a high mobility group domain) commonly found in aggressive, poorly differentiated tumors (Ben-Porath et al. 2008) .
Besides Hedgehog, Wnt, and Notch pathways, additional ones are PI3K and NFκB (Alison et al. 2012) . Phosphoinositide 3-kinase (PI3K) is linked to the mammalian target of rapamycin (mTOR) that relates to cellular energetics. The signaling pathway that leads to the activation of nuclear factor kappa B (NFκB) is important for cytokine expression and the inflammatory response. Activation of these pathways results in the expression of stemness phenotype. The CSCs proliferate and differentiate, and may become resistant to treatments by chemicals and radiation. Molecules targeting these pathways, especially dietary phytochemicals , are discussed with respect to the CSCs.
Niche and Metastasis
Normal stem cells/CSCs reside in specific places. All cells are influenced by their surrounding environment, a local ecosystem. This concept is known as the stem cell microenvironment, or the stem cell niche. Thus, for “normal stem cells and cancer stem cells : the niche matters” (Li and Neaves 2006) ; and “location, location, location: the cancer stem cell niche” (Sneddon and Werb 2007) .
Each resident cell is affected by its niche. The best characterized normal stem cell is the HSC. It behaves differently depending on the niche. In its bone niche, HSC is quiescent in the bone marrow. In its vascular niche, HSC undergoes proliferation and differentiation within the blood vessels. Similarly, the niche concept applies to the CSCs. In addition to the primary tumor site, there are secondary tumor sites after tumor metastasis. Each tumor site consists of a variety of cellular and extracellular components. In addition to cancer cells and CSCs, other cell types include blood and lymphatic endothelial cells, fibroblasts, adipocytes, and immune and inflammatory cells. These cells may secrete molecules that can either promote or inhibit the tumor depending on changes in the microenvironment. The cytokines, chemotactic factors, and extracellular matrix produced by the various cells affect the behavior of cancer cells. Changes always occur within the tumor; the niche is dynamic. For example, there are cancer-associated fibroblasts and tumor-infiltrating macrophages. Separating the macrophage subsets as M1 and M2 with pro- and anti-inflammatory attributes, respectively, M1 protects against and M2 facilitates tumor development. Furthermore, the niche within the tumor is hypoxic. Hypoxia can be a determinant for CSCs, as CSCs may express hypoxia-inducible factors (Fábián et al. 2013) .
Biology of a tumor can only be understood by studying tumor cell heterogeneity and the niche that is constructed during the course of carcinogenesis. Hence, the situation becomes “the bed and the bugs: interactions between the tumor microenvironment and cancer stem cells” (Castaño et al. 2012) . There is interdependence between the CSC and its niche. For example, paracrine signaling between carcinoma cells and mesenchymal stem cells (MSCs) has been reported (Li et al. 2012; Rasanen and Herlyn 2012) . MSCs are recruited to the tumor niche. Carcinoma cells produce the cytokine interleukin-1, which induces the MSCs to produce the lipid molecule prostaglandin E2 (PGE2). PGE2induces changes in carcinoma cells by the activation of the beta-catenin signaling pathway and the formation of CSCs, via a process known as epithelial–mesenchymal transition (EMT) . This example suggests that the niche allows CSC development.
Cells communicate within an epithelial cell layer. This comprises gap junctional intercellular communication (GJIC) , an essential feature for cellular homeostasis (Trosko 2009) . During EMT, epithelial cells undergo a transformation to become more mesenchymal in nature, from the tightly packed, highly differentiated, and immobile cells into the more loosely packed, less differentiated, and mobile cells. A special group of transcription factors is activated, including Snail and Slug (named after fruit fly zinc finger gene regulatory proteins). The adhesive protein E-cadherin (E for epithelial) is downregulated, leading to more motile cells. The absence of E-cadherin also allows for drug resistance, because its elevated expression has been associated with high drug sensitivity. Within the tumor, activation of the EMT program generates a reservoir of CSCs. EMT enhances cell survival by expressing genes that aid cells in avoiding apoptosis (programmed cell death) , anoikis (cell death due to loss of cell adhesion), cellular senescence (the process of limited cell replication), and even host immune response (Tiwaria et al. 2012; Scheel and Weinberg 2012) .
In addition to the contribution to the expression of stem cell phenotypes, EMT facilitates early stages of cancer cell invasion and metastasis. CSCs within the primary tumor leave this location to colonize distant sites; thus, metastasis is the major cause of human cancer deaths. The CSCs may be genetically unstable; multiple CSC clones may be present within the same tumor. The intrinsic genetic alterations keep the cells in the EMT and in the stemness state (Baccelli and Trumpp 2012) . Once they have migrated to secondary sites, the cells may convert back to the epithelial state. Mesenchymal–epithelial transition (MET) can occur, but this reverse process is less well characterized. Nonetheless, there is phenotypic plasticity between the EMT–MET processes (Brabletz 2012) .
Cancer, Cancer Stem Cells, and Mitochondria
Are there interactions among cancer, CSCs, and mitochondria? Almost all mammalian cells have mitochondria as their subcellular organelles, except mature erythrocytes. Mitochondria are essential for cell metabolism. In tumor cells, their mitochondria resist apoptosis-associated permeabilization. Tumor cell mitochondria also contribute to altered cell metabolism by stimulating cell growth and anabolic metabolism. Tumor cell metabolism can become a target for cancer treatment (Kroemer and Pouyssegur 2008) .
An early discovery on tumor metabolism became known as the Warburg effect, after Otto Warburg, who reported the increased glycolysis in leukemic and solid tumor cells. The dominance of this work may have delayed investigations on cancer and mitochondria for some time. However, recent studies on mitochondrial enzymes and metabolism have shown that they play a significant pathogenic role in cancer (Scatena 2012) .
When the cell experiences stress, its mitochondria respond. The rapid mitochondrial fission and fusion depend on GTPases, notably dynamin-related protein-1 (Drp-1) promoting fission and mitofusin-2 (Mfn-2) promoting fusion. The processes lead to elimination of damage (via fission) or compensation of damage (via fusion). Defective mitochondria are destroyed via autophagy (known as mitophagy). If all else fails, the stressed cell undergoes apoptosis (Youle and van der Bliek 2012) .
That mitochondria fission is important for cancer has been demonstrated in lung cancer. Using human lung adenocarcinoma cell line A549 xenografts in nude mice, Mfn-2 gene therapy (with adenovirus vector) leads to smaller tumors as compared to control cancer cells. Similar tumor size reduction is also seen with the pharmacological approach using Mdiv-1, a selective inhibitor of mitochondrial division in yeasts and mammalian cells. Thus, impaired fusion and enhanced fission contribute to the proliferation/apoptosis imbalance in cancer (Rehman et al. 2012) . Targeting cancer cell mitochondria has generated the development of a group of compounds known as mitocans (mitochondrially targeted anticancer drugs). One such example is alpha-tocopheryl succinate, an analogue of vitamin E (Neuzil et al. 2007) .
Thus far, metabolic differences between normal and cancer cell mitochondria have been shown, and the latter may be a therapeutic target of cancer. Is there a relationship to CSCs? Breast CSCs, derived from the human breast cancer cell line MCF7, is susceptible to mitochondrially targeted vitamin E succinate (named MitoVES) in vitro. Additionally, it has been proposed that the CSC marker CD133 may select the stem cells by both evading host immune response and by overcoming stress-induced apoptosis via a mitochondrial connection (Zobalova et al. 2011) . In a study using the CD44+/CD24−/low breast CSCs from MCF7 as xenografts to NOD/SCID mice, the mitochondrial targeting liposome carrying daunorubicin and quinacrine inhibits tumor development (Zhang et al. 2012a) . In summary, CSCs can be targeted by mitochondrial drugs.
Chemoprevention and Phytochemicals
Occupational cancer was first identified by Percivall Pott in 1775: young chimney sweeps later got scrotal cancer because they worked naked to avoid dirtying their clothes. The remedy was to wear clothes during chimney work and to clean and bathe afterwards. Such measures were followed in Holland but not in England, resulting in individuals with less and more scrotal cancers, respectively. Prevention of cancer became a topic of interest; concepts of chemoprophylaxis and chemoprotection began being proposed. In 1976, Michael Sporn introduced the word “chemoprevention” while investigating vitamin A and its analogues, and this became the commonly accepted term (Sporn and Suh 2000; Lippman and Hawk 2009) . Chemoprevention refers to the use of agents to inhibit, reverse, or delay tumorigenesis .
Fruit and vegetable intake has been part of the human diet. Being omnivorous, humans also consume various meats and other foods. Bioactive components from plants are nonnutrient dietary phytochemicals that may modulate gene expression and signal transduction pathways (Manson 2003; Surh 2003; Tan et al. 2011) . Phytochemicals have been isolated and characterized from fruits such as grapes and apples, vegetables such as broccoli and onions, spices such as turmeric, and beverages such as green tea and red wine, as well as numerous other sources. Collectively, these compounds became known as chemopreventive agents. As chemopreventive agents, phytochemicals have been shown to interact with multiple cellular targets (Aggarwal and Shishodia 2006a; Lee et al. 2011) and even with the epigenome (Vanden Berghe 2012) . With respect to chemoprevention , phytochemicals target inflammation (Murakami and Ohigashi 2007; Kim et al. 2009) . This is important because inflammation may initiate cancer, which is the loss of cell cycle control (Meeran and Katiyar 2008) . In addition, the chemopreventive phytochemicals are applicable to cancer therapy, because molecular mechanisms may be common to both chemoprevention and cancer therapy. Phytochemicals may modulate cancer development and even metastasis (Pan and Ho 2008; Gupta et al. 2010) .
Among the diverse phytochemicals implicated in the inhibition of carcinogenesis, one major group comprises polyphenols. These compounds are virtually ubiquitous in plant materials and may occur at very high levels. Mostly synthesized from phenylalanine, plant phenolics serve as structural polymer components and are also responsible for ultraviolet light screening. They have antioxidant activity and act as signaling molecules. The most important role of plant phenolics may be in plant defense against pathogens and herbivore predators. With the discovery of health benefits of plant polyphenols, it has been proposed to optimize the phenolic content of the diet so as to obtain favorable consequences for the general health of the population (Parr and Bolwell 2000) . Within the plant phenolics, one group of importance for chemoprevention is known as the flavonoids (Yao et al. 2011) . Another group has been classified as phytoestrogens (Moutsatsou 2007) . Phytochemical action on CSCs is discussed in the following sections. CSCs are known to be resistant to chemotherapy . Previously we reviewed “overcoming cancer drug resistance by phytochemicals” (Chan and Fong 2009) . Relevant information on phytochemicals discussed earlier has been updated and incorporated in this chapter.
Dietary Phytochemicals Targeting Cancer Stem Cells
Dietary phytochemicals that target CSCs are timely research areas, as seen in the abundance of publications and reviews on the topic. Before covering the major phytochemicals involved, we discuss recent reviews on this subject.
Aptly the very first review we found has the title: “Targeting Cancer Stem Cells with Phytochemicals” (Kawasaki et al. 2008) . Stem cell pathways are presented, as well as a list of clinical trials to evaluate various phytochemicals. A review on CSCs concluded that “dietary phytochemicals are natural products found in our diet and can be used to target cancer stem cells” (Subramaniam et al. 2010) . Another review suggested the tissue-specific stem cells as a target for chemoprevention and concludes that “known chemoprevention agents including sulforaphane , vitamin D3, curcumin , quercetin, genistein , vitamin E, and EGCG may attribute their success at least in part to regulating self-renewal and differentiation of tissue-specific stem cells” (Maund and Cramer 2011) . In a chapter titled “Towards New Anticancer Strategies by Targeting Cancer Stem Cells with Phytochemical Compounds” (Tanveer et al. 2011) , a suggestion was to target Oct4 function in CSCs by phytochemicals . In a review titled “Implications of Cancer Stem Cell Theory for Cancer Chemoprevention by Natural Dietary Compounds” (Li et al. 2011b), stem cell pathways and phytochemicals were discussed; the authors concluded that:
Naturally-occurring dietary compounds are advantageous in several aspects as chemoprevention agents: (1) they are present in commonly consumed food, which is readily available to most people in daily life; (2) they usually have very low or no toxicity, in contrast to most chemotherapy drugs ; (3) many of these compounds have shown potential as an adjunct to chemotherapy drugs in some clinical trials. Although the reports were very limited for dietary compounds to inhibit CSCs, many of them have been shown to be involved in modulation of CSC self-renewal pathways.
The same conclusions were reiterated in their contributed book chapter (Li et al. 2011a).
In addition to dietary phytochemicals , CSCs can be targeted by phytochemical analogues (as reviewed by Dandawate et al. 2013) , other natural products (Efferth 2012) , and even Chinese herbs (Weber et al. 2012) . To end this section, an appropriate quote is from a review titled “Cancer Stem Cells: Potential Target for Bioactive Food Components” (Kim et al. 2012) : “Unquestionably, a diet-induced shift from deregulation to regulation in cancer stem cells could have profound influence on cancer relapses and therefore is of immense societal importance.”
Curcumin
Health Effects
Curcumin is a diferuloylmethane from the Indian spice turmeric. (See Figure 1 for the chemical structure of curcumin.) It is responsible for the yellow color of curry powder. Turmeric is produced from the rhizome of the plant Curcuma longa . In addition to its use as a spice, turmeric has multiple applications in traditional Asian Indian medicine, ranging from treating insect bites to wound healing. Multiple health benefits have been associated with curcumin and turmeric, as summarized in a monograph (Aggarwal et al. 2007) and many reviews (Strimpakos and Sharma 2008; Goel et al. 2008) . Multiple and diverse bioactivities have been attributed to curcumin , including cancer chemopreventive and anti-inflammatory activities (Bisht et al. 2010) . Curcumin acts on many cellular targets (Ravindran et al. 2009) . These include enzymes such as cyclooxygenase, lipoxygenase, and inducible nitric oxide synthase (iNOS) ; transcription factors such as NFκB and activating protein-1 (AP1); cytokines such as tumor necrosis factor (TNF); and many gene products linked with cell survival, proliferation, invasion, and angiogenesis . For example, we found that curcumin inhibits NFκB and TNF in a human monocytic cell line Mono Mac 6. With the endotoxin-induced murine sepsis model, curcumin ingestion by gavage induces bioactivity as shown by downregulating iNOS gene expression in the murine liver (Chan 1995; Chan et al. 1998) . Synergistic effects between curcumin and chemotherapeutic drugs have been demonstrated: we have shown that curcumin enhances the cisplatin-mediated killing of the human ovarian cancer cell lines CAOV3 and SKOV3 in vitro (Chan et al. 2003; Chan and Fong 2007) .

Chemical structure of phytochemicals
We end this section with a note on curcumin and patentability. In March 1995 the USPTO awarded the patent “Use of turmeric in wound healing” (US Patent 5,401,504) to Suman Das and Hari Har Cohly, with the University of Mississippi as assignee. The patent was withdrawn in August 1997, after protest from India’s Council of Scientific and Industrial Research (CSIR) and demonstration of the lack of novelty criterion required for patenting, inasmuch as turmeric has been used for centuries by the Indian population for wound healing (Jayaraman 1997) . Whereas CSCs are drug- and radiation-resistant, curcumin is a chemosensitizer and radiosensitizer for tumors (Goel and Aggarwal 2010) . However, we should also point out the “dark side of curcumin” (Burgos-Morón et al. 2010) , as we note the potential of turmeric consumption in reducing the risk of cancer (Hutchins-Wolfbrandt and Mistry 2011) .
Effects on Cancer Stem Cells
Effects of curcumin on CSCs have been shown. Using SP as selection for CSCs in the rat glioma cell line C6 in vitro, we found that curcumin (5 µM daily for 10 days via medium change) reduced SP, likely via the inhibition of drug transporter (Fong et al. 2010) . Curcumin targets human glioblastoma CSCs in vitro (2 µM) and in vivo, the latter via intracranial implantation of human CSCs with intraperitoneal injections of curcumin (300 mg/kg every 3 days for 5 weeks); the investigators proposed the mechanism as induction of autophagy and promotion of differentiation (Zhuang et al. 2012) . Curcumin targets CSCs of human esophageal squamous carcinoma cell lines in vitro (40–60 µM), showing a decrease in aldehyde dehydrogenase (ALDH) activity as well as CD44 and NFκB expression (Almanaa et al. 2012) .
Curcumin (30–60 µM) inhibits spheroid culture of human esophageal CSCs in vitro via the Notch signaling pathway (Subramaniam et al. 2012) . Both human breast CSCs (from cell lines MCF7 and SUM159) and normal breast stem cells are targeted by curcumin (5–10 µM) together with another phytochemical piperine (from black pepper, 5–10 µM). There was a decrease in spheroid culture numbers as well as a decrease in ALDH activity (Kakarala et al. 2010) . Both normal and CSCs were affected, therefore as a potential phytochemical for cancer therapy, its effect on normal stem cells must be clearly differentiated from CSCs.
Curcumin can complement other drugs. In vitro, cotreatment of curcumin (20 µM) and FOLFOX/FUOX (50 µM fluorouracil and 1.12 µM oxaliplatin) for 5 days inhibited the spheroid cultures of human colon CSCs from the cell lines HCT116 and HT29 (Yu et al. 2009) . Induction of apoptosis in human brain tumor stem cells (commercially available from Celprogen) in vitro has been shown with cotreatment of curcumin (20 µM) and paclitaxel (10 nM) (Hossain et al. 2012) . These results show the feasibility of combining conventional drugs with phytochemicals to target CSCs in clinical cancer.
However, the solubility of curcumin is a concern. To overcome this hurdle, various carriers have been developed. A polymeric nanoparticle formulation of curcumin, nanocurcumin trademark as NanoCurc (5–20 µM), induced apoptosis and reduced the CD133+ stem cells in malignant brain tumors in vitro (Lim et al. 2011) . A hyaluronic acid (HA)-based nanogel-curcumin conjugate, using cholesteryl-HA, had 8 times higher cytotoxicity than curcumin alone towards human pancreatic CSCs expressing CD44 (Wei et al. 2013) . The authors also reported similar effects with nanogel-drug conjugates of two other compounds targeting CSCs, namely etoposide and salinomycin. A novel micelle formulation of curcumin, composed of stearic acid-g-chitosan oligosaccharide (CSO-SA) as a polymeric, biocompatible, and biodegradable drug carrier, inhibited CD44+/CD24+ colorectal CSCs in vitro and in vivo (nude mice xenografts), the latter at a dose of 2 mg/kg daily for 14 days in a 15-day experiment (Wang et al. 2012a) . Another approach for curcumin solubility is to synthesize analogues. Difluorinated-curcumin (CDF) inhibited spheroid cultures of human colon CSCs from the cell line HCT116 (at 2–8 µM) in vitro (Kanwar et al. 2011) . CDF (2 µM) plus FOLFOX/FUOX (100 µM fluorouracil and 2.5 µM oxaliplatin) inhibited the growth of the HT-29 colorectal cancer cell line CSCs in vitro via downregulation of microRNA-21 (miR-21) and promotion of differentiation (Yu et al. 2013) . GO-Y030, a monoketone analogue of curcumin (Sato et al. 2011) , inhibited human colorectal CSCs from the cell lines HCT116 and SW480 in vitro (induced apoptosis at 0.5–2.5 µM) and in vivo (NOD/SCID mice xenografts), the latter at a dose of 50 mg/kg daily for 14 days in a 29-day experiment (Lin et al. 2011) . These results show that human CSCs can be targeted by curcumin and analogues in vivo using mouse models.
Resveratrol
Health Effects
Resveratrol is a stilbene (3,4′,5-tri-hydroxy-trans-stilbene to be exact) from red wine, grapes, peanuts, and pine nuts. (See Fig. 1 for the chemical structure of resveratrol.) Red wine is made from whole grapes (including grape skin) from the fruits of the grape vine plant Vitis venifera and related species. Resveratrol is present in red wine because the molecule is associated with the skin portions of the fruit to fight against fungal infections of the plant. It is an example of the phytoalexins, compounds produced by plants against pathogens. Similar to plant fungal pathogens, we demonstrated it is active against human dermal fungal pathogens: resveratrol may be useful for treating athlete’s foot (Chan 2002) . Health benefits associated with resveratrol are summarized in a monograph (Aggarwal and Shishodia 2006b) and many reviews (Athar et al. 2007; Gescher 2008; Harikumar and Aggarwal 2008; Kundu and Surh 2008; Saiko et al. 2008) . It is “one molecule, many targets” (Pirola and Fröjdo 2008) . Resveratrol’s recent claim to fame involved its action as an activator of sirtuin deacetylases, leading to lifespan extension from organisms as diverse as yeasts, nematodes, fruit flies, and fish (Baur and Sinclair 2006) . Resveratrol’s prior claim to fame involved a phenomenon known as the “French Paradox” (Opie and Lecour 2007) , referring to the observation that the French benefit from a relatively low risk of developing cardiovascular disease despite a diet that is high in saturated fat.
Resveratrol improves general health in mice and seems to delay their ageing parameters (Barger et al. 2008; Pearson et al. 2008) . In this sense, it mimics a phenomenon known as caloric restriction, a method that retards aspects of the ageing process in mammals (Maxmen 2012) . Indeed, it has anti-inflammatory activity (Bisht et al. 2010) and shows health benefits, especially cardioprotection and cancer chemoprevention . With respect to the French Paradox, it is believed resveratrol and alcohol in red wine may be part of the reason. We have shown a synergistic effect between resveratrol and alcohol in the inhibition of iNOS (Chan et al. 2000) . Moderate consumption of red wine may be beneficial to human health.
We end this section with a commercial note. In April 2008, the FDA granted Orphan Drug Status to resveratrol , for the treatment of MELAS syndrome: mitochondrial encephalomyopathy with lactic acidosis and strokelike episodes, caused by mitochondrial DNA mutations. The sponsor was Sirtris Pharmaceuticals, a company in Massachusetts with expertise in resveratrol and its analogues (SRT501 was their resveratrol formulation). Orphan drug designation provides a company with seven years of marketing exclusivity. In late April 2008, Sirtris was purchased by GlaxoSmithKline for $ 720 million. In December 2010, Sirtris announced that the clinical trial of SRT501 was halted but studies with other synthetic resveratrol-mimicking drugs would continue. Currently, the role of resveratrol as an antiageing molecule seems to be in doubt (Wade 2011) . However, we must emphasize that resveratrol possesses cancer chemopreventive properties. The difficulty seems to be deciding on dietary-relevant doses (Scott et al. 2012) .
Effects on Cancer Stem Cells
Effects of resveratrol on CSCs have been shown. These include CSCs from acute myeloid leukemia (AML), brain, breast, head and neck, and pancreatic cancer. Inasmuch as resveratrol has multiple cellular targets, several of the cellular targets in CSCs have been revealed. In AML patients there is an increase in the cytokine interleukin 6 (IL-6) in their plasma. IL-6 induced activation of the Sonic Hedgehog signaling pathway has been shown in the human HL-60 cell line, and resveratrol (25 µM) inhibits this activation and decreases cell viability (Su et al. 2013) . In malignant brain tumor medulloblastoma CSCs that were grown in spheroid cultures, resveratrol (100 µM) inhibited proliferation in vitro and enhanced radiosensitivity (Lu et al. 2009) . In primary central nervous system atypical teratoid/rhabdoid tumors, resveratrol (150–200 µM) induced apoptosis of the isolated CD133+CSCs in vitro and also enhanced radiosensitivity. The in vitro resveratrol-treated and irradiated CD133+CSCs resulted in better survival of SCID mice in a 14-week xenograft experiment (Kao et al. 2009) . In glioblastoma multiforma, comparison of CD133+ and CD133− cells has shown the involvement of signal transducer and activator of transcription 3 (STAT3). Resveratrol acts via inhibiting the STAT3 cytokine signaling pathway. CSCs treated in vitro with resveratrol (200 µM) and irradiation resulted in better survival of SCID mice; this effect of resveratrol can be mimicked with small hairpin (sh) RNA of STAT3 (Yang et al. 2012a) .
Moving from the central nervous system to other organs, breast CSCs have been targeted by resveratrol. Using CD44+/CD24− CSCs isolated from cell lines MCF7, MDA-MB231, and 231LM, resveratrol (50–100 µM) inhibited spheroid cultures by inducing apoptosis. The mechanism involved the action of resveratrol on lipid synthesis, by its downregulation of fatty acid synthase (FAS). Luciferase-expressing CSCs from MDA-MB231 showed smaller-sized tumors in nude mice xenografts with resveratrol administration (22.4 mg/kg via oral gavage or intraperitoneal injection) every 2 days in a 28-day experiment (Pandey et al. 2011) . Similar results have been reported by the same investigators using CSCs isolated from another human breast cancer cell line, MCF10DCIS.com (Pandey et al. 2012) . In human head and neck cancer, resveratrol (25–100 µM) eliminates aldehyde dehydrogenase (ALDH) activity, spheroid formation, and epithelial mesenchymal transition (EMT) in vitro, as well as reduces tumor xenograft volume and weight in vivo in immunodeficient mice (40 mg/kg daily for 20 days) (Hu et al. 2012) . Human pancreatic CD133+/CD44+/CD24+CSCs formed spheroid cultures and expressed stemness genes such as Oct4, Nanog, and Sox2. These activities were inhibited by resveratrol (10–30 µM) in vitro. In a special transgenic mouse model, KRASG12D, the mice were designed to develop pancreatic ductal adenocarcinoma. The increase in size of the pancreas was inhibited by resveratrol (40 mg/kg) administered 5 days a week for 10 months. In addition to inhibiting the stemness genes, alias pluripotency maintaining factors, resveratrol inhibited EMT (Shankar et al. 2011) . In summary, resveratrol aims at different cellular targets of CSCs, depending on the cancer type.
Epigallocatechin Gallate
Health Effects
EGCG is a polyphenol from green tea. (See Figure 1 for the chemical structure of EGCG.) Tea is made from leaves of the plant Camellia sinensis , originally from China. EGCG is the major green tea polyphenol, accounting for 30–40% dry weight of the water-extractable material of green tea. It is converted to dimers and polymers during black tea production. Tea is one of the most popular beverages consumed in the world. Multiple health benefits have been associated with EGCG and tea consumption (Zaveri 2006; Yang et al. 2007; Khan and Mukhtar 2008) . These include cancer chemoprevention , as well as the reduction of atherosclerosis, hypercholesterolemia, Alzheimer’s and other ageing-related diseases. EGCG targets multiple signaling pathways. For example, we reported EGCG inhibition of iNOS both at the gene expression level and at the enzyme activity level (Chan et al. 1997) . EGCG acts synergistically with cisplatin to kill ovarian cancer cells. Even in the cisplatin-resistant cell line C200, enhanced cell death by cisplatin was seen in the presence of EGCG in cell culture (Chan et al. 2006) .
We end this section with a commercial note. In October 2006, the FDA approved an extract of green tea as a prescription drug for topical treatment of genital warts (Gross et al. 2008) . The extract is Polyphenon E (Veregen) ointment (from a German biotech company MediGene and its marketing partner Bradley Pharmaceuticals, Inc.). This was the first prescription botanical (herbal) drug approved by the FDA both under its current regulatory policy for botanical products as well as the FDA’s new Drug Application Process (Blumenthal 2007) .
Effects on Cancer Stem Cells
Effects of EGCG on CSCs have been shown. Actually, early reports used normal stem cells. Spheroid cultures for CSCs followed the initial work with normal neurospheres. In rat neurospheres, EGCG (10–50 µg/ml) and related compounds inhibited neurosphere adhesion and migration (Chen et al. 2003) . Extending to CSC spheroid cultures, epigallocatechin (EGCG without the gallate moiety) and polyphenon 60 (characterized green tea extract, 10 µM) prevented cell shedding from mouse mammary cancer mammospheres in vitro. The mechanisms proposed are decreased ROS production and downregulation of matrix metalloproteinase-9 (Günther et al. 2007) . EGCG (50 µM) inhibited spheroid cultures of human neuroblastoma BE(2)-C cells in vitro. These CSCs exhibit ALDH activity as well as the expression of stemness genes such as Oct4 and Nanog (Nishimura et al. 2012) .
Both EGCG and its analogues act on CSCs. Several chemically synthesized analogues (5–40 µM) target human breast CSCs cultured as mammospheres from the cell line MDA-MB231. The mechanism has been attributed to activation of the AMP-activated protein kinase (AMPK) pathway, because a known activator of this pathway, metformin (5–10 mM), behaves in the same way (Chen et al. 2012c) .
Synergistic effects of EGCG and another phytochemical, quercetin, on CSCs have been investigated. In human pancreatic CSCs, EGCG (20–60 µM) inhibited spheroid cultures, EMT, and the Hedgehog pathway in vitro. This activity was synergistic with quercetin (20 µM), as shown by increased apoptosis (Tang et al. 2012) . In another publication, the same investigators also reported similar effects on human prostate CSCs (Tang et al. 2010) . In summary, EGCG can act on CSCs either alone or in concert with another phytochemical.
Genistein
Health Effects
Genistein is an isoflavone (4,5,7-trihydroxyisoflavone to be exact) from soybeans. (See Fig. 1 for the chemical structure of genistein.) Soybeans are seeds from the legume plant Glycine max , originally from China, although China currently imports soybeans from the United States. It is believed that the isoflavone in soy-rich foods contributes to lower rates of prostate and breast cancers observed in China and Japan as compared to Western countries. Genistein is a molecule with many cellular targets, including inhibition of the NFκB pathway, modulation of the cell cycle, and induction of apoptosis (Banerjee et al. 2008) . In addition, genistein has been classified as a phytoestrogen. However, it is lipid soluble and exhibits low oral bioavailability in pharmacokinetics studies (Yang et al. 2012b) . Furthermore, there is a “dark side” to “the whole soy story” (Daniel 2005) . Soybeans contain proteinase inhibitors that can be “toxic” to digestion. Also, people may develop allergic reactions to soy.
We end this section with an interesting demonstration of the transgenerational effect of genistein . The agouti mouse model , as noted by color change in the fur, is useful as an epigenetic biosensor for nutritional and environmental alterations on the fetal genome (Dolinoy 2008) . Maternal dietary supplementation with genistein (250 mg/kg in the diet) led to DNA hypermethylation in the embryo and the methylation state was maintained until adulthood. Thus, phytochemicals such as genistein modify the epigenome, and the effect starts early in embryogenesis (Dolinoy et al. 2006) .
Effects on Cancer Stem Cells
Effects of genistein on CSCs have been shown. Genistein was reported as a protein tyrosine kinase inhibitor. In an in vitro study on chronic myelogenic leukemia in 1996, the stem and progenitor cells were more sensitive to genistein (200 µM) than the normal progenitor cells (Carlo-Stella et al. 1966a, 1996b) . In human prostate cancer, the CSCs from the cell line PC3 had been targeted by genistein (15 µM) in vitro, which aided in the death of these CSCs by docetaxel. In this work, the CSCs have been named “the Achilles’ heel of cancer” (McCubrey et al. 2011) . Another study on prostate cancer extended its focus to in vivo xenografts, and discovered the cellular target as inhibiting the Hedgehog pathway (Zhang et al. 2012b) . Genistein (30 µM) inhibited spheroid cultures of prostate CSCs from the cell lines 22RV1 and DU145, and inhibited tumor development in vivo in nude mice (twice weekly of genistein and/or docetaxel at 10 mg/kg) in a 28-day experiment. Although the CSCs were resistant to docetaxel, genistein plus docetaxel decreased tumor volume more than genistein alone. Another interesting report involved the feeding of genistein to mice, “at concentrations present in soy protein isolate (250 mg/kg food;” Montales et al. 2012) . Sera from the genistein-fed animals were used to treat human breast CSCs from the cell lines MCF7 and MDA-MB231. When 1–5% of these sera was added to the medium, spheroid formation was inhibited, suggesting “factors found in sera of mice consuming” genistein were active against CSCs . Molecular details of the factors were not characterized. However, this last investigation is interesting with respect to chemoprevention : genistein in the diet leads to the animals having factors in the circulation that inhibit CSCs.
Sulforaphane
Health Effects
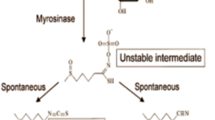
Sulforaphane , alias 1-isothiocyanato-4-methylsulfinylbutane, is an isothiocyanate found in brassicaceous vegetables. (See Fig. 1 for the chemical structure of sulforaphane.) Broccoli, cabbage, cauliflower, Brussel sprouts, and kale are cultivars of the plant known as Brassica oleracea . These common vegetables belong to the plant order Brassicales. An older name is Cruciferae; they are also named crucifers or the cabbage family. The wild cabbage originally came from Europe. Broccoli and related vegetables contain many phytochemicals (Latte et al. 2011) . In addition to sulforaphane , there are other compounds, such as indole-3-carbinol (I3C) and quercetin. Sulforaphane is released from the plant cell wall by activity of the enzyme myrosinase via cooking or chewing. The compound has multiple cellular targets: as an antioxidant, an activator of phase II detoxification enzymes, an anti-inflammatory agent, an inducer of apoptosis, and other roles (James et al. 2012) . Broccoli has been used, as an example, for a “nutritional genomics” approach “in conveying to the consumer” that “the dietary regime being marketed will positively affect their health” (Ferguson and Schlothauer 2012) .
We end this section with a quote on broccoli from a former US president (Dowd 1990) . In March 1990, President George H. W. Bush, in response to queries about a broccoli ban he had imposed upon Air Force One, said: “I do not like broccoli. And I haven’t liked it since I was a little kid and my mother made me eat it. And I’m President of the United States, and I’m not going to eat any more broccoli!” In fact, the president might be a “supertaster.” People who find Brussel sprouts unbearably bitter carry a variant of the bitter taste receptor (T2R38) which results in their enhanced innate immunity (Lee et al. 2012a) .
Effects on Cancer Stem Cells
Effects of sulforaphane on CSCs have been shown. Sulforaphane has been shown to inhibit human breast CSCs from the cell lines MCF7 and SUM159 in vitro and in vivo (Li et al. 2010). Spheroid cultures were inhibited (10–20 µM), as well as ALDH expression. In NOD/SCID mouse xenografts, two successive generations elegantly showed a decrease in tumor volume and an increase in survival (50 mg/kg daily for 14 days) in a 35-day experiment. Sulforaphane inhibited spheroid cultures in human pancreatic CSCs from the cell line MIA-PaCa2 in vitro (20 µM) and in vivo (Rausch et al. 2010) . Sulforaphane inhibits renewal of pancreatic cancer stem cells in vitro (20 µM) by inhibiting the Sonic Hedgehog signaling pathway (Rodova et al. 2012) . In nude mice xenograft experiments, sulforaphane (60 mg/kg) or cotreatment with the multikinase inhibitor sorafenib (3 mg/kg), daily for 3 days in a 5-day experiment, reduced tumor size via inhibition of the NFκB pathway (Rausch et al. 2010) . The same research group also extended their report on pancreatic CSCs to include results on prostate CSCs derived from the cell line DU145 (Kallifatidis et al. 2011) . Sulforaphane was added in vitro to taxol, cisplatin, gemcitabine, flurouracil, or doxorubicin. Targeting was specific for CSCs, because normal cells (primary human fibroblasts and umbilical vein endothelial cells) were not affected. In vivo nude mice xenografts for pancreatic CSCs used sulforaphane in combination with gemcitabine (3 mg/kg consequently for 3 days; treatment started after tumor development was observed, in a 40-day experiment). In a different experiment, the pancreatic CSCs had been pretreated with the sulforaphane–gemcitabine combination before xenograft. Both protocols resulted in decreased tumor volume (Kallifatidis et al. 2011) . In fact, sulforaphane alone could reduce human pancreatic CSC growth in immunodeficient mice, through inhibiton of the Sonic Hedgehog signaling pathway (20 mg/kg once a day for 5 days in a 6-week experiment; Li et al. 2013). In addition to solid tumors, sulforaphane enhancement of drug efficacy has been reported for leukemia. The CD34+ chronic myeloid leukemia CSCs previously resistant to imatinib were killed by a combination of imatinib (0.1–1 µM) and sulforaphane (20–30 µM) in vitro, via activation of the Wnt pathway and ROS induction (Lin et al. 2012) .
Other Dietary Phytochemicals
Many phytochemicals may serve as chemopreventive agents. In some cases, their activities on CSCs have been investigated. In addition to sulforaphane, another important phytochemical found in the brassicaceous vegetables is indole-3-carbinol (I3C). I3C is a derivative of glucobrassicin, a secondary plant metabolite. I3C can be further metabolized into a dimer (3,3”-diindolmethane, DIM). I3C, DIM, and related synthetic analogues are potential agents of chemoprevention and cancer therapy (Weng et al. 2008; Safe et al. 2008) . DIM inhibited spheroid culture formation, for CSCs from human and mouse cancer cell lines (the human non-small–cell lung carcinoma H460 and the murine melanoma B16/F10) in vitro. DIM-treated melanoma CSCs showed smaller tumor volume in vivo (Semov et al. 2012) .
Quercetin is a flavonol found in common fruits and vegetables such as apples, broccoli, and onions. It has antioxidant activity and has been reviewed as safe in vivo, although mutagenic in vitro (Harwood et al. 2007; Boots et al. 2008; Gibellini et al. 2011) . We reported that quercetin sensitized the killing by cisplatin of the human ovarian cancer cell line SKOV3 (Chan et al. 2003) . Quercetin–EGCG’s synergistic activity on human pancreatic CSCs has been discussed (Tang et al. 2011) . In fact, quercetin by itself inhibited spheroid cultures of pancreatic CSCs, and it exhibited a synergistic effect with sulforaphane . In nude mice xenografts, quercetin and sulforaphane decreased tumor size (Zhou et al. 2010) . Quercetin also targets the heat shock protein Hsp27. In human breast CSCs, inhibition of spheroid cultures by the Hsp90 inhibitor geldanamycin was enhanced by quercetin (Lee et al. 2012b) . In human lung CSCs from the cell line A549 as xenografts to NOD/SCID mice, tumor volume was decreased by treatment with quercetin, cisplatin, and gemcitabine (Hsu et al. 2011a) .
Cucurbitacin I , a cell-permeable triterpenoid compound, is found in members of the Cucurbitacea family (cucumber, squash, melons, pumpkins) as well as brassicaceous vegetables. Investigators from Taiwan showed its effect on CD44+/ALDH+ CSCs from head and neck squamous cell carcinoma (Chen et al. 2010) , as well as CD133+ CSCs from non-small–cell lung cancer (Hsu et al. 2011b) , thyroid cancer (Tseng et al. 2012), and pediatric brain cancer medulloblastoma (Chang et al. 2012) . Its cellular target is the STAT3 cytokine pathway.
Lupeol is another triterpene found in fruits and vegetables. A research group in Hong Kong demonstrated its effect on CSCs from liver cancer (Lee et al. 2011) . Its cellular target is the phosphatase and tensin homologue (PTEN)-protein kinase B (Akt)-ABCG2 pathway.
Extending from purified phytochemicals to plant extracts, a blueberry preparation fed to mice has been shown to be effective towards CSCs. Sera from blueberry-fed animals inhibited spheroid cultures of human breast CSCs (Montales et al. 2012) . A pomegranate extract also inhibited mouse mammary CSCs in vitro (Dai et al. 2010)
In summary, individual phytochemicals and plant extracts from fruits, vegetables, spices, and beverages may be harnessed to inhibit CSCs, but their cellular targets may vary depending on the cancer type. A list of probable molecular targets for phytochemicals in CSCs can be found in Table 2.
Other Molecules Targeting Cancer Stem Cells
In addition to dietary phytochemicals , many other molecules target CSCs . We start with compounds isolated from plant species. Parthenolide (PTL), from the feverfew plant (Tanacetum parthenium) , is commonly used in herbal medicine. PTL targets human acute myelogenous leukemia stem cells (Guzman et al. 2005) . As an inhibitor of NFκB, it inhibited human breast CSCs from the cell line MCF7 (Zhou et al. 2008) . Screening in silico for compounds with similar effects on gene expression as PTL has revealed others, including celastrol, found in thunder god vine (Tripterygium wilfordii) and used in Chinese herbal medicine (Hassane et al. 2008) . Another compound, berberine, from plants such as goldenseal (Hydrastis canadensis) and the Chinese goldthread (Coptis chinensis), also used in herbal medicine, targets MCF7 SP cells (Kim et al. 2008) . MCF7 SP cells were also inhibited by the alkaloid oxymatrine, from the Chinese herbal medical plant Sophora flavescens (Zhang et al. 2011) . Eriocalyxin, an analogue of the natural diterpene from the Chinese herbal medical plant Isodon eriocalyx , inhibited human ovarian CSCs via NFκB inhibition (Leizer et al. 2011) . Cyclopamine from corn lily (Veratrum californicum) targets glioblastoma CSCs by inhibiting the Hedgehog pathway (Bar et al. 2007) . However, it should be noted that cyclopamine is a teratogen that causes holoprosencephaly (cyclopia). Silybin, alias silibinin, is a flavonoid compound from the herbal medical plant milk thistle (Silybum marianum). The phytochemical targets colorectal CSCs (Wang et al. 2012b) . Butein is a chalconoid compound from the Chinese lacquer tree (Toxicodendron vernicifluum) with aromatase-inhibiting effects. It targets human breast CSCs (Cioce et al. 2010) . Gossypol is a polyphenol from the cotton plant (genus Gossypium) and at one time had been tested as a male contraceptive. It targets prostate CSCs via activation of the p53 gene regulatory protein (Volate et al. 2010) . From the above studies, it is obvious that many phytochemicals can target CSCs of different cancers.
In addition to plants, molecules targeting CSCs have been found in other sources. 3-O-methylfunicone (OMF) , a secondary metabolite of the soil fungus Penicillium pinophilum, inhibited spheroid cultures of human breast CSCs from the cell line MCF7 (Buommino et al. 2011) . Thus, fungal metabolites may act on CSCs.
Vitamins target CSCs. Retinoic acid, a metabolite of vitamin A, is a molecule essential for growth and development; it also induces cell differentiation. All-trans retinoic acid (ATRA) inhibited human breast CSCs (Ginestier et al. 2009) ; ATRA stealth liposomes prevented relapse of breast cancer arising from breast CSCs (Li et al. 2011). Gemini vitamin D, a vitamin D analogue, targets human breast CSCs (So et al. 2011) . Gamma-tocotrienol, a vitamin E constituent, targets human prostate CSCs (Luk et al. 2011) .
Some drugs may also target CSCs. The telomerase inhibitor imetelstat inhibited both breast and pancreatic CSCs (Joseph et al. 2010) . Statins, including simvastatin and lovastatin, suppressed abnormal human embryonic stem cells and thus may play a role in CSC inhibition (Gauthaman et al. 2009) . Probably the most significant drug reported thus far for targeting CSCs is metformin (Hirsch et al. 2009; Vazquez-Martin et al. 2011; Iliopoulos et al. 2011) . Metformin is currently being used to treat diabetes. There is a definite future in its application to CSCs (Dowling et al. 2012) .
High-throughput screening assays on currently available compounds have been established for embryonic and CSCs. In one human embryonic stem cell screen, retinoic acid and others have been found to modulate Oct4 expression (Desbordes et al. 2008) . In a breast CSC screen, salinomycin has been found to target CSCs. This compound, originally isolated from Streptomyces albus , is an ionophore and has been added to chicken feed as a coccidiostat to control protozoan parasites in poultry (Gupta et al. 2009b) . In another ovarian CSC screen from 1,200 clinically approved drugs, the compound niclosamide has been found to target CSCs (Yo et al. 2012) . Originally developed as a molluscicide (for snails that are intermediate hosts of blood flukes, parasites causing schistosomiasis) and anthelminthic (for cestodes or tapeworms), niclosamide has a proton carrier mode of action and is also active against viruses (such as the one that causes severe acute respiratory syndrome, SARS; Jurgeit et al. 2012) . In a combined normal and CSC screen, the antipsychotic drug thioridazine has been shown to target only CSCs, whereas salinomycin targets both normal and CSCs (Sachlos et al. 2012) . It is expected that more compounds targeting CSCs will be uncovered in future screenings.
A complementary approach is to look for novel sources that target CSCs. Along this line of research, an extract from the marine sponge Crambe crambe has been shown to target pancreatic and prostate CSCs (Ottinger et al. 2012) . Thus, unusual animal and plant species may yield novel molecules targeting CSCs.
Desirable Properties of Compounds Targeting Cancer Stem Cells
Phytochemicals and other compounds that target CSCs have just been reviewed. Here we discuss desirable properties needed for application to cancer therapy. Topics of concern are antagonistic effects, combination therapy , multiple molecular targets, safety, and bioavailability.
Whereas synergistic effects among phytochemicals are welcome in cancer prevention and therapy (Liu 2004; HemaIswarya and Doble 2006) , antagonistic effects are not. We can use curcumin , resveratrol, and EGCG as examples. Curcumin is synergistic with resveratrol in colon cancer inhibition (Majumdar et al. 2009) , but antagonistic with EGCG towards keratinocytes. EGCG enhances keratinocyte differentiation whereas curcumin inhibits its differentiation; curcumin activates apoptosis whereas EGCG does not (Eckert et al. 2006) . Potential antagonistic effects among compounds must be analyzed before combination use.
Despite the potential opposing effects between individual phytochemicals , their combinatorial applications have been promoted. It has been suggested that “Specific combinations of phytochemicals may be far more effective in protecting against cancer than isolated compounds” because “combinations of dietary chemopreventive agents can sometimes result in significant activities at concentrations where any single agent is inactive” (De Kok et al. 2008) . Thus, combinatorial use of phytochemicals is an attractive approach in targeting CSCs.
Each CSC has multiple molecular targets. Dietary phytochemicals such as curcumin , resveratrol, and EGCG tend to aim at a multitude of cellular targets. Perhaps it is because of these characteristics that definitive mechanisms of action are not available despite decades of research (Francy-Guilford and Pezzuto 2008) . This multitarget nature of phytochemicals may be advantageous in targeting CSCs because the multifaceted mode of action may hinder the cell’s ability to develop resistance to the phytochemicals.
How safe are the dietary phytochemicals? Because they can interact with drug transporters and drug metabolizing enzymes (known as phase I and phase II enzymes), the potential toxicity and safety of phytochemicals are major concerns. As an example, they have antioxidant capacities against ROS. However, under special conditions they also exhibit pro-oxidant capacity. Whereas ROS is harmful in general, the removal of too much ROS interferes with body functions, as seen in the warning presented by scientists to the food industry (Finley et al. 2011) . With an emphasis on their putative cancer chemopreventive uses, the safety of a group of phytochemicals has been reviewed (Verschoyle et al. 2007) . For curcumin, given to cancer patients at 3,600 mg/day for 4 months or 800 mg/day for 3 months, only minor adverse effects were seen. For EGCG, daily oral doses for 4 weeks at 800 mg/day in 40 volunteers caused only minor adverse effects. For resveratrol , a single oral dose at 5 g in 10 volunteers caused only minor adverse effects (Boocock et al. 2007) . However, “High consumption of dietary phytochemicals should be considered with caution taking into account their dosage regimes, toxicity, metabolic conversion, transport mechanisms, tissue availability, synergistic interaction with drugs, and interferences with key enzymes, receptors, metabolic pathways, and normal GI microflora” (Priyadarsini and Nagini 2012) . More extensive toxicological testing is recommended, if phytochemicals are to be taken as cancer chemopreventive agents on a long-term basis by healthy individuals.
How bioavailable are the dietary phytochemicals? Bioavailability may differ, depending on conditions varying from pure compounds and plant extracts, to mixtures with other food components (EGCG vs. green tea extract, genistein vs. soymilk or tofu, quercetin vs. apple cider or apples; Manach et al. 2005) . For phytochemicals such as plant polyphenols, the effects in vivo, although significant, are more limited than those observed in vitro (Williamson and Manach 2005) . However, it is possible to design appropriate interactions that affect the bioavailability of phytochemicals in vivo (Scholz and Williamson 2007) , such as enhancement of curcumin efficacy with piperine, as previously mentioned. We have used oral gavage plus starvation (an empty stomach) to enhance curcumin uptake in mice (Chan et al. 1998; Adapala and Chan 2008) so as to demonstrate its in vivo inhibition of the inflammatory iNOS in the murine liver (beneficial outcome) and the exacerbation of murine visceral leishmaniasis, a protozoan parasitic disease (detrimental outcome). The last report recommends caution with regard to the potential clinical use of curcumin : it may re-activate latent infections.
A second look at bioavailability relates to the concept of hormesis. The effect of curcumin has been described as an example of xeno-hormesis (Salvioli et al. 2007) . The concept of hormesis suggests that the fundamental nature of the dose–response curve to be neither linear nor threshold, but rather U-shaped or J-shaped. This shows a low dose stimulatory response known as the hormetic effect, representing overcompensation in response to disruptions in homeostasis (Calabrese and Baldwin 2001) . For phytochemicals among an estimated 10,000 secondary products (natural pesticides), it has been proposed that human ancestors evolved a generalized defense mechanism against low levels of phytochemicals to enable their consumption of many different plant species containing variable levels of natural pesticides (carcinogens) without subsequent ill health. Traces of phytochemicals found in fruits and vegetables may potentiate the immune system and help to protect against cancer (Trewavas and Stewart 2003) . A biphasic dose response is observed. At high concentrations, phytochemicals can be toxic, whereas subtoxic doses may induce adaptive stress responses. There is the activation of signaling pathways that results in increased expression of genes encoding cytoprotective proteins. It is suggested that hormetic mechanisms of action may underlie many of the health benefits of phytochemicals (Mattson 2008) . These benefits would include their action against CSCs.
Implementation of the Phytochemical Approach Towards Chemoprevention
Among all the compounds undergoing clinical trials , the success rate of anticancer drugs is less than 5%, the lowest as compared to other drugs for cardiovascular, central nervous system, and infectious diseases; and it takes $ 2 billion to bring a drug to market (Bhattacharjee 2012). Examples of anticancer drugs from plants are taxol and vinblastine. Plants are a rich source of drugs. “Only a small fraction of the immense diversity of plant metabolism has been explored for the production of new medicines and other products important to human well-being” (De Luca et al. 2012) . Therefore, new phytochemicals targeting CSCs are expected in the future.
In the meantime, currently available phytochemicals need more definitive studies, because an evidence-based approach is favored (Higdon 2007) . “Facts and fiction” of each phytochemical must be differentiated (Espín et al. 2007) . A phytochemical may act in concert with conventional drugs, as visualized by the following metaphor. When used in low concentrations, the phytochemical causes multiple wounds without killing the CSC. Nevertheless, the treatment increases the CSC’s vulnerability to the magic bullet represented by the chemotherapeutic drug (Russo 2007) . In this sense, dietary phytochemicals may serve as adjuvant anticancer agents with practically minimal or no side effects.
There is an urgent need to bridge the gap between apparent in vitro efficacy and clinical use of dietary phytochemicals (Manson et al. 2007) . Biomarkers and decisions on dosages for clinical trials still have to be worked out (Scott et al. 2009) . As an example, transcription analyses (microarrays of gene expression) of peripheral blood mononuclear cells (PMBCs) from dietary intervention studies “have not resulted yet in clear confirmation of candidate genes related to disease risk” (De Mello et al. 2012) .
What is the current status of dietary phytochemicals for cancer chemoprevention? The focus lies in the identification of molecular mechanisms and cellular targets. The “concept of combination chemoprevention by multiple agents or by the consumption of ‘whole foods’ has become an increasingly attractive area of study” (Mehta et al. 2010) .
Concerning “whole foods” consumption, the current US dietary guidelines have been illustrated as MyPlate, with advice to cover half the plate with fruits and vegetables (Willett and Ludwig 2011) . From Europe, there is Mediterranean Diet Pyramid Today, with a plant-based core of the dietary pattern for consuming fruits and vegetables everyday (Bach-Faig et al. 2011) .
How effective are fruits and vegetables for cancer chemoprevention? The early case-control studies tout the reduction in cancer risk, but the more recent prospective cohort studies show only a weak association. For example, a pooled analysis of 14 cohort studies of pancreatic cancer concludes that “fruit and vegetable intake during adulthood is not associated with a reduced pancreatic cancer risk” (Koushik et al. 2012) . Therefore, this is “turmoil in the produce section” but: “A very weak or undetectable association between fruits and vegetables and risk of cancer does not exclude the possibility that one or a small group of fruits or vegetables, or a specific substance in some of these foods, has an important protective effect” (Willett 2010) . This latter point has been well proven by the catalogue of dietary phytochemicals that can target CSCs covered in this review. Furthermore, “although the evidence for benefits of fruits and vegetables against cancer was waning, data supporting benefits for cardiovascular disease were accumulating” (Willett 2010) .
Perspectives
In September 2011, the United Nations General Assembly convened a High Level Meeting on Non-communicable Diseases (NCDs) because the four main NCDs —cardiovascular disease, cancer, chronic lung diseases, and diabetes—kill three in five people worldwide, and cause great socioeconomic harm within all countries, particularly developing nations. In September 2012, Science magazine published a special section including a cover photo on the topic of disease prevention (Ash et al. 2012) . One contributed article titled: “Can Noncommunicable Diseases Be Prevented?” suggested “effective approaches for large-scale NCD prevention include,” among these, “increasing the consumption of fresh fruits and vegetables” (Ezzati and Riboli 2012) .
When is the best time to start eating fruits and vegetables? First Lady Michelle Obama started a campaign against childhood obesity (Willett and Ludwig 2011) . It has been suggested that “research should focus more sharply on specific fruits and vegetables and their constituents and on earlier periods of life” (Willett 2010) . We have already mentioned genistein’s effect on fetal DNA methylation. Thus, here comes “the maternal womb: a novel target for cancer prevention” (Simmen and Simmen 2011) . Dietary interventions prior to and during pregnancy may confer cancer chemoprevention . The first sentence in a discussion on the “myth or reality” aspects of dietary molecules begins with the 400 BC statement of Hippocrates: “Let thy food be thy medicine and thy medicine be thy food” (Neergheen-Bhujun et al. 2012) .
We end this chapter with a comment on cell differentiation and the CSC. In October 2012 the Nobel Prize in Physiology or Medicine was announced for the discovery of cell reprogramming (Abbott 2012) . In 1962 John Gurdon published his work on the plasticity of a differentiated nucleus from an amphibian: the nucleus from a tadpole’s intestinal cell replaced the egg nucleus and generated a frog. In 2006 Shinya Yamanaka published his work on the reprogramming of a differentiated cell type from a mouse: the mouse fibroblast became an embryonic stem cell by adding just four genes. The latter work is transferable to humans and the induced pluripotent stem (iPS) cells will be important for future clinical applications. The differentiated state of a cell is dynamic. A CSC plays its essential role in maintaining the particular tumor. If dietary phytochemicals , or specific fruits and vegetables, are available to destroy this CSC, or reverse its stemness characteristics, or induce its cell differentiation, there will be chemoprevention of cancer.
References
Abbott A (2012) Cell rewind wins medicine nobel. Nature 490:151–152
Adapala N, Chan MM (2008) Long-term use of an anti-inflammatory, curcumin, suppressed type 1 immunity and exacerbated visceral leishmaniasis in a chronic experimental model. Lab Invest 88:1329–1339
Aggarwal BB, Shishodia S (2006a) Molecular targets of dietary agents for prevention and therapy of cancer. Biochem Pharmacol 71:1397–1421
Aggarwal BB, Shishodia S (eds) (2006b) Resveratrol in health and disease. Taylor & Francis, Boca Raton
Aggarwal BB, Surh YJ, Shishodia S (eds) (2007) The molecular targets and therapeutic uses of curcumin in health and disease. Springer, New York
Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, Clarke MF (2003) Prospective identification of tumorigenic breast cancer cells. Proc Natl Acad Sci USA 100:3983–3988
Alison MR, Lim SM, Nicholson LJ (2011) Cancer stem cells: problems for therapy? J Pathol 223:147–161
Alison MR, Lin WR, Lim SM, Nicholson LJ (2012) Cancer stem cells: in the line of fire. Cancer Treat Rev 38:589–598
Allan AL (ed) (2011) Cancer stem cells in solid tumors. Springer, New York
Almanaa TN, Geusz ME, Jamasbi RJ (2012) Effects of curcumin on stem-like cells in human esophageal squamous carcinoma cell lines. BMC Complement Altern Med 12(1):195
Ash C, Kiberstis P, Marshall E, Travis J (2012) It takes more than an apple a day. Science 337:1467
Athar M, Back JH, Tang X, Kim KH, Kopelovich L, Bickers DR, Kim AL (2007) Resveratrol: a review of preclinical studies for human cancer prevention. Toxicol Appl Pharmacol 224:274–283
Baccelli I, Trumpp A (2012) The evolving concept of cancer and metastasis stem cells. J Cell Biol 198:281–293
Bach-Faig A, Berry EM, Lairon D, Reguant J, Trichopoulou A, Dernini S, Medina FX, Battino M, Belahsen R, Miranda G, Serra-Majem L, Mediterranean Diet Foundation Expert Group (2011) Mediterranean diet pyramid today: science and cultural updates. Public Health Nutr 14:2274–2284
Banerjee S, Li Y, Wang Z, Sarkar FH (2008) Multi-targeted therapy of cancer by genistein. Cancer Lett 269:226–242
Bapat SA (ed) (2009) Cancer Stem Cells: identification and Targets. John Wiley, New York
Bapat SA (2010) Human ovarian cancer stem cells. Reproduction 140:33–41
Bapat SA, Mali AM, Koppikar CB, Kurrey NK (2005) Stem and progenitor-like cells contribute to the aggressive behavior of human epithelial ovarian cancer. Cancer Res 65:3025–3029
Bar EE, Chaudhry A, Lin A, Fan X, Schreck K, Matsui W, Piccirillo S, Vescovi AL, DiMeco F, Olivi A, Eberhart CG (2007) Cyclopamine-mediated hedgehog pathway inhibition depletes stem-like cancer cells in glioblastoma. Stem Cells 25:2524–2533
Barger JL, Kayo T, Vann JM, Arias EB, Wang J, Hacker TA, Wang Y, Raederstorff D, Morrow JD, Leeuwenburgh C, Allison DB, Saupe KW, Cartee GD, Weindruch R, Prolla TA (2008) A low dose of dietary resveratrol partially mimics caloric restriction and retards aging parameters in mice. PloS ONE 3:e2264
Baur JA, Sinclair DA (2006) Therapeutic potential of resveratrol: the in vivo evidence. Nat Rev Drug Discov 5:493–506
Ben-Porath I, Thomson MW, Carey VJ, Ge R, Bell GW, Regev A, Weinberg RA (2008) An embryonic stem cell-like gene expression signature in poorly differentiated aggressive human tumors. Nat Genet 40:499–507
Bhattacharjee Y (2012) Pharma firms push for sharing of cancer trial data. Science 338:29
Bisht K, Wagner KH, Bulmer AC (2010) Curcumin, resveratrol and flavonoids as anti-inflammatory, cyto- and DNA-protective dietary compounds. Toxicology 278:88–100
Blackburn JS, Liu S, Langenau DM (2011) Quantifying the frequency of tumor-propagating cells using limiting dilution cell transplantation in syngeneic zebrafish. J Vis Exp 14(53):e2790.
Blumenthal M (2007) FDA approves special green tea extract as a new topical drug for genital warts: expert says development marks the birth of a “new industry.” HerbalGram 74:62–63
Bonnet D, Dick JE (1997) Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell. Nat Med 3:730–737
Boocock DJ, Faust GE, Patel KR, Schinas AM, Brown VA, Ducharme MP, Booth TD, Crowell JA, Perloff M, Gescher AJ, Steward WP, Brenner DE (2007) Phase I dose escalation pharmacokinetic study in healthy volunteers of resveratrol, a potential cancer chemopreventive agent. Cancer Epidemiol Biomarkers Prev 16:1246–1252
Boots AW, Haenen GR, Bast A (2008) Health effects of quercetin: from antioxidant to nutraceutical. Eur J Pharmacol 585:325–337
Brabletz T (2012) To differentiate or not—routes towards metastasis. Nat Rev Cancer 12:425–436
Buommino E, Tirino V, De Filippis A, Silvestri F, Nicoletti R, Ciavatta ML, Pirozzi G, Tufano MA (2011) 3-O-methylfunicone, from Penicillium pinophilium, is a selective inhibitor of breast cancer stem cells. Cell Prolif 44:401–409
Burgos-Morón E, Calderón-Montaño JM, Salvador J, Robles A, López-Lázaro M (2010) The dark side of curcumin. Int J Cancer 126:1771–1775
Calabrese EJ, Baldwin LA (2001) Hormesis: u-shaped dose responses and their centrality in toxicology. Trends Pharmacol Sci 22:285–291
Carlo-Stella C, Regazzi E, Garau D, Mangoni L, Rizzo MT, Bonati A, Dotti G, Almici C, Rizzoli V (1996) Effect of the protein tyrosine kinase inhibitor genistein on normal and leukaemic haemopoietic progenitor cells. Br J Haematol 93:551–557
Carlo-Stella C, Dotti G, Mangoni L, Regazzi E, Garau D, Bonati A, Almici C, Sammarelli G, Savoldo B, Rizzo MT, Rizzoli V (1996) Selection of myeloid progenitors lacking BCR/ABL mRNA in chronic myelogenous leukemia patients after in vitro treatment with the tyrosine kinase inhibitor genistein. Blood 88:3091–3100
Castaño Z, Fillmore CM, Kim CF, McAllister SS (2012) The bed and the bugs: interactions between the tumor microenvironment and cancer stem cells. Semin Cancer Biol 22:462–470
Chan MM (1995) Inhibition of tumor necrosis factor by curcumin, a phytochemical. Biochem Pharmacol 49:1551–1556
Chan MM (2002) Antimicrobial effect of resveratrol on dermatophytes and bacterial pathogens of the skin. Biochem Pharmacol 63:99–104
Chan MM, Fong D (2007) Overcoming ovarian cancer drug resistance with phytochemicals and other compounds. In: Torres LS (ed) Cancer drug resistance research perspectives. Hauppauge: Nova Science Publishers, pp 173–201
Chan MM, Fong D (2009) Overcoming drug resistance by phytochemicals. In: Mehta K, Siddik ZH (eds) Drug resistance in cancer cells. Springer, New York, pp 315–342
Chan MM, Fong D, Ho CT, Huang HI (1997) Inhibition of inducible nitric oxide synthase gene expression and enzyme activity by epigallocatechin gallate, a natural product from green tea. Biochem Pharmacol 54:1281–1286
Chan MM, Huang HI, Fenton MR, Fong D (1998) In vivo inhibition of nitric oxide synthase gene expression by curcumin, a cancer preventive natural product with anti-inflammatory properties. Biochem Pharmacol 55:1955–1962
Chan MM, Soprano KJ, Weinstein K, Fong D (2006) Epigallocatechin-3-gallate deliver hydrogen peroxide to induce death of ovarian cancer cells and enhances their cisplatin susceptibility. J Cell Physiol 207:38996
Chan MM, Fong D, Soprano KJ, Holmes WF, Heverling H (2003) Inhibition of growth and sensitization to cisplatin-mediated killing of ovarian cancer cells by polyphenolic chemopreventive agents. J Cell Physiol 194:63–70
Chan MM, Mattiacci JA, Hwang HS, Shah A, Fong D (2000) Synergy between ethanol and grape polyphenols, quercetin, and resveratrol, in the inhibition of the inducible nitric oxide synthase pathway. Biochem Pharmacol 60:1539–1548
Chang CJ, Chiang CH, Song WS, Tsai SK, Woung LC, Chang CH, Jeng SY, Tsai CY, Hsu CC, Lee HF, Huang CS, Yung MC, Liu JH, Lu KH (2012) Inhibition of phosphorylated STAT3 by cucurbitacin I enhances chemoradiosensitivity in medulloblastoma-derived cancer stem cells. Childs Nerv Syst 28:363–73
Chen CN, Liang CM, Lai JR, Tsai YJ, Tsay JS, Lin JK (2003) Capillary electrophoretic determination of theanine, caffeine, and catechins in fresh tea leaves and oolong tea and their effects on rat neurosphere adhesion and migration. J Agric Food Chem 51:7495–7503
Chen D, Pamu S, Cui Q, Chan TH, Dou QP (2012a) Novel epigallocatechin gallate (EGCG) analogs activate AMP-activated protein kinase pathway and target cancer stem cells. Bioorg Med Chem 20:3031–3037
Chen J, Li Y, Yu TS, McKay RM, Burns DK, Kernie SG, Parada LF (2012b) A restricted cell population propagates glioblastoma growth after chemotherapy. Nature 488:522–526
Chen SF, Chang YC, Nieh S, Liu CL, Yang CY, Lin YS (2012c) Nonadhesive culture system as a model of rapid sphere formation with cancer stem cell properties. PLoS ONE 7:e31864
Chen YW, Chen KH, Huang PI, Chen YC, Chiou GY, Lo WL, Tseng LM, Hsu HS, Chang KW, Chiou SH (2010) Cucubitacin I suppressed stem-like property and enhanced radiation-induced apoptosis in head and neck squamos carcinoma—derived CD44(+)ALDH1(+)cells. Mol Cancer Ther 9:2879–2892
Cheng L, Ramesh AV, Flesken-Nikitin A, Choi J, Nikitin AY (2010) Mouse models for cancer stem cell research. Toxicol Pathol 38:62–71
Cioce M, Gherardi S, Viglietto G, Strano S, Blandino G, Muti P, Ciliberto G (2010) Mammosphere-forming cells from breast cancer cell lines as a tool for the identification of CSC-like- and early progenitor-targeting drugs. Cell Cycle 9:2878–2887
Clarke MF, Dick JE, Dirks PB, Eaves CJ, Jamieson CH, Jones DL, Visvader J, Weissman IL, Wahl GM (2006) Cancer stem cells—perspectives on current status and future directions: AACR Workshop on cancer stem cells. Cancer Res 66:9339–9344
Clevers H (2011) The cancer stem cell: premises, promises, and challenges. Nat Med 17:313–319
Crea F, Duhagon MA, Farrar WL, Danesi R (2011) Pharmacogenomics and cancer stem cells: a changing landscape? Trends Pharmacol Sci 32:487–494
Creighton CJ, Li X, Landis M, Dixon JM, Neumeister VM, Sjolund A, Rimm DL, Wong H, Rodriguez A, Herschkowitz JI, Fan C, Zhang X, He X, Pavlick A, Gutierrez MC, Renshaw L, Larionov AA, Faratian D, Hilsenbeck SG, Perou CM, Lewis MT, Rosen JM, Chang JC (2009) Residual breast cancers after conventional therapy display mesenchymal as well as tumor-initiating features. Proc Natl Acad Sci USA 106:13820–13825
Dai Z, Nair V, Khan M, Ciolino HP (2010) Pomegranate extract inhibits the proliferation and viability of MMTV-Wnt-1 mouse mammary cancer stem cells in vitro. Oncol Rep 24:1087–1091
Dandawate P, Padhye S, Ahmad A, Sarkar FH (2013) Novel strategies targeting cancer stem cells through phytochemicals and their analogs. Drug Delivery Transl Res 3(2):165–182
Daniel KT (2005) The whole soy story: the dark side of America’s favorite health food. New Trends Publishing, Washington D.C.
De Kok TM, Breda SG van, Manson MM (2008) Mechanisms of combined action of different chemopreventive dietary compounds: a review. Eur J Nutr 47(Suppl 2):51–59
De Mello VD, Kolehmanien M, Schwab U, Pulkkinen L, Uusitupa M (2012) Gene expression of peripheral blood mononuclear cells as a tool in dietary intervention studies: what do we know so far? Mol Nutr Food Res 56:1160–1172
De Luca V, Salim V, Atsumi SM, Yu F (2012) Mining the biodiversity of plants: a revolution in the making. Science 336:1658–1661
Desbordes SC, Placantonakis DG, Ciro A, Socci ND, Lee G, Djaballah H, Studer L (2008) High-throughput screening assay for the identification of compounds regulating self-renewal and differentiation in human embryonic stem cells. Cell Stem Cell 2:602–612
Diehn M, Cho RW, Lobo NA, Kalisky T, Dorie MJ, Kulp AN, Qian D, Lam JS, Ailles LE, Wong M, Joshua B, Kaplan MJ, Wapnir I, Dirbas FM, Somlo G, Garberoglio C, Paz B, Shen J, Lau SK, Quake SR, Brown JM, Weissman IL, Clarke MF (2009) Association of reactive oxygen species levels and radioresistance in cancer stem cells. Nature 458:780–783
Dolinoy DC (2008) The agouti mouse model: an epigenetic biosensor for nutritional and environmental alterations on the fetal epigenome. Nutr Rev 66(Suppl 1):S7–S11
Dolinoy DC, Weidman JR, Waterland RA, Jirtle RL (2006) Maternal genistein alters coat color and protects Avy mouse offspring from obesity by modifying the fetal epigenome. Environ Health Perspect 114:567–572
Donnenberg VS, Donnenberg AD (2005) Multiple drug resistance in cancer revisited: the cancer stem cell hypothesis. J Clin Pharmacol 45:872–877
Dowd M (1990) ‘I’m President,’ so no more broccoli! The New York Times. 23 March
Dowling RJ, Niraula S, Stambolic V, Goodwin PJ (2012) Metformin in cancer: translational challenges. J Mol Endocrinol 48:R31–R43
Drewa T, Czerwinski M, Olkowska J (2011) Stem cells within established cancer stem cell lines: an impact on in vitro experiments. Stem Cell Studies Volume 1:e7
Driessens G, Beck B, Caauwe A, Simons BD, Blanpain C (2012) Defining the mode of tumour growth by clonal analysis. Nature 488:527–530
Eckert RL, Crish JF, Efimova T, Balasubramanian S (2006) Opposing action of curcumin and green tea polyphenol in human keratinocytes. Mol Nutr Food Res 50:123–129
Efferth T (2012) Stem cells, cancer stem-like cells, and natural products. Planta Med 78:935–942
Espín JC, García-Conesa MT, Tomás-Barberán FA (2007) Nutraceuticals: facts and fiction. Phytochemistry 68:2986–3008
Ezzati M, Riboli E (2012) Can noncommunicable diseases be prevented? Lessons from studies of populations and individuals. Science 337:1482–1487
Fábián Á, Vereb G, SzöllÓ§si J (2013) The hitchhikers guide to cancer stem cell theory: markers, pathways and therapy. Cytometry Part A 83(1):62–71
Farrar WL (ed) (2010) Cancer stem cells. Cambridge University Press, New York
Ferguson LR, Schlothauer RC (2012) The potential role of nutritional genomics tools in validating high health foods for cancer control: broccoli as example. Mol Nutr Food Res 56:126–146
Finley JW, Kong AN, Hintze KJ, Jeffery EH, Ji LL, Lei XG (2011) Antioxidants in foods: state of the science important to the food industry. J Agric Food Chem 59:6837–6846
Fong D, Chan MM (2012) Targeting cancer stem cells with phytochemicals: inhibition of the rat C6 glioma side population by curcumin. In: Hayat MA (ed) Stem cells and cancer stem cells. Springer, New York, pp 61–68
Fong D, Yeh A, Naftalovich R, Choi TH, Chan MM (2010) Curcumin inhibits the side population (SP) phenotype of the rat C6 glioma cell line: towards targeting of cancer stem cells with phytochemicals. Cancer Lett 293:65–72
Francy-Guilford J, Pezzuto JM (2008) Mechanisms of cancer chemopreventive agents: a perspective. Planta Med 74:1644–1650
Gao MQ, Choi YP, Kang S, Youn JH, Cho NH (2010) CD24 + cells from hierarchically organized ovarian cancer are enriched in cancer stem cells. Oncogene 29:2672–2680
Garvalov BK, Acker T (2011) Cancer stem cells: a new framework for the design of tumor therapies. J Mol Med 89:95–107
Gauthaman K, Manasi N, Bongso A (2009) Statins inhibit the growth of variant human embryonic stem cells and cancer cells in vitro but not normal human embryonic stem cells. Br J Pharmacol 157:962–973
Gescher AJ (2008) Resveratrol from red grapes—pedestrian polyphenol or useful anticancer agent? Planta Med 74:1651–1655
Ghiaur G, Gerber J, Jones RJ (2012) Concise review: cancer stem cells and minimal residual disease. Stem Cells 30:89–93
Gibellini L, Pinti M, Nasi M, Montagna JP, De Biasi S, Roat E, Bertoncelli L, Cooper EL, Cossarizza A (2011) Quercetin and cancer chemoprevention. Evid Based Complement Alternat Med 2011:591356
Gilbertson RJ (2012) Meeting the parents. Nature 488:462–463
Ginestier C, Wicinski J, Cervera N, Monville F, Finetti P, Bertucci F, Wicha MS, Birnbaum D, Charafe-Jauffret E (2009) Retinoid signaling regulates breast cancer stem cell differentiation. Cell Cycle 8:3297–3302
Ginestier C, Hur MH, Charafe-Jauffret E, Monville F, Dutcher J, Brown M, Jacquemier J, Viens P, Kleer CG, Liu S, Schott A, Hayes D, Birnbaum D, Wicha MS, Dontu G (2007) ALDH1 is a marker of normal and malignant human mammary stem cells and a predictor of poor clinical outcome. Cancer Stem Cell 1:555–567
Goel A, Aggarwal BB (2010) Curcumin, the golden spice from Indian saffron, is a chemosensitizer and radiosensitizer for tumors and chemoprotector and radioprotector for normal organs. Nutr Cancer 62:919–930
Goel A, Kunnumakkara AB, Aggarwal BB (2008) Curcumin as "Curecumin": from kitchen to clinic. Biochem Pharamcol 75:787–809
Graham TA (2012) Stemming tumour evolution. Nature 488:463
Greaves M, Maley CC (2012) Clonal evolution in cancer. Nature 481:306–313
Gross G, Meyer K-G, Pres H, Thielert C, Tawfik H, Mescheder A (2008) Green tea extract Polyphenon E treats external genital warts. HerbalGram 21:1404–1412
Günther S, Ruhe C, Derikito MG, Böse G, Sauer H, Wartenberg M (2007) Polyphenols prevent cell shedding from mouse mammary cancer spheroids and inhibit cancer cell invasion in confrontation cultures derived from embryonic stem cells. Cancer Lett 250:25–35
Gupta PB, Chaffer CL, Weinberg RA (2009a) Cancer stem cells: mirage or reality? Nat Med 15:1010–1012
Gupta PB, Onder TT, Jiang G, Tao K, Kuperwasser C, Weinberg RA, Lander ES (2009b) Identification of selective inhibitors of cancer stem cells by high-throughput screening. Cell 138:645–659
Gupta SC, Kim JH, Prasad S, Aggarwal BB (2010) Regulation of survival, proliferation, invasion, angiogenesis, and metastasis of tumor cells through modulation of inflammatory pathways by nutraceuticals. Cancer Metastasis Rev 29:405–434
Guzman ML, Rossi RM, Karnischky L, Li X, Peterson DR, Howard DS, Jordan CT (2005) The sesquiterpene lactone parthenolide induces apoptosis of human acute myelogenous leukemia stem and progenitor cells. Blood 105:4163–4169
Hanahan D, Weinberg RA (2000) The hallmarks of cancer. Cell 100:57–70
Hanahan D, Weinberg RA (2011) Hallmarks of cancer: the next generation. Cell 144:646–674
Harikumar KB, Aggarwal BB (2008) Resveratrol: a multitargeted agent for age-associated chronic diseases. Cell Cycle 7:1020–1035
Harwood M, Danielewska-Nikiel B, Borzelleca JF, Flamm GW, Williams GM, Lines TC (2007) A critical review of the data related to the safety of quercetin and lack of evidence of in vivo toxicity, including lack of genotoxic/carcinogenic properties. Food Chem Toxicol 45:2179–2205
Hassane DC, Guzman ML, Corbett C, Li X, Abboud R, Young F, Liesveld JL, Carrol M, Jordan CT (2008) Discovery of agents that eradicate leukemia stem cells using an in silico screen of public gene expression data. Blood 111:5654–5662
Hayat MA (ed) (2012) Stem cells and cancer stem cells, therapeutic application in disease and injury, Vol 1–7. Springer, New York
HemaIswarya S, Doble M (2006) Potential synergism of natural products in the treatment of cancer. Phytother. Res 20:239–249
Higdon J (2007) An evidence-based approach to dietary phytochemicals. Thieme, New York
Hirsch HA, Iliopoulos D, Tsichlis PN, Struhl K (2009) Metformin selectively targets cancer stem cells, and acts together with chemotherapy to block tumor growth and prolong remission. Cancer Res 69:7507–7511
Hossain M, Banik NL, Ray SK (2012) Synergistic anti-cancer mechanisms of curcumin and paclitaxel for growth inhibition of human brain tumor stem cells and LN18 and U138MG cells. Neurochem Int 61(7):1102–1113
Hsu HS, Lin JH, Huang WC, Hsu TW, Su K, Chiou SH, Tsai YT, Hung SC (2011a) Chemoresistance of lung cancer stemlike cells depends on activation of Hsp27. Cancer 117:1516–1528
Hsu HS, Huang PI, Chang YL, Tzao C, Chen YW, Shih HC, Hung SC, Chen YC, Tseng LM, Chiou SH (2011b) Cucurbitacin I inhibits tumorigenic ability and enhances radiochemosensitivity in nonsmall cell lung cancer-derived CD133-positive cells. Cancer 117:2970–2985
Hu FW, Tsai LL, Yu CH, Chen PN, Chou MY, Yu CC. (2012) Impairment of tumor-initiating stem-like property and reversal of epithelial-mesenchymal transdifferentiation in head and neck cancer by resveratrol treatment. Mol Nutr Food Res 56(8):1247–1258
Hu Y, Fu L (2012) Targeting cancer stem cells: a new therapy to cure cancer patients. Am J Cancer Res 2:340–356
Hutchins-Wolfbrandt A, Mistry AM (2011) Dietary turmeric potentially reduces the risk of cancer. Asian Pac J Cancer Prev 12:3169–3173
Iliopoulos D, Hirsch HA, Struhl K (2011) Metformin decreases the dose of chemotherapy for prolonging tumor remission in mouse xenografts involving multiple cancer cell types. Cancer Res 71:3196–3201
Jaggupilli A, Elkord E (2012) Significance of CD44 and CD24 as cancer stem cell markers: an enduring ambiguity. Clin Dev Immunol 2012:708036
James D, Devaraj S, Bellur P, Lakkanna S, Vicini J, Boddupalli S (2012) Novel concepts of broccoli sulforaphanes and disease: induction of phase II antioxidant and detoxification enzymes by enhanced-glucoraphanin broccoli. Nut Rev 70:654–665
Jayaraman KS (1997) US patent office withdraws patent on Indian herb. Nature 389:6
Joseph I, Tressler R, Bassett E, Harley C, Buseman CM, Pattamatta P, Wright WE, Shay JW, Go NF (2010) The telomerase inhibitor imetelstat depletes cancer stem cells in breast and pancreatic cancer cell lines. Cancer Res 70:9494–9504
Jurgeit A, McDowell R, Moese S, Meldrum E, Schwendener R, Greber UF (2012) Niclosamide is a proton carrier and targets acidic endosomes with broad antiviral effects. PLoS Pathog 8:e1002976
Kakarala M, Brenner DE, Korkaya H, Cheng C, Tazi K, Ginestier C, Liu S, Dontu G, Wicha MS (2010) Targeting breast stem cells with the cancer preventive compounds curcumin and piperine. Breast Cancer Res Treat 122:777–785
Kao CL, Huang PI, Tsai PH, Tsai ML, Lo JF, Lee YY, Chen YJ, Chen YW, Chiou SH (2009) Resveratrol-induced apoptosis and increased radiosensitivity in CD133-positive cells derived from atypical teratoid/rhabdoid tumor. Int J Radiat Oncol Biol Phys 74:219–228
Kallifatidis G, Labsch S, Rausch V, Mattern J, Gladkich J, Moldenhauer G, Büchler MW, Salnikov AV, Herr I (2011) Sulforaphane increases drug-mediated cytotoxicity toward cancer stem-like cells of pancreas and prostate. Mol Ther 19:188–195
Kanwar SS, Yu Y, Nautiyal J, Patel BB, Padhye S, Sarkar FH, Majumdar AP (2011) Difluorinated-curcumin (CDF): a novel curcumin analog is a potent inhibitor of colon cancer stem-like cells. Pharm Res 28:827–838
Kawasaki BT, Hurt EM, Mistree T, Farrar WL (2008) Targeting cancer stem cells with phytochemicals. Mol Interv 8:174–184
Keysar SB, Jimeno A (2010) More than markers: biological significance of cancer stem cell-defining molecules. Mol Cancer Ther 9:2450–2457
Khan N, Mukhtar H (2008) Multitargeted therapy of cancer by green tea polyphenols. Cancer Lett 269:269–280
Kim JB, Ko E, Han W, Shin I, Park SY, Noh DY (2008) Berberine diminishes the side population and ABCG2 transporter expression in MCF-7 breast cancer cells. Planta. Med 74:1693–1700
Kim YS, Farrar W, Colburn NH, Milner JA (2012) Cancer stem cells: potential target for bioactive food components. J Nutr Biochem 23:691–698
Kim YS, Young MR, Bobe G, Colburn NH, Milner JA (2009) Bioactive food components, inflammatory targets, and cancer prevention. Cancer Prev Res 2:200–208
Koushik A, Spiegelman D, Albanes D, Anderson KE, Bernstein L, Brandt PA van den, Bergkvist L, English DR, Freudenheim JL, Fuchs CS, Genkinger JM, Giles GG, Goldbohm RA, Horn-Ross PL, Männistö S, McCullough ML, Millen AE, Miller AB, Robien K, Rohan TE, Schatzkin A, Shikany JM, Stolzenberg-Solomon RZ, Willett WC, Wolk A, Ziegler RG, Smith-Warner SA (2012) Intake of fruits and vegetables and risk of pancreatic cancer in a pooled analysis of 14 cohort studies. Am J Epidemiol 176:373–386
Krause M, Yaromina A, Eicheler W, Koch U, Baumann M (2011) Cancer stem cells: targets and potential biomarkers for radiotherapy. Clin Cancer Res 17:7224–7229
Kroemer G, Pouyssegur J (2008) Tumor cell metabolism: cancer’s Achilles’ heel. Cancer Cell 13:472–482
Kundu JK, Surh YJ (2008) Cancer chemopreventive and therapeutic potential of resveratrol: mechanistic perspectives. Cancer Lett 269:243–261
Latte KP, Appel KE, Lampen A (2011) Health benefits and possible risks of broccoli—a review. Food Chem. Toxicol 49:3287–3309
Lee KW, Bode AM, Dong Z (2011) Molecular targets of phytochemicals for cancer prevention. Nat Rev Cancer 11:211–218
Lee CH, Hong HM, Chang YY, Chang WW (2012a) Inhibition of heat shock protein (Hsp) 27 potentiates the suppressive effect of Hsp90 inhibitors in targeting breast cancer stem-like cells. Biochimie 94:1382–1389
Lee RJ, Xiong G, Kofonow JM, Chen B, Lysenko A, Jiang P, Abraham V, Doghramji L, Adappa ND, Palmer JN, Kennedy DW, Beauchamp GK, Doulias PT, Ischiropoulos H, Kreindler JL, Reed DR, Cohen NA (2012b) T2R38 taste receptor polymorphisms underlie susceptibility to upper respiratory infection. J Clin Invest 122(11):4145–4159 10.1172/JCI64240.
Lee TK, Castilho A, Cheung VC, Tang KH, Ma S, Ng IO (2011) Lupeol targets liver tumor-initiating cells through phosphatase and tensin homolog modulation. Hepatology 53:160–170
Leizer AL, Alvero AB, Fu HH, Holmberg JC, Cheng YC, Silasi DA, Rutherford T, Mor G (2011) Regulation of inflammation by the NF-κB pathway in ovarian cancer stem cells. Am J Reprod Immunol 65(4):438–447
Li HJ, Reinhardt F, Herschman HR, Weinberg RA (2012) Cancer-stimulated mesenchymal stem cells create a carcinoma stem cell niche via prostaglandin E2 signaling. Cancer Discov 2:840–855
Li L, Neaves WB (2006) Normal stem cells and cancer stem cells: the niche matters. Cancer Res 66:4553–4557
Li RJ, Ying X, Zhang Y, Ju RJ, Wang XX, Yao HJ, Men Y, Tian W, Yu Y, Zhang L, Huang RJ, Lu WL (2011) All-trans retinoic acid stealth liposomes prevent the relapse of breast cancer arising from the cancer stem cells. J. Control Release 149:281–291
Li SH, Fu J, Watkins DN, Srivastava RK, Shankar S (2013) Sulforaphane regulates self-renewal of pancreatic cancer stem cells through the modulation of Sonic hedgehog–GLI pathway. Mol Cell Biochem 373(1–2):217–227
Li Y, Schwartz S, Sun D (2011) Potential application of natural dietary components to target cancer stem cells. In: Shostak S (ed) Cancer stem cells—the cutting edge. InTech, Rijeka, Croatia, pp 407–430
Li Y, Wicha MS, Schwartz SJ, Sun D (2011) Implications of cancer stem cell theory for cancer chemoprevention by natural dietary compounds. J Nutr Biochem 22:799–806
Li Y, Zhang T, Korkaya H, Liu S, Lee HF, Newman B, Yu Y, Clouthier SG, Schwartz SJ, Wicha MS, Sun D (2010) Sulforaphane, a dietary component of broccoli/broccoli sprouts, inhibits breast cancer stem cells. Clin Cancer Res 16:2580–2590
Lim KJ, Bisht S, Bar EE, Maitra A, Eberhart CG (2011) A polymeric nanoparticle formulation of curcumin inhibits growth, clonogenicity and stem-like fraction in malignant brain tumors. Cancer Biol Ther 11:464–473
Lin L, Liu Y, Li H, Li PK, Fuchs J, Shibata H, Iwabuchi Y, Lin J (2011) Targeting colon cancer stem cells using a new curcumin analogue, GO-Y030. Br J Cancer 105:212–220
Lin LC, Yeh CT, Kuo CC, Lee CM, Yen GC, Wang LS, Wu CH, Yang WC, Wu AT (2012) Sulforaphane potentiates the efficacy of imatinib against chronic leukemia cancer stem cells through enhanced abrogation of Wnt/β-catenin function. J Agric Food Chem 60:7031–7039
Lippman SM, Hawk ET (2009) Cancer prevention: from 1727 to milestones of the past 100 years. Cancer Res 69(13):5269–5284
Liu RH (2004) Potential synergy of phytochemicals in cancer prevention: mechanism of action. J Nutr 134:3479S–3485S
Lu KH, Chen YW, Tsai PH, Tsai ML, Lee YY, Chiang CY, Kao CL, Chiou SH, Ku HH, Lin CH, Chen YJ (2009) Evaluation of radiotherapy effect in resveratrol-treated medulloblastoma cancer stem-like cells. Childs Nerv Syst 25:543–550
Luk SU, Yap WN, Chiu YT, Lee DT, Ma S, Lee TK, Vasireddy RS, Wong YC, Ching YP, Nelson C, Yap YL, Ling MT (2011) Gamma-tocotrienol as an effective agent in targeting prostate cancer stem cell-like population. Int J Cancer 128:2182–2191
Majumdar AP, Banerjee S, Nautiyal J, Patel BB, Patel V, Du J, Yu Y, Elliott AA, Levi E, Sarkar FH (2009) Curcumin synergizes with resveratrol to inhibit colon cancer. Nutr Cancer 61:544–553
Manach C, Williamson G, Morand C, Scalbert A, Rémésy C (2005) Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am J Clin Nutr 81:230S–242S
Manson MM (2003) Cancer prevention—the potential for diet to modulate molecular signalling. Trends Mol Med 9:11–18
Manson MM, Foreman BE, Howells LM, Moiseeva EP (2007) Determining the efficacy of dietary phytochemicals in cancer prevention. Biochem Soc Trans 35:1358–1363
Mather JP (2012) In vitro models. Stem Cells 30:95–99
Mattson MP (2008) Dietary factors, hormesis and health. Ageing Res Rev 7:43–48
Maund SL, Cramer SD (2011) The tissue-specific stem cell as a target for chemoprevention. Stem Cell Rev 7:307–314
Maxmen A (2012) Calorie restriction falters in the long run. Nature 488:569
McCubrey JA, Chappell WH, Abrams SL, Franklin RA, Long JM, Sattler JA, Kempf CR, Laidler P, Steelman LS (2011) Targeting the cancer initiating cell: the Achilles’ heel of cancer. Adv Enzyme Regul 51:152–162
Meeran SM, Katiyar SK (2008) Cell cycle control as a basis for cancer chemoprevention through dietary agents. Front Biosci 13:2191–2202
Mehta RG, Murillo G, Naithani R, Peng X (2010) Cancer chemoprevention by natural products: how far have we come? Pharm Res 27:950–961
Mo W, Zhang J (2012) Human ABCG2: structure Function and its role in multidrug resistance. Int J Biochem Mol Biol 1:1–27
Montales MT, Rahal OM, Kang J, Rogers TJ, Prior RL, Wu X, Simmen RC (2012) Repression of mammosphere formation of human breast cancer cells by soy isoflavone genistein and blueberry polyphenolic acids suggests diet-mediated targeting of cancer stem-like/progenitor cells. Carcinogenesis 33:652–660
Moutsatsou P (2007) The spectrum of phytoestrogens in nature: our knowledge is expanding. Hormones 6:173–193
Murakami A, Ohigashi H (2007) Targeting NOX, INOS and COX-2 in inflammatory cells: chemoprevention using food phytochemicals. Int J Cancer 121:2357–2363
Neergheen-Bhujun VS, Kang KS, Aruoma OI, Bahorun T (2012) Dietary biofactors in the management of cancer: myth or reality? In: Shankar S, Srivastava RK (eds) Nutrition, diet and cancer. Springer, New York, pp 109–127
Neuzil J, Tomasetti M, Zhao Y, Dong LF, Birringer M, Wang XF, Low P, Wu K, Salvatore BA, Ralph SJ (2007) Vitamin E analogs, a novel group of “mitocans,” as anticancer agents: the importance of being redox-silent. Mol Pharmacol 71:1185–1199
Nguyen LV, Vanner R, Dirks P, Eaves CJ (2012) Cancer stem cells: an evolving concept. Nat Rev Cancer 12:133–143
Nishimura N, Hartomo TB, Pham TV, Lee MJ, Yamamoto T, Morikawa S, Hasegawa D, Takeda H, Kawasaki K, Kosaka Y, Yamamoto N, Kubokawa I, Mori T, Yanai T, Hayakawa A, Takeshima Y, Iijima K, Matsuo M, Nishio H (2012) Epigallocatechin gallate inhibits sphere formation of neuroblastoma BE(2)-C cells. Environ Health Prev Med 17:246–251
Opie LH, Lecour S (2007) The red wine hypothesis: from concepts to protective signalling molecules. Eur Heart J 28:1683–1693
Ottinger S, Klöppel A, Rausch V, Liu L, Kallifatidis G, Gross W, Gebhard MM, Brümmer F, Herr I (2012) Targeting of pancreatic and prostate cancer stem cell characteristics by Crambe crambe marine sponge extract. Int J Cancer 130:1671–1681
Pan MH, Ho CT (2008) Chemopreventive effects of natural dietary compounds on cancer development. Chem Soc Rev 37:2558–2574
Pandey PR, Okuda H, Watabe M, Pai SK, Liu W, Kobayashi A, Xing F, Fukuda K, Hirota S, Sugai T, Wakabayashi G, Koeda K, Kashiwaba M, Suzuki K, Chiba T, Endo M, Fujioka T, Tanji S, Mo YY, Cao D, Wilber AC, Watabe K (2011) Resveratrol suppresses growth of cancer stem-like cells by inhibiting fatty acid synthase. Breast Cancer Res Treat 130:387–398
Pandey PR, Xing F, Sharma S, Watabe M, Pai SK, Iiizumi-Gairani M, Fukuda K, Hirota S, Mo Y-Y, Watabe K (2013) Elevated lipogenesis in epithelial stem-like cell confers survival advantage in ductal carcinoma in situ of breast cancer, Oncogene 32:511–522.
Parr A, Bolwell G (2000) Phenols in the plant and in man. The potential for possible nutritional enhancement of the diet by modifying the phenols content or profile. J Sci Food Agric 80:985–1012
Pastrana E, Silva-Vargas V, Doetsch F (2011) Eyes wide open: a critical review of sphere-formation as an assay for stem cells. Cell Stem Cell 8:486–498
Pearson KJ, Baur JA, Lewis KN, Peshkin L, Price NL, Labinskyy N, Swindell WR, Kamara D, Minor RK, Perez E, Jamieson HA, Zhang Y, Dunn SR, Sharma K, Pleshko N, Woollett LA, Csiszar A, Ikeno Y, Le Couteur D, Elliott PJ, Becker KG, Navas P, Ingram DK, Wolf NS, Ungvari Z, Sinclair DA, Cabo R de (2008) Resveratrol delays age-related deterioration and mimics transcriptional aspects of dietary restriction without extending life span. Cell Metab 8:157–168
Pece S, Tosoni D, Confalonieri S, Mazzarol G, Vecchi M, Ronzoni S, Bernard L, Viale G, Pelicci PG, Di Fiore PP (2010) Biological and molecular heterogeneity of breast cancers correlates with their cancer stem cell content. Cell 140:62–73
Pirola L, Fröjdö S (2008) Resveratrol: one molecule, many targets. IUBMB Life 60:323–332
Priyadarsini RV, Nagini S (2012) Cancer chemoprevention by dietary phytochemicals: promises and pitfalls. Curr Pharm Biotechnol 13:125–136
Quintana E, Shackleton M, Sabel MS, Fullen DR, Johnson TM, Morrison SJ (2008) Efficient tumour formation by single human melanoma cells. Nature 456:593–598
Räsänen K, Herlyn M (2012) Paracrine Signaling between Carcinoma Cells and Mesenchymal Stem Cells Generates Cancer Stem Cell Niche via Epithelial-Mesenchymal Transition. Cancer Discov 2:775–777
Rausch V, Liu L, Kallifatidis G, Baumann B, Mattern J, Gladkich J, Wirth T, Schemmer P, Büchler MW, Zöller M, Salnikov AV, Herr I (2010) Synergistic activity of sorafenib and sulforaphane abolishes pancreatic cancer stem cell characteristics. Cancer Res 70:5004–5013
Ravindran J, Prasad S, Aggarwal BB (2009) Curcumin and cancer cells: how many ways can curry kill tumor cells selectively? AAPS J 11:495–510
Rehman J, Zhang HJ, Toth PT, Zhang Y, Marsboom G, Hong Z, Salgia R, Husain AN, Wietholt C, Archer SL (2012) Inhibition of mitochondrial fission prevents cell cycle progression in lung cancer. FASEB J 26:2175–2186
Reya T, Morrison SJ, Clarke MF, Weissman IL (2001) Stem cells, cancer, and cancer stem cells. Nature 414:105–111
Rodova M, Fu J, Watkins DN, Srivastava RK, Shankar S (2012) Sonic hedgehog signaling inhibition provides opportunities for targeted therapy by sulforaphane in regulating pancreatic cancer stem cell self-renewal. PLoS One 7(9):e46083
Rosen JM, Jordan CT (2009) The increasing complexity of the cancer stem cell paradigm. Science 324:1670–1673
Russo GL (2007) Ins and outs of dietary phytochemicals in cancer chemoprevention. Biochem Pharmacol 74:533–544
Sachlos E, Risueño RM, Laronde S, Shapovalova Z, Lee JH, Russell J, Malig M, McNicol JD, Fiebig-Comyn A, Graham M, Levadoux-Martin M, Lee JB, Giacomelli AO, Hassell JA, Fischer-Russell D, Trus MR, Foley R, Leber B, Xenocostas A, Brown ED, Collins TJ, Bhatia M (2012) Identification of drugs including a dopamine receptor antagonist that selectively target cancer stem cells. Cell 149:1284–1297
Safe S, Papineni S, Chintharlapalli S (2008) Cancer chemotherapy with indole-3-carbinol, bis(3’-indolyl)methane and synthetic analogs. Cancer Lett 269:326–338
Saiko P, Szakmary A, Jaeger W, Szekeres T (2008) Resveratrol and its analogs: defense against cancer, coronary disease and neurodegenerative maladies or just a fad? Mutat. Res 658:68–94
Salvioli S, Sikora E, Cooper EL, Franceschi C (2007) Curcumin in cell death processes: a challenge for CAM of age-related pathologies. Evid Based Complement Alternat Med 4:181–190
Sato A, Kudo C, Yamakoshi H, Uehara Y, Ohori H, Ishioka C, Iwabuchi Y, Shibata H (2011) Curcumin analog GO-Y030 is a novel inhibitor of IKKβ that suppresses NF-κB signaling and induces apoptosis. Cancer Sci 102:1045–1051
Scatena R (2012) Mitochondria and cancer: a growing role in apoptosis, cancer cell metabolism and dedifferentiation. Adv Exp Med Biol 942:287–308
Scatena R, Mordente A, Giardina B (eds) (2012) Advances in cancer stem cell biology. Springer, New York
Scheel C, Weinberg RA (2012) Cancer stem cells and epithelial-mesenchymal transition: concepts and molecular links. Semin Cancer Biol 22:396–403
Schepers AG, Snippert HJ, Stange DE, Born M van den, Es JH van, Wetering M van de, Clevers H (2012) Lineage tracing reveals Lgr5 + stem cell activity in mouse intestinal adenomas. Science 337:730–735
Scholz S, Williamson G (2007) Interactions affecting the bioavailability of dietary polyphenols in vivo. Int J Vitam Nutr Res 77:224–235
Schreiber RD, Old LJ, Smyth MJ (2011) Cancer immunoediting: integrating immunity’s roles in cancer suppression and promotion. Science 331:1565–1570
Scott E, Steward WP, Gescher AJ, Brown K (2012) Resveratrol in human cancer chemoprevention–choosing the ‘right’ dose. Mol Nutr Food Res 56:7–13
Scott EN, Gescher AJ, Steward WP, Brown K (2009) Development of dietary phytochemical chemopreventive agents: biomarkers and choice of dose for early clinical trials. Cancer Prev Res 2:525–530
Semov A, Iourtchenco L, Lin Fang L, Shengmin L, Xu Y, Su X, Muyjnek E, Kiselev V, Alakhov V (2012) Diindolilmethane (DIM) selectively inhibits cancer stem cells. Biochem Biophys Res Commun 424:45–51
Shankar S, Nall D, Tang SN, Meeker D, Passarini J, Sharma J, Srivastava RK (2011) Resveratrol inhibits pancreatic cancer stem cell characteristics in human and KrasG12D transgenic mice by inhibiting pluripotency maintaining factors and epithelial-mesenchymal transition. PloS ONE 6:e16530
Shostak S (ed) (2011a) Cancer stem cells—the cutting edge. InTech.
Shostak S (ed) (2011b) Cancer stem cells theories and practice. InTech, Rijeka, Croatia
Simmen FA, Simmen RC (2011) The maternal womb: a novel target for cancer prevention in the era of the obesity pandemic? Eur J Cancer Prev 20:539–548
Singh SK, Clarke ID, Terasaki M, Bonn VE, Hawkins C, Squire J, Dirks PB (2003) Identification of a cancer stem cell in human brain tumors. Cancer Res 63:5821–5828
Sneddon J, Werb Z (2007) Location, Location, Location: the cancer stem cell niche. Cell Stem Cell 1:607–611
So JY, Lee HJ, Smolarek AK, Paul S, Wang CX, Maehr H, Uskokovic M, Zheng X, Conney AH, Cai L, Liu F, Suh N (2011) A novel Gemini vitamin D analog represses the expression of a stem cell marker CD44 in breast cancer. Mol Pharmacol 79:360–367
Sporn MB, Suh N (2000) Chemoprevention of cancer. Carcinogenesis 21:525–530
Strimpakos AS, Sharma RA (2008) Curcumin: preventive and therapeutic properties in laboratory studies and clinical trials. Antioxid Redox Signal 10:511–545
Subramaniam D, Ponnurangam S, Ramamoorthy P, Standing D, Battafarano RJ, Anant S, Sharma P (2012) Curcumin induces cell death in esophageal cancer cells through modulating Notch signaling. PloS one 7(2):e30590
Subramaniam D, Ramalingam S, Houchen CW, Anant S (2010) Cancer stem cells: a novel paradigm for cancer prevention and treatment. Mini Rev Med Chem 10:359–371
Su Y-C, Li S-C, Wu Y-C, Wang L-M, Chao KSC, Liao HF (2013) Resveratrol Downregulates Interleukin-6-Stimulated Sonic Hedgehog Signaling in Human Acute Myeloid Leukemia. Evid Based Complement Alternat Med 2013:547430
Surh YJ (2003) Cancer chemoprevention with dietary phytochemicals. Nat Rev Cancer 3:768–780
Tan A, Konczak I, Sze D, Ramzan I (2011) Molecular pathways for cancer chemoprevention by dietary phytochemicals. Nut Cancer 63:495–505
Tang SN, Singh C, Nall D, Meeker D, Shankar S, Srivastava RK (2010) The dietary bioflavonoid quercetin synergizes with epigallocathechin gallate (EGCG) to inhibit prostate cancer stem cell characteristics, invasion, migration and epithelial-mesenchymal transition. J Mol Signal 5:14
Tang SN, Fu J, Nall D, Rodova M, Shankar S, Srivastava RK (2012) Inhibition of sonic hedgehog pathway and pluripotency maintaining factors regulate human pancreatic cancer stem cell characteristics. Int J Cancer 131:30–40
Tanveer S, Fathi E, Guy F (2011) Towards new anticancer strategies by targeting cancer stem cells with phytochemical compounds. In: Shostak S (ed) Cancer stem cells—the cutting edge. InTech, Rijeka, Croatia, pp 431–456
Tirino V, Desiderio V, Paino F, De Rosa A, Papaccio F, La Noce M, Laino L, De Francesco F, Papaccio G (2013) Cancer stem cells in solid tumors: an overview and new approaches for their isolation and characterization. The FASEB J 27(1):13–24
Tiwaria N, Gheldof A, Tatari M, Christofori G (2012) EMT as the ultimate survival mechanism of cancer stem cells. Sem Cancer Biol 22:194–207
Trewavas A, Stewart D (2003) Paradoxical effects of chemicals in the diet on health. Curr Opin Plant Biol 6:185–190
Trosko JE (2009) Review paper: cancer stem cells and cancer nonstem cells: from adult stem cells or from reprogramming of differentiated somatic cells. Vet Pathol 46:176–193
Tseng LM, Huang PI, Chen YR, Chen YC, Chou YC, Chen YW, Chang YL, Hsu HS, Lan YT, Chen KH, Chi CW, Chiou SH, Yang DM, Lee CH (2012) Targeting signal transducer and activator of transcription 3 pathway by cucurbitacin I diminishes self-renewing and radiochemoresistant abilities in thyroid cancer-derived CD133 + cells. J Pharmacol Exp Ther 341:410–423
Vanden Berghe W (2012) Epigenetic impact of dietary polyphenols in cancer chemoprevention: lifelong remodeling of our epigenomes. Pharmacol Res 65:565–576
Vazquez-Martin A, Oliveras-Ferraros C, Del Barco S, Martin-Castillo B, Menendez JA (2011) The anti-diabetic drug metformin suppresses self-renewal and proliferation of trastuzumab-resistant tumor-initiating breast cancer stem cells. Breast Cancer Res Treat 126:355–364
Vermeulen L, Sprick MR, Kemper K, Stassi G, Medema JP (2008) Cancer stem cells—old concepts, new insights. Cell Death Differ 15:947–958
Verschoyle RD, Steward WP, Gescher AJ (2007) Putative cancer chemopreventive agents of dietary origin-how safe are they? Nutr Cancer 59(2):152–162
Volate SR, Kawasaki BT, Hurt EM, Milner JA, Kim YS, White J, Farrar WL (2010) Gossypol induces apoptosis by activating p53 in prostate cancer cells and prostate tumor-initiating cells. Mol Cancer Ther 9:461–470
Wade N (2011) Doubt on anti-aging molecule as drug trial stops. The New York Times. 10 January
Wang JY, Chang CC, Chiang CC, Chen WM, Hung SC (2012a) Silibinin suppresses the maintenance of colorectal cancer stem-like cells by inhibiting PP2A/AKT/mTOR pathways. J Cell Biochem 113:1733–1743
Wang K, Zhang T, Liu L, Wang X, Wu P, Chen Z, Ni C, Zhang J, Hu F, Huang J (2012b) Novel micelle formulation of curcumin for enhancing antitumor activity and inhibiting colorectal cancer stem cells. Int J Nanomedicine 7:4487–4497
Weber DA, Wheat JM, Currie GM (2012) Cancer stem cells and the impact of Chinese herbs, isolates and other complementary medical botanicals: a review. Zhong Xi Yi Jie He Xue Bao 10:493–503
Wei X, Senanayake TH, Warren G, Vinogradov SV (2013) Hyaluronic acid-based nanogel–drug conjugates with enhanced anticancer activity designed for the targeting of CD44-positive and drug-resistant tumors. Bioconjug chem 24(4):658–668
Weng JR, Tsai CH, Kulp SK, Chen CS (2008) Indole-3-carbinol as a chempreventive and anti-cancer agent. Cancer Lett 262:153–163
Willett WC (2010) Fruits, vegetables, and cancer prevention: turmoil in the produce section. J Natl Cancer Inst 102:510–511
Willett WC, Ludwig DS (2011) The 2010 Dietary Guidelines—the best recipe for health? N Engl J Med 365:1563–1565
Williamson G, Manach C (2005) Bioavailability and bioefficacy of polyphenols in humans. II. Review of 93 intervention studies. Am J Clin Nutr 81:243S–255S
Woodward WA, Sulman EP (2008) Cancer stem cells: markers or biomarkers? Cancer Metastasis Rev 27:459–470
Xin HW, Hari DM, Mullinax JE, Ambe CM, Koizumi T, Ray S, Anderson AJ, Wiegand GW, Garfield SH, Thorgeirsson SS, Avital I (2012) Tumor-initiating label-retaining cancer cells in humangastrointestinal cancers undergo asymmetric cell division. Stem Cells 30:591–598
Yang CS, Lambert JD, Ju J, Lu G, Sang S (2007) Tea and cancer prevention: molecular mechanisms and human relevance. Toxicol Appl Pharmacol 224:265–273
Yang YP, Chang YL, Huang PI, Chiou GY, Tseng LM, Chiou SH, Chen MH, Chen MT, Shih YH, Chang CH, Hsu CC, Ma HI, Wang CT, Tsai LL, Yu CC, Chang CJ (2012a) Resveratrol suppresses tumorigenicity and enhances radiosensitivity in primary glioblastoma tumor initiating cells by inhibiting the STAT3 axis. J Cell Physiol 227:976–993
Yang Z, Kulkarni K, Zhu W, Hu M (2012b) Bioavailability and pharmacokinetics of genistein: mechanistic studies on its ADME. Anti-Cancer Agents in Medicinal Chemistry-Anti-Cancer Agents 12(10):1264–1280
Yao H, Xu W, Shi X, Zhang Z (2011) Dietary flavonoids as cancer prevention agents. J Environ Sci Health C Environ Carcinog Ecotoxicol Rev 29:1–31
Yo YT, Lin YW, Wang YC, Balch C, Huang RL, Chan MW, Sytwu HK, Chen CK, Chang CC, Nephew KP, Huang T, Yu MH, Lai HC (2012) Growth inhibition of ovarian tumor-initiating cells by niclosamide. Mol Cancer Ther 11:1703–1712
Youle RJ, Bliek AM van der (2012) Mitochondrial fission, fusion, and stress. Science 337:1062–1065
Yu JS (ed) (2009) Cancer stem cells: methods and protocols. Springer, New York
Yu Y, Kanwar SS, Patel BB, Nautiyal J, Sarkar FH, Majumdar AP (2009) Elimination of colon cancer stem-like cells by the combination of curcumin and FOLFOX. Transl Oncol 2:321–328
Yu Y, Sarkar FH, Majumdar AP (2013) Down-regulation of miR-21 induces differentiation of chemoresistant colon cancer cells and enhances susceptibility to therapeutic regimens. Transl oncol 6(2):180
Zaveri NT (2006) Green tea and its polyphenolic catechins: medicinal uses in cancer and noncancer applications. Life Sci 78:2073–2080
Zhou J, Zhang H, Gu P, Bai J, Margolick JB, Zhang Y (2008) NF-kappaB pathway inhibitors preferentially inhibit breast cancer stem-like cells. Breast Cancer Res Treat 111:pp 419–427
Zhang Y, Piao B, Zhang Y, Hua B, Hou W, Xu W, Qi X, Zhu X, Pei Y, Lin H (2011) Oxymatrine diminishes the side population and inhibits the expression of β-catenin in MCF-7 breast cancer cells. Med Oncol 28(Suppl 1):S99–S107
Zhang L, Li L, Jiao M, Wu D, Wu K, Li X, Zhu G, Yang L, Wang X, Hsieh JT, He D (2012a) Genistein inhibits the stemness properties of prostate cancer cells through targeting Hedgehog-Gli1 pathway. Cancer Lett 323:48–57
Zhang L, Yao HJ, Yu Y, Zhang Y, Li RJ, Ju RJ, Wang XX, Sun MG, Shi JF, Lu WL (2012b) Mitochondrial targeting liposomes incorporating daunorubicin and quinacrine for treatment of relapsed breast cancer arising from cancer stem cells. BioMaterials 33:565–582
Zhou BB, Zhang H, Damelin M, Geles KG, Grindley JC, Dirks PB (2009) Tumour initiating cells: challenges and opportunities for anticancer drug discovery. Nat Rev Drug Discov 8:806–823
Zhou W, Kallifatidis G, Baumann B, Rausch V, Mattern J, Gladkich J, Giese N, Moldenhauer G, Wirth T, Büchler MW, Salnikov AV, Herr I (2010) Dietary polyphenol quercetin targets pancreatic cancer stem cells. Int J Oncol 37:551–561
Zhuang W, Long L, Zheng B, Ji W, Yang N, Zhang Q, Liang Z (2012) Curcumin promotes differentiation of glioma-initiating cells by inducing autophagy. Cancer Sci 103(4):684–690
Zobalova R, Prokopova K, Stantic M, Stapelberg M, Dong LF, Ralph SJ, Akporiaye E, Neuzil J (2011) The potential role of CD133 in immune surveillance and apoptosis: a mitochondrial connection? Antioxid Redox Signal 15:2989–3002
Acknowledgments
We thank Drs. Siddavaram Nagini (Annamlai University, Tamil Nadu, India), Roberto Scatena (Catholic University, Rome, Italy), and Zhen Yang (University of Houston, Texas, USA) for preprints and reprints. The chapter was completed during the disruptive events on the eastern United States of Hurricane Sandy in late October 2012. For their dedication, we are grateful to Jennifer Hostettler for improving the text, Jay Patel for figure and references, and Rensa Chen for going over the manuscript. We also thank the editor and the anonymous reviewers for their helpful suggestions in improving this chapter. The writing of this chapter was supported, in part, by an award from the American Institute for Cancer Research to Marion Chan.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer Science+Business Media New York
About this chapter
Cite this chapter
Fong, D., Chan, M. (2013). Dietary Phytochemicals Target Cancer Stem Cells for Cancer Chemoprevention. In: Chandra, D. (eds) Mitochondria as Targets for Phytochemicals in Cancer Prevention and Therapy. Springer, New York, NY. https://doi.org/10.1007/978-1-4614-9326-6_5
Download citation
DOI: https://doi.org/10.1007/978-1-4614-9326-6_5
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4614-9325-9
Online ISBN: 978-1-4614-9326-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)