
Abstract
Hemagglutinin protease (HAP) produced by Vibrio cholerae is the most well characterized protease in pathogenic Vibrio. The matured 45-kDa and 35-kDa processed forms of HAP were purified from a ctx gene negative Vibrio cholerae O1 strain. The 35-kDa HAP showed hemorrhagic fluid response in a dose dependent manner in the rabbit ileal loop assay (RIL). Histopthological examination of purified protease treated rabbit ileum showed the presence of erythrocytes and neutrophils in the upper part of the villous lamina propria, gross damage of the villous epithelium with inflammation, hemorrhage and necrosis. The 35-kDa form of HAP, when added to the luminal surface of the rat ileum loaded in an Ussing chamber, showed a decrease in the intestinal short-circuit current and a cell rounding effect on HeLa cells. Almost all results of earlier studies suggest an indirect pathogenic role of HAP but this study showed the possibility of a direct role of HAP in pathogenesis. The other well characterized protease secreted by Vibrio cholerae is PrtV a metalloprotease which has a role in the protection from predator grazing in natural aquatic environments and also has a role in human pathogenicity. PrtV also modulated hemolysin which plays a role in inflammatory response in human epithelial cells. A novel 59-kDa serine protease was identified from a ∆hapA∆prtV V. cholerae O1 strain. The serine protease has been shown to cause hemorrhagic fluid response in RIL assay. Proteases secreted in Vibrio cholerae play a role in its pathogenesis.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Vibrio cholerae pathogenesis
- Hemagglutinin protease (HAP)
- Vibrio cholerae protease (PrtV)
- 59-kDa Serine protease
1 Introduction
The major features of pathogenesis of cholera are well established. Infection due to Vibrio cholerae begins with the ingestion of contaminated water or food. After passage through the acid barrier of the stomach, the organism colonizes the epithelium of the small intestine by means of the toxin-coregulated pili [1] and possibly other colonization factors such as the different hemagglutinins, accessory colonization factor and core-encoded pilus, all of which are thought to play a role. Cholera enterotoxin produced by the adherent vibrios is secreted across the bacterial outer membrane into the extracellular environment and disrupts ion transport by intestinal epithelial cells. The subsequent loss of water and electrolytes leads to severe diarrhea which is characteristic of cholera [2].
Proteases produced by pathogenic microorganisms play an important role in virulence [3]. Proteases are enzymes that catalyze the hydrolysis of peptide bonds in proteins or peptides. They are either exopeptidases, whose actions are restricted to the N- or C-termini of proteins, or endopeptidases which cleave internal peptide bonds. Microbial peptides are predominantly secreted enzymes and can be classified based on the essential catalytic residue at their active site. They include serine proteases, cysteine proteases, aspartate proteases and metalloproteases. Tissue barriers to pathogen invasion, such as extracellular matrices, epidermal keratinocyte layers and blood vessel walls, may be targeted by bacterial proteases. Proteolysis of host tissue components such as extracellular matrix proteins, including collagen, laminin, fibronectin and elastin, can induce necrotic tissue damage which may aid the bacteria in host cell entry [4, 5]. V. cholerae O1, the causative agent of epidemic cholera, secretes a 32-kDa zinc-containing hemagglutinin protease. Most results suggest that HAP plays an indirect role in pathogenesis. Our study is the first study to show direct role of HAP in pathogenesis of V. cholerae [6]. A metalloprotease, other than HAP, Vibrio cholerae protease (PrtV) plays a role in protection from predator grazing in natural aquatic environments. Using a ∆hapA∆prtV V. cholerae strain a novel 59 kDa serine protease was identified and shown to play a role in hemorrhagic response in rabbit ileal loop [7].
2 Proteases in Vibrio cholerae
2.1 Hemagglutinin Protease
V. cholerae O1 El Tor biotype strains, as well as the non-O1, non-O139 serogroup strains produce several hemagglutinins [8–11] which may potentially be involved in adherence of the vibrios to the human gut. In particular, the soluble hemagglutinin, which also has proteolytic activity [12] is called hemagglutinin/protease (HAP). HAP (vibriolysin) belongs to a family of metalloprotese (M4: M04/003) which is largely distributed among pathogenic bacteria. Comparison of multiple sequences shows considerable sequence similarity among primary structures of all known neutral proteases of M4 family. The active site and metal binding residues are highly conserved. HEXXH is the putative signature of Zn-metalloprotease. This motif contains all of the residues that have been positively identified as part of the active site [13, 14].
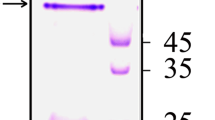
When HAP is purified in the presence of EDTA, a larger 45-kDa form is isolated while in absence of EDTA, a 32-kDa form is purified [6]. The protease undergoes several steps of processing, including cleavage of the signal peptide and further processing of the N terminus and the C terminus to generate the mature protein [14]. The 45-, 37-, 32-, and 9-kDa bands are the major bands bound to anti-HAP antibodies, which suggests that they are the proteolytic degradation products of the 45-kDa polypeptide [15]. Purified HAP in the absence of EDTA from V. cholerae non-O1, non-O139 strain PL-21 in TSB medium showed the presence of both 45- and 35-kDa forms [6]. In the presence of EDTA, the 45-kDa form of HAP was purified in larger amounts but in an in-active state. This is first study in which the two major forms of HAP were purified and studied in Ussing’s chamber, rabbit ileal loop and tissue culture assays. The functional activity of HAP is related to its protease activity, and HAP being a metalloprotease, it shows no protease and HA activity in the presence of EDTA. The mature 45-kDa form of HAP purified in the presence of EDTA showed no effects in RIL, Ussing chamber, and tissue culture assays. Functional forms were generated when 45-kDa HAP was dialyzed in EDTA-free buffer. Once the protease is functional, it is processed, as is evident by the presence of 45, 42, and 35-kDa bands on SDS-polyacrylamide gels when HAP was purified in the absence of EDTA. V. anguillarum secretes a metalloprotease that also exists in two forms, Pa and Pb [16]. The Pb form was converted to the Pa form when the protease was incubated overnight at 4 °C. HAP exists in both the 45-kDa and 35-kDa forms but is not completely converted to the smaller processed form at 4 °C. Functional studies with only the 45-kDa form are not possible, as one cannot prevent its processing to the 35-kDa form in the absence of EDTA. However, the 45-kDa form can be studied through in vitro models such as Ussing chamber and tissue culture assays. This protein, at lower concentrations of 2 μg/ml and 4 μg/ml, exerts an enterotoxic response in the Ussing chamber in the form of an increase in I SC (Fig. 1a) and a cell distending effect in the HeLa cell assay (Fig. 2b). At higher concentrations, it shows a decrease in the I SC in the Ussing chamber (Fig. 1a), suggesting a cytotoxic response and a cell rounding effect on HeLa cells (Fig. 2a). The most probable explanation for this differential effect is that, at lower concentrations of functional 45-kDa HAP, the amount of 35-kDa form is less abundant, while the amount of the processed 35-kDa form is elevated at higher concentrations of 45-kDa HAP and may be higher than a critical level when it exerts a cytotoxic effect. The I SC changes peaked at 4 μg/ml, while the effect observed at 6 μg/ml revealed that the enterotoxic response of the 45-kDa form was masked by the cytotoxic effect of the 35-kDa form, resulting in a decrease in I SC . The enterotoxic response of the 45-kDa HAP started after a delay of 45–60 min and was equivalent to the effect of 1 μg/ml of CT in the rat small intestine [17]. Figure 1a shows that all protein concentrations of 45-kDa form upto 4 μg/ml the I SC exhibited a gradual increase in rabbit ileal mucosa, with the peak being reached at different time intervals, depending on the protein concentration used, indicative of Cl− secretion. The addition of glucose at the end of the experiment showed equivalent responses in treated and untreated tissue, indicating that glucose-Na+ active co-transport function was unaffected. Enterotoxicity is caused by an abnormal flux of negatively charged ions from the serosal side to the mucosal side [18]. Interestingly, at a higher concentration (6 μg/ml), there was a significant decrease in I SC (Fig. 1a). The dose dependent effect of the purified 35-kDa HAP showed a sharp decrease in the intestinal short circuit current (Fig. 1b). The purified 35-kDa form of HAP also showed cell rounding effect (Fig. 2a). It was earlier reported that culture supernatant prepared from the reactogenic strains of V. cholerae cause a decrease in the transcellular epithelial resistance of cultured T84 cells [19]. This decrease correlated with the presence of HAP but not with the presence of other potential accessory toxins or proteases [19].

Dose dependent response of HAP on intestinal short circuit current (Isc) of rat ileum loaded in Ussing’s chamber. HAP was added in absence or presence of EDTA to the Ringer’s solution in Ussing’s chamber and the change in Isc was recorded for next 2 h from the time the sample was added. The data represents the mean and standard deviation for triplicate experiments. Proteins purified from DH5α were used as controls. (a). Response of Isc to increasing doses of 45-kDa HAP. (b). Response of Isc to increasing doses of 35-kDa HAP. There is an initial increase, but the Isc falls as the dose reaches beyond 40 μg/ml. Values are the mean standard deviations (error bars) for triplicate experiments. Proteins purified from E. coli DH5α were used as controls

Effects of HAP on HeLa cells. HeLa cells grown in tissue culture dishes were treated overnight with different concentrations of either the 35-kDa HAP or the 45-kDa HAP. Cells were observed under phase contrast microscope. (a). Cell rounding effect of 35-kDa HAP and higher concentration of 45-kDa HAP. (b). Cell distending effect of lower concentration of 45-kDa HAP (c). Normal HeLa cells grown in tissue culture plates
Purified HAP showed a dose-dependent hemorrhagic fluid response in the rabbit ileal loop assay. At 20 μg, a weak hemorrhagic response was elicited. At HAP concentrations increasing up to 60 μg there was a gradual increase in the fluid accumulation ratio, with hemorrhagic fluid present at all the different concentrations. The protease incubated in the presence of EDTA failed to show any response in RIL assay. The results of the RIL assay were substantiated by histopathological analysis of the ileal loop samples. In all the experiments, the rabbits were sacrificed 18 h postinoculation. The villous architecture and mucosa in the buffer control sections were normal (Fig. 3a). Purified HAP at a concentration of 20 μg resulted in little distortion of the villous pattern. The blood vessels were congested, and inflammatory cells were present in the lamina propia. High-powered photomicrograph showed the presence of erythrocytes in the upper part of the villous lamina propria. After treatment with 40 μg of purified HAP, there were marked alterations of the villous contour with red blood cell and inflammatory cell infiltrates in the villous epithelium and lamina propria and in damaged crypt epithelium (Fig. 3b). Higher magnification of portions of Fig. 3a, b revealed the clear presence of erythrocytes, eosinophils, and neutrophil polymorphs in the lamina propia (Fig. 3c, d). Washed V. cholerae 569B cultures, on the other hand, showed non-hemorrhagic fluid accumulation in the RIL assay. The active dialyzed 45-kDa form tested for entertoxigenicity in the RIL assay also showed hemorrhagic fluid response

Effect of purified HAP in the RIL assay. Purified HAP treated ileal tissues were processed for histopathological analysis and photomicrographs were taken. (a) Villus architecture observed in 25 mM Tris-HCl treated ileal tissues (10× magnification). Figure shows normal villous structure. (b) 40 μg of purified 35-kDa HAP treated rabbit ileal tissues show disruption of normal villus architecture with shortening of the villi (20× magnification). (c) 40× magnification of Fig. 4b shows infiltration of polymorphonuclear neutrophils, eosinophils and erythrocytes. (d) The inset box in Fig. 4c was further magnified (200×) to show the presence of RBC (R), neutrophils (N) and eosinophils (E). Magnification 200×
HAP can proteolytically activate CT-A subunit [20] and the El Tor cytolysin/hemolysin [21] and can hydrolyze several physiologically important proteins such as mucin, fibronectin, and lactoferrin [22]. HAP perturbs the paracellular barrier of cultured intestinal epithelial cells [19, 23] by acting on tight-junction-associated proteins [23] and promotes the detachment of vibrios from monolayers and mucin [24, 25].
2.2 Vibrio cholerae Protease (PrtV)
The second most well characterized protease in Vibrio cholerae is the Vibrio cholerae protease (PrtV). Using reverse molecular genetics techniques an extracellular protease was identified as PrtV protein as being necessary for killing nematodes by Vibrio cholerae [26]. The killing effect is associated with colonization of the C. elegans intestine. The nematode Caenorhabditis elegans has been used successfully as an invertebrate model to screen for virulence factors of several human pathogens e.g. P. aeruginosa and S. typhimurium [27, 28]. V. cholerae lethal infection of the C. elegans depends on a HapR-regulated protease. HAP encoded by hapA gene is regulated by hapR, and is the major extracellular protease. However, when hapA mutant V. cholerae was tested in C. elegans killing assay no attenuation was found in comparison with the wild type. The culture supernatant from hapA mutant contained some proteins not found in the supernatant of hapR mutant [26]. The three most abundant putative hapR-regulated proteins detected in the supernatants from the hapA mutant V. cholerae by mass spectrophotometer revealed that the protein bands corresponded to proteins encoded by the ORFs VCA0812, VCA0813 and VCA0223. The protein products are a leucine aminopeptidase-related protease, leucine amino aminopeptidase (Lap) [29] and the PrtV protease [30] respectively. Knock mutants ∆prtV, ∆lap and ∆lapX mutants were tested in C. elegans model and of all the mutants only ∆prtV mutant showed complete attenuation in comparison to wild type. Neither of the ∆lap and ∆lapX mutants showed significant attenuation. Furthermore, the cloned prtV gene resulted in an increased killing effect by the wild type V. cholerae strain C6706, and it restored a significant killing effect in the case of the hapR mutant derivative. PrtV is important for V. cholerae inhibition of natural Bacteriovorous predators. V. cholerae caused a marked reduction in the number of viable protozoa (Cafeteria roenbergensis and Tetrahymena pyriformis) during prolonged co-cultivation, whereas the bacterial density remained virtually unaltered. Furthermore V. cholerae with mutations abolishing the hapR or prtV genes appeared strongly attenuated in the predator grazing test, and most of the bacteria were consumed by the predator within 3 days. The metalloprotease PrtV was purified from the culture supernatants of a Vibrio cholerae derivative that is deficient in several other secreted peptidases, including the otherwise abundant hemagglutinin protease [31]. The PrtV is synthesized as a 102 kDa protein, but undergoes several N-and C-terminal processing steps during V. choleraeenvelope translocation and prolonged incubation [31]. Purified V. cholerae PrtV protease forms of 81 or 73-kDa were stabilized by calcium ions. Removal of calcium resulted in further rapid autoproteolysis. In assay using cultured cells of the human intestinal cell line HCTS, the PrtV protein showed a cytotoxic effect leading to cell death. PrtV also degraded the extracellular matrix components fibronectin and fibrinogen. PrtV mutants were not attenuated in the infant mouse model, nor did they exhibit reduced colonization potential compared with wild type in competition experiments [30]. PrtV also modulates the response of Vibrio cholerae hemolysin in human intestinal epithelial cells. Vibrio cholerae hemolysin is capable of causing an inflammatory response characterized by increased permeability and production of IL-8 and TNF-alpha in tight monolayers. The inflammatory response was totally abolished by treatment with PrtV [32].
2.3 -kDa Serine Protease
Although HAP is a very active virulence factor, an isogenic strain of V. cholerae mutated in the hap gene was no less virulent in infant rabbits than the parental strain [24]. An earlier study by Hase et al. [33] showed that a hapA-deleted mutant of V. cholerae O1 had reduced extracellular proteolytic activity compared with the parental strain in a skim milk assay, indicating that the mutant still produces some extracellular proteolytic activity. In addition, residual proteolytic activity expressed by the hapA-deleted mutant is distinct from HAP, as demonstrated by failure of anti-HAP serum to inhibit the activity of this secondary protease on milk agar. The mutant strain also failed to agglutinate chicken erythrocytes [24]. Young and Broadbent [34] described several extracellular proteases in V. cholerae that could explain the residual proteolytic activity of the hap-negative V. cholerae mutant. Besides HAP, the other major well-characterized protease in V. cholerae is a 97-kDa Vibrio cholerae protease, PrtV. PrtV plays a role in virulence in a C. elegans infection model [26].
A hapA and prtV double knock out mutant of V. cholerae strain CHA6.8∆prtV still had residual protease activity [7]. This protease was partially purified from strain CHA6.8∆prtV. The ammonium sulphate precipitated proteins from culture supernatants of CHA6.8∆prtV were loaded onto an anion exchange chromatography column (DE-52) [7]. The proteins in the non-binding (NB) fraction of the column (Fig. 4a) were pooled and concentrated. The bound proteins were eluted with 0.1 M (Fig. 4b) and 0.3 M (Fig. 4c) NaCl, dialyzed against 25 mM Tris-HCl buffer and concentrated. When protease activity in the NB, 0.1 M and 0.3 M NaCl eluted fractions were tested by azocasein assay, the major protease activity was present in the NB fraction (Fig. 4d). The NB-pooled fraction was concentrated and run on a native PAGE (Fig. 4e). The major protein band was excised and analyzed by MS/MS sequencing (Fig. 4e). The sequences highlighted showed homology with a 59-kDa serine protease encoded by the gene VC1649 (Fig. 4f). The sequence GDSGGP (underlined) flanks the serine residue in trypsin-like serine proteases (Fig. 4f). To determine the nature of the partially purified protease from CHA6.8∆prtV eluted in the non-binding fraction of a DE52 anion-exchange column, we performed protease inhibition assays with several inhibitors (Fig. 4g). The protease was partially inhibited in the presence of EDTA (60.3 %), EGTA (59.2 %) and PMSF (60.3 %). The partially purified protease was completely inhibited when PMSF and EDTA are used together (Fig. 4g). There was significantly less inhibition of protease activity in the presence of 1, 10-phenanthroline (9 %), aprotinin (10.5 %), leupeptin (8.7 %) and E64 (1.8 %). Although EDTA inhibited protease activity by 60.3 %, EDTA in the presence of CaCl2 inhibited activity by only 3.6 % inhibition (Fig. 4g). The serine protease secreted byCHA6.8∆prtV is a calcium-dependent serine protease.

Partial purification and identification of protease. Chromatographic profile of ammonium sulphate precipitated crude proteins from culture supernatants of CHA6.8∆prtV strain loaded onto an anion exchange column (DE-52). (a) Proteins eluted in the non-binding fraction (NB), (b) proteins eluted with 0.1 M NaCl, (c) proteins eluted with 0.3 M NaCl, +/− shows presence or absence of protease activity, (d) azocasein assay with pooled samples (30 μg) NB, 0.1M#1, 0.1M#2, 0.3 M and crude proteins. (e) Native PAGE profile (lane 1) of crude proteins of CHA6.8∆prtV strain and (lane 2) of partially purified protease (NB) from DE-52 column. The marked protein band was analyzed by MS/MS sequencing and the peptides highlighted showed homology with a 59-kDa trypsin-like serine protease encoded by VC1649. (f) The underlined GDSGGP are the amino acid sequences around the serine residue present in trypsin-like serine proteases. (g) Protease inhibition test of NB fraction (5 μg) with protease inhibitors 10 mM EDTA, 25 mM PMSF, 25 mM PMSF and 10 mM EDTA, 10 mM EDTA and 20 mM CaCl2, 10 mM EGTA, 1 μg/ml aprotinin, 28 mM E64, 1 μg/ml leupeptin and 10 mM 1,10-phenanthroline incubated for 30 min at 37 °C. Residual protease activity was assayed by azocasein assay. Twenty-five mM Tris-HCl was used as a negative control. The values shown are the means with standard deviations from three experiments
To study the role of the 59-kDa serine protease in virulence, 50 μg of the partially purified protease was injected into the rabbit ileum, which induced significant hemorrhagic fluid accumulation (FA ratio 1.2+/20.2, n = 3, Fig. 5a). When a similar concentration of the protease was incubated in the presence of PMSF and EDTA and injected into the rabbit ileum, there was a significant decrease in fluid accumulation (FA ratio 0.3+/20.05, n = 3, Fig. 5a). Histopathological analysis of the rabbit ileum revealed that the protease caused extensive damage to all the layers of the mucosa. There was damage to the villus surface structure with hemorrhage in all layers of the mucosa. On the other hand, analysis of the ileal tissues treated with the protease in presence of both PMSF and EDTA, revealed normal microvillus structure with no gross alteration in villus structure, although the villus lamina propria was slightly dilated and RBCs had accumulated in a few places in the basal area. PMSF and EDTA completely inhibited protease activity (Fig. 5a), but still we observed some residual effect in the rabbit ileal loop. This effect could be due to some other domain in the protease, which may not be its proteolytic domain, and could be responsible for causing damage to the ileal tissue. Tissues treated with 25 mM Tris-HCl and PMSF + EDTA did not cause fluid accumulation in RIL (FA ratio 0.12+/20.002 n = 3, Fig. 5a)

Rabbit ileal loop assay. (a) RIL response of partially purified protease (50 μg, NB) showing significant hemorrhagic fluid accumulation (FA ratio 1.2 +/−0.2 n = 3) and its effect after inhibition with 25 mM PMSF and 10 mM EDTA (NB + PMSF + EDTA) shows significant decrease in fluid accumulation (FA ratio 0.3 +/−0.05 n = 3). Twenty five mM Tris-HCl with 25 mM PMSF + 10 mM EDTA was used as a negative control (FA ratio = 0.12 +/−0.002, n = 3). (b) RIL response with culture supernatants of C6709 (FA ratio 1.1 +/− 0.3, n = 3), CHA6.8 (FA ratio 1.08 +/− 0.2, n = 3), CHA6.8∆prtV (FA ratio 1.02 +/− 0.2, n = 3), CHA6.8∆prtV∆VC1649 (FA ratio 0.11 +/− 0.005, n = 3) and Tryptic soy broth as negative control (FA ratio 0.09 +/− 0.002, n = 3)
Rabbit ileal loop assay with one ml of culture supernatant of C6709, CHA6.8 and CHA6.8∆prtV strain induced significant fluid accumulation (Fig. 5b) in RIL model (FA ratio 1.1+/20.3, n = 3; 1.08+/20.2, n = 3; and 1.02+/20.2, n = 3 respectively) where as CHA6.8∆prtV∆VC1649 strain which is devoid of the serine protease gene and sterile tryptic soy broth, TSB (negative control) did not induce any fluid accumulation (Fig. 5b) (FA ratio 0.11+/20.005, n = 3 and 0.09+/20.002, n = 3 respectively). Almost similar results were observed when one ml of washed bacterial cells (109 cfu/ml) of C6709, CHA6.8 and CHA6.8∆prtV induced significant fluid accumulation (FA ratio 1.2+/20.35, n = 3; 1.1+/20.3, n = 3; and 1.0+/20.2, n = 3 respectively). Bacterial cells of CHA6.8∆prtV∆VC1649 strain did not induce any fluid accumulation (FA ratio 0.15+/20.005, n = 3). Ileal tissues treated with culture supernatant of CHA6.8∆prtV strain also showed dilated villi with gross hemorrhage in all layers of the mucosa. The ileal tissues treated with culture supernatant of CHA6.8∆prtV∆VC1649 strain protease showed villous architecture almost normal with minimum hemorrhage in mucosa and sub-mucosa.
V. vulnificus also secretes a 59-kDa serine protease, which is the free form, while the 69-kDa protein may be a complex form associated non-covalently with small peptide(s) [35]. The proteolytic activity of the final preparation was almost completely abolished by treatment with 5 mM PMSF, a well-known inhibitor of serine proteases. By contrast, tetraethylenepentamine, a specific inhibitor of metalloproteases including VVP (Vibrio vulnificus protease), showed no inhibitory effect on the proteolytic activity. Vibrio vulnificus strain NCIMB 2137, in which vvp has been deleted, secretes a serine protease, VVA0302. V. vulnificus serine protease may be a virulence factor in vibriosis, which is characterized by external and internal hemorrhages affecting the major organs [36], or human wound infection with necrotic tissue damage [37].
The proteomic analysis of the V. cholerae type II secretome revealed the presence of three related serine proteases named as (ves: vibrio extracellular serine proteases) vesA, vesB and vesC [38]. Experiments suggest that vesA, vesB and vesC do not contribute to the intestinal survival and colonization capability of V. cholerae in mice. VesA has shown to be responsible for processing the A subunit of cholera toxin during in-vitro growth of V. cholerae strain N 16961. VesC encoded by VC1649 (59-kDa serine protease) is not capable of processing cholera toxin under tested conditions but has been shown by us to play a role in hemorrhagic response in rabbit ileal loop and it may play a role in pathogenesis [7].
3 Leucine Aminopeptidase
The aminopeptidases (APs) form a group of zinc-dependent metalloproteases that catalyze the removal of amino acids from the N terminus of a protein. These enzymes are of biological and medical importance because of their role in protein degradation and in the metabolism of biologically active peptides. Leucine APs (LAPs) preferably cleave leucyl substrates, although substantial rates of enzymatic cleavage with most amino-terminal residues are observed [39]. The structural gene for the Vibrio cholerae leucine aminopeptidase (lap) has been cloned and sequenced [29]. The molecular weight of LAP is 54.4 kDa. The deduced amino acid sequence of the entire protein showed high homology with the sequence of Vibrio proteolyticus leucine aminopeptidase. The protease showed maximum activity at pH 9.0 and was thermostable at 70 °C. The substrate leucyl-p-nitroanilide was cleaved by the protease, and its activity was inhibited by EDTA and bestatin. LAP may play an important role in releasing the free amino acids needed for the regulation of expression of other virulence factors [40].
4 Conclusions
V. cholerae, secretes several proteases like HAP, PrtV and 59-kDa serine protease. The major protease secreted by V. cholerae strains is the HAP. Almost all results suggest indirect pathogenic roles of HAP, the possibility of a direct role is also reported. Ghosh et al. [6] assayed the enterotoxic activity of HAP purified from a cholera toxin-negative V. cholerae strain. The purified HAP showed a positive hemorrhagic fluid response in the rabbit ileal loop assay and an increase in the intestinal short circuit current in the Ussing chamber assay. A metalloprotease, other than HAP, PrtV is thought to play a role in aquatic environment. Vaitkevicius et al. [26] showed the effect of PrtV against predator grazing in aquatic environment. PrtV also plays a role in pathogenicity of V. cholerae. The serine protease from a ∆hapA∆prtV V. cholerae O1 strain induced hemorrhagic response in rabbit ileal loop [7]. Role of proteases in pathogenicity in V. cholerae could also depend on the expression levels of proteases. In an earlier study by Young and Broadbent [34], 100 strains of V. cholerae El Tor from different parts of the world were screened for protease production by a rapid assay with gelatin agar plates. Based on protease production, the strains were classified as high, medium and low protease producers. The expression of these proteases may play a role in pathogenicity of V. cholerae strains. Our results suggest that the expression of proteases in V. cholerae may follow a cascade of events. HAP controls secretion of the 59-kDa serine protease, which in turn may control secretion of other proteases.
References
Taylor RK, Miller VL, Furlong DB et al (1987) Use of phoA gene fusions to identify a pilus colonization factor coordinately regulated with cholera toxin. Proc Natl Acad Sci U S A 84:2833–2837
Kaper JB, Morris JG, Levine MM (1995) Cholera. Clin Microbiol Rev 8:48–86
Hase CC, Finkelstein RA (1993) Bacterial extracellular zinc-containing metalloproteases. Microbiol Rev 57:823–837
Harrington DJ (1996) Bacterial collagenases and collagen degrading enzymes and their potential role in human disease. Infect Immun 64:1885–1891
Miyoshi S, Shinoda S (2000) Microbial metalloproteases and pathogenesis. Microbes Infect 2:91–98
Ghosh A, Saha DR, Hoque KM et al (2006) Enterotoxigenicity of mature 45-kilodalton and processed 35-kilodalton forms of hemagglutinin protease purified from a cholera toxin gene-negative Vibrio cholerae non-O1, non-O139 strain. Infect Immun 74:2937–2946
Syngkon A, Elluri S, Koley H et al (2010) Studies on a novel serine protease of a ∆hapA ∆prtV Vibrio cholerae O1 strain and its role in hemorrhagic response in the rabbit ileal loop model. PLoS One 5(9):e13122
Finkelstein RA, Arita M, Clements J et al (1978) Proceedings of the 13th joint conference on cholera, U.S.-Japan cooperative medical science program. National Institutes of Health publication number 78-1590. National Institutes of Health, Bethesda, MD, pp 137–151
Finkelstein RA, Hanne LF (1982) Purification and characterization of the soluble hemagglutinin (cholera lectin) produced by Vibrio cholerae. Infect Immun 36:1199–1208
Booth BA, Finkelstein RA (1986) Presence of the HA/protease and other potential virulence factors in O1 and non-O1 Vibrio cholerae. J Infect Dis 154:183–186
Honda T, Booth BA, Boesman-Finkelstein M et al (1987) Comparative study of Vibrio cholerae non-O1 protease and soluble hemagglutinin with those of Vibrio cholerae O1. Infect Immun 55:451–454
Booth BA, Boesman-Finkelstein M, Finkelstein RA (1983) Vibrio cholerae soluble hemagglutinin/protease is a metalloenzyme. Infect Immun 42:639–644
Jongeneel CV, Bouvier J, Bairoch A (1989) A unique signature identifies a family of zinc-dependent metallopeptidases. FEBS Lett 242:211–214
Domann E, Leimeister-Wachter M, Goebel W et al (1991) Molecular cloning, sequencing, and identification of a metalloprotease gene from Listeria monocytogenes that is species specific and physically linked to the listeriolysin gene. Infect Immun 59:65–72
Kimsey HH, Waldor MK (1998) Vibrio cholerae hemagglutinin/protease in-activates CTX. Infect Immun 66:4025–4029
Norqvist A, Norman B, Wolf-Watz H (1990) Identification and characterization of a zinc metalloprotease associated with invasion by the fish pathogen Vibrio anguillarum. Infect Immun 58:3731–3736
Lu L, Baldear ME, Savidge T et al (2003) Development of microbial-human enterocyte interaction: cholera toxin. Pediatr Res 54:211–218
Field M, McCall I (1971) Ion transport in rabbit ileal mucus: I, Na and Cl fluxes and short-circuit current. Am J Physiol 220:1388–1396
Mel SF, Fullner KA, Wimer-Mackin S et al (2000) Association of protease activity in Vibrio cholerae vaccine strains with decrease in transcellular epithelial resistance of polarized T84 intestinal epithelial cells. Infect Immun 68:6487–6492
Booth BA, Boesman-Finkelstein M, Finkelstein RA (1984) Vibrio cholerae hemagglutinin/protease nicks cholera enterotoxin. Infect Immun 45:558–560
Nagamune K, Yamamoto K, Naka A et al (1996) In vitro proteolytic processing and activation of the recombinant precursor of El Tor cytolysin/hemolysin (pro-HlyA) of Vibrio cholerae by soluble hemagglutinin/protease of V. cholerae, trypsin, and other proteases. Infect Immun 64:4655–4658
Finkelstein RA, Boesman-Finkelstein M, Holt P (1983) Vibrio cholerae hemagglutinin/lectin/protease hydrolyzes fibronectin and ovomucin: F.M. Burnet revisited. Proc Natl Acad Sci U S A 180:1092–1095
Wu Z, Milton D, Nybom P et al (1996) Vibrio cholerae hemagglutinin/protease (HA/protease) causes morphological changes in cultured epithelial cells and perturbs their paracellular barrier function. Microb Pathog 21:111–123
Finkelstein RA, Boesman-Finkelstein M, Chang Y et al (1992) Vibrio cholerae hemagglutinin/protease, colonial variation, virulence, and detachment. Infect Immun 60:472–478
Benítez JA, Garcia A, Silva H et al (1999) Preliminary assessment of the safety and immunogenicity of a new CTX - negative, hemagglutinin/protease-defective El Tor strain as a cholera vaccine candidate. Infect Immun 67:539–545
Vaitkevicius K, Lindmark B, Ou G et al (2006) A Vibrio cholerae protease needed for killing of Caenorhabditis elegans has a role in protection from natural predator grazing. Proc Natl Acad Sci U S A 103:9280–9285
Alegado RA, Campbell MC, Chen WC et al (2003) Characterization of mediators of microbial virulence and innate immunity using the Caenorhabditis elegans host-pathogen model. Cell Microbiol 5(7):435–444
Labrousse A, Chauvet S, Couillault C et al (2000) Caenorhabditis elegans is a model host for Salmonella typhimurium. Curr Biol 10(23):1543–1545
Toma C, Honma Y (1996) Cloning and genetic analysis of Vibrio cholerae aminopeptidase gene. Infect Immun 64:4495–4500
Ogierman MA, Fallarino A, Riess T et al (1997) Characterization of Vibrio choleras El Tor lipase operon lipAB and a protease gene of the downstream of the hly region. J Bacteriol 179:7072–7080
Vaitkevicius K, Rompikuntal PK, Lindmark B et al (2008) The metalloprotease PrtV from Vibrio cholerae purification and properties. FEBS J 275:3167–3177
Ou G, Rompikuntal PK, Bitar A et al (2009) Vibrio cholerae cytolysin causes an inflammatory response in human intestinal epithelial cells that is modulated by the PrtV protease. PLoS One 4:e7806
Hase CC, Finkelstein RA (1991) Cloning and nucleotide sequence of the Vibrio cholerae hemagglutinin/protease (HA/protease) gene and construction of an HA/protease negative strain. J Bacteriol 173:3311–3317
Young DB, Broadbent DA (1982) Biochemical characterization of extracellular proteases from Vibrio cholerae. Infect Immun 37:875–883
Wang J, Sasaki T, Maehara Y et al (2008) Variation of extracellular proteases produced by Vibrio vulnificus clinical isolates: genetic diversity of the metalloprotease gene (vvp), and serine protease secretion by vvp-negative strains. Microb Pathog 44:494–500
Biosca EG, Amaro C, Esteve C et al (1991) First record of Vibrio vulnificus biotype 2 from diseased European eel, Anguilla Anguilla L. J Fish Dis 14:103–109
Miyoshi S (2006) Vibtio vulnificus infection and metalloprotease. J Dermatol 33:589–595
Sikora AE, Zielke RA, Lawrence DA et al (2011) Proteomic analysis of the Vibrio cholerae type II secretome reveals new proteins, including three related serine proteases. J Biol Chem 286:16555–16566
Burley SK, David PR, Taylor A et al (1990) Molecular structure of leucine aminopeptidase at 2.7-Ǻ resolution. Proc Natl Acad Sci U S A 87:6878–6882
Miller VL, Mekalanos JL (1988) A novel suicide vector and its use in construction of insertion mutation, osmoregulation of outer membrane and virulence determinants in Vibrio cholerae requires toxR. J Bacteriol 170:2575–2583
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer Science+Business Media New York
About this chapter
Cite this chapter
Pal, A., Wai, S.N. (2013). Proteases in Vibrio cholerae and Their Role in Pathogenesis. In: Chakraborti, S., Dhalla, N. (eds) Proteases in Health and Disease. Advances in Biochemistry in Health and Disease, vol 7. Springer, New York, NY. https://doi.org/10.1007/978-1-4614-9233-7_14
Download citation
DOI: https://doi.org/10.1007/978-1-4614-9233-7_14
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4614-9232-0
Online ISBN: 978-1-4614-9233-7
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)