
Abstract
In addition to chemical modifications such as acetylation or methylation, histones are modified by the addition of small polypeptides such as ubiquitin. Whereas poly-ubiquitylation generally targets proteins for proteasomal degradation, histones are predominantly mono-ubiquitylated. Histone mono-ubiquitylation regulates nuclear processes that include transcription and DNA damage responses. The carboxy-terminal tails of histones H2A and H2B provide the major sites for mono-ubiquitylation. However, recent studies have identified mono-ubiquitylated residues on the amino-terminal tails of H2A and H2B. These include novel lysines that are ubiquitylated in response to DNA damage. In this review, first the enzymes that deposit and remove ubiquitin from histones are described. Although these enzymes are best characterized with respect to histone ubiquitylation, a number of these enzymes have other substrates that might play an important role in their cellular activities. Second, the role of H2A and H2B mono-ubiquitylation in gene expression is discussed; whereas H2B mono-ubiquitylation activates transcription initiation and elongation, H2A mono-ubiquitylation promotes transcription repression. Finally, regulation of histone ubiquitylation during the DNA damage response and in the cell cycle is described.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Ataxia Telangiectasia Mutate
- Nucleosome Occupancy
- High Order Chromatin Structure
- Nucleosome Stability
- Histone Ubiquitylation
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
6.1 Ubiquitylation Is a Reversible Post-translational Modification
Proteins are subject to a variety of post-translational modifications through the attachment of specific moieties to target amino acids. While many of these post-translational modifications involve small chemical molecules, polypeptides can also be conjugated specifically to substrate proteins. The 76-amino acid protein ubiquitin was the first of these protein-based post-translational modifications to be identified (Hicke 2001). Ubiquitin can be attached to protein substrates either as a monomer or as a poly-ubiquitin chain (Hicke 2001). Poly-ubiquitylation has a well-characterized role in targeting proteins for proteasomal degradation [reviewed in Clague and Urbe (2010), Weissman et al. (2011)]. In contrast, mono-ubiquitylation regulates distinct cellular processes that include transcription and endocytosis [reviewed in Hicke (2001)].
6.1.1 Mechanism of Ubiquitylation
In most substrates, the carboxy-terminal glycine (Gly77) of ubiquitin is conjugated to the ε-amino NH2 side chain of an internal lysine (Lys) residue in the target protein through an isopeptide bond. However, ubiquitin can also be attached to other amino acids including serine (Ser), threonine (Thr), cysteine (Cys), and the α-amino NH2 group of substrate proteins (Weissman et al. 2011). Thus, different types of mono-ubiquitylation are possible. Poly-ubiquitylation also differs depending on which of the seven different lysines within ubiquitin is linked in the poly-ubiquitin chain (Weissman et al. 2011). Thus, whereas poly-ubiquitin chains formed through the linkage of ubiquitin via Lys48 are characteristically associated with proteasomal degradation, poly-ubiquitin chains containing Lys63 linkages are involved in cellular processes that include endosomal sorting and the DNA damage response (Clague and Urbe 2010; Weissman et al. 2011; Mattiroli et al. 2012).
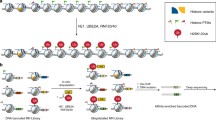
Ubiquitin is conjugated to substrate proteins in a series of reactions involving three separate enzymatic activities [reviewed in Weissman et al. (2011)]. First, ubiquitin is activated by an E1 ubiquitin-activating enzyme in an ATP-dependent reaction (Fig. 6.1). This activation results in the generation of an ubiquitin–AMP product that remains bound to the E1 enzyme. Following activation, ubiquitin is conjugated via a thioester bond to a cysteine residue within an E2 ubiquitin-conjugating enzyme, which functions as an intermediate acceptor for the activated ubiquitin. Lastly, an E3 ubiquitin-protein isopeptide ligase transfers activated ubiquitin from the E2 enzyme to the substrate. There are two classes of E3 ubiquitin ligases that have different mechanisms of action and contain either a HECT or RING finger domain. Whereas HECT domain-containing E3 ligases form a temporary thioester bond with activated ubiquitin prior to transferring it to substrate proteins, RING finger domain-containing E3 ligases promote direct transfer of ubiquitin from the E2 enzyme to the substrate. The last step of this enzymatic cascade provides the majority of the specificity to substrate protein selection; thus there are many different E3 ubiquitin ligases, which play an important role in substrate identification (Clague and Urbe 2010; Weissman et al. 2011).

The 76-amino acid polypeptide ubiquitin is activated and attached to substrate proteins in a series of enzymatic reactions involving E1, E2, and E3 enzymes. First, ubiquitin is conjugated to the active site cysteine of an E1 ubiquitin-activating enzyme via a thioester bond, in an ATP-dependent reaction. In this reaction, the carboxy-terminal glycine (Gly77) of ubiquitin is adenylated and activated, resulting in an ubiquitin–AMP that is bound to the E1 enzyme. Next, the activated-ubiquitin is transferred to the active site cysteine of an E2 ubiquitin-conjugating enzyme. Lastly, an E3 ubiquitin-protein isopeptide ligase transfers the Gly77 of ubiquitin to the ε-amino NH2 side chain of an internal lysine in the target protein through an isopeptide bond. Whereas RING finger domain-containing E3 ligases transfer ubiquitin directly from the E2 enzyme to the substrate protein, HECT domain-containing E3 ligases form a temporary thioester bond with activated ubiquitin before transferring it to the substrate. Ubiquitylation is reversible, and specific proteases known as de-ubiquitylating enzymes (DUBs) remove ubiquitin from target proteins, generating free ubiquitin
6.1.2 Reversal of Protein Ubiquitylation
Ubiquitylation is reversible, and ubiquitin is removed from substrate proteins by de-ubiquitylating enzymes (DUBs) [reviewed in Komander et al. (2009), Reyes-Turcu et al. (2009)] (Fig. 6.1). There are five families of DUBs (Komander et al. 2009; Reyes-Turcu et al. 2009). Four DUB families contain papain-like cysteine protease domains including the ubiquitin carboxy-terminal hydrolase (UCH), the ubiquitin-specific protease (USP/UBP), the ovarian tumor domain (OTU), and the Machado-Josephin domain (MJD) DUBs. The fifth family of DUBs is the JAB1/MPN/Mov34 metalloenzyme (JAMM/MPN+) DUBs. Members of the JAMM/MPN + DUB family contain a zinc-dependent metalloprotease domain. The USP/UBP family is the largest class of DUBs and includes members with specificity for ubiquitylated histones (Reyes-Turcu et al. 2009). Members within this DUB family contain a highly conserved USP domain fold that resembles the finger, palm, and thumb of a right hand (Reyes-Turcu et al. 2009). Ubiquitin binds within this right-hand structure with its carboxy terminus situated in a cleft between the thumb and palm subdomains, while its globular domain interacts with the finger subdomain (Reyes-Turcu et al. 2009). In addition to the core USP domain fold, many USP/UBP family members contain protein-interaction domains that define substrate specificity and mediate association with adaptors or scaffold proteins (Reyes-Turcu et al. 2009). Several of these protein-interaction domains within USP/UBPs are predicted to bind ubiquitin, such as the zinc finger ubiquitin-specific protease (ZnF-UBP) domain (Komander et al. 2009).
6.2 Ubiquitylation of Histones
Histones were first identified as targets for ubiquitylation more than three decades ago [reviewed in Osley (2006)]. In contrast to most ubiquitylated proteins, histones are predominantly mono-ubiquitylated rather than poly-ubiquitylated [reviewed in Osley (2006), Weake and Workman (2008)]. H2A and H2B are the most prominent ubiquitylated histones. However, histone variants and the other histones can also be both mono- and poly-ubiquitylated.
6.2.1 Mono-ubiquitylation of Histones
Mono-ubiquitylated histone H2A (ubH2A) was first identified in the late 1970s as a unique histone-like chromosomal protein, A24 (Goldknopf et al. 1975, 1977; Goldknopf and Busch 1975, 1977; Hunt and Dayhoff 1977; Ballal et al. 1975). Soon after the discovery of ubH2A, mono-ubiquitylated histone H2B (ubH2B) was detected in mouse cells (West and Bonner 1980; Pina and Suau 1985). In mammalian cells, approximately 10 % of histone H2A and 1 % of histone H2B are mono-ubiquitylated (West and Bonner 1980; Pina and Suau 1985). UbH2B is present in all eukaryotes. UbH2A is present in vertebrates, plants, and invertebrates but has not been detected in Saccharomyces cerevisiae (Swerdlow et al. 1990; Robzyk et al. 2000; Sridhar et al. 2007; Sanchez-Pulido et al. 2008; Bratzel et al. 2010; Chen et al. 2010; Alatzas and Foundouli 2006; Gorfinkiel et al. 2004; Gutierrez et al. 2012; Wang et al. 2004; Calonje et al. 2008; de Napoles et al. 2004). The carboxy-terminal tails of histones H2A and H2B provide the major sites for mono-ubiquitylation (Fig. 6.2). However, recent studies have identified residues on the amino-terminal tails of both histones H2A and H2B that are also mono-ubiquitylated.

Specific residues within the amino- and carboxy-terminal tails of human histones H2A and H2B are mono-ubiquitylated. The amino acid sequences of human histones H2B (panel a) and H2A (panel b) are displayed schematically to illustrate the position of ubiquitylation sites on the amino- and carboxy-terminal tails relative to the core histone-fold domains. The human E3 ubiquitin-protein isopeptide ligases involved in mono-ubiquitylation of specific residues are indicated by arrows next to the target lysines
The major site of mono-ubiquitylation on histone H2A is Lys119 in mammalian cells, which corresponds to Lys121 in Arabidopsis thaliana, and Lys118 in Drosophila melanogaster (Goldknopf and Busch 1977; Sanchez-Pulido et al. 2008; Bratzel et al. 2010; Wang et al. 2004; de Napoles et al. 2004; Lagarou et al. 2008). Recent studies have shown that Lys13 and Lys15 on histone H2A are mono-ubiquitylated in response to DNA damage (Mattiroli et al. 2012). The predominant residue mono-ubiquitylated on histone H2B is Lys120 in mammalian cells, which corresponds to Lys143 in A. thaliana, Lys123 in S. cerevisiae, and Lys119 in Schizosaccharomyces pombe (Robzyk et al. 2000; Sridhar et al. 2007; Thorne et al. 1987; Tanny et al. 2007). Additional residues in mammalian histone H2B are also subject to mono-ubiquitylation. These residues include Lys34 and Lys125 in mammalian histone H2B (Wu et al. 2011; Minsky and Oren 2004). However, transfection of plasmids expressing histone H2B point mutations in human cells demonstrates that Lys120 is the major site of H2B mono-ubiquitylation in vivo (Wu et al. 2011).
In this chapter, ubH2A will refer to H2A mono-ubiquitylated at Lys119 in mammalian cells or at its equivalent residues in A. thaliana and D. melanogaster unless otherwise stated. Correspondingly, unless otherwise specified, ubH2B will refer to Lys123-ubiquitylated histone H2B in S. cerevisiae or to its equivalent, Lys120-ubiquitylated histone H2B in mammalian cells.
In addition to the canonical histones, histone variants are ubiquitylated (Hatch et al. 1983; Nickel et al. 1987). The mammalian histone H2A variants H2A.Z and macroH2A1.2 are mono-ubiquitylated at Lys120/Lys121 and Lys115, respectively (Sarcinella et al. 2007; Chu et al. 2006). Furthermore, histone variants such as H2AX that are involved in the response to DNA damage are also subject to mono-, di-, and poly-ubiquitylation (Mattiroli et al. 2012; Huen et al. 2007; Ikura et al. 2007; Kolas et al. 2007; Wang and Elledge 2007).
6.2.2 Poly-ubiquitylation of Histones
Although histone mono-ubiquitylation is more prevalent, histones and histone variants are also poly-ubiquitylated. Poly-ubiquitylated histones H2A and H2B have been detected (Nickel et al. 1987; Sung et al. 1988). In addition, histone poly-ubiquitylation plays important roles in the DNA damage response (for further discussion, see Sect. 6.6 ).
Poly-ubiquitylated histone H3 has been detected both in vitro and in rat testes (Chen et al. 1998; Haas et al. 1990). Furthermore, the centromere-specific histone H3 variant Cse4 (CENP-A in humans) is poly-ubiquitylated in S. cerevisiae (Hewawasam et al. 2010; Ranjitkar et al. 2010). Whereas mono-ubiquitylation of histones is more generally involved in signaling processes, histone H3 poly-ubiquitylation regulates histone protein levels, through degradation, during the cell cycle in S. cerevisiae (Gunjan and Verreault 2003). In addition, poly-ubiquitylation of histone variants can regulate their chromosomal distribution. For example, ubiquitylation of the histone variant Cse4 by the E3 ubiquitin-protein isopeptide ligase Psh1 regulates its chromosomal distribution (Hewawasam et al. 2010; Ranjitkar et al. 2010). This centromere-specific histone is redistributed to euchromatin in psh1Δ yeast (Hewawasam et al. 2010; Ranjitkar et al. 2010). Thus, ubiquitylation-mediated degradation is important both for proper cell cycle regulation and for the correct chromosomal distribution of histones and histone variants.
6.3 Mono-ubiquitylation of Histones Is Catalyzed by Specific E2/E3 Enzymes
Although ubH2A was identified in the late 1970s, the enzymes that catalyze mono-ubiquitylation of histones H2A and H2B remained uncharacterized for another two decades. Early hints as to the identity of enzymes involved in histone ubiquitylation came from in vitro studies in S. cerevisiae (Sung et al. 1988; Jentsch et al. 1987). Four years following the discovery of yeast H2B-specific E2 ubiquitin conjugase, the first complex containing the E3 ubiquitin-protein isopeptide ligase specific for mono-ubiquitylation of histone H2A was isolated from human cells (Robzyk et al. 2000; Wang et al. 2004; Cao et al. 2005). Since then, a number of E2 and E3 enzymes involved in histone mono-ubiquitylation have been discovered. However, the relative contribution of some of these E2 and E3 enzymes to in vivo histone mono-ubiquitylation still remains to be determined. In addition, some mammalian E3 enzymes have been recently discovered that mono-ubiquitylate residues on histones H2A and H2B that differ from the major sites at Lys119 and Lys120, respectively (Mattiroli et al. 2012; Wu et al. 2011; Minsky and Oren 2004).
6.3.1 Mono-ubiquitylation of Histone H2B
The first enzyme identified as a mediator of histone mono-ubiquitylation was the S. cerevisiae DNA repair protein Rad6 (Ubc2), which is an E2 ubiquitin-conjugating enzyme. Rad6 initially exhibited poly-ubiquitylation activity on both histones H2A and H2B in vitro (Sung et al. 1988; Jentsch et al. 1987). However, studies soon showed that in vivo Rad6 catalyzes mono-ubiquitylation, rather than poly-ubiquitylation, of yeast histone H2B at Lys123 (Robzyk et al. 2000). Homologs of Rad6 include Rhp6 in S. pombe, Dhr6 in D. melanogaster, AtUBC2 in A. thaliana, and UBE2A (HHR6A/RAD6A) and UBE2B (HHR6B/RAD6B) in humans (Table 6.1) (Reynolds et al. 1990; Koken et al. 1991a, b; Zwirn et al. 1997; Kim et al. 2009). Another human E2 enzyme, UBE2E1 (UbcH6), has been implicated in H2B ubiquitylation in vitro (Zhu et al. 2005; Pavri et al. 2006). However, cells treated with small interfering RNA (siRNA) against UBE2A and UBE2B, but not UBE2E1, exhibit decreased ubH2B levels (Kim et al. 2009). This finding indicates that these two human RAD6 homologs rather than UBE2E1 are the bona fide E2 conjugases for H2B ubiquitylation in vivo (Kim et al. 2009).
Shortly following the identification of Rad6 as the E2 ubiquitin conjugase for ubH2B, the RING finger domain-containing protein Bre1 was identified as the E3 ubiquitin-protein isopeptide ligase specific for ubH2B at Lys123 in S. cerevisiae (Wood et al. 2003a; Hwang et al. 2003). Bre1 homologs have since been identified in multiple species and include Brl1 (Rfp2/Spcc1919.15) and Brl2 (Rfp1/Spcc970.10c) in S. pombe, Bre1 in D. melanogaster, HUB1 in A. thaliana, and RNF20 and RNF40 in humans (Tanny et al. 2007; Kim et al. 2005, 2009; Zhu et al. 2005; Zofall and Grewal 2007; Bray et al. 2005; Fleury et al. 2007). In both S. cerevisiae and S. pombe, Rad6 and Bre1 exist as a complex together with a third protein, Lge1 (Shf1 in S. pombe), that is also required for H2B mono-ubiquitylation (Tanny et al. 2007; Hwang et al. 2003; Zofall and Grewal 2007). In mammals, a third protein, WAC, also interacts with RNF20 and RNF40 and is necessary for H2B mono-ubiquitylation in vivo (Zhang and Yu 2011).
In addition to the mammalian Bre1 orthologs, RNF20/40, other mammalian E3 ubiquitin-protein isopeptide ligases are implicated in histone H2B mono-ubiquitylation. These E3 ligases include the human RING finger domain-containing proteins MSL2, BRCA1, and MDM2 (Wu et al. 2011; Minsky and Oren 2004; Chen et al. 2002; Mallery et al. 2002; Xia et al. 2003). All three of these E3 ligases target lysines within human histone H2B other than Lys120 (Wu et al. 2011; Minsky and Oren 2004; Chen et al. 2002; Mallery et al. 2002; Xia et al. 2003). MSL2 is a component of the human MOF-MSL complex and mono-ubiquitylates nucleosomal histone H2B on Lys34 both in vitro and in vivo (Wu et al. 2011). BRCA1 exhibits ubiquitylation activity on both histones H2A and H2B in vitro (Chen et al. 2002; Mallery et al. 2002; Xia et al. 2003; Zhu et al. 2011). However, in vivo BRCA1 is implicated in the regulation of heterochromatic ubiquitylation of H2A (Chen et al. 2002; Mallery et al. 2002; Xia et al. 2003; Zhu et al. 2011). MDM2 ubiquitylates both histones H2A and H2B in vitro (Minsky and Oren 2004). Overexpression of MDM2 results in an increase in ubH2B levels in vivo. Both Lys120 and Lys125 of histone H2B are required for this ubiquitylation, suggesting both lysines are targets (Minsky and Oren 2004).
6.3.2 Mono-ubiquitylation of Histone H2A
Following the identification of the E2/E3 enzymes involved in histone H2B mono-ubiquitylation, biochemical studies identified Ring finger protein 2 (RNF2, also known as Ring2/Ring1B) as an E3 ubiquitin-protein isopeptide ligase that possesses in vitro activity against histone H2A at Lys119 (Wang et al. 2004; Cao et al. 2005). RNF2 was first identified as a component of the Polycomb repressive complex 1 (PRC1) (Table 6.1) (Wang et al. 2004; Cao et al. 2005). The PRC1 class complexes consist of four core subunits that are homologous to the Drosophila Polycomb (Pc), Sex combs extra (Sce/dRing), Polyhomeotic (Ph), and Posterior sex combs (Psc) proteins (Kerppola 2009). Loss of Sce in Drosophila results in a global decrease in ubH2A levels (Gutierrez et al. 2012; Lagarou et al. 2008). There are multiple homologs of each of these four PRC1 proteins in mammals that are classified into the Cbx, Ring1, Phc, and Bmi1/Mel18 families (Kerppola 2009). Thus mammalian cells have an increased diversity in PRC1 complexes that are capable of mediating H2A mono-ubiquitylation as compared with flies. In addition, other factors have been shown to interact with the PRC1 core in mammals and in flies. For example, RNF2 is a subunit of two other repressive complexes in mammalian cells, E2F-6.com-1 and FBXL10-BcoR, that contain the PRC1 core and additional subunits (Gearhart et al. 2006; Ogawa et al. 2002; Sanchez et al. 2007). Furthermore, the Drosophila RNF2-ortholog Sce is a subunit of the Drosophila Ring-Associated Factor (dRAF) complex, in addition to being a subunit of PRC1 (Gorfinkiel et al. 2004; Gutierrez et al. 2012; Wang et al. 2004; Lagarou et al. 2008).
In addition to RNF2, there are other RING finger domain-containing subunits within the PRC1 class complexes, suggesting that these proteins could also catalyze H2A Lys119 ubiquitylation. For example, the PRC1 core itself contains two additional RING finger domain-containing proteins, Ring1 (Rnf1/Ring1A) and Bmi1 (Wang et al. 2004; Cao et al. 2005). However, only RNF2 possesses in vitro E3 ubiquitin-protein isopeptide ligase activity specific for histone H2A (Wang et al. 2004; Cao et al. 2005). These findings indicate that the other RING finger domain-containing proteins within the RNF2-containing complexes in mammals, such as Bmi1, are not directly involved in H2A mono-ubiquitylation (Wang et al. 2004; Cao et al. 2005; Li et al. 2006; Wei et al. 2006). Nonetheless, Bmi1 does stimulate the E3 ubiquitin-protein isopeptide ligase activity of RNF2 (Wang et al. 2004; Cao et al. 2005; Li et al. 2006; Wei et al. 2006). Furthermore, this interplay between subunits in stimulating RNF2 activity is not limited to the PRC1 complex. NSPC1, a subunit of the FBXL10–BcoR complex that has homology to Bmi1, also stimulates RNF2 E3 ubiquitin-protein isopeptide ligase activity on histone H2A (Sanchez et al. 2007). Thus, other RING finger domain-containing subunits within the PRC1 family of multi-subunit complexes can enhance RNF2-mediated mono-ubiquitylation of histone H2A.
Recently, homologs of RNF2 and the other PRC1 complex subunits were identified in A. thaliana (Sanchez-Pulido et al. 2008; Bratzel et al. 2010; Chen et al. 2010; Xu and Shen 2008). Prior to this discovery, it had been unclear whether Arabidopsis histone H2A could in fact be mono-ubiquitylated. This is because the consensus ubiquitylation sequence, PKKT, is present in only one of the 13 isoforms of histone H2A in Arabidopsis, H2A.1 (Sanchez-Pulido et al. 2008; Bratzel et al. 2010). However, recently it has been shown that the Arabidopsis homologs of Bmi1 and RNF2, AtBMI1A (DRIP2), AtBMI1B (DRIP1), AtRING1A, and AtRING1B, are all capable of mono-ubiquitylating H2A.1 in vitro (Bratzel et al. 2010). Furthermore, in vitro experiments suggest that these E3 ubiquitin-protein isopeptide ligases interact with the other PRC1L subunit orthologs, EMBRYONIC FLOWER1 (EMF1), and the Pc ortholog, LIKE-HETEROCHROMTAIN PROTEIN1 (LHP1) (Bratzel et al. 2010). Lastly, ubH2A levels are reduced in both Atbmi1a-1/Atbmi1b and emf1-2 mutant backgrounds, suggesting that the Arabidopsis PRC1L complex indeed mediates H2A ubiquitylation in vivo (Bratzel et al. 2010).
In addition to RNF2, other E3 ubiquitin-protein isopeptide ligases specific for histone H2A have been identified in mammalian cells. Several of these are implicated in mono-ubiquitylation of residues in H2A that differ from Lys119. These ligases include DZIP3, HUWE1, DDB1-CUL4DDB2, RNF8, RNF168, and BRCA1. DZIP3 (DAZ-interacting protein 3 zinc finger, also known as 2A-HUB/hRUL138) is a component of the N-CoR/HDAC1/3 repressive complex (Zhou et al. 2008). HUWE1 (HECT, UBA, and WWE domain containing 1, E3 ubiquitin protein ligase, also known as LASU1) is a testis-specific HECT domain-containing protein that exhibits mono-ubiquitylation activity on histone H2A in vitro (Liu et al. 2005; Rajapurohitam et al. 1999). However a role of HUWE1 in H2A mono-ubiquitylation in vivo has not yet been demonstrated. DDB1-CUL4DDB2, RNF8, and RNF168 are implicated in DNA damage-induced ubiquitylation of histones H2A and H2AX (Mattiroli et al. 2012; Bergink et al. 2006; Kapetanaki et al. 2006; Mailand et al. 2007; Doil et al. 2009; Stewart et al. 2009) (for further discussion, see Sect. 6.6 ). Recent studies have shown that RNF168 specifically mono-ubiquitylates nucleosomal histones H2A and H2AX at Lys13 and Lys15 (Mattiroli et al. 2012). The tumor suppressor BRCA1 regulates H2A ubiquitylation at heterochromatic loci (Zhu et al. 2011). BRCA1 mono-ubiquitylates histone H2A and the histone variant H2AX in vitro, and its activity is stimulated by a second RING domain-containing protein BARD1 (Chen et al. 2002; Mallery et al. 2002; Xia et al. 2003). Loss of Brca1 in mice causes loss of ubH2A at satellite repeats and a reduction in condensed DNA regions in the genome (Zhu et al. 2011). Furthermore, these effects are reversed upon ectopic expression of H2A fused to ubiquitin at its Lys119 residue, suggesting that the effects of BRCA1 are mediated via H2A-Lys119 ubiquitylation (Zhu et al. 2011). BRCA1 might also be involved in H2A ubiquitylation at other loci because it has been shown to regulate ubiquitylation of H2A at the promoters of progesterone receptor target genes (Calvo and Beato 2011). Thus, several other E3 ubiquitin ligases ubiquitylate histone H2A, at a variety of lysine residues, in addition to RNF2.
Despite the characterization of a number of E3 ubiquitin-protein isopeptide ligases specific for histone H2A, identification of the corresponding E2 ubiquitin-conjugating enzymes has been less successful. Several different E2 ubiquitin-conjugating enzymes including UBE2D1/2/3 (UbcH5a/b/c) and UBE2E1 can function together with RNF2 to catalyze H2A ubiquitylation in vitro (Li et al. 2006; Buchwald et al. 2006). Notably, there is a crystal structure of the Bmi1/RNF2 heterodimer together with the UBE2D3 E2 ubiquitin-conjugating enzyme, supporting a role for UBE2D3 in H2A ubiquitylation (Bentley et al. 2011). In addition, the E2 ubiquitin-conjugating enzymes UBC4-1 and UBC4-testis specific catalyze H2A ubiquitylation in vitro in the presence of HUWE1 (Liu et al. 2005; Rajapurohitam et al. 1999). Furthermore, the E2 ubiquitin conjugase UBE2N (UBC13) is associated with RNF8 and is required for ubiquitylation of the phosphorylated H2AX (γH2AX) histone variant (Huen et al. 2007; Ikura et al. 2007; Kolas et al. 2007; Wang and Elledge 2007). The numbers of E2 ubiquitin-conjugating enzymes implicated in H2A ubiquitylation suggest that these enzymes might function redundantly to catalyze H2A ubiquitylation in vivo. Thus, E2 enzymes might have a limited role in regulating the specificity of H2A ubiquitylation.
6.3.3 Non-histone Targets of E2/E3 Enzymes Involved in Histone Ubiquitylation
Several of the E2 and E3 enzymes, which mono-ubiquitylate histones H2A and H2B, also have other protein substrates. Here, we describe three examples of E2 and E3 enzymes involved in histone ubiquitylation that have additional protein substrates. The finding that histone-specific E2 and E3 enzymes have other protein substrates demonstrates that careful analysis is required when examining phenotypes resulting from loss of these enzymes, as these phenotypes might only partially result from their role in histone ubiquitylation.
First, the Arabidopsis H2A-specific E3 ubiquitin-protein isopeptide ligases, AtBMI1A and AtBMI1B, also ubiquitylate the transcription factor DEHYDRATION-RESPONSIVE ELEMENT-BINDING PROTEIN 2A (DREB2A) in vitro (Qin et al. 2008). However, it is worth noting that in mammals, Bmi1 does not directly ubiquitylate histone H2A (Wang et al. 2004; Cao et al. 2005; Li et al. 2006; Wei et al. 2006). Second, another E3 ligase implicated in H2A ubiquitylation, BRCA1, also ubiquitylates the progesterone receptor targeting it for degradation in the absence of hormone (Calvo and Beato 2011). Thus, H2A-specific E3 ubiquitin ligases may also ubiquitylate non-histone targets. In addition, some of the E2 ubiquitin-conjugating enzymes involved in histone ubiquitylation regulate ubiquitylation of other non-histone substrates. Most notably, Rad6 has a number of functions that are distinct from H2B ubiquitylation, including DNA damage repair and protein degradation pathways. For example, Rad6 in conjunction with the E3 ubiquitin-protein isopeptide ligases, Rad18 and Rad5, mono-ubiquitylates proliferating cell nuclear antigen (PCNA) at replication forks that have stalled due to DNA damage [reviewed in Lee and Myung (2008)]. In addition, Rad6 together with Ubr1 targets short-lived protein substrates for degradation via the 26S proteasome (Sung et al. 1991; Watkins et al. 1993; Xie and Varshavsky 1999). Furthermore, studies suggest that the mammalian Rad6 homologs might regulate ubiquitylation of substrates as diverse as beta-catenin and p53 (Gerard et al. 2012; Chen et al. 2012). Thus, non-histone substrates should be considered when examining the effects on gene expression of the E2 and E3 enzymes that mediate histone ubiquitylation.
6.4 Histone Mono-ubiquitylation Is Reversed by De-ubiquitylating Enzymes
Following the discovery of the enzymes that catalyze the addition of ubiquitin to histones H2A and H2B, it soon became apparent that DUBs that reverse histone ubiquitylation also exist. The DUBs with activity against ubH2B were the first of these histone-specific DUBs to be identified, first in yeast and later in other eukaryotes. Soon after, a number of proteins with de-ubiquitylation activity against ubH2A were identified in mammalian cells and in Drosophila. Whereas these DUBs catalyze de-ubiquitylation of mono-ubiquitylated histones, recent work shows that there are other substrates for these DUBs in vivo that might also play important roles in regulating aspects of gene expression.
6.4.1 De-ubiquitylation of ubH2B
The first two distinct DUBs specific for ubH2B were identified in S. cerevisiae: Ubp8 and Ubp10 (Henry et al. 2003; Daniel et al. 2004; Emre et al. 2005; Gardner et al. 2005). Whereas Ubp8 is a subunit of the Spt-Ada-Gcn5 acetyltransferase (SAGA) transcription coactivator complex, Ubp10 functions independently of SAGA and was originally implicated in telomeric silencing (Henry et al. 2003; Daniel et al. 2004; Emre et al. 2005; Gardner et al. 2005; Kahana and Gottschling 1999). Ubp8 orthologs include Nonstop in D. melanogaster and USP22 in humans (Table 6.1) (Weake et al. 2008; Zhang et al. 2008; Zhao et al. 2008). Structural studies have shown that Ubp8 resides within a four-subunit module within SAGA and that complex structural interactions with the other proteins in this module, Sgf11, Sgf73, and Sus1, are required for its de-ubiquitylating activity (Kohler et al. 2010; Samara et al. 2010, 2012). These interactions are conserved in humans, and the respective orthologs in human SAGA, ATXN7L3, ATXN7, and ENY2, are also required for USP22 activity (Lang et al. 2011). Orthologs of the second DUB specific for ubH2B, Ubp10, include Scrawny in D. melanogaster and SUP32 (UBP26) in A. thaliana (Sridhar et al. 2007; Buszczak et al. 2009). Other members of the USP/UBP family of DUBs have also been implicated in ubH2B de-ubiquitylation. In D. melanogaster, USP7 catalyzes de-ubiquitylation of ubH2B in vitro and in vivo (van der Knaap et al. 2005, 2010). USP7 associates with a biosynthetic enzyme, GMP synthetase, which is required for its de-ubiquitylase activity (van der Knaap et al. 2005, 2010). In addition, human USP3 might also play a role in ubH2B de-ubiquitylation (Nicassio et al. 2007).
The two major ubH2B DUBs, Ubp8 and Ubp10, remove ubiquitin from ubH2B at different locations on genes in yeast. Early evidence supporting the activity of these two DUBs on distinct pools of ubH2B was provided by the observation that deletion of both UBP8 and UBP10 results in a greater increase in the global level of ubH2B relative to either of the single deletions (Emre et al. 2005; Gardner et al. 2005). Genome-wide studies of ubH2B distribution in yeast lacking either Ubp8 or Ubp10 confirmed that these DUBs indeed act on ubH2B at different locations (Schulze et al. 2011). Whereas Ubp8 functions primarily in 5′ transcribed regions that are marked by H3 Lys4 tri-methylation, Ubp10 acts on ubH2B within the body of transcribed genes that contain H3 Lys79 tri-methylation (Schulze et al. 2011). Although early studies suggested that Ubp10 was involved in telomeric silencing through interactions with Sir4, genome-wide studies of ubH2B indicate that this modification is absent from telomeric regions (Emre et al. 2005; Gardner et al. 2005; Kahana and Gottschling 1999; Schulze et al. 2011). Thus, both Ubp8 and Ubp10 are involved in de-ubiquitylation of ubH2B at different locations on actively transcribed genes.
6.4.2 De-ubiquitylation of ubH2A
Several of the DUBs involved in ubH2B de-ubiquitylation also exhibit activity, at least in vitro, on ubH2A. USP22 and USP3 de-ubiquitylate ubH2A in vitro, and knockdown of human ATXN7L3, which is required for USP22 activity, results in a global increase in ubH2A levels (Zhao et al. 2008; Lang et al. 2011; Nicassio et al. 2007). Some evidence suggests that USP3 might de-ubiquitylate ubH2A at DNA damage sites (Doil et al. 2009; Nicassio et al. 2007) (for further discussion, see Sect. 6.6.4 ). In addition, USP12 and USP46 are implicated in de-ubiquitylation of both ubH2A and ubH2B in Xenopus (Joo et al. 2011).
In addition to USP22 and USP3, three DUBs specific for ubH2A have been identified in mammals: USP16 (Ubp-M), MYSM1 (2A-DUB/KIAA1915), and USP21 (Table 6.1) (Joo et al. 2007; Nakagawa et al. 2008; Zhu et al. 2007). Transient transfection of USP16 results in a reduction in ubH2A levels in human cells (Cai et al. 1999; Mimnaugh et al. 2001). In addition, USP16 de-ubiquitylates nucleosomal ubH2A in vitro, and knockdown of USP16 results in a global increase in ubH2A levels (Joo et al. 2007). MYSM1 differs from many of the other known histone DUBs in that it contains a JAMM/MPN + zinc-dependent metalloprotease domain rather than the USP/UBP domain commonly found in histone DUBs (Zhu et al. 2007). Similarly to Ubp8/USP22, MYSM1 also interacts with the histone acetyltransferase, p300/CBP-associated factor (PCAF/KAT2B), and preferentially de-ubiquitylates acetylated nucleosomes in vitro (Zhu et al. 2007). Moreover, knockdown of MYSM1 in cultured cells results in an increase in global ubH2A levels (Zhu et al. 2007). USP21 de-ubiquitylates ubH2A in vitro and relieves transcriptional repression in vivo (Nakagawa et al. 2008). The relative contribution of the three ubH2A-specific DUBs to ubH2A de-ubiquitylation is not clear, and these DUBs might have both redundant and specific functions in different cell types.
Recently, the UCH domain-containing Polycomb Group protein Calypso was also identified as an ubH2A-specific DUB in Drosophila (Scheuermann et al. 2010). Calypso is a subunit of the Polycomb repressive de-ubiquitylating enzyme (PR-DUB) complex, together with another Polycomb Group protein, ASX (Scheuermann et al. 2010). Moreover, calypso mutant flies have increased global levels of ubH2A (Scheuermann et al. 2010). Notably, the mammalian homolog of Calypso, BRCA1-associated protein (BAP1), also interacts with the ASX homolog, ASXL1, in vitro (Scheuermann et al. 2010). Although the DUB activity of mammalian BAP1 on ubH2A has not yet been characterized, knockdown of BAP1 in malignant pleural mesothelioma cell lines results in the misregulation of Polycomb-target genes (Bott et al. 2011). Thus, the human PR-DUB complex might also de-ubiquitylate ubH2A and could contribute a fourth ubH2A-specific DUB activity in mammalian cells.
6.4.3 Non-histone Targets of Histone DUBs
Similar to the non-histone substrates identified for the E2 and E3 enzymes involved in histone ubiquitylation, other substrates have also been identified as targets of the DUBs which de-ubiquitylate histones H2A and H2B. For example, in addition to ubH2B, Ubp8/USP22 de-ubiquitylates several other substrates. These substrates include the mammalian telomeric-repeat-binding factor 1 (TRF1), the sucrose non-fermenting 1 (Snf1) AMP protein kinase in S. cerevisiae, and the human transcriptional regulator (FUSE)-binding protein 1 (FBP1) (Atanassov et al. 2009; Wilson et al. 2011; Atanassov and Dent 2011). In addition, Ubp10 de-ubiquitylates PCNA in S. cerevisiae (Gallego-Sanchez et al. 2012). Furthermore, the mammalian homolog of the ubH2A DUB Calypso, BAP1, has been shown to bind and de-ubiquitylate Host Cell Factor (HCF) (Machida et al. 2009; Misaghi et al. 2009). Thus, de-ubiquitylation of non-histone substrates by the DUBs that catalyze histone de-ubiquitylation should be considered when examining the effects of these enzymes on cellular processes.
6.5 Transcription Is Regulated by Histone Mono-ubiquitylation
In general, while mono-ubiquitylation of histone H2A is involved in transcriptional repression, ubH2B plays important roles in transcription initiation and elongation. Intriguingly, recent studies suggest that both the addition and the removal of ubiquitin from histones H2A and H2B might play an important role in regulating transcriptional status.
6.5.1 ubH2B Correlates with Active Transcription
H2B mono-ubiquitylation occurs at actively transcribed genes, and high levels of ubH2B are present on the transcribed region of abundantly expressed human genes (Minsky et al. 2008; Jung et al. 2012). Although ubH2B plays an important role in the events that take place during the early stages of the transcription cycle, recent work suggests that ubH2B can also have repressive roles at specific gene loci. Thus, ubH2B has multiple separable roles in regulating transcription. First, ubH2B regulates the recruitment and/or activity of complexes that mediate several of the sequential events that occur during transcription activation at promoters and at the 5′ end of genes (see Sects. 6.5.1.4 – 6.5.1.8 ). Second, ubH2B directly promotes nucleosome stability and/or occupancy, which has differential effects on gene expression at promoters and transcribed regions (see Sect. 6.5.1.11 ). Thus, ubH2B regulates gene expression differentially both at individual genes and at specific locations within the promoter and transcribed region of those genes.
6.5.1.1 ubH2B Is Enriched on Transcribed Regions of Highly Expressed Genes
The highest ubH2B levels on expressed human genes are observed at a region extending from just downstream of the transcription start site (tss) into the middle of the coding/transcribed region (Kim et al. 2009; Minsky et al. 2008; Jung et al. 2012). The level of ubH2B on the gene then decreases toward the 3′ end of the transcribed region (Kim et al. 2009; Minsky et al. 2008; Jung et al. 2012). In S. cerevisiae, ubH2B is enriched on the coding regions of highly expressed genes and is also present on the coding region of long genes (Shieh et al. 2011). Notably, levels of ubH2B are low at the tss of most genes, despite higher levels of the factors that mediate H2B mono-ubiquitylation at these gene regions (Kim et al. 2009; Jung et al. 2012). Thus, in addition to ubiquitylation of H2B, active de-ubiquitylation of ubH2B by DUBs plays an important role in regulating steady-state levels of ubH2B over gene regions. Intriguingly, there is an enrichment of ubH2B at the exon–intron boundary of highly expressed genes in humans, suggesting that this modification could play a role in alternative splicing (Jung et al. 2012). Supporting this hypothesis, ubH2B is enriched on introns in S. cerevisiae and on skipped exons in mammals, indicating that its presence might be a negative determinant of exon selection (Shieh et al. 2011). The spatial and temporal distribution of ubH2B throughout gene regions during transcription activation indicates that H2B mono-ubiquitylation is highly regulated and could regulate multiple functions during the transcription cycle. Indeed, recent studies on the role of ubH2B in nucleosome organization suggest that ubH2B plays distinct roles at the 5′ end and on the transcribed regions of actively expressed genes.
6.5.1.2 H2B Mono-ubiquitylation Is Required for Transcription Activation at Specific Genes
At a subset of genes, H2B mono-ubiquitylation is required for efficient transcription activation. Mono-ubiquitylation of histone H2B is required for activation of transcription at the inducible GAL1 and SUC2 genes in S. cerevisiae and at the retinoic acid-activated RARβ2 gene in mammalian cells (Pavri et al. 2006; Henry et al. 2003). Notably, at some of these genes, the removal of ubiquitin from ubH2B is also important for transcription activation. Thus, the DUB Ubp8 and its orthologs are required for high expression of specific subsets of genes in yeast, Drosophila, and humans (Henry et al. 2003; Weake et al. 2008; Zhang et al. 2008; Zhao et al. 2008). Hence, both the addition and removal of ubiquitin from histone H2B are necessary for transcription activation at specific genes. A role for ubH2B in transcription activation might not be limited to the specific ubiquitylated site on histone H2B, Lys120. MSL2-mediated ubiquitylation of H2B Lys34 activates transcription cooperatively with the MOF acetyltransferase at the HOXA9 and MEIS1 loci in human cells (Wu et al. 2011). Moreover, the Drosophila ortholog of MSL2 that regulates dosage compensation of the male X chromosome also exhibits in vitro ubiquitylation activity on mammalian histone H2B Lys34 (Wu et al. 2011). Thus, it is possible that mono-ubiquitylation of the equivalent lysine in Drosophila histone H2B, Lys31, promotes the twofold upregulation in expression of X-linked genes in males (Wu et al. 2011).
6.5.1.3 Transcription Activators Recruit the E3 Ligases that Deposit ubH2B
Supporting a role for H2B mono-ubiquitylation in transcription activation at specific genes, the E3 ubiquitin-protein isopeptide ligase for ubH2B, Bre1, interacts directly with transcription activators such as Gal4 in S. cerevisiae and p53 in mammalian cells (Wood et al. 2003a, b; Hwang et al. 2003; Kao et al. 2004). Bre1 is recruited to its target promoters upon gene activation (Wood et al. 2003a, b; Hwang et al. 2003; Kao et al. 2004). Once bound at its target promoter, Bre1 then recruits the E2 conjugase Rad6 (Wood et al. 2003a, b; Kao et al. 2004). The third subunit in the Rad6/Bre1 complex, Lge1, stimulates the activator-dependent recruitment of Bre1 to promoters (Song and Ahn 2010). In mammalian cells, the RNF20/40-interacting protein WAC binds directly to RNA polymerase II targeting the E3 ligases to regions of active transcription (Zhang and Yu 2011). However, the recruitment of Bre1 and Rad6 in yeast is not sufficient for mono-ubiquitylation of histone H2B (Fig. 6.3a). Instead, additional factors are necessary for the catalytic activity of Rad6/Bre1 at gene promoters and on transcribed regions [reviewed in Osley (2006), Weake and Workman (2008)].

Early steps in transcription are required for H2B mono-ubiquitylation. (a) The yeast H2B ubiquitylating enzymes, Rad6, Bre1, and Lge1, are recruited to target promoters through the interaction of Bre1 with activators. However, recruitment of the H2B ubiquitylating enzymes is not sufficient for efficient H2B mono-ubiquitylation; tss: transcription start site. (b) Instead, efficient H2B mono-ubiquitylation requires the presence of factors that regulate the initial stages of the transcription cycle such as the PAF complex, the BUR complex, and the elongating form of RNA polymerase II that has been phosphorylated on Ser5 of the carboxy-terminal domain by Kin28. Phosphorylation of Rad6 at Ser120 by the BUR complex might stimulate its E2 ubiquitin conjugase activity against histone H2B and other targets. (c) Low levels of H2B mono-ubiquitylation at promoters are required for the efficient di- and tri-methylation of histone H3 at Lys4 by the Set1 methyltransferase within the COMPASS complex. In addition to ubH2B, Rad6 and Bre1 mono-ubiquitylate the Swd2 subunit of COMPASS at Lys68 and Lys69, and this ubiquitylation is promoted by prior H2B mono-ubiquitylation. Swd2 mono-ubiquitylation then stimulates the interaction of Spp1 with COMPASS, enabling di- and tri-methylation activity of Set1 toward histone H3 at Lys4. The maintenance of high ubH2B levels is not required for methylation of histone H3 at Lys4 following the initial recruitment/interaction of Spp1 with COMPASS at promoters
6.5.1.4 Early Steps in Transcription Initiation and Elongation Are Required for Mono-ubiquitylation of Histone H2B
Genetic screens in S. cerevisiae have identified a number of factors that are involved in the initial stages of transcription initiation and elongation as being important for H2B mono-ubiquitylation. These include components of the Polymerase-Associated Factor (PAF) complex, the Bur1/Bur2 (BUR) cyclin-dependent protein kinase complex, and the phosphorylation status of RNA polymerase II itself (Fig. 6.3b). The PAF complex associates with the initiating and elongating forms of RNA polymerase II and regulates a number of transcription-associated processes [reviewed in Jaehning (2010)]. Mutations in PAF subunits such as Rtf1 and Paf1 result in a loss of ubH2B (Wood et al. 2003b; Ng et al. 2003). In PAF mutants, Rad6 is recruited to promoters but does not catalyze mono-ubiquitylation of histone H2B (Wood et al. 2003b; Xiao et al. 2005). Moreover, both yeast and human Bre1 directly interact with purified PAF complex in vitro through the Paf1 subunit (Kim et al. 2009; Kim and Roeder 2009). Thus, the PAF complex is required for the ubiquitylation activity of Rad6 and Bre1, but not for their recruitment to gene promoters. In addition to the PAF complex, the BUR kinase complex is also important for mono-ubiquitylation of histone H2B (Wood et al. 2005; Laribee et al. 2005). Similar to PAF mutants, deletion of bur2 reduces ubH2B levels, but does not affect recruitment of Rad6 (Wood et al. 2005; Laribee et al. 2005). However, loss of Bur2 does reduce recruitment of the PAF complex to gene promoters (Wood et al. 2005; Laribee et al. 2005). Does the BUR complex affect ubH2B levels via its effect on PAF complex recruitment or through additional PAF-independent mechanisms? Studies in S. cerevisiae have shown that the BUR complex directly phosphorylates Ser120 of Rad6 in vitro and that mutation of this residue reduces ubH2B levels without affecting localization of Rad6 to chromatin (Wood et al. 2005). Furthermore, phosphorylation of human UBE2A (HHR6A/RAD6A) by CDK2 stimulates its in vitro E2 ubiquitin conjugase activity (Sarcevic et al. 2002). Thus the BUR complex might affect ubH2B levels by two distinct mechanisms: directly via phospho-regulation of Rad6 activity and indirectly through its effect on PAF complex recruitment.
In addition to the PAF and BUR complexes, events that occur during transcription by RNA polymerase II, as well as transcription itself, are important for efficient mono-ubiquitylation of histone H2B. During in vitro transcription on a chromatin template, the appearance of ubH2B is dependent on the addition of nucleotide triphosphates (Pavri et al. 2006). Furthermore, PAF-mediated transcription is required for efficient H2B mono-ubiquitylation in vitro (Kim et al. 2009). Although these in vitro transcription studies suggest that the majority of the effect of the PAF complex on ubH2B levels results from its stimulation of transcription, low levels of PAF-independent transcription are not sufficient for ubH2B deposition (Kim et al. 2009). Thus the PAF complex also has transcription-independent roles in regulating H2B mono-ubiquitylation.
During the early stages of the transcription cycle, a series of events take place in which the carboxy-terminal heptapeptide repeat sequences (also known as the carboxy-terminal domain, CTD) on the largest subunit of RNA polymerase II become sequentially phosphorylated [reviewed in Buratowski (2009)]. These CTD phosphorylations provide binding sites for a number of factors involved in regulating the release of RNA polymerase II from the promoter and its efficient transition into transcription elongation. First, Kin28 (CDK7 in humans) within the general transcription factor TFIIH phosphorylates the CTD at the Ser5 position. This coincides with the introduction of a single-stranded DNA template into the active site of RNA polymerase II and its subsequent dissociation from the general transcription factors. Second, Ctk1 (P-TEFb/CDK9 and CDK12 in humans) phosphorylates the CTD at Ser2. In S. cerevisiae, Ser5 phosphorylation by Kin28, but not Ser2 phosphorylation by Ctk1, is required for ubH2B (Xiao et al. 2005). Thus, the first phosphorylation of the CTD at Ser5, which occurs during the initial stages of transcription by RNA polymerase II, is required for efficient histone H2B mono-ubiquitylation. Together, these results from both yeast and in vitro studies show that the beginning stages of transcription initiation, which involve the PAF and BUR complexes, play important roles in regulating H2B mono-ubiquitylation (Fig. 6.3b).
6.5.1.5 Mono-ubiquitylation of Histone H2B Is a Prerequisite for H3 Lys4 Methylation
Genetic screens for mutations affecting histone methylation in S. cerevisiae have identified a unidirectional histone cross talk between ubH2B and methylation of histone H3 on Lys4, an important modification for transcription activation [reviewed in Shukla et al. (2009)]. Mutations that disrupt the enzymes required for H2B mono-ubiquitylation, Rad6 and Bre1, or point mutations in histone H2B itself (H2B-K123R) reduce the global level of di- and tri-methylated histone H3 Lys4 (Sun and Allis 2002; Dover et al. 2002; Shahbazian et al. 2005; Schneider et al. 2005; Dehe et al. 2005). However, deletion of the H3 Lys4 methyltransferase Set1 or point mutations in Lys4 of histone H3 (H3-K4R) do not affect ubH2B levels (Sun and Allis 2002). Thus, whereas ubH2B is necessary for histone H3 Lys4 methylation, methylation of H3 Lys4 is not required for H2B mono-ubiquitylation. Notably, ubH2B is only necessary for di- and tri-methylation of histone H3 Lys4 and does not affect mono-methylation (Shahbazian et al. 2005; Schneider et al. 2005; Dehe et al. 2005).
A recent genetic screen in S. cerevisiae for factors influencing ubH2B levels has demonstrated that this histone cross talk might not be as clear-cut as the original studies had suggested (Lee et al. 2012). This genetic screen identified the chromatin remodeler Chd1 as a novel factor required for maintaining high levels of ubH2B (Lee et al. 2012). In contrast to all other factors identified thus far that affect ubH2B levels, loss of chd1 does not affect H3 Lys4 methylation (Lee et al. 2012). Residual low levels of ubH2B remain in the chd1∆ strain, and it appears likely that these low levels of ubH2B are sufficient for wild-type level of H3 Lys4 methylation (Lee et al. 2012). This conclusion is supported by the observation that in cultured mouse myoblasts, ubH2B is reduced during myogenic differentiation without a corresponding decrease in H3 Lys4 methylation (Vethantham et al. 2012).
Despite these recent findings, it is clear that at least a low level of initial deposition of ubH2B is important for H3 Lys4 methylation. How might the initial deposition of ubH2B at 5´ gene regions regulate H3 Lys4 methylation? H3 Lys4 methylation is catalyzed by the methyltransferase Set1 in S. cerevisiae that resides within Complex Proteins Associated with Set1 (COMPASS) [reviewed in Malik and Bhaumik (2010)]. Studies using in vitro transcription reactions have shown that the di- and tri-methyltransferase activity of human Set1 is directly stimulated by ubH2B (Kim et al. 2009). In S. cerevisiae, deletion of the Spp1 (Cps40) subunit of COMPASS eliminates di- and tri-methylation of H3 Lys4 (Dehe et al. 2006; Morillon et al. 2005; Shi et al. 2007). In addition, loss of the ubiquitin conjugase Rad6, or loss of ubH2B, weakens the association of another COMPASS subunit, Swd2 (Cps35), with both chromatin and the COMPASS complex (Lee et al. 2007). Could these two subunits within COMPASS regulate the cross talk between ubH2B and Set1 di- and tri-methylation activity? This indeed appears to be the case: COMPASS purified from rad6∆ yeast lacks di- and tri-methylation activity (Lee et al. 2007). Notably, ubH2B promotes mono-ubiquitylation of Swd2 at Lys68 and Lys69 by Rad6 and Bre1 (Vitaliano-Prunier et al. 2008). This Swd2 mono-ubiquitylation regulates recruitment of the Spp1 subunit of COMPASS that regulates di- and tri-methylation activity of Set1 (Vitaliano-Prunier et al. 2008). Thus, these findings collectively suggest a model in which initial mono-ubiquitylation of histone H2B by activator-recruited Rad6 and Bre1 at 5′ gene regions regulates ubiquitylation of the Swd2 subunit within COMPASS. This Swd2 ubiquitylation is necessary for efficient interaction of the Spp1 subunit of COMPASS that regulates di- and tri-methylation activity of Set1 with chromatin (Vitaliano-Prunier et al. 2008) (Fig. 6.3c). It appears likely that the subsequent maintenance of high ubH2B levels is not required for COMPASS activity following this initial recruitment of Swd2.
Strikingly, the cross talk between ubH2B and H3 Lys4 methylation is not restricted to H2B mono-ubiquitylated at Lys123 (Lys120 in humans). H2B-Lys34 mono-ubiquitylation also promotes histone H3 Lys4 di- and tri-methylation, both in vitro and in human cells (Wu et al. 2011). However, H2B-Lys34 mono-ubiquitylation also promotes RNF20/40 recruitment to chromatin, thus increasing local ubH2B-Lys120 levels, and could therefore affect H3 Lys4 methylation indirectly (Wu et al. 2011).
6.5.1.6 Mono-ubiquitylation of Histone H2B Signals for H3 Lys79 Methylation
Similar to the cross talk between ubH2B and H3 Lys4 methylation, mono-ubiquitylation of histone H2B is also a prerequisite for H3 Lys79 methylation [reviewed in Shukla et al. (2009)]. In human cells, ubH2B and H3 Lys79 are both enriched on the 5′ to middle regions of actively transcribed genes (Jung et al. 2012). In S. cerevisiae H3 Lys79 methylation is catalyzed by the methyltransferase Dot1 (Ng et al. 2002; van Leeuwen et al. 2002). Dot1-mediated di- and tri-methylation, but not mono-methylation, of H3 Lys79 requires prior ubH2B (Shahbazian et al. 2005; Briggs et al. 2002). In human cells, mono-ubiquitylation of histone H2B at both Lys120 and Lys34 is required for efficient H3 Lys79 methylation (Wu et al. 2011; Shahbazian et al. 2005; Briggs et al. 2002). However, as observed for the cross talk between ubH2B and H3 Lys4 methylation, low levels of ubH2B are sufficient for H3 Lys79 methylation, and the levels of this methylated histone residue remain constant in chd1∆ yeast that have reduced ubH2B levels (Lee et al. 2012). Furthermore, Swd2 might also mediate the cross talk between ubH2B and H3 Lys79 methylation. In addition to COMPASS, Swd2 is present in other protein complexes in S. cerevisiae and physically interacts with Dot1 in co-immunoprecipitation studies (Lee et al. 2007). Thus, the mechanism for establishing the cross talk between ubH2B and H3 Lys79 methylation might also utilize Swd2 mono-ubiquitylation in an unknown manner.
6.5.1.7 Methylation of the Kinetochore Protein Dam1 Is Regulated by Cross Talk with ubH2B
Notably, the cross talk between ubH2B and lysine methylation is not restricted to histone substrates (Latham et al. 2011). Set1 di-methylates the kinetochore protein Dam1 at Lys233, in addition to histone H3 Lys4 (Zhang et al. 2005). Dam1 is a component of the ten-subunit DASH complex that anchors kinetochores to microtubules and is important for the correct attachment of sister chromatids to opposing spindle poles (biorientation) during mitosis (Zhang et al. 2005). Phosphorylation of Dam1 by the Ipl1 kinase has been proposed to reduce its affinity for microtubules to enable the dissociation of faulty kinetochore–microtubule interactions [reviewed in Smolle and Workman (2011)]. Di-methylation of Dam1 at Lys233 by Set1 appears to inhibit Ipl1-mediated phosphorylation of Dam1 at Ser235 (Zhang et al. 2005). Thus, Dam1 methylation could stabilize kinetochore–microtubule interactions once proper biorientation is achieved by inhibiting Ipl1-mediated phosphorylation (Latham et al. 2011; Smolle and Workman 2011). Similar to the cross talk between ubH2B and H3 Lys4 tri-methylation, Rad6- and Bre1-mediated mono-ubiquitylation of H2B at the centromere is required for Set1 to methylate Dam1 (Latham et al. 2011). Furthermore, the PAF complex is also required for Dam1 methylation, but this is independent of active transcription and does not require Kin28 (Latham et al. 2011). Intriguingly, de-ubiquitylation of ubH2B by Ubp8, but not by Ubp10, limits Dam1 methylation because methylated Dam1 levels increase in ubp8Δ yeast (Latham et al. 2011). Overall, these findings demonstrate that in addition to histone H3, prior mono-ubiquitylation of histone H2B is a prerequisite for methylation of some non-histone substrates. Furthermore, the cross talk between ubH2B and non-histone substrates can occur independently of active transcription.
6.5.1.8 De-ubiquitylation of ubH2B Is Required for Efficient Transcription
In addition to the deposition of ubH2B, the removal of ubiquitin from ubH2B is required for efficient transcription at a subset of genes. Thus, there is a sequential addition and removal of ubiquitin from histone H2B that plays an important role in facilitating transcription activation at specific genes. This cycle of H2B mono-ubiquitylation and de-ubiquitylation is best exemplified by the GAL1 gene in S. cerevisiae. Full transcription activation at GAL1 requires both an intact Lys123 on histone H2B, mono-ubiquitylation by Rad6/Bre1, and the DUB Ubp8 (Henry et al. 2003). How might this cycle of histone H2B mono-ubiquitylation function during transcription activation?
The removal of ubiquitin from histone H2B has been shown to regulate the phosphorylation status of the RNA polymerase II CTD and the transition from transcription initiation to elongation at a subset of genes in S. cerevisiae (Fig. 6.4a). Whereas the first phosphorylation of the RNA polymerase II CTD on Ser5 is required for Rad6/Bre1 to ubiquitylate H2B, the removal of ubiquitin from ubH2B by Ubp8 appears to be important for the second CTD phosphorylation at Ser2 by Ctk1 (Xiao et al. 2005; Wyce et al. 2007). Recruitment of Ctk1 and Ser2-phosphorylated RNA polymerase II to the coding regions of the GAL1, ADH1, and PMA1 genes is reduced in ubp8∆ yeast (Wyce et al. 2007) (Fig. 6.4b). Moreover, localization of Ctk1 is restored in ubp8∆, bre1∆ yeast, indicating that the de-ubiquitylated form of histone H2B is required for efficient Ctk1 recruitment (Wyce et al. 2007) (Fig. 6.4c). Thus, de-ubiquitylation of ubH2B is important for the transition of RNA polymerase II into efficient transcription elongation. These studies illustrate that mono-ubiquitylation and de-ubiquitylation of histone H2B are both required at different stages within the transcription cycle for optimal gene expression. However, it is unclear whether this cycle of H2B ubiquitylation is generally utilized during transcription activation at all genes or is specific to particular types of genes that are highly induced in response to stimuli such as GAL1.

De-ubiquitylation of ubH2B regulates early steps in transcription elongation. (a) Ubp8, the DUB within the yeast SAGA transcriptional coactivator, removes ubiquitin from ubH2B. De-ubiquitylation of ubH2B facilitates recruitment and/or activity of the Ctk1 kinase, which then phosphorylates Ser2 of the carboxy-terminal domain of RNA polymerase II (Pol II); tss: transcription start site. (b) In the absence of Ubp8, persistent ubH2B inhibits the recruitment and/or activity of the Ctk1 kinase against Pol II. (c) Loss of the E3 ubiquitin ligase, Bre1, rescues Ctk1 activity in the absence of Ubp8
6.5.1.9 Mono-ubiquitylation of Histone H2B on Coding Regions Requires Factors Involved in Transcription Elongation
Although ubH2B at promoters is important for transcription activation, the highest levels of ubH2B are observed on the transcribed regions of expressed genes (Minsky et al. 2008). How is ubH2B deposited during transcription elongation? In S. cerevisiae, Rad6 associates with the elongating form of RNA polymerase II (Xiao et al. 2005). Thus, mono-ubiquitylation of histone H2B occurs as RNA polymerase II transcribes the body of the gene. Notably, mutations in factors that regulate transcription elongation, such as the PAF complex (rtf1∆), disrupt the association of Rad6 with RNA polymerase II preventing it from moving into the transcribed region of the gene (Xiao et al. 2005). Thus, the global decrease in ubH2B observed upon loss of the PAF complex might result largely from the disruption of the interaction between Rad6 and the elongating RNA polymerase II. Although the role of ubH2B at promoter regions has been well studied, the majority of ubH2B is present on the transcribed regions of genes rather than at promoters. What, therefore, is the role of ubH2B on the transcribed regions of genes?
6.5.1.10 Nucleosome Occupancy and/or Stability Is Regulated by ubH2B
Several recent studies have provided insight into the role of ubH2B on the transcribed regions of genes. These studies examined the role of ubH2B in regulating nucleosomal organization using a combination of in vitro assays and high-resolution micrococcal nuclease chromatin immunoprecipitation and sequencing (Batta et al. 2011; Chandrasekharan et al. 2009). In general, ubH2B promotes nucleosome stability and/or occupancy (Batta et al. 2011; Chandrasekharan et al. 2009; Davies and Lindsey 1994). Individual nucleosomes that contain ubH2B are more resistant to micrococcal nuclease digestion and salt extraction, consistent with increased stability of the ubH2B-containing nucleosome (Chandrasekharan et al. 2009). In addition, in vitro-reconstituted ubH2B-containing nucleosomes exhibit enhanced resistance to DNase I digestion when compared to nucleosomes containing unmodified H2B (Davies and Lindsey 1994). These in vitro observations are supported by results from an examination of genome-wide nucleosome organization in H2B-K123A and wild-type S. cerevisiae using high-resolution micrococcal nuclease chromatin immunoprecipitation and sequencing (Batta et al. 2011). The H2B-K123A mutant shows a genome-wide decrease in nucleosome occupancy when compared to the wild type that is most apparent on transcribed regions downstream from the +1 nucleosome position (Batta et al. 2011). Furthermore, this decrease in nucleosome occupancy is also observed upon deletion of the enzymes that catalyze H2B mono-ubiquitylation: Rad6 and its regulator Lge1 (Batta et al. 2011). It is possible that stabilization in the body of genes may protect against disruption by the passage of the elongating RNA polymerase II.
Does the effect of ubH2B on nucleosome occupancy or stability depend upon its cross talk with histone methylation? Surprisingly, although ubH2B regulates methylation of histone H3 at Lys4, Lys79, and Lys36, the effects of ubH2B on nucleosome occupancy occur independently of these H3 methylations (Batta et al. 2011). Consistent with this observation, chd1∆ yeast have reduced ubH2B levels and a genome-wide decrease in nucleosome occupancy in transcribed regions downstream from the +1 nucleosome that is independent of changes in H3 Lys4 or H3 Lys79 methylation (Lee et al. 2012).
Intriguingly, whereas ubH2B promotes stability and/or occupancy of individual nucleosomes, ubH2B is disruptive to higher order chromatin structure (Fierz et al. 2011). Analysis of the conformation and accessibility of chromatin fibers containing chemically synthesized ubH2B shows that ubH2B impairs compaction of the chromatin fiber (Fierz et al. 2011). Models of the chromatin fiber show that nucleosome stacking is unlikely to occur in the presence of ubH2B (Fierz et al. 2011). These in vitro observations are supported by the finding that the A. thaliana ubH2B DUB, SUP32, is required for heterochromatin spreading and DNA methylation (Sridhar et al. 2007). Furthermore, ubH2B also restricts heterochromatin spreading into the chicken β-globin gene cluster (Ma et al. 2011). Thus, the association of ubH2B with actively transcribed genes might promote the euchromatic state of these genes by enhancing the accessibility of the chromatin fiber. However, at the single nucleosome level, ubH2B promotes nucleosome stability and/or occupancy.
6.5.1.11 ubH2B-Containing Nucleosomes Affect Transcription Differentially at Promoters and on Coding Regions
What is the effect of the ubH2B-mediated enhancement of nucleosome stability and/or occupancy on transcription? Although ubH2B is generally associated with active transcription, recent studies using the H2B-K123A and chd1∆ yeast strains have shown that ubH2B has both activating and repressing functions with regard to expression of individual genes (Lee et al. 2012; Batta et al. 2011). These studies demonstrate that ubH2B has an activating function at highly expressed genes, but has a repressive function at lowly expressed genes (Lee et al. 2012; Batta et al. 2011).
How can ubH2B have both activating and repressive roles at different genes? This observation can be explained by a model in which the ubH2B-mediated increase in nucleosome stability/occupancy has different effects on transcription at promoters and on transcribed regions (Batta et al. 2011; Chandrasekharan et al. 2009; Davies and Lindsey 1994) (Fig. 6.5). These effects on transcription are independent of the role of ubH2B in regulating the events that occur during transcription activation such as H3 Lys4 tri-methylation. Instead, in this model, ubH2B promotes nucleosome assembly on transcribed regions following passage of RNA polymerase II (Lee et al. 2012; Batta et al. 2011). Supporting this model, ubH2B is required for the efficient reassembly of nucleosomes during transcription elongation in S. cerevisiae (Fleming et al. 2008). Furthermore, this efficient nucleosome reassembly also requires the Facilitates Chromatin Transcription (FACT) histone chaperone composed of Spt16 and SSRP1 (Fleming et al. 2008). Moreover, FACT activity is stimulated by ubH2B in vitro (Pavri et al. 2006). Proper nucleosome occupancy and positioning might be important to enable RNA polymerase II to clear the promoter region and begin elongation, especially during successive rounds of transcription. Thus, enhanced nucleosome stability and/or occupancy would promote transcription elongation by RNA polymerase II. Consistent with this model, loss of ubH2B in the H2B-K123A mutant results in decreased levels of RNA polymerase II on the transcribed region of highly expressed genes (Batta et al. 2011).

H2B mono-ubiquitylation regulates transcription differentially at promoters and on coding regions. (a) At lowly expressed genes, ubH2B enhances nucleosome stability at promoters and inhibits recruitment of RNA polymerase II (Pol II) and transcription initiation; tss: transcription start site. (b) At highly expressed genes, ubH2B promotes nucleosome stability and/or assembly on coding regions following the passage of Pol II. This enhances transcription elongation by Pol II and thus promotes gene expression
At lowly expressed genes, ubH2B appears to play a repressive role. Indeed, loss of ubH2B in the H2B-K123A mutant increases levels of RNA polymerase II bound at the promoter of lowly expressed genes (Batta et al. 2011). Under normal conditions, these lowly expressed genes lack RNA polymerase II at their promoters. Thus, in the proposed model, the loss of ubH2B from promoter nucleosomes facilitates recruitment of RNA polymerase II and the subsequent transcription initiation at these lowly expressed genes (Batta et al. 2011). Supporting this model, ubH2B-mediated nucleosome stabilization suppresses transcription initiation from cryptic weak promoters within coding regions (Chandrasekharan et al. 2009; Fleming et al. 2008). Hence ubH2B stabilization of nucleosomes at promoters is primarily repressive at lowly expressed genes or at cryptic promoters because it prevents recruitment of RNA polymerase II and transcription initiation. In contrast, ubH2B stabilization of nucleosomes is activating on the transcribed regions of highly expressed genes where it promotes reassembly of nucleosomes following passage of RNA polymerase II, thus enabling successive rounds of transcription to efficiently occur.
6.5.1.12 Mono-ubiquitylation of Histone H2B Plays Multiple Roles in Transcriptional Regulation
Whereas ubH2B is most commonly associated with transcriptional activation, the evidence discussed previously in Sects. 6.5.1.1 – 6.5.1.11 demonstrates that ubH2B can have both activating and repressive roles in regulating gene expression. In summary, although ubiquitylation of H2B occurs at promoters during transcription activation, high levels of ubH2B are not observed at the promoters of actively expressed genes. Instead, the highest levels of ubH2B are found on the transcribed region of actively expressed genes. This is because there is a sequential addition and removal of ubH2B that plays an important role in regulating the initial stages of the transcription cycle that include tri-methylation of histone H3 Lys4 and the recruitment of kinases that phosphorylate the RNA polymerase II CTD. However, the formation and/or removal of ubH2B might not be essential for high levels of expression at all genes. Instead, the greatest effects of ubH2B on gene expression result from its role in promoting nucleosome stability and/or occupancy. This promotion of nucleosome stability and/or occupancy by ubH2B has repressive effects at genes with weak promoters, where it prevents recruitment of RNA polymerase II. However, at highly expressed genes, ubH2B promotes nucleosome occupancy following passage of the elongating RNA polymerase II and has an activating effect on transcription. Thus, ubH2B has multiple functions in regulating transcription events and gene expression at both promoters and on transcribed regions.
6.5.2 Transcriptional Repression by ubH2A
While ubH2B is associated predominantly with actively expressed genes, H2A mono-ubiquitylation is generally regarded as being repressive to gene expression. Supporting this statement, two of the E3 ubiquitin ligases specific for ubH2A at Lys119, RNF2 and DZIP3, are subunits of complexes that repress gene expression. Furthermore, ubH2A is found at silenced regions of the genome such as the heterochromatin in DNA satellite repeats and the inactive X chromosome (de Napoles et al. 2004; Fang et al. 2004). However, recent studies have provided evidence that ubH2A, like ubH2B, might play a more complicated role in regulating gene expression. Similar to findings that both mono-ubiquitylation and de-ubiquitylation of histone H2B are required for transcription activation at specific genes, both the addition and removal of ubiquitin from histone H2A are required for transcriptional repression at some Polycomb-target genes.
6.5.2.1 Polycomb Complex-Associated Repression by ubH2A
The first ubH2A-specific E3 ligase to be identified, RNF2, is a subunit of the PRC1 class complexes (Wang et al. 2004; Cao et al. 2005; Kerppola 2009). This observation provided the first clue that H2A ubiquitylation at Lys119 is a repressive mark because PRC1 class complexes have a well-characterized role in transcriptional repression (Wang et al. 2004; Cao et al. 2005; Kerppola 2009). RNF2 in mammals and its ortholog Sce in flies are required for ubiquitylation of H2A and transcriptional repression at the promoters of Polycomb-target genes such as the Hox cluster (Gutierrez et al. 2012; Wang et al. 2004; Cao et al. 2005; Wei et al. 2006). In addition, the Arabidopsis RNF2 and Bmi1 orthologs repress genes that regulate embryonic development (Bratzel et al. 2010; Xu and Shen 2008). Moreover, RNF2 is also a subunit of the PRC1-related E2F-6.com-1 repressive complex. The E2F-6.com-1 complex represses E2F- and Myc-responsive genes in quiescent cells (Ogawa et al. 2002; Sanchez et al. 2007). Thus, mono-ubiquitylation of H2A by the E3 ligase subunits of the Polycomb-related transcriptional repressor complexes correlates with transcriptional silencing in plants, flies, and mammals.
The second hint as to a repressive role for ubH2A in gene expression came from studies of X inactivation in mice in which PRC1 and ubH2A were shown to colocalize on the inactive X chromosome (de Napoles et al. 2004; Fang et al. 2004). RNAi-mediated knockdown of RNF2 and Ring1 depletes ubH2A from the inactive X chromosome, suggesting that this mark is added by the PRC1 class complexes (de Napoles et al. 2004; Fang et al. 2004). Notably, RNF2 also ubiquitylates the histone variant H2A.Z on the inactive X chromosome in female cells, and unmodified H2A.Z is excluded from this chromosome (Sarcinella et al. 2007).
Thus, H2A ubiquitylation is associated with transcriptional silencing by PRC1-related complexes both at individual gene targets and on the inactive X chromosome. Notably, the correlation between ubH2A and transcriptional silencing is not limited to RNF2-mediated ubiquitylation. Brca1 mutant mice have a reduced number of condensed DNA regions in the genome and also show loss of ubH2A at regions that are usually silenced, such as DNA satellite repeats (Zhu et al. 2011). These observations suggest that the presence of ubH2A generally correlates with transcriptional repression and silenced chromatin.
Does H2A mono-ubiquitylation directly repress transcription? Consistent with a direct role for ubH2A in repressing transcription, RNF2/Ring1-deficient cells have decreased ubH2A levels and de-repression of Polycomb-target genes (Wang et al. 2004; Cao et al. 2005). In addition, Drosophila Sce is required for repression at a subset of Polycomb-target genes that include the Hox genes (Gutierrez et al. 2012) (Fig. 6.6a). Furthermore, the USP16 (Ubp-M) DUB antagonizes Polycomb repression at the HoxD10 gene by downregulating ubH2A levels (Joo et al. 2007). Overexpression of another ubH2A DUB, USP21, results in reduced ubH2A levels and increased expression of the Serpina6 gene (Nakagawa et al. 2008). More direct evidence is provided by the observation that expression of histone H2A fused to ubiquitin restores silencing of DNA satellite repeats in brca1-deficient human cell lines (Zhu et al. 2011). Together, these observations are consistent with a direct role for ubH2A in repressing gene expression.

H2A mono-ubiquitylation and de-ubiquitylation are required for repression of PRC1-target genes in Drosophila. (a) Mono-ubiquitylation of histone H2A by the E3 ubiquitin ligase Sce/dRing within the Drosophila PRC1 complex is associated with repression of expression of Polycomb-target genes such as Ultrabithorax (Ubx). (b) De-ubiquitylation of ubH2A by Calypso within the PR-DUB complex is required for repression of expression of the Polycomb-target gene Ubx
However, the recent identification of the PR-DUB complex in Drosophila poses a problem with regard to the mechanism through which ubH2A represses gene expression. Prior to the identification of PR-DUB in Drosophila, studies examining the role of ubH2A were consistent with a model in which ubH2A represses transcription, and de-ubiquitylation of ubH2A facilitates transcription activation. However, in flies the ubH2A de-ubiquitylating enzyme PR-DUB is associated with transcriptional repression rather than activation (Scheuermann et al. 2010) (Fig. 6.6b). The identification of a ubH2A-DUB that represses, rather than activates, transcription poses several problems for the current models of how ubH2A represses gene expression. These problems are unresolved and provide an important area for future research.
What is the evidence supporting a role for the PR-DUB in transcription repression? PR-DUB is bound at the Polycomb response element of the Hox gene Ultrabithorax (Ubx) both in cells where Ubx is repressed and in cells where it is expressed (Scheuermann et al. 2010). However, the catalytic activity of the DUB within PR-DUB, Calypso, is required for repression of Ubx in larval imaginal discs (Scheuermann et al. 2010). Furthermore, simultaneous loss of both the E3 ligase Sce and the DUB Calypso results in a more rapid and severe loss of HOX gene repression than loss of either individual activity alone (Gutierrez et al. 2012; Scheuermann et al. 2010). Thus, both the mono-ubiquitylation and de-ubiquitylation of H2A are required for full transcriptional repression, at least at a subset of Polycomb-target genes in Drosophila. These recent findings indicate that further study is necessary to elucidate the mechanism of transcriptional silencing by ubH2A, especially with regard to PRC1-target genes.
6.5.2.2 The 2A-HUB and 2A-DUB Complexes Function Antagonistically to Regulate Gene Expression
Although the mechanism of ubH2A-mediated transcriptional silencing might vary by genomic region and method of deposition, some specific examples of the mechanism of ubH2A-mediated repression have been revealed for certain target loci. For example, one mechanism through which ubH2A might repress transcription is via the inhibition of transcription elongation by RNA polymerase II.
The 2A-HUB complex containing the DZIP3 H2A E3 ligase represses transcription at a subset of chemokine genes in mammalian cells (Zhou et al. 2008). In addition to DZIP3, this 2A-HUB complex contains the nuclear co-receptor N-CoR and the histone deacetylases HDAC1 and HDAC3 (Zhou et al. 2008). Knockdown of DZIP3 results in a decrease in ubH2A levels at target promoters, but does not affect levels of RNA polymerase II present at these promoters (Zhou et al. 2008). Instead, knockdown of DZIP3 results in increased levels of FACT and the Ser2-phosphorylated form of RNA polymerase II on the transcribed region of target genes such as RANTES (Zhou et al. 2008). How does loss of DZIP3 and ubH2A promote FACT recruitment? The Spt16 subunit of FACT preferentially interacts with histone H2A, but not ubH2A, in pull-downs from cultured cells (Zhou et al. 2008). Thus, H2A mono-ubiquitylation by the 2A-HUB complex inhibits recruitment of the Spt16 subunit of FACT, decreasing transcription elongation by RNA polymerase II (Zhou et al. 2008) (Fig. 6.7a).

Multiple mechanisms involved in transcriptional repression by H2A mono-ubiquitylation. (a) The human 2A-HUB complex containing the DZIP3 E3 ubiquitin ligase mono-ubiquitylates histone H2A and inhibits recruitment of the Spt16 subunit of FACT, thus repressing transcription elongation by RNA polymerase II (Pol II); tss: transcription start site. (b) Fig. 6.7 (continued) Multiple De-ubiquitylation of ubH2A by MYSM1 within the 2A-DUB complex activates transcription, in part, through enabling recruitment of FACT, which facilitates subsequent transcription elongation by Pol II. (c) The presence of ubH2A at promoters prior to recruitment of Pol II inhibits MLL3-mediated tri-methylation of histone H3 Lys4 and transcription initiation. (d) H2A mono-ubiquitylation enhances association of the linker histone H1 with chromatin. Conversely, nucleosomes that lack ubH2A do not have histone H1 associated with them
Does de-ubiquitylation of ubH2A then activate transcription? Indeed, several ubH2A-specific DUBs are associated with transcriptional activation. Introduction of USP21 into regenerating hepatocytes correlates with increased expression of Serpina6, as determined by immunostaining (Nakagawa et al. 2008). Furthermore, the 2A-DUB complex consisting of the ubH2A DUB, MYSM1, together with the histone acetyltransferase PCAF/KAT2B, is required for transcription activation at androgen-responsive genes in mammalian cells (Zhu et al. 2007). Knockdown of MYSM1 reduces recruitment of RNA polymerase II to promoters and decreases levels of the elongating form of RNA polymerase II on transcribed regions (Zhu et al. 2007). Thus, ubH2A and histone acetylation function antagonistically to regulate transcription repression and activation at a subset of genes in mammalian cells (Fig. 6.7b).
6.5.2.3 Inhibition of H3 Lys4 Methylation by ubH2A
An alternative mechanism of transcriptional repression by ubH2A has been proposed based on the observation that there is a negative cross talk between ubH2A at Lys119 and H3 Lys4 methylation in vitro (Nakagawa et al. 2008; Vissers et al. 2008). When ubH2A is present on reconstituted chromatin templates in an in vitro transcription system, MLL3-mediated di- and tri-methylation of histone H3 Lys4 is inhibited (Nakagawa et al. 2008) (Fig. 6.7c). Furthermore, the presence of ubH2A in this in vitro system also inhibits transcription by RNA polymerase II (Nakagawa et al. 2008). Additionally, substitution of histone H3 with an H3-K4R mutant relieves the inhibition of transcription caused by ubH2A (Nakagawa et al. 2008). These observations suggest a model in which the presence of ubH2A at promoters, prior to recruitment of RNA polymerase II, could inhibit the subsequent tri-methylation of H3 Lys4 that is associated with transcription initiation. Importantly, an effect of ubH2A on transcription elongation was not observed in this study, suggesting that an inhibitory role of ubH2A on transcription using in vitro chromatin templates occurs during the early stages of transcription initiation (Nakagawa et al. 2008).
Despite this in vitro observation, both transcription initiation and elongation could be affected by ubH2A in vivo. For example, the same cytokine genes that are repressed by ubH2A through an inhibition of FACT-mediated transcription elongation are activated by the ubH2A-DUB, USP21 (Nakagawa et al. 2008). Studies on other genes suggest that the MYSM1-containing 2A-DUB activates transcription through, in part, increased recruitment of the initiating form of RNA polymerase II (Zhu et al. 2007). However, at genes in mouse embryonic stem cells that are poised for transcription, but are not being actively transcribed, RNF2, Ring1, and ubH2A are present together with the initiating form of RNA polymerase II (Stock et al. 2007). Furthermore, in addition to ubH2A, both H3 Lys4 and repressive H3 Lys27 methylated nucleosomes are present at the promoters of these bivalent genes (Stock et al. 2007). These observations show that an inhibitory role for ubH2A in regulating H3 Lys4 methylation is not a genome-wide phenomenon in vivo and might be restricted to a subset of gene promoters.
Notably, some of the complexes that contain E3 ubiquitin ligases specific for ubH2A also contain de-methylase subunits with activity toward methylated histone H3. For example, the human FBXL10-BcoR complex and the dRAF complex both contain de-methylases specific for methylated histone H3: KDM2 in Drosophila and KDM2B (FBXL10/JHDM1B) in humans (Lagarou et al. 2008; Gearhart et al. 2006; Sanchez et al. 2007). However, whereas Drosophila KDM2 is specific for di-methylated H3 Lys36, human KDM2B is a nucleolar protein with specificity for tri-methylated H3 Lys4 (Lagarou et al. 2008; Frescas et al. 2007). Thus it is unclear whether the human FBXL10-BcoR complex and the dRAF complex are functionally related. Despite this discrepancy, these observations suggest that histone H3 Lys4 methylation and ubH2A might be co-regulated at specific target loci in vivo because of the presence of E3 ubiquitin ligase and histone de-methylase subunits within common complexes.
6.5.2.4 Higher Order Chromatin Structure Is Regulated by ubH2A
Yet another mechanism through which ubH2A might repress transcription is via the regulation of higher order chromatin structure. Structural studies of the nucleosome show that the carboxy-terminal tail of histone H2A containing the ubiquitylated lysine residue is able to contact the linker histone H1 (Luger et al. 1997). Furthermore, the presence of ubH2A enhances binding of the linker histone H1 to reconstituted nucleosomes in vitro (Jason et al. 2005). Thus, ubH2A-containing nucleosomes might be more compacted in chromatin due to enhanced binding of the linker histone H1 (Fig. 6.7d). These in vitro observations are supported by the in vivo finding that purified mono-nucleosomes from H2A-K119R cells lack histone H1 (Zhu et al. 2007). The effects of ubH2A in regulating higher order chromatin structure might play an important role in repression at large silenced genomic regions such as DNA satellite repeats and the inactive X chromosome.
6.5.2.5 Multiple Mechanisms of Transcription Repression by ubH2A
In Sects. 6.5.2.1 – 6.5.2.4 , we have described evidence to support a role for mono-ubiquitylation of histone H2A in transcriptional repression. We have summarized studies showing that the mechanism of repression by ubH2A appears to differ at individual gene loci. Notably, the enzymes that ubiquitylate H2A are frequently found in transcription repressor complexes. Some evidence suggests that ubH2A directly inhibits transcription by restricting the recruitment of, or elongation by, RNA polymerase II. The repressive effect of ubH2A on transcription elongation could result in part from an inhibitory action toward the FACT histone chaperone. At promoters, ubH2A has been shown to interfere with other histone modifications that correlate with active transcription such as histone H3 Lys4 tri-methylation. Lastly, ubH2A regulates higher order chromatin structure by enhancing binding of the linker histone H1. Thus, at large silenced genomic regions, ubH2A-mediated heterochromatin generation could play an important role in silencing.
6.6 Histone Ubiquitylation and DNA Repair
Although ubiquitylation of both histones H2A and H2B is primarily associated with the regulation of gene expression, histone ubiquitylation also plays an important role in other cellular processes. Notably, histones are ubiquitylated during the cellular response to DNA damage, and many studies have implicated ubH2A and ubH2B in the regulation of DNA repair processes [reviewed in Ulrich and Walden (2010)]. Ubiquitylation of histones H2A and H2B regulates recruitment of downstream effectors of the DNA damage response and also modulates the local chromatin environment at sites of DNA damage to facilitate efficient repair. Recent work has clarified how histone H2A is mono-ubiquitylated at sites of DNA damage. Rather than being mono-ubiquitylated on Lys119, histone H2A is mono-ubiquitylated at Lys13 and Lys15 by the RNF168 E3 ubiquitin ligase in response to DNA damage (see Sect. 6.6.3 ). The subsequent extension of Lys63-linked poly-ubiquitin chains on this mono-ubiquitylated residue by RNF8 provides binding sites for factors that regulate later stages of the DNA damage response. Whereas previous studies had implicated ubH2A-Lys119 in the response to DNA damage, these new results indicate that other residues on histone H2A and the histone variant H2AX play a more important role in this particular cellular process.
6.6.1 Histones Are Ubiquitylated at Sites of DNA Damage
Histones are ubiquitylated at sites of DNA damage, and mono-ubiquitylated forms of the histones H2A, H3, and H4 accumulate in response to UV irradiation in mammalian cells (Bergink et al. 2006; Kapetanaki et al. 2006; Wang et al. 2006). In addition, DNA double-strand breaks induce ubiquitylation of both histones H2A and H2B in human cells (Bergink et al. 2006; Moyal et al. 2011; Nakamura et al. 2011). Furthermore, histone variants such as H2AX that are associated with specific aspects of the DNA damage response pathway are also mono-ubiquitylated (Huen et al. 2007; Ikura et al. 2007; Kolas et al. 2007; Wang and Elledge 2007). Several studies suggest that histones H2A and H2AX become di- or poly-ubiquitylated in response to DNA damage and that ubiquitylation occurs at sites other than the canonical Lys119 on H2A (Mattiroli et al. 2012; Huen et al. 2007; Ikura et al. 2007; Mailand et al. 2007; Zhao et al. 2007). Thus, a variety of species of ubiquitylated histones are present locally on chromatin at sites of DNA damage.
6.6.2 Histone Ubiquitylation Occurs Downstream of ATM/ATR Signaling
How is histone ubiquitylation regulated at sites of DNA damage? Studies suggest that histones are ubiquitylated as part of the DNA damage response. The response to DNA damage is mediated in part by the ataxia telangiectasia mutated (ATM), ataxia telangiectasia and Rad3-related (ATR), and DNA-PK kinases [reviewed in Ciccia and Elledge (2010)]. While ATM and DNA-PK are activated directly by DNA-damaging agents such as UV irradiation, ATR is activated following its recruitment to single-stranded DNA regions generated at stalled replication forks (Ciccia and Elledge 2010). Following DNA damage, ATM and ATR then phosphorylate a number of substrate proteins that are involved in mediating DNA repair processes (Ciccia and Elledge 2010). One of the critical early targets for phosphorylation at DNA damage sites is Ser139 of the histone variant H2AX, generating phosphorylated H2AX (γH2AX) (Ciccia and Elledge 2010). Many of the later steps in the DNA damage response pathway rely on effectors recruited through interactions with γH2AX.
Notably, the ATM and ATR signaling kinases are required for the appearance of ubH2A and ubH2B at DNA double-strand breaks (Bergink et al. 2006; Moyal et al. 2011). How do these kinases regulate histone ubiquitylation at sites of DNA damage? In mammalian cells, ATM directly phosphorylates the ubH2B E3 ubiquitin-protein isopeptide ligases RNF20 and RNF40 at DNA lesions (Moyal et al. 2011). Moreover, the phosphorylation sites on RNF20 and RNF40 are required for ubH2B accumulation in response to DNA damage (Moyal et al. 2011). Thus, DNA damage response-mediated mono-ubiquitylation of histone H2B is regulated through direct phosphorylation of the E3 ligases by the ATM kinase (Moyal et al. 2011).
In contrast, the deposition of ubH2A in response to DNA damage is regulated by factors that act downstream of the ATM and ATR kinases. The E3 ligases RNF2, DDB1-CUL4ADDB2, RNF8, and RNF168 have all been implicated in H2A ubiquitylation in response to DNA damage (Mattiroli et al. 2012; Huen et al. 2007; Ikura et al. 2007; Kolas et al. 2007; Wang and Elledge 2007; Bergink et al. 2006; Kapetanaki et al. 2006; Mailand et al. 2007; Doil et al. 2009; Stewart et al. 2009; Marteijn et al. 2009; Pinato et al. 2009; Ismail et al. 2010). Until recently, several studies had provided evidence that RNF8 was the predominant E3 ubiquitin-protein isopeptide ligase involved in H2A and H2AX ubiquitylation at DNA double-strand breaks (Huen et al. 2007; Ikura et al. 2007; Kolas et al. 2007; Wang and Elledge 2007; Mailand et al. 2007). Supporting its role as the predominant E3 ligase involved in DNA damage-induced H2A ubiquitylation, RNF8 ubiquitylates both free histones H2A and γH2AX in vitro and is required for the accumulation of ubH2A and conjugated ubiquitin at DNA damage sites (Huen et al. 2007; Ikura et al. 2007; Kolas et al. 2007; Wang and Elledge 2007; Mailand et al. 2007). Furthermore, RNF8 is the first E3 ligase to be recruited to sites of DNA damage by events that occur downstream of ATM- and ATR-dependent phosphorylation (Huen et al. 2007; Kolas et al. 2007; Mailand et al. 2007; Ciccia and Elledge 2010). The recruitment of RNF8 is mediated through interactions with phosphorylated MDC1, which itself binds γH2AX through its BRCT repeats (Huen et al. 2007; Kolas et al. 2007; Mailand et al. 2007; Ciccia and Elledge 2010). The RING finger domain of RNF8, and thus its E3 ligase activity, is required for the subsequent recruitment of a second E3 ligase, RNF168, via its ubiquitin-binding domains (Doil et al. 2009; Stewart et al. 2009; Pinato et al. 2009). This ordered recruitment suggested that RNF8, rather than RNF168 or another E3 ligase, was likely to be the enzyme responsible for mono-ubiquitylation of histone H2A at DNA damage sites.
6.6.3 RNF168 Ubiquitylates H2A Lys13,15 at DNA Damage Sites
However, the order of recruitment of RNF8 and RNF168 does not reflect their roles in H2A ubiquitylation. Instead, recent studies have shown that RNF168 is the E3 ligase responsible for H2A mono-ubiquitylation at DNA damage sites. There was some initial confusion in determining which of these E3 ligases mono-ubiquitylates H2A at DNA damage sites due to both the substrates used to characterize ubiquitylation in vitro and the specificity of the ubiquitylation itself. Importantly, recent studies have shown that RNF8 and RNF168 have differential mono-ubiquitylation activity on nucleosomal versus free histone H2A. While RNF8 mono-ubiquitylates free histone H2A in vitro, RNF8 does not mono-ubiquitylate nucleosomal H2A (Mattiroli et al. 2012; Huen et al. 2007; Ikura et al. 2007; Kolas et al. 2007; Wang and Elledge 2007; Mailand et al. 2007). In contrast, RNF168 mono-ubiquitylates nucleosomal histone H2A (Mattiroli et al. 2012). Notably, the specificity of this H2A ubiquitylation differs from PRC1-mediated H2A mono-ubiquitylation at Lys119. In fact, RNF168 mono-ubiquitylates Lys13 and Lys15 of nucleosomal histone H2A (Mattiroli et al. 2012; Gatti et al. 2012). While RNF8 does not mono-ubiquitylate H2A, it can extend Lys63-linked ubiquitin chains on histone H2A from the mono-ubiquitylated Lys13,15 of H2A (Mattiroli et al. 2012). These Lys63-linked ubiquitin chains do not form on histone H2A that is mono-ubiquitylated at Lys119 (Mattiroli et al. 2012).
Why, then, is RNF8 required for ubiquitylation of H2A and γH2AX at DNA damage sites? The Lys63-linked ubiquitin chains generated by RNF8 provide binding sites for RNF168 (Doil et al. 2009; Stewart et al. 2009). Notably, these Lys63-linked ubiquitin chains can be conjugated to proteins other than the histones. A model has been proposed in which RNF8 ubiquitylates non-nucleosomal substrate(s) at sites of DNA damage, which are required for the subsequent recruitment of RNF168 (Mattiroli et al. 2012) (Fig. 6.8). Although the identity of these non-nucleosomal substrates is unknown, candidates for these include valosin-containing protein (VCP, also known as p97), L3MBTL1, KU80, CHK2, and KDM4A (Mallette et al. 2012; Acs et al. 2011; Feng and Chen 2012; Meerang et al. 2011). In the proposed model, RNF168 binds the ubiquitylated chains on one or more of these unknown protein substrates through its ubiquitin-binding domains (Doil et al. 2009; Stewart et al. 2009; Pinato et al. 2009). Once recruited to sites of DNA damage, RNF168 then mono-ubiquitylates histone H2A at Lys13,15, followed by RNF8-mediated extension of these ubiquitin chains through Lys63 linkages (Fig. 6.8).

RNF8/RNF168 ubiquitylate histone H2A at Lys13,15. (a) Human RNF8 ubiquitylates unknown non-nucleosomal substrate(s) at sites of DNA damage. (b) RNF168 subsequently binds the ubiquitylated chains on one or more of these protein substrates through its ubiquitin-binding domains. Following this, RNF168 then mono-ubiquitylates histone H2A at Lys13,15. (c) RNF8 then extends these ubiquitin chains through ubiquitin Lys63 linkages. These Lys63-linked poly-ubiquitin chains on histone H2A are involved in the recruitment of the BRCA1 complex and other downstream regulators of the DNA damage response pathway such as 53BP1. In addition, mono-ubiquitylation of H2A at Lys13,15 might also affect nucleosomal structure, thus providing a permissive chromatin environment in which DNA repair can take place
What is the downstream function of these Lys63-linked ubiquitin chains on histones H2A and γH2AX in the DNA damage response? Lys63-linked ubiquitin chains resulting from RNF8 and RNF168 activity can be bound by the ubiquitin-interacting motif domain of RAP80 (Huen et al. 2007; Kolas et al. 2007; Wang and Elledge 2007). RAP80 interacts with the scaffold protein ABRA1 to recruit the BRCA1 complex, which itself is an E3 ligase (Huen et al. 2007; Kolas et al. 2007; Wang and Elledge 2007). Thus, RNF168/RNF8-mediated histone H2A ubiquitylation results in the subsequent recruitment of the BRCA1 complex and other downstream regulators of the DNA damage response pathway such as 53BP1 [reviewed in Ulrich and Walden (2010)]. In addition to the recruitment of downstream effectors of the DNA damage response, mono-ubiquitylation of H2A at Lys13,15 might also affect the structure of the nucleosome, thus providing a permissive chromatin environment in which DNA repair can occur (for further discussion, see Sect. 6.6.5 ).
6.6.4 Histone Ubiquitylation at DNA Damage Sites Is Reversible
The addition of ubiquitin to histones during the DNA damage response is reversible, and several DUBs have been suggested as potential candidates for involvement in this process. For example, the DUB BRCC3 (BRCC36) is a subunit of the BRCA1 complex that is recruited by interactions with RAP80. BRCC3 can de-ubiquitylate Lys63-linked poly-ubiquitin chains (Sobhian et al. 2007). In addition, the DUB USP3 regulates histone ubiquitylation at DNA damage sites. Supporting its involvement, knockdown of USP3 results in persistent ubH2A foci at DNA damage sites, whereas its overexpression causes the loss of these foci (Doil et al. 2009; Nicassio et al. 2007). Although the lysine specificity of USP3 is not known, the role of this DUB at DNA damage sites suggests that USP3 might target mono-ubiquitylated Lys13 and Lys15 of histone H2A. Thus, both the addition and removal of ubiquitin from histones at DNA damage sites are regulated during the response to DNA damage.
6.6.5 Histone Ubiquitylation Generates a Permissive Chromatin Environment at DNA Damage Sites
Does histone ubiquitylation have a functional role in regulating the DNA damage response in addition to its role in recruiting downstream effectors? Several studies suggest that ubiquitylation of both histones H2A and H2B might play a role in generating a local chromatin environment that is permissive to assembly of the machinery required for DNA repair [reviewed in Shiloh et al. (2011)]. The efficiency of DNA damage repair is reduced in cells depleted for the H2B E3 ligases, RNF20 and RNF40 (Moyal et al. 2011; Chernikova et al. 2010; Kari et al. 2011). Evidence suggests that ubH2B might affect the efficiency of the DNA damage response by altering the local chromatin environment, either through ubH2B itself or through the recruitment of chromatin remodeling factors. For instance, RNF20 recruits the chromatin remodeling factor SNF2h to DNA double-strand breaks (Nakamura et al. 2011). Depletion of SNF2h or expression of H2B-K120R compromises resection of DNA ends and the recruitment of downstream factors such as RAD51 and BRCA1 (Nakamura et al. 2011). Furthermore, the recruitment of chromatin remodeling factors by the E3 ligases involved in histone ubiquitylation is not limited to ubH2B. For example, RNF8 interacts with the NuRD chromatin remodeling complex subunit CHD4 independent of its E3 ubiquitin ligase activity to recruit CHD4 to DNA lesions (Luijsterburg et al. 2012). Thus, H2B- and H2A-specific E3 ligases can regulate both chromatin remodeling and histone ubiquitylation at DNA lesions, generating a local chromatin environment that is permissive to DNA repair.
Studies indicate that although ubH2B promotes the stability of individual nucleosomes, ubH2B is disruptive to higher order chromatin structure (Fierz et al. 2011) (for further discussion, see Sect. 6.5.1.10 ). The generation of a euchromatic chromatin environment might be important for efficient repair by facilitating rapid recruitment of factors to the local site of DNA damage. Notably, mono-ubiquitylation of histone H2A during the DNA damage response might have similar effects on the chromatin fiber to those observed for ubH2B. The two lysine residues that are ubiquitylated by RNF168, Lys13 and Lys15, are located on the amino-terminal tail of histone H2A, close to the histone-fold domain (Fig. 6.2). Importantly, Lys13 and Lys15 lie close to histone H2B Lys120 within the nucleosome structure. Thus, the effects on higher order chromatin structure resulting from mono-ubiquitylation of histone H2A Lys13,15 may be similar to those caused by mono-ubiquitylation of histone H2B Lys120.
6.6.6 Histone Ubiquitylation Is Involved in the DNA Damage Response
To summarize what has been described in Sects. 6.6.1 – 6.6.5 , histones H2A and H2B are both mono-ubiquitylated during the response to DNA damage. Whereas histone H2B is mono-ubiquitylated at Lys120, histone H2A is mono-ubiquitylated at Lys13 and Lys15. Subsequently, Lys63-linked poly-ubiquitin chains are extended on histone H2A, and these chains recruit downstream effectors involved in the DNA damage response. Notably, some evidence suggests that in addition to recruiting downstream effectors, histone mono-ubiquitylation at sites of DNA damage might generate a local chromatin environment that is permissive to repair. However, further studies are required to examine the mechanism of action of ubH2B-Lys120 and ubH2A-Lys13,15 in the DNA damage response.
6.7 Cell Cycle Regulation by Histone Mono-ubiquitylation
In addition to its role in DNA repair, H2A mono-ubiquitylation is also involved in the regulation of cell cycle progression. During the early 1980s, soon after the initial studies that identified ubH2A, it became apparent that ubH2A was absent from isolated metaphase chromosomes (Matsui et al. 1979; Mueller et al. 1985; Wu et al. 1981). In addition, it was observed that the levels of ubH2A and H3 Ser10 phosphorylation inversely correlate during the cell cycle (Joo et al. 2007; Matsui et al. 1979; Mueller et al. 1985; Wu et al. 1981). The identification of the USP16 DUB has provided a potential mechanism to explain how ubH2A levels are regulated during the cell cycle (Joo et al. 2007). De-ubiquitylation of ubH2A by USP16 is required for cell cycle progression (Joo et al. 2007). Furthermore, efficient chromosome segregation during mitosis requires USP16, which regulates Aurora B kinase-mediated phosphorylation of H3 Ser10 (Joo et al. 2007). How is USP16 activity regulated during the cell cycle? USP16 is sequentially phosphorylated and de-phosphorylated during the cell cycle, potentially by the Cdc2/Cyclin B complex that phosphorylates USP16 in vitro (Cai et al. 1999). Moreover, an enzymatically inactive form of USP16 fails to dissociate from mitotic chromosomes, suggesting that de-ubiquitylation of ubH2A and/or the subsequent phosphorylation of H3 Ser10 are required for its localization to chromatin (Cai et al. 1999). Other DUBs might also be involved in the removal of ubiquitin from ubH2A during the cell cycle. For example, knockdown of USP3 is required for S phase progression (Nicassio et al. 2007). However, these DUBs also regulate aspects of the DNA damage response and it is therefore difficult to separate their effects on DNA damage repair from their role in the cell cycle. In addition, mono-ubiquitylation of histone H2B could also be involved in regulating aspects of the cell cycle. Recent work suggests that ubH2B promotes replication fork progression, perhaps due to its role in enhancing nucleosome stability and/or occupancy (Trujillo and Osley 2012).
6.8 Conclusion
Ubiquitylation of histones plays an important role in cellular processes that include transcription, the DNA damage response, and the cell cycle. Intriguingly, mono-ubiquitylation of histones H2A and H2B appears to have opposite effects on many different aspects of gene expression. First, whereas histone H2A mono-ubiquitylation is generally repressive to gene expression, histone H2B is associated with active transcription. Second, ubH2B and ubH2A have opposite effects on methylation at histone H3 Lys4. Third, ubH2B promotes transcription elongation by RNA polymerase II, while ubH2A inhibits transcription elongation. Lastly, while ubH2B facilitates more open higher order chromatin structure, ubH2A enhances heterochromatin formation by promoting association of the linker histone H1. Thus, ubH2A and ubH2B can be regarded as having opposite effects in regulating many aspects of gene expression.
Notably, however, there are also a number of intriguing similarities between ubH2A and ubH2B. For instance, both ubH2B- and ubH2A-specific DUBs are associated with histone acetyltransferase complexes. In addition, the sequential addition and removal of ubiquitin from histones H2A and H2B appear to be important for repression and activation respectively of particular target genes. Thus, mechanistically there might be many parallels between ubH2A and ubH2B that could provide insight into their function in regulating gene expression.
The recent identification of E3 ligases that ubiquitylate novel residues on the amino-terminal tails of histones H2A and H2B raises new questions regarding the mechanisms through which histone ubiquitylation regulates gene expression and other cellular processes. In particular, the role of the novel lysines that are ubiquitylated on histone H2A during the DNA damage response is an important topic for future investigation. Future studies on the function of the novel ubiquitylated lysines on the amino-terminal tails of histones H2A and H2B may provide new insight into how histone ubiquitylation regulates both chromatin structure and gene expression. Overall, these studies support the role of histone ubiquitylation as a critical histone modification that has important regulatory functions in chromatin structure, gene expression, DNA repair, and the cell cycle.
Abbreviations
- BUR:
-
Bur1/Bur2 cyclin-dependent protein kinase complex
- COMPASS:
-
Complex Proteins Associated with Set1
- CTD:
-
Carboxy-terminal heptapeptide repeat sequences/domain
- Cys:
-
Cysteine
- dRAF:
-
Drosophila Ring-Associated Factor complex
- DUB:
-
De-ubiquitylating enzyme
- FACT:
-
Facilitates Chromatin Transcription complex
- Gly:
-
Glycine
- JAMM/MPN+:
-
JAB1/MPN/Mov34 metalloenzyme
- Lys:
-
Lysine
- MJD:
-
Machado-Josephin domain
- OTU:
-
Ovarian tumor domain
- PAF:
-
Polymerase-Associated Factor complex
- Pol II:
-
RNA polymerase II
- PRC1:
-
Polycomb repressive complex 1
- PR-DUB:
-
Polycomb repressive de-ubiquitylating enzyme complex
- SAGA:
-
Spt-Ada-Gcn5 acetyltransferase complex
- Ser:
-
Serine
- siRNA:
-
Small interfering RNA
- Thr:
-
Threonine
- tss:
-
Transcription start site
- ubH2A:
-
Mono-ubiquitylated histone H2A
- ubH2B:
-
Mono-ubiquitylated histone H2B
- UCH:
-
Ubiquitin carboxy-terminal hydrolase
- USP/UBP:
-
Ubiquitin-specific protease
- ZnF-UBP:
-
Zinc finger ubiquitin-specific protease
- γH2AX:
-
Phosphorylated histone variant H2AX
References
Acs K et al (2011) The AAA-ATPase VCP/p97 promotes 53BP1 recruitment by removing L3MBTL1 from DNA double-strand breaks. Nat Struct Mol Biol 18(12):1345–1350
Alatzas A, Foundouli A (2006) Distribution of ubiquitinated histone H2A during plant cell differentiation in maize root and dedifferentiation in callus culture. Plant Sci 171(4):481–487
Atanassov BS, Dent SY (2011) USP22 regulates cell proliferation by deubiquitinating the transcriptional regulator FBP1. EMBO Rep 12(9):924–930
Atanassov BS et al (2009) Gcn5 and SAGA regulate shelterin protein turnover and telomere maintenance. Mol Cell 35(3):352–364
Ballal NR et al (1975) Changes in nucleolar proteins and their phosphorylation patterns during liver regeneration. J Biol Chem 250(15):5921–5925
Batta K et al (2011) Genome-wide function of H2B ubiquitylation in promoter and genic regions. Genes Dev 25(21):2254–2265
Bentley ML et al (2011) Recognition of UbcH5c and the nucleosome by the Bmi1/Ring1b ubiquitin ligase complex. EMBO J 30(16):3285–3297
Bergink S et al (2006) DNA damage triggers nucleotide excision repair-dependent monoubiquitylation of histone H2A. Genes Dev 20(10):1343–1352
Bott M et al (2011) The nuclear deubiquitinase BAP1 is commonly inactivated by somatic mutations and 3p21.1 losses in malignant pleural mesothelioma. Nat Genet 43(7):668–672
Bratzel F et al (2010) Keeping cell identity in Arabidopsis requires PRC1 RING-finger homologs that catalyze H2A monoubiquitination. Curr Biol 20(20):1853–1859
Bray S, Musisi H, Bienz M (2005) Bre1 is required for Notch signaling and histone modification. Dev Cell 8(2):279–286
Briggs SD et al (2002) Gene silencing: trans-histone regulatory pathway in chromatin. Nature 418(6897):498
Buchwald G et al (2006) Structure and E3-ligase activity of the Ring-Ring complex of polycomb proteins Bmi1 and Ring1b. EMBO J 25(11):2465–2474
Buratowski S (2009) Progression through the RNA polymerase II CTD cycle. Mol Cell 36(4):541–546
Buszczak M, Paterno S, Spradling AC (2009) Drosophila stem cells share a common requirement for the histone H2B ubiquitin protease scrawny. Science 323(5911):248–251
Cai SY, Babbitt RW, Marchesi VT (1999) A mutant deubiquitinating enzyme (Ubp-M) associates with mitotic chromosomes and blocks cell division. Proc Natl Acad Sci USA 96(6):2828–2833
Calonje M et al (2008) EMBRYONIC FLOWER1 participates in polycomb group-mediated AG gene silencing in Arabidopsis. Plant Cell 20(2):277–291
Calvo V, Beato M (2011) BRCA1 counteracts progesterone action by ubiquitination leading to progesterone receptor degradation and epigenetic silencing of target promoters. Cancer Res 71(9):3422–3431
Cao R, Tsukada Y, Zhang Y (2005) Role of Bmi-1 and Ring1A in H2A ubiquitylation and Hox gene silencing. Mol Cell 20(6):845–854
Chandrasekharan MB, Huang F, Sun ZW (2009) Ubiquitination of histone H2B regulates chromatin dynamics by enhancing nucleosome stability. Proc Natl Acad Sci USA 106(39):16686–16691
Chen HY et al (1998) Ubiquitination of histone H3 in elongating spermatids of rat testes. J Biol Chem 273(21):13165–13169
Chen A et al (2002) Autoubiquitination of the BRCA1*BARD1 RING ubiquitin ligase. J Biol Chem 277(24):22085–22092
Chen D et al (2010) The Arabidopsis PRC1-like ring-finger proteins are necessary for repression of embryonic traits during vegetative growth. Cell Res 20(12):1332–1344
Chen S et al (2012) RAD6 regulates the dosage of p53 by a combination of transcriptional and posttranscriptional mechanisms. Mol Cell Biol 32(2):576–587
Chernikova SB et al (2010) Deficiency in Bre1 impairs homologous recombination repair and cell cycle checkpoint response to radiation damage in mammalian cells. Radiat Res 174(5):558–565
Chu F et al (2006) Mapping post-translational modifications of the histone variant MacroH2A1 using tandem mass spectrometry. Mol Cell Proteomics 5(1):194–203
Ciccia A, Elledge SJ (2010) The DNA damage response: making it safe to play with knives. Mol Cell 40(2):179–204
Clague MJ, Urbe S (2010) Ubiquitin: same molecule, different degradation pathways. Cell 143(5):682–685
Daniel JA et al (2004) Deubiquitination of histone H2B by a yeast acetyltransferase complex regulates transcription. J Biol Chem 279(3):1867–1871
Davies N, Lindsey GG (1994) Histone H2B (and H2A) ubiquitination allows normal histone octamer and core particle reconstitution. Biochim Biophys Acta 1218(2):187–193
de Napoles M et al (2004) Polycomb group proteins Ring1A/B link ubiquitylation of histone H2A to heritable gene silencing and X inactivation. Dev Cell 7(5):663–676
Dehe PM et al (2005) Histone H3 lysine 4 mono-methylation does not require ubiquitination of histone H2B. J Mol Biol 353(3):477–484
Dehe PM et al (2006) Protein interactions within the Set1 complex and their roles in the regulation of histone 3 lysine 4 methylation. J Biol Chem 281(46):35404–35412
Doil C et al (2009) RNF168 binds and amplifies ubiquitin conjugates on damaged chromosomes to allow accumulation of repair proteins. Cell 136(3):435–446
Dover J et al (2002) Methylation of histone H3 by COMPASS requires ubiquitination of histone H2B by Rad6. J Biol Chem 277(32):28368–28371
Emre NC et al (2005) Maintenance of low histone ubiquitylation by Ubp10 correlates with telomere-proximal Sir2 association and gene silencing. Mol Cell 17(4):585–594
Fang J et al (2004) Ring1b-mediated H2A ubiquitination associates with inactive X chromosomes and is involved in initiation of X inactivation. J Biol Chem 279(51):52812–52815
Feng L, Chen J (2012) The E3 ligase RNF8 regulates KU80 removal and NHEJ repair. Nat Struct Mol Biol 19(2):201–206
Fierz B et al (2011) Histone H2B ubiquitylation disrupts local and higher-order chromatin compaction. Nat Chem Biol 7(2):113–119
Fleming AB et al (2008) H2B ubiquitylation plays a role in nucleosome dynamics during transcription elongation. Mol Cell 31(1):57–66
Fleury D et al (2007) The Arabidopsis thaliana homolog of yeast BRE1 has a function in cell cycle regulation during early leaf and root growth. Plant Cell 19(2):417–432
Frescas D et al (2007) JHDM1B/FBXL10 is a nucleolar protein that represses transcription of ribosomal RNA genes. Nature 450(7167):309–313
Gallego-Sanchez A et al (2012) Reversal of PCNA ubiquitylation by Ubp10 in Saccharomyces cerevisiae. PLoS Genet 8(7):e1002826
Gardner RG, Nelson ZW, Gottschling DE (2005) Ubp10/Dot4p regulates the persistence of ubiquitinated histone H2B: distinct roles in telomeric silencing and general chromatin. Mol Cell Biol 25(14):6123–6139
Gatti M et al (2012) A novel ubiquitin mark at the N-terminal tail of histone H2As targeted by RNF168 ubiquitin ligase. Cell Cycle 11(13):2538–2544
Gearhart MD et al (2006) Polycomb group and SCF ubiquitin ligases are found in a novel BCOR complex that is recruited to BCL6 targets. Mol Cell Biol 26(18):6880–6889
Gerard B et al (2012) Lysine 394 is a novel Rad6B-induced ubiquitination site on beta-catenin. Biochim Biophys Acta 1823(10):1686–1696
Goldknopf IL, Busch H (1975) Remarkable similarities of peptide fingerprints of histone 2A and nonhistone chromosomal protein A24. Biochem Biophys Res Commun 65(3):951–960
Goldknopf IL, Busch H (1977) Isopeptide linkage between nonhistone and histone 2A polypeptides of chromosomal conjugate-protein A24. Proc Natl Acad Sci USA 74(3):864–868
Goldknopf IL et al (1975) Isolation and characterization of protein A24, a “histone-like” non-histone chromosomal protein. J Biol Chem 250(18):7182–7187
Goldknopf IL et al (1977) Presence of protein A24 in rat liver nucleosomes. Proc Natl Acad Sci USA 74(12):5492–5495
Gorfinkiel N et al (2004) The Drosophila polycomb group gene sex combs extra encodes the ortholog of mammalian Ring1 proteins. Mech Dev 121(5):449–462
Gunjan A, Verreault A (2003) A Rad53 kinase-dependent surveillance mechanism that regulates histone protein levels in S. cerevisiae. Cell 115(5):537–549
Gutierrez L et al (2012) The role of the histone H2A ubiquitinase Sce in polycomb repression. Development 139(1):117–127
Haas A et al (1990) Ubiquitin-mediated degradation of histone H3 does not require the substrate-binding ubiquitin protein ligase, E3, or attachment of polyubiquitin chains. J Biol Chem 265(35):21664–21669
Hatch CL, Bonner WM, Moudrianakis EN (1983) Minor histone 2A variants and ubiquinated forms in the native H2A:H2B dimer. Science 221(4609):468–470
Henry KW et al (2003) Transcriptional activation via sequential histone H2B ubiquitylation and deubiquitylation, mediated by SAGA-associated Ubp8. Genes Dev 17(21):2648–2663
Hewawasam G et al (2010) Psh1 is an E3 ubiquitin ligase that targets the centromeric histone variant Cse4. Mol Cell 40(3):444–454
Hicke L (2001) Protein regulation by monoubiquitin. Nat Rev Mol Cell Biol 2(3):195–201
Huen MS et al (2007) RNF8 transduces the DNA-damage signal via histone ubiquitylation and checkpoint protein assembly. Cell 131(5):901–914
Hunt LT, Dayhoff MO (1977) Amino-terminal sequence identity of ubiquitin and the nonhistone component of nuclear protein A24. Biochem Biophys Res Commun 74(2):650–655
Hwang WW et al (2003) A conserved RING finger protein required for histone H2B monoubiquitination and cell size control. Mol Cell 11(1):261–266
Ikura T et al (2007) DNA damage-dependent acetylation and ubiquitination of H2AX enhances chromatin dynamics. Mol Cell Biol 27(20):7028–7040
Ismail IH et al (2010) BMI1-mediated histone ubiquitylation promotes DNA double-strand break repair. J Cell Biol 191(1):45–60
Jaehning JA (2010) The Paf1 complex: platform or player in RNA polymerase II transcription? Biochim Biophys Acta 1799(5–6):379–388
Jason LJ et al (2005) Histone H2A ubiquitination does not preclude histone H1 binding, but it facilitates its association with the nucleosome. J Biol Chem 280(6):4975–4982
Jentsch S, McGrath JP, Varshavsky A (1987) The yeast DNA repair gene RAD6 encodes a ubiquitin-conjugating enzyme. Nature 329(6135):131–134
Joo HY et al (2007) Regulation of cell cycle progression and gene expression by H2A deubiquitination. Nature 449:1068–1072
Joo HY et al (2011) Regulation of histone H2A and H2B deubiquitination and Xenopus development by USP12 and USP46. J Biol Chem 286(9):7190–7201
Jung I et al (2012) H2B monoubiquitylation is a 5'-enriched active transcription mark and correlates with exon-intron structure in human cells. Genome Res 22:1026–1035
Kahana A, Gottschling DE (1999) DOT4 links silencing and cell growth in Saccharomyces cerevisiae. Mol Cell Biol 19(10):6608–6620
Kao CF et al (2004) Rad6 plays a role in transcriptional activation through ubiquitylation of histone H2B. Genes Dev 18(2):184–195
Kapetanaki MG et al (2006) The DDB1-CUL4ADDB2 ubiquitin ligase is deficient in xeroderma pigmentosum group E and targets histone H2A at UV-damaged DNA sites. Proc Natl Acad Sci USA 103(8):2588–2593
Kari V et al (2011) The H2B ubiquitin ligase RNF40 cooperates with SUPT16H to induce dynamic changes in chromatin structure during DNA double-strand break repair. Cell Cycle 10(20):3495–3504
Kerppola TK (2009) Polycomb group complexes – many combinations, many functions. Trends Cell Biol 19(12):692–704
Kim J, Roeder RG (2009) Direct Bre1-Paf1 complex interactions and RING finger-independent Bre1-Rad6 interactions mediate histone H2B ubiquitylation in yeast. J Biol Chem 284(31):20582–20592
Kim J, Hake SB, Roeder RG (2005) The human homolog of yeast BRE1 functions as a transcriptional coactivator through direct activator interactions. Mol Cell 20(5):759–770
Kim J et al (2009) RAD6-mediated transcription-coupled H2B ubiquitylation directly stimulates H3K4 methylation in human cells. Cell 137(3):459–471
Kohler A et al (2010) Structural basis for assembly and activation of the heterotetrameric SAGA histone H2B deubiquitinase module. Cell 141(4):606–617
Koken M et al (1991a) Dhr6, a Drosophila homolog of the yeast DNA-repair gene RAD6. Proc Natl Acad Sci USA 88(9):3832–3836
Koken MH et al (1991b) Structural and functional conservation of two human homologs of the yeast DNA repair gene RAD6. Proc Natl Acad Sci USA 88(20):8865–8869
Kolas NK et al (2007) Orchestration of the DNA-damage response by the RNF8 ubiquitin ligase. Science 318(5856):1637–1640
Komander D, Clague MJ, Urbe S (2009) Breaking the chains: structure and function of the deubiquitinases. Nat Rev Mol Cell Biol 10(8):550–563
Lagarou A et al (2008) dKDM2 couples histone H2A ubiquitylation to histone H3 demethylation during Polycomb group silencing. Genes Dev 22(20):2799–2810
Lang G et al (2011) The tightly controlled deubiquitination activity of the human SAGA complex differentially modifies distinct gene regulatory elements. Mol Cell Biol 31(18):3734–3744
Laribee RN et al (2005) BUR kinase selectively regulates H3 K4 trimethylation and H2B ubiquitylation through recruitment of the PAF elongation complex. Curr Biol 15(16):1487–1493
Latham JA et al (2011) Chromatin signaling to kinetochores: transregulation of Dam1 methylation by histone H2B ubiquitination. Cell 146(5):709–719
Lee KY, Myung K (2008) PCNA modifications for regulation of post-replication repair pathways. Mol Cells 26(1):5–11
Lee JS et al (2007) Histone crosstalk between H2B monoubiquitination and H3 methylation mediated by COMPASS. Cell 131(6):1084–1096
Lee JS et al (2012) Codependency of H2B monoubiquitination and nucleosome reassembly on Chd1. Genes Dev 26(9):914–919
Li Z et al (2006) Structure of a Bmi-1-Ring1B polycomb group ubiquitin ligase complex. J Biol Chem 281(29):20643–20649
Liu Z, Oughtred R, Wing SS (2005) Characterization of E3Histone, a novel testis ubiquitin protein ligase which ubiquitinates histones. Mol Cell Biol 25(7):2819–2831
Luger K et al (1997) Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature 389(6648):251–260
Luijsterburg MS et al (2012) A new non-catalytic role for ubiquitin ligase RNF8 in unfolding higher-order chromatin structure. EMBO J 31(11):2511–2527
Ma MK et al (2011) Histone crosstalk directed by H2B ubiquitination is required for chromatin boundary integrity. PLoS Genet 7(7):e1002175
Machida YJ et al (2009) The deubiquitinating enzyme BAP1 regulates cell growth via interaction with HCF-1. J Biol Chem 284(49):34179–34188
Mailand N et al (2007) RNF8 ubiquitylates histones at DNA double-strand breaks and promotes assembly of repair proteins. Cell 131(5):887–900
Malik S, Bhaumik SR (2010) Mixed lineage leukemia: histone H3 lysine 4 methyltransferases from yeast to human. FEBS J 277(8):1805–1821
Mallery DL, Vandenberg CJ, Hiom K (2002) Activation of the E3 ligase function of the BRCA1/BARD1 complex by polyubiquitin chains. EMBO J 21(24):6755–6762
Mallette FA et al (2012) RNF8- and RNF168-dependent degradation of KDM4A/JMJD2A triggers 53BP1 recruitment to DNA damage sites. EMBO J 31(8):1865–1878
Marteijn JA et al (2009) Nucleotide excision repair-induced H2A ubiquitination is dependent on MDC1 and RNF8 and reveals a universal DNA damage response. J Cell Biol 186(6):835–847
Matsui SI, Seon BK, Sandberg AA (1979) Disappearance of a structural chromatin protein A24 in mitosis: implications for molecular basis of chromatin condensation. Proc Natl Acad Sci USA 76(12):6386–6390
Mattiroli F et al (2012) RNF168 Ubiquitinates K13-15 on H2A/H2AX to drive DNA damage signaling. Cell 150(6):1182–1195
Meerang M et al (2011) The ubiquitin-selective segregase VCP/p97 orchestrates the response to DNA double-strand breaks. Nat Cell Biol 13(11):1376–1382
Mimnaugh EG et al (2001) Caspase-dependent deubiquitination of monoubiquitinated nucleosomal histone H2A induced by diverse apoptogenic stimuli. Cell Death Differ 8(12):1182–1196
Minsky N, Oren M (2004) The RING domain of Mdm2 mediates histone ubiquitylation and transcriptional repression. Mol Cell 16(4):631–639
Minsky N et al (2008) Monoubiquitinated H2B is associated with the transcribed region of highly expressed genes in human cells. Nat Cell Biol 10(4):483–488
Misaghi S et al (2009) Association of C-terminal ubiquitin hydrolase BRCA1-associated protein 1 with cell cycle regulator host cell factor 1. Mol Cell Biol 29(8):2181–2192
Morillon A et al (2005) Dynamic lysine methylation on histone H3 defines the regulatory phase of gene transcription. Mol Cell 18(6):723–734
Moyal L et al (2011) Requirement of ATM-dependent monoubiquitylation of histone H2B for timely repair of DNA double-strand breaks. Mol Cell 41(5):529–542
Mueller RD et al (1985) Identification of ubiquitinated histones 2A and 2B in Physarum polycephalum. Disappearance of these proteins at metaphase and reappearance at anaphase. J Biol Chem 260(8):5147–5153
Nakagawa T et al (2008) Deubiquitylation of histone H2A activates transcriptional initiation via trans-histone cross-talk with H3K4 di- and trimethylation. Genes Dev 22(1):37–49
Nakamura K et al (2011) Regulation of homologous recombination by RNF20-dependent H2B ubiquitination. Mol Cell 41(5):515–528
Ng HH et al (2002) Lysine methylation within the globular domain of histone H3 by Dot1 is important for telomeric silencing and Sir protein association. Genes Dev 16(12):1518–1527
Ng HH, Dole S, Struhl K (2003) The Rtf1 component of the Paf1 transcriptional elongation complex is required for ubiquitination of histone H2B. J Biol Chem 278(36):33625–33628
Nicassio F et al (2007) Human USP3 is a chromatin modifier required for S phase progression and genome stability. Curr Biol 17(22):1972–1977
Nickel BE et al (1987) Changes in the histone H2A variant H2A.Z and polyubiquitinated histone species in developing trout testis. Biochemistry 26(14):4417–4421
Ogawa H et al (2002) A complex with chromatin modifiers that occupies E2F- and Myc-responsive genes in G0 cells. Science 296(5570):1132–1136
Osley MA (2006) Regulation of histone H2A and H2B ubiquitylation. Brief Funct Genomic Proteomic 5(3):179–189
Pavri R et al (2006) Histone H2B monoubiquitination functions cooperatively with FACT to regulate elongation by RNA polymerase II. Cell 125(4):703–717
Pina B, Suau P (1985) Core histone variants and ubiquitinated histones 2A and 2B of rat cerebral cortex neurons. Biochem Biophys Res Commun 133(2):505–510
Pinato S et al (2009) RNF168, a new RING finger, MIU-containing protein that modifies chromatin by ubiquitination of histones H2A and H2AX. BMC Mol Biol 10:55
Qin F et al (2008) Arabidopsis DREB2A-interacting proteins function as RING E3 ligases and negatively regulate plant drought stress-responsive gene expression. Plant Cell 20(6):1693–1707
Rajapurohitam V et al (1999) Activation of a UBC4-dependent pathway of ubiquitin conjugation during postnatal development of the rat testis. Dev Biol 212(1):217–228
Ranjitkar P et al (2010) An E3 ubiquitin ligase prevents ectopic localization of the centromeric histone H3 variant via the centromere targeting domain. Mol Cell 40(3):455–464
Reyes-Turcu FE, Ventii KH, Wilkinson KD (2009) Regulation and cellular roles of ubiquitin-specific deubiquitinating enzymes. Annu Rev Biochem 78:363–397
Reynolds P et al (1990) The rhp6+ gene of Schizosaccharomyces pombe: a structural and functional homolog of the RAD6 gene from the distantly related yeast Saccharomyces cerevisiae. EMBO J 9(5):1423–1430
Robzyk K, Recht J, Osley MA (2000) Rad6-dependent ubiquitination of histone H2B in yeast. Science 287(5452):501–504
Samara NL et al (2010) Structural insights into the assembly and function of the SAGA deubiquitinating module. Science 328(5981):1025–1029
Samara NL, Ringel AE, Wolberger C (2012) A role for intersubunit interactions in maintaining SAGA deubiquitinating module structure and activity. Structure 20(8):1414–1424
Sanchez C et al (2007) Proteomics analysis of Ring1B/Rnf2 interactors identifies a novel complex with the Fbxl10/Jhdm1B histone demethylase and the Bcl6 interacting corepressor. Mol Cell Proteomics 6(5):820–834
Sanchez-Pulido L et al (2008) RAWUL: a new ubiquitin-like domain in PRC1 ring finger proteins that unveils putative plant and worm PRC1 orthologs. BMC Genomics 9:308
Sarcevic B et al (2002) Regulation of the ubiquitin-conjugating enzyme hHR6A by CDK-mediated phosphorylation. EMBO J 21(8):2009–2018
Sarcinella E et al (2007) Monoubiquitylation of H2A.Z distinguishes its association with euchromatin or facultative heterochromatin. Mol Cell Biol 27(18):6457–6468
Scheuermann JC et al (2010) Histone H2A deubiquitinase activity of the polycomb repressive complex PR-DUB. Nature 465(7295):243–247
Schneider J et al (2005) Molecular regulation of histone H3 trimethylation by COMPASS and the regulation of gene expression. Mol Cell 19(6):849–856
Schulze JM et al (2011) Splitting the task: Ubp8 and Ubp10 deubiquitinate different cellular pools of H2BK123. Genes Dev 25(21):2242–2247
Shahbazian MD, Zhang K, Grunstein M (2005) Histone H2B ubiquitylation controls processive methylation but not monomethylation by Dot1 and Set1. Mol Cell 19(2):271–277
Shi X et al (2007) Proteome-wide analysis in Saccharomyces cerevisiae identifies several PHD fingers as novel direct and selective binding modules of histone H3 methylated at either lysine 4 or lysine 36. J Biol Chem 282(4):2450–2455
Shieh GS et al (2011) H2B ubiquitylation is part of chromatin architecture that marks exon-intron structure in budding yeast. BMC Genomics 12:627
Shiloh Y et al (2011) RNF20-RNF40: a ubiquitin-driven link between gene expression and the DNA damage response. FEBS Lett 585(18):2795–2802
Shukla A, Chaurasia P, Bhaumik SR (2009) Histone methylation and ubiquitination with their cross-talk and roles in gene expression and stability. Cell Mol Life Sci 66(8):1419–1433
Smolle M, Workman JL (2011) Signaling through chromatin: setting the scene at kinetochores. Cell 146(5):671–672
Sobhian B et al (2007) RAP80 targets BRCA1 to specific ubiquitin structures at DNA damage sites. Science 316(5828):1198–1202
Song YH, Ahn SH (2010) A Bre1-associated protein, large 1 (Lge1), promotes H2B ubiquitylation during the early stages of transcription elongation. J Biol Chem 285(4):2361–2367
Sridhar VV et al (2007) Control of DNA methylation and heterochromatic silencing by histone H2B deubiquitination. Nature 447(7145):735–738
Stewart GS et al (2009) The RIDDLE syndrome protein mediates a ubiquitin-dependent signaling cascade at sites of DNA damage. Cell 136(3):420–434
Stock JK et al (2007) Ring1-mediated ubiquitination of H2A restrains poised RNA polymerase II at bivalent genes in mouse ES cells. Nat Cell Biol 9(12):1428–1435
Sun ZW, Allis CD (2002) Ubiquitination of histone H2B regulates H3 methylation and gene silencing in yeast. Nature 418(6893):104–108
Sung P, Prakash S, Prakash L (1988) The RAD6 protein of Saccharomyces cerevisiae polyubiquitinates histones, and its acidic domain mediates this activity. Genes Dev 2(11):1476–1485
Sung P et al (1991) Yeast RAD6 encoded ubiquitin conjugating enzyme mediates protein degradation dependent on the N-end-recognizing E3 enzyme. EMBO J 10(8):2187–2193
Swerdlow PS, Schuster T, Finley D (1990) A conserved sequence in histone H2A which is a ubiquitination site in higher eucaryotes is not required for growth in Saccharomyces cerevisiae. Mol Cell Biol 10(9):4905–4911
Tanny JC et al (2007) Ubiquitylation of histone H2B controls RNA polymerase II transcription elongation independently of histone H3 methylation. Genes Dev 21(7):835–847
Thorne AW et al (1987) The structure of ubiquitinated histone H2B. EMBO J 6(4):1005–1010
Trujillo KM, Osley MA (2012) A role for H2B ubiquitylation in DNA replication. Mol Cell 48:734–746
Ulrich HD, Walden H (2010) Ubiquitin signalling in DNA replication and repair. Nat Rev Mol Cell Biol 11(7):479–489
van der Knaap JA et al (2005) GMP synthetase stimulates histone H2B deubiquitylation by the epigenetic silencer USP7. Mol Cell 17(5):695–707
van der Knaap JA et al (2010) Biosynthetic enzyme GMP synthetase cooperates with ubiquitin-specific protease 7 in transcriptional regulation of ecdysteroid target genes. Mol Cell Biol 30(3):736–744
van Leeuwen F, Gafken PR, Gottschling DE (2002) Dot1p modulates silencing in yeast by methylation of the nucleosome core. Cell 109(6):745–756
Vethantham V et al (2012) Dynamic loss of H2B ubiquitylation without corresponding changes in H3K4 trimethylation during myogenic differentiation. Mol Cell Biol 32(6):1044–1055
Vissers JH et al (2008) The many faces of ubiquitinated histone H2A: insights from the DUBs. Cell Div 3:8
Vitaliano-Prunier A et al (2008) Ubiquitylation of the COMPASS component Swd2 links H2B ubiquitylation to H3K4 trimethylation. Nat Cell Biol 10(11):1365–1371
Wang B, Elledge SJ (2007) Ubc13/Rnf8 ubiquitin ligases control foci formation of the Rap80/Abraxas/Brca1/Brcc36 complex in response to DNA damage. Proc Natl Acad Sci USA 104(52):20759–20763
Wang H et al (2004) Role of histone H2A ubiquitination in polycomb silencing. Nature 431(7010):873–878
Wang H et al (2006) Histone H3 and H4 ubiquitylation by the CUL4-DDB-ROC1 ubiquitin ligase facilitates cellular response to DNA damage. Mol Cell 22(3):383–394
Watkins JF et al (1993) The extremely conserved amino terminus of RAD6 ubiquitin-conjugating enzyme is essential for amino-end rule-dependent protein degradation. Genes Dev 7(2):250–261
Weake VM, Workman JL (2008) Histone ubiquitination: triggering gene activity. Mol Cell 29(6):653–663
Weake VM et al (2008) SAGA-mediated H2B deubiquitination controls the development of neuronal connectivity in the Drosophila visual system. EMBO J 27(2):394–405
Wei J et al (2006) Role of Bmi1 in H2A ubiquitylation and Hox gene silencing. J Biol Chem 281(32):22537–22544
Weissman AM, Shabek N, Ciechanover A (2011) The predator becomes the prey: regulating the ubiquitin system by ubiquitylation and degradation. Nat Rev Mol Cell Biol 12(9):605–620
West MH, Bonner WM (1980) Histone 2B can be modified by the attachment of ubiquitin. Nucleic Acids Res 8(20):4671–4680
Wilson MA et al (2011) Ubp8 and SAGA regulate Snf1 AMP kinase activity. Mol Cell Biol 31(15):3126–3135
Wood A et al (2003a) Bre1, an E3 ubiquitin ligase required for recruitment and substrate selection of Rad6 at a promoter. Mol Cell 11(1):267–274
Wood A et al (2003b) The Paf1 complex is essential for histone monoubiquitination by the Rad6-Bre1 complex, which signals for histone methylation by COMPASS and Dot1p. J Biol Chem 278(37):34739–34742
Wood A et al (2005) The Bur1/Bur2 complex is required for histone H2B monoubiquitination by Rad6/Bre1 and histone methylation by COMPASS. Mol Cell 20(4):589–599
Wu RS, Kohn KW, Bonner WM (1981) Metabolism of ubiquitinated histones. J Biol Chem 256(11):5916–5920
Wu L et al (2011) The RING finger protein MSL2 in the MOF complex is an E3 ubiquitin ligase for H2B K34 and is involved in crosstalk with H3 K4 and K79 methylation. Mol Cell 43(1):132–144
Wyce A et al (2007) H2B ubiquitylation acts as a barrier to Ctk1 nucleosomal recruitment prior to removal by Ubp8 within a SAGA-related complex. Mol Cell 27(2):275–288
Xia Y et al (2003) Enhancement of BRCA1 E3 ubiquitin ligase activity through direct interaction with the BARD1 protein. J Biol Chem 278(7):5255–5263
Xiao T et al (2005) Histone H2B ubiquitylation is associated with elongating RNA polymerase II. Mol Cell Biol 25(2):637–651
Xie Y, Varshavsky A (1999) The E2-E3 interaction in the N-end rule pathway: the RING-H2 finger of E3 is required for the synthesis of multiubiquitin chain. EMBO J 18(23):6832–6844
Xu L, Shen WH (2008) Polycomb silencing of KNOX genes confines shoot stem cell niches in Arabidopsis. Curr Biol 18(24):1966–1971
Zhang F, Yu X (2011) WAC, a functional partner of RNF20/40, regulates histone H2B ubiquitination and gene transcription. Mol Cell 41(4):384–397
Zhang K et al (2005) The Set1 methyltransferase opposes Ipl1 aurora kinase functions in chromosome segregation. Cell 122(5):723–734
Zhang XY et al (2008) The putative cancer stem cell marker USP22 is a subunit of the human SAGA complex required for activated transcription and cell-cycle progression. Mol Cell 29(1):102–111
Zhao GY et al (2007) A critical role for the ubiquitin-conjugating enzyme Ubc13 in initiating homologous recombination. Mol Cell 25(5):663–675
Zhao Y et al (2008) A TFTC/STAGA module mediates histone H2A and H2B deubiquitination, coactivates nuclear receptors, and counteracts heterochromatin silencing. Mol Cell 29(1):92–101
Zhou W et al (2008) Histone H2A monoubiquitination represses transcription by inhibiting RNA polymerase II transcriptional elongation. Mol Cell 29(1):69–80
Zhu B et al (2005) Monoubiquitination of human histone H2B: the factors involved and their roles in HOX gene regulation. Mol Cell 20(4):601–611
Zhu P et al (2007) A histone H2A deubiquitinase complex coordinating histone acetylation and H1 dissociation in transcriptional regulation. Mol Cell 27(4):609–621
Zhu Q et al (2011) BRCA1 tumour suppression occurs via heterochromatin-mediated silencing. Nature 477(7363):179–184
Zofall M, Grewal SI (2007) HULC, a histone H2B ubiquitinating complex, modulates heterochromatin independent of histone methylation in fission yeast. J Biol Chem 282(19):14065–14072
Zwirn P et al (1997) Arabidopsis thaliana RAD6 homolog AtUBC2 complements UV sensitivity, but not N-end rule degradation deficiency, of Saccharomyces cerevisiae rad6 mutants. Curr Genet 32(5):309–314
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer Science+Business Media New York
About this chapter
Cite this chapter
Weake, V.M. (2014). Histone Ubiquitylation Control of Gene Expression. In: Workman, J., Abmayr, S. (eds) Fundamentals of Chromatin. Springer, New York, NY. https://doi.org/10.1007/978-1-4614-8624-4_6
Download citation
DOI: https://doi.org/10.1007/978-1-4614-8624-4_6
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4614-8623-7
Online ISBN: 978-1-4614-8624-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)