
Abstract
A significant increase in lipogenesis is a metabolic hallmark of proliferating tumor cells and is required for oncogenic transformation of epithelial cells. Although most normal cells acquire the bulk of their fatty acids from the circulation, tumor cells synthesize more than 90 % of required lipids de novo. Consistent with an increased demand for lipid synthesis, diverse human cancer cells express high levels of lipogenic enzymes, such as fatty acid synthase (FASN) and stearoyl-CoA desaturase 1 (SCD1). The sterol regulatory element-binding protein 1 (SREBP1) and peroxisome proliferator-activated receptor γ (PPARγ) are master regulators of lipogenesis in diverse organisms. Previous studies have established that FASN and SCD1, the major transcriptional targets of SREBP1 and PPARγ, promote synthesis of fatty acids, which then serve as ligands for PPARγ activation. This review focuses on the potential therapeutic value of these lipogenic transcription factors as targets in cancer treatment.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
- Lipogenesis
- Transcription factor
- Sterol regulatory element-binding protein 1 (SREBP1)
- Peroxisome proliferator-activated receptor gamma (PPARγ)
- Cancer
- Lipid metabolism
- Cell proliferation
1 Introduction
Lipogenesis is almost universally upregulated in human cancers [1]. Consistent with an essential role of the sterol regulatory element-binding protein 1 (SREBP1) in sensing and regulating intracellular lipid homeostasis, increased expression of SREBP1 has been detected in colorectal carcinoma, breast and prostate cancer, and hepatocarcinoma [2–5]. Moreover, elevated expression of SREBP1 is closely correlated with malignant transformation, cancer progression, and metastasis for several cancer types, particularly hormone responsive tissue-derived cancers, such as breast and prostate cancers [2, 4, 6, 7]. SREBP1 expression correlates with the expression of FASN (encodes fatty acid synthase or FAS) and Ki-67(a nuclear marker for cellular proliferation) in colorectal cancer, suggesting a role for SREBP1 in supporting rapid cellular proliferation [7]. SREBP1 is elevated in clinical prostate cancer samples compared to benign prostatic hypertrophy [3]. Gene expression profiling of hepatocellular carcinoma (HCC) tissue and non-cancerous liver tissue showed increased lipogenic signaling in HCC. ElevatedSREBP1 expression in hepatocellular carcinoma is a known predictor of increased mortality [4, 6]. Overexpression of SREBP1 in human hepatoma HuH7 and Hep3B cells enhanced cellular proliferation and foci formation, while knockdown of SREBP1 in these cells reduced cell replication and anchorage-independent cell growth [6]. A dramatic increase of SREBP1 has been correlated with the progression of prostate cancer towards androgen-independence [3]. Oncogenic transformation of normal breast epithelial cells was accompanied by increased SREBP1 and FASN expression, consistent with the observation of increased SREBP1 levels in human breast cancers [8–10]. Previous studies have established that SREBP1, through induction of FASN and subsequent fatty acids production, regulates PPARγ transactivation [11, 12].
Dietary carbohydrates are digested into glucose, the major source of energy for many tissues. Once transported into cells, glucose is converted into pyruvate through glycolysis and subsequently acetyl Co-enzyme A (acetyl-coA), which is then re-engineered into palmitate, the major fatty acid, by Acetyl-coA carboxylase (ACC), the rate-limiting enzyme, and FAS, the major enzyme, both of which are required for fatty acid biosynthesis. Palmitate is further converted into triglycerides for energy storage and phospholipids, the major components of cell membrane. The key steps in lipogenesis in mammalian hepatocytes are summarized in Fig. 10.1. The enzymatic reactions that govern carbohydrate and lipid metabolism, as well as the allosteric regulation of the activities of these enzymes, also known as the “short-term regulation”, have been elucidated by many pioneering biochemists during the first half of the twentieth century.

The key biochemical reactions and enzymes involved in de novo lipogenesis in mammalian hepatocytes. This process is highly conserved in evolution. Transcription of many metabolic enzymes in this process is directly regulated by several transcription factors, such as PPARγ, SREBP, ChREBP, and LXR etc.
Compared to the short-term quick regulation of the enzymes, however, the transcriptional regulation of the metabolic enzymes in vivo, known as the “long-term regulation” of metabolism, is less well-understood [13]. Since defects in short-term regulation of enzymes are likely detrimental to survival at the cellular or organismal levels, the aberrant regulation of the long-term regulation contributes to a number of major diseases in adults, collectively known as the metabolic syndrome [14–17].
Because of the fundamental importance of FAS, ACC, ACS and SCD1 in regulating lipid metabolism, it is essential to understand the transcriptional regulation of these enzymes in response to physiological stimuli by key transcription factors, including SREBP, PPARγ, liver X receptor (LXR), and carbohydrate-responsive element-binding protein (ChREBP) [13, 18–20]. In this chapter, we summarize the recent advances in studies linking deregulated lipogenesis in cancers, and then focus on our understanding of SREBPs and PPARγ in regulating lipid homeostasis. Finally, we will discuss potential therapeutic approaches to target lipid metabolism in treating cancer.
2 Deregulation of Lipogenic Signaling in Cancer
2.1 Elevated FASN Expression and Enhanced De Novo Fatty Acid Synthesis in Cancer
Most normal human tissues preferentially use circulating lipids for synthesis of new structural lipids, and de novo fatty-acid synthesis is normally suppressed due to the low levels of FASN expression. In cancer cells, however, fatty-acid supply becomes highly dependent on de novo lipogenesis. Deregulation of de novo fatty-acid synthesis leads to cellular fatty-acid accumulation and affects cellular processes, including signal transduction and gene expression.
FASN over-expression occurs in a variety of human cancers [21–25]. In cancer cells, FASN gene expression is upregulated in response to multiple signaling pathways, including growth factors, steroid hormone receptors such as the estrogen receptor alpha (ERα), androgen receptor (AR) and progesterone receptor (PR), as well as oncoproteins including ErbB2, Ras and Akt [9, 25–30]. In addition to the essential role in cancer cell growth and survival, FAS is involved in other phases of cancer development. FAS over-expression confers resistance to adriamycin and mitoxantrone in breast cancer cells [31] and increased lipogenesis and FAS has been reported to be associated with invasive phenotype and cancer metastasis [24, 27, 32–36]. Elevated expression of FASN leads to increased cell proliferation, migration and invasion of prostate cancer cells [27, 32] and FAS inhibition reduces cellular migration and invasiveness [25, 26, 29, 35]. For example, Orlistat, an anti-obesity drug, inhibits FAS function and suppresses endothelial cell proliferation and angiogenesis, suggesting a novel role of FAS in endothelial cell in tumor growth in vivo [37]. It is still unclear how the level and activity of FAS are regulated during tumor progression towards metastasis.
2.2 Stearoyl-CoA Desaturase (SCD) and Cancer
SCD is a regulatory enzyme in lipogenesis, catalyzing the rate-limiting step in the de novo synthesis of monounsaturated fatty acids (MUFAs), mainly palmitic and stearic acids. Increased content of the MUFA products, palmitoleic and oleic acids, occurs in a variety of transformed cells and cancers [38–41], suggesting that the high rate of fatty acid synthesis in cancer is coupled to the conversion of saturated fatty acids (SFAs) into MUFAs. Elevated expression and activity of SCD1, the endoplasmic reticulum-resident Δ9 desaturase that converts SFA into MUFA, has been reported in several types of cancers, including colonic and oesophageal carcinoma, liver cancer, and mammary gland tumor [42–45]. SV40-transformed human lung fibroblasts show significantly increased protein and activity levels of SCD1 compared to their parental normal cell line [46]. This is consistent with a model in which a high rate of MUFA synthesis is required for producing membrane lipids in order to sustain the proliferation of transformed cells. Deficiency or inhibition of SCD1 reduces cell proliferation and anchorage-independent growth, and enhances apoptosis in several different cancer cell types [40, 47]. We and others have shown that SCD1 is a transcriptional target of SREBP1 and PPARγ [48–54].
3 Cellular Regulation of SREBP1 Function
3.1 SREBP1 Signaling in Lipogenesis and Tumorigenesis
SREBPs are a family of transcription factors that control lipid homeostasis by regulating the expression of enzymes required for cholesterol and fatty acids (FAs) synthesis. The three SREBP isoforms, SREBP-1a, SREBP-1c and SREBP-2, have distinct roles in lipid synthesis [55, 56]. In vivo studies using transgenic and knockout mice suggest that SREBP-1c is involved in FA synthesis and insulin-induced glucose metabolism (particularly in lipogenesis), whereas SREBP-2 is relatively specific in controlling cholesterol synthesis. The SREBP-1a isoform is implicated in regulating both cholesterol and FA pathways [57, 58].
Extensive studies in the past two decades have revealed an elegant paradigm to understand how SREBPs maintain the intracellular lipid and cholesterol homeostasis. SREBP transcription factors are synthesized as inactive precursors bound to the endoplasmic reticulum (ER) membranes and their processing is mainly controlled by cellular sterol content: when sterol level decreases, the precursor undergoes a sequential two-step cleavage process to release the NH2-terminal active domain in the nucleus (designated as the nSREBPs), which then activates SREBP target genes whose products are required for the de novo biosynthesis of cholesterol and FAs [59–66]. The major SREBP targets include FASN [12, 67] and stearoyl-CoA desaturase (SCD) [49–51, 68]. This sterol-sensitive process appears to be a major point of regulation for the SREBP-1a and SREBP-2 isoforms, but not for SREBP-1c. Moreover, the SREBP-1c isoform is mainly regulated at the transcriptional level by insulin. The unique regulation and activation properties of each SREBP isoform facilitate the coordinated regulation of lipid and energy metabolism.
3.2 Regulation of the Transcriptional Activity of SREBP1
As summarized above, SREBPs are family of transcription factors that play critical roles in regulating intracellular lipid and cholesterol homeostasis. Using SREBP-1a/-1c as an example, here we focus on the recent advances in our understanding of how SREBP-1 activates lipogenic gene expression and how the transcriptional activity of SREBP is regulated.
3.2.1 Transcription Activation by SREBP
In response to cholesterol depletion, the N-terminus of SREBP that contains the transactivation domain and the basic helix-loop-helix leucine zipper (bHLH-Zip) DNA binding domain, is cleaved from its precursor, which is localized in ER and Golgi apparatus, and then translocates to the nucleus and activates the expression of SREBP-target genes [69, 70]. Through the bHLH-Zip DNA-binding domain, the nuclear SREBP fragments bind to the SREBP-target gene promoters that contain either palindromic E-boxes (CAXXTG) or nonpalindromic sterol regulatory elements (SREs) [71].
The transactivation domain of SREBPs can directly interact with transcription coactivators including CBP/p300, PGC-1β, MED14/DRIP150, and MED15/ARC105 [72]. Recruitment of CBP/p300 via the KIX domain of SREBP may alter chromatin structure through the intrinsic histone acetyltransferase activity of CBP/p300, thereby facilitating gene activation [73]. The interaction between PGC-1β and SREBP is required for SREBP-dependent lipogenic gene expression and contributes to the effect of saturated fat in stimulating hyperlipidemia and atherogenesis [74]. In addition, SREBPs directly interact with the MED14/DRIP150 and the MED15/ARC105 subunits of the Mediator complex in mammals and C. elegans, which provides an elegant model to explain how transcription activator SREBPs interact with the general transcription machinery [73, 75–77]. The interactions between SREBP-1c and MED14 or MED15 are weaker than the interactions between SREBP-1a and MED14 or MED15, which may explain why SREBP-1a is more potent than SREBP-1c in activating gene expression [73, 75]. Since the nuclear SREBPs bind to DNA as homo-dimers, it is unclear whether the two transactivation domains of the SREBP homo-dimer can bind to MED14 and MED15 simultaneously.
3.2.2 Inactivation of SREBP-Mediated Transcription
Because of the fundamental roles of SREBPs in regulating the expression of lipogenic and cholesterogenic genes, the mechanisms that restrain SREBP transactivation are also important. CDK8, the enzymatic subunit of the Mediator complex, directly phosphorylates a conserved Threonine residue in SREBP (Thr402 in SREBP-1c), thereby promoting nuclear SREBP degradation [78]. Consistent with this model, the mutants of CDK8 and its regulatory partner Cyclin C (CycC) in Drosophila larvae, as well as depletion of CDK8 in cultured mammalian cells and mouse liver, display significantly increased expression of SREBP-target genes and dramatic increase of triglyceride accumulation [78]. Feeding and activation of the insulin-signaling pathway can down-regulate CDK8-CycC thus allow the activation of nuclear SRBEP, providing a mechanism for the lipogenic effect of insulin [78]. Together with the previous works on MED14 and MED15 in activating SREBP-dependent gene expression, this recent work on the inhibitory effect of CDK8-CycC on SREBP-regulated de novo lipogenes further highlights the importance of the Mediator complexes in modulating the activation and subsequent degradation of nuclear SREBPs.
Interestingly, GSK3β also negatively regulates SREBP by phosphorylating SREBP-1a at Thr 426 and Ser430 (corresponding to Thr402 and Ser406 in SREBP-1c), thereby providing a docking site for the ubiquitin ligase FBW7 [79–81]. It is still not known whether CDK8 and GSK3β play redundant roles in phosphorylating and thereby promoting SREBP destruction, however, these studies suggest a model to explain how activation of SREBP-dependent transcription is coupled to its degradation. This mechanism is consistent with a general theme by coupling transactivation with their destruction for many transactivators in yeasts and multicellular organisms [82].
Both CDK8 and CycC are amplified, mutated or deleted in a variety of cancers, and CDK8 is identified as an oncoprote in melanoma and colorectal cancers [83]. In addition, the MED12 subunit of the CDK8 module, which is composed of CDK8, CycC, MED12 and MED13, is mutated in prostate cancer, colorectal cancer, and ~70 % of uterine leiomyomas [84–87]. Importantly, MED12, but not MED13, is required for human CDK8 kinase activity [88]. Therefore, although it is still unclear how dysregulation of the CDK8 module contributes to tumorigenesis, it is conceivable that dysregulation of CDK8 submodules may compromise CDK8 activity, thereby potentiating SREBP activity, increasing SREBP target gene expression and promoting lipogenesis in cancer cells. This model may explain the mechanisms underlying aberrantly increased lipogenesis in human cancer cells and provide the rationale for developing pharmaceutical approaches to block de novo lipogenesis in tumor cells.
4 Modulation of PPARγ Activation for Cancer Therapeutics
4.1 The Function of PPARγ in Lipogenesis
Besides SREBPs, the peroxisome proliferator-activator receptor gamma (PPARγ) also plays a critical role in both lipid metabolism and tumorigenesis. The PPARs are ligand-activated nuclear receptors, which include PPARα, PPARγ and PPARδ [89]. Their modular structure resembles other nuclear hormone receptors with an N-terminal activation function 1 (AF-1), a DNA binding domain, and a C-terminal ligand-binding domain that harbors AF2. PPARγ was initially cloned as a transcription factor involved in adipocyte differentiation. Subsequent studies suggested a broad spectrum of PPARγ functions in lipid metabolism, inflammation, atherogenesis, cell differentiation, as well as tumorigenesis. The endogenous PPARγ ligands include derivatives of fatty acids produced through lipogenesis (Fig. 10.1).
PPARγ regulates lipogenesis and adipocyte differentiation, and ectopic PPARγ expression promotes cell adipogenesis in an NIH 3 T3 cell model [90, 91]. Synthetic PPARγ ligands enhance de novo lipogenesis [92, 93], which was further supported by the genetic evidence that PPARγ -/− ES cells and embryonic fibroblasts are resistant to induction of adipogenesis [94, 95]. Aberrant hepatic expression of PPARγ2 stimulates murine hepatic lipogenesis [96, 97]. The screening for PPARγ-regulated genes in mammary epithelial cells, identified that Scd1 (SCD1) as a transcriptional target of PPARγ [98]. SCD1 production of unsaturated fatty acids may thereby serve as PPARγ agonist ligands, providing a feedback loop to PPARγ. Reciprocal up-regulation of PPARγ and SREBP-1 has been reported. Ectopic expression of SREBP-1 in pre-adipocyte 3 T3-L1 cells and hepatic cancer HepG2 cells induced endogenous PPARγ mRNA expression [99]. SREBP-1 activation increased the production of lipids as endogenous ligands for PPARγ, which binds to PPARγ and augments the transcriptional activity of PPARγ [11, 12]. PPARγ, upon ligand binding, up-regulates the expression of INSIG1, the key regulator in the processing of SREBPs [100].
4.2 Contradictory Role of PPARγ in Tumorigenesis
PPARγ has been shown to function either as an oncoprote in, or as a tumor suppressor. PPARγ is expressed in breast, prostate and colonic epithelium and ligand-dependent activation of PPARγ in cell lines derived from these tumors inhibits cellular proliferation [101–105]. Consistent with the role of PPARγ as a tumor suppressor, PPARγ ligand reduced tumorigenesis in the ApcMin model of familial adenomatous polyposis. Carcinogen, N-nitroso-N-methylurea (NMU)-induced mammary tumorigenesis is prevented by PPARγ agonists [106], and 7, 12-dimethylbenz(a)anthracene (DMBA)-induced mammary tumorigenesis was inhibited by troglitazone [107–109]. A chromosomal translocation between PAX8 and PPARγ in follicular thyroid cancer served as a dominant inhibitor of endogenous PPARγ expression [110]. PPARγ expression is reduced in human breast cancers compared with normal breast tissue [111, 112] and PPARγ over expression in tumor cells inhibits cell proliferation in tissue culture [112]. PPARγ levels are reduced in mouse transgenic mammary tumors induced by distinct oncogenes, compared with normal adjacent non tumorous mammary epithelium [111, 112].
In contrast, evidence that PPARγ is an oncogene includes observations that PPARγ ligands promote colonic tumor growth in ApcMin mice when fed a high fat diet [113]. Heterozygous mutations of PPARγ have been detected in 4/55 patients with colon cancer [114]. Although genetic analyses failed to show that PPARγ-deficient mice develop enhanced tumor phenotypes in prostate epithelium induced by the SV40 large-T antigen oncogene [113]. A constitutively active mutant of PPARγ (PγCA) enhanced ErbB2-induced tumor in vivo in immune-competent animals (Fig. 10.2) and in transgenic mice [115]. PγCA promoted ErbB2-induced tumor growth in immune-competent animals. Increased angiogenesis is associated with enhanced tumor growth in vivo [116]. Collectively, these studies suggest cell-type specific functions of PPARγ in the tumor induction versus inhibition.

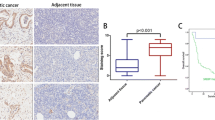
PγCA promotes tumor growth in vivo. (a) NAFA cells transduced with MSCV-IRES-GFP vector encoding either PPARγ, PγCA, or empty vector were implanted into FVB by injecting 2 × 106 cells subcutaneously. Tumor growth was measured every 3 days by digital caliper and tumor volume was calculated. (b) Tumor volumes were logarithm-transformed and analyzed using a linear mixed model. Separate slope and intercepts were computed for each group (GFP, PPARγ, and PγCA), then compared across groups using a global test followed by pair-wise comparisons via linear contrasts (This figure was reproduced from our previous publication [116])
4.3 Can PPARγ Be Targeted to Block the Tumor Growth?
In cell culture, PPARγ expression and/or activation repressed tumor cell growth by inhibiting cell proliferation, promoting apoptotic and autophagic cell death, and inducing terminal differentiation of cancer cells [112, 117, 118]. In whole animal studies, the picture appears more complex, which is consistent with the importance of heterotypic signals in cancer progression, and the presence of PPARγ in a variety of cell types including the inflammatory system. Clinical trials have been undertaken in a variety of tumor types including liposarcomas, prostate, pancreatic, colorectal, breast, thyroid, head and neck cancers, as well as melanoma and leukemia [119]. Overall, PPARγ agonists failed to yield positive clinical outcome in most cancer types. PPARγ is increased in ERα-negative breast cancer, but reduced expression in ERα-positive breast cancers [120]. Breast cancer genetic subtypes (Luminal A, Luminal B, Triple negative/basal-like, HER2 subtypes [121–123]) may have different response to PPARγ ligands. Our analysis of PPARG gene expression in a combined dataset comprising of over 2,000 breast cancers [124] showed a strong heterogeneous distribution of PPARG expression among the subtypes (unpublished data). Consistent with our previous IHC result showing reduced PPARγ expression in breast cancer comparing to normal breast tissue, this analysis demonstrated that the gene expression of PPARG was also reduced (data not shown). The higher PPARG expression predicts a better clinical outcome, which again holds the promise that PPARγ could serve as a therapeutic target. Given the variability in PPARγ expression in patient populations, clinical trials using PPARγ expression or function as a companion diagnostic may be warranted.
5 Conclusions and Future Directions
Given the importance of lipogenesis in cancer development, targeting lipogenic signaling, particularly lipogenic enzymes, is an attractive strategy. The inhibitors of the rate-limiting or key lipogenic enzymes, including HMGCR, ACC, FASN, and SCD, are summarized in Table 10.1. In addition to inhibiting these lipogenic enzymes, pharmaceutical inhibition of SREBPs and PPARγ, may also be effective.
Taken together, these studies suggest that dysregulated lipogenic signaling in cancer is required for oncogenic transformation, thus targeting the dysregulated lipogenesis in câncer cells may represent an attractive therapeutic approach. Current research efforts have been focused on repressing the activity of lipogenic enzymes (such as FASN, HMG-CoA reductase, ACC, ACLY, and SCD). Future studies are required to provide a deeper understanding of the following three major aspects. First, it would be important to understand how alterations in molecular mechanisms of lipogenic signaling occur in cancer. Second, a compendium of metabolic profiling in different cancer types and subtypes may allow for more accurate patient selection for specific lipogenic pathway targeted therapies. Third, it may be important to simultaneously target multiple lipogenic factors rather than a single molecule, to ensure therapy effectiveness.
References
Swinnen JV, Brusselmans K, Verhoeven G (2006) Increased lipogenesis in cancer cells: new players, novel targets. Curr Opin Clin Nutr Metab Care 9:358
Yang YA (2003) Regulation of fatty acid synthase expression in breast cancer by sterol regulatory element binding protein-1c. Exp Cell Res 282:132
Ettinger SL (2004) Dysregulation of sterol response element-binding proteins and downstream effectors in prostate cancer during progression to androgen independence. Cancer Res 64:2212
Yahagi N (2005) Co-ordinate activation of lipogenic enzymes in hepatocellular carcinoma. Eur J Cancer 41:1316
Calvisi DF (2011) Increased lipogenesis, induced by AKT-mTORC1-RPS6 signaling, promotes development of human hepatocellular carcinoma. Gastroenterology 140:1071
Yamashita T (2009) Activation of lipogenic pathway correlates with cell proliferation and poor prognosis in hepatocellular carcinoma. J Hepatol 50:100
Li JN, Mahmoud MA, Han WF, Ripple M, Pizer ES (2000) Sterol regulatory element-binding protein-1 participates in the regulation of fatty acid synthase expression in colorectal neoplasia. Exp Cell Res 261:159
Furuta E (2008) Fatty acid synthase gene is up-regulated by hypoxia via activation of Akt and sterol regulatory element binding protein-1. Cancer Res 68:1003
Yoon S (2007) Up-regulation of acetyl-CoA carboxylase alpha and fatty acid synthase by human epidermal growth factor receptor 2 at the translational level in breast cancer cells. J Biol Chem 282:26122
Yang YA, Han WF, Morin PJ, Chrest FJ, Pizer ES (2002) Activation of fatty acid synthesis during neoplastic transformation: role of mitogen-activated protein kinase and phosphatidylinositol 3-kinase. Exp Cell Res 279:80
Kim JB, Wright HM, Wright M, Spiegelman BM (1998) ADD1/SREBP1 activates PPAR gamma through the production of endogenous ligand. Proc Natl Acad Sci USA 95:4333
Kim JB, Spiegelman BM (1996) ADD1/SREBP1 promotes adipocyte differentiation and gene expression linked to fatty acid metabolism. Genes Dev 10:1096
Desvergne B, Michalik L, Wahli W (2006) Transcriptional regulation of metabolism. Physiol Rev 86:465
Lusis AJ, Attie AD, Reue K (2008) Metabolic syndrome: from epidemiology to systems biology. Nat Rev Genet 9:819
Eckel RH, Grundy SM, Zimmet PZ (2005) The metabolic syndrome. Lancet 365:1415
Cornier MA et al (2008) The metabolic syndrome. Endocr Rev 29:777
Braun S, Bitton-Worms K, LeRoith D (2011) The link between the metabolic syndrome and cancer. Int J Biol Sci 7:1003
Postic C, Dentin R, Denechaud PD, Girard J (2007) ChREBP, a transcriptional regulator of glucose and lipid metabolism. Annu Rev Nutr 27:179
Laurencikiene J, Ryden M, Liver X (2012) Receptors and fat cell metabolism. Int J Obes (Lond). Dec; 36(12):1494–502
Horton JD, Goldstein JL, Brown MS (2002) SREBPs: activators of the complete program of cholesterol and fatty acid synthesis in the liver. J Clin Invest 109:1125
Mashima T, Seimiya H, Tsuruo T (2009) De novo fatty-acid synthesis and related pathways as molecular targets for cancer therapy. Br J Cancer 100:1369
Little JL, Kridel SJ (2008) Fatty acid synthase activity in tumor cells. Subcell Biochem 49:169
Kridel SJ, Lowther WT, Pemble CW (2007) Fatty acid synthase inhibitors: new directions for oncology. Expert Opin Investig Drugs 16:1817
Menendez JA, Lupu R (2007) Fatty acid synthase and the lipogenic phenotype in cancer pathogenesis. Nat Rev Cancer 7:763
Lupu R, Menendez JA (2006) Targeting fatty acid synthase in breast and endometrial cancer: an alternative to selective estrogen receptor modulators? Endocrinology 147:4056
Grunt TW et al (2009) Interaction between fatty acid synthase- and ErbB-systems in ovarian cancer cells. Biochem Biophys Res Commun 385:454
Migita T et al (2009) Fatty acid synthase: a metabolic enzyme and candidate oncogene in prostate cancer. J Natl Cancer Inst 101:519
Vazquez-Martin A, Ortega-Delgado FJ, Fernandez-Real JM, Menendez JA (2008) The tyrosine kinase receptor HER2 (erbB-2): from oncogenesis to adipogenesis. J Cell Biochem 105:1147
Uddin S et al (2008) Fatty acid synthase and AKT pathway signaling in a subset of papillary thyroid cancers. J Clin Endocrinol Metab 93:4088
Swinnen JV et al (2000) Stimulation of tumor-associated fatty acid synthase expression by growth factor activation of the sterol regulatory element-binding protein pathway. Oncogene 19:5173
Liu H, Liu Y, Zhang JT (2008) A new mechanism of drug resistance in breast cancer cells: fatty acid synthase overexpression-mediated palmitate overproduction. Mol Cancer Ther 7:263
Fiorentino M et al (2008) Overexpression of fatty acid synthase is associated with palmitoylation of Wnt1 and cytoplasmic stabilization of beta-catenin in prostate cancer. Lab Invest 88:1340
Ashida S et al (2004) Molecular features of the transition from prostatic intraepithelial neoplasia (PIN) to prostate cancer: genome-wide gene-expression profiles of prostate cancers and PINs. Cancer Res 64:5963
Vazquez-Martin A, Colomer R, Brunet J, Lupu R, Menendez JA (2008) Overexpression of fatty acid synthase gene activates HER1/HER2 tyrosine kinase receptors in human breast epithelial cells. Cell Prolif 41:59
Carvalho MA et al (2008) Fatty acid synthase inhibition with orlistat promotes apoptosis and reduces cell growth and lymph node metastasis in a mouse melanoma model. Int J Cancer 123:2557
Montgomery RB et al (2008) Maintenance of intratumoral androgens in metastatic prostate cancer: a mechanism for castration-resistant tumor growth. Cancer Res 68:4447
Browne CD, Hindmarsh EJ, Smith JW (2006) Inhibition of endothelial cell proliferation and angiogenesis by orlistat, a fatty acid synthase inhibitor. FASEB J J20:2027
Bagnato C, Igal RA (2003) Overexpression of diacylglycerol acyltransferase-1 reduces phospholipid synthesis, proliferation, and invasiveness in simian virus 40-transformed human lung fibroblasts. J Biol Chem 278:52203
Scaglia N, Chisholm JW, Igal RA (2009) Inhibition of stearoylCoA desaturase-1 inactivates acetyl-CoA carboxylase and impairs proliferation in cancer cells: role of AMPK. PLoS One 4:e6812
Scaglia N, Igal RA (2008) Inhibition of stearoyl-CoA desaturase 1 expression in human lung adenocarcinoma cells impairs tumorigenesis. Int J Oncol 33:839
Tsubura A, Yuri T, Yoshizawa K, Uehara N, Takada H (2009) Role of fatty acids in malignancy and visual impairment: epidemiological evidence and experimental studies. Histol Histopathol 24:223
Li J et al (1994) Partial characterization of a cDNA for human stearoyl-CoA desaturase and changes in its mRNA expression in some normal and malignant tissues. Int J Cancer 57:348
Paton CM, Ntambi JM (2008) Role of stearoyl-CoA desaturase-1 expression in cancer proliferation. FASEB J 22:794.1
Lu J, Pei H, Kaeck M, Thompson HJ (1997) Gene expression changes associated with chemically induced rat mammary carcinogenesis. Mol Carcinog 20:204
Falvella FS et al (2002) Stearoyl-CoA desaturase 1 (Scd1) gene overexpression is associated with genetic predisposition to hepatocarcinogenes is in mice and rats. Carcinogenesis 23:1933
Scaglia N, Caviglia JM, Igal RA (2005) High stearoyl-CoA desaturase protein and activity levels in simian virus 40 transformed-human lung fibroblasts. Biochim Biophys Acta 1687:141
Morgan-Lappe SE et al (2007) Identification of Ras-related nuclear protein, targeting protein for xenopus kinesin-like protein 2, and stearoyl-CoA desaturase 1 as promising cancer targets from an RNAi-based screen. Cancer Res 67:4390
Shimano H et al (1999) Sterol regulatory element-binding protein-1 as a key transcription factor for nutritional induction of lipogenic enzyme genes. J Biol Chem 274:35832
Ntambi JM (1999) Regulation of stearoyl-CoA desaturase by polyunsaturated fatty acids and cholesterol. J Lipid Res 40:1549
Tabor DE, Kim JB, Spiegelman BM, Edwards PA (1999) Identification of conserved cis-elements and transcription factors required for sterol-regulated transcription of stearoyl-CoA desaturase 1 and 2. J Biol Chem 274:20603
Shimomura I, Shimano H, Korn BS, Bashmakov Y, Horton JD (1998) Nuclear sterol regulatory element-binding proteins activate genes responsible for the entire program of unsaturated fatty acid biosynthesis in transgenic mouse liver. J Biol Chem 273:35299
Tabor DE, Kim JB, Spiegelman BM, Edwards PA (1998) Transcriptional activation of the stearoyl-CoA desaturase 2 gene by sterol regulatory element-binding protein/adipocyte determination and differentiation factor 1. J Biol Chem 273:22052
Kim YC, Gomez FE, Fox BG, Ntambi JM (2000) Differential regulation of the stearoyl-CoA desaturase genes by thiazolidinediones in 3T3-L1 adipocytes. J Lipid Res 41:1310
Yao-Borengasser A et al (2008) Stearoyl-coenzyme a desaturase 1 gene expression increases after pioglitazone treatment and is associated with peroxisomal proliferator-activated receptor-gamma responsiveness. J Clin Endocrinol Metab 93:4431
Hua X, Wu J, Goldstein JL, Brown MS, Hobbs HH (1995) Structure of the human gene encoding sterol regulatory element binding protein-1 (SREBF1) and localization of SREBF1 and SREBF2 to chromosomes 17p11.2 And 22q13. Genomics 25:667
Edwards PA, Tabor D, Kast HR, Venkateswaran A (2000) Regulation of gene expression by SREBP and SCAP. Biochim Biophys Acta 1529:103
Shimano H et al (1996) Overproduction of cholesterol and fatty acids causes massive liver enlargement in transgenic mice expressing truncated SREBP-1a. J Clin Invest 98:1575
Horton JD et al (1998) Activation of cholesterol synthesis in preference to fatty acid synthesis in liver and adipose tissue of transgenic mice overproducing sterol regulatory element-binding protein-2. J Clin Invest 101:2331
Wang X et al (1996) Cleavage of sterol regulatory element binding proteins (SREBPs) by CPP32 during apoptosis. EMBO J J15:1012
Wang X, Sato R, Brown MS, Hua X, Goldstein JL (1994) SREBP-1, a membrane-bound transcription factor released by sterol-regulated proteolysis. Cell 77:53
Hua X et al (1993) SREBP-2, a second basic-helix-loop-helix-leucine zipper protein that stimulates transcription by binding to a sterol regulatory element. Proc Natl Acad Sci USA A90:11603
Yokoyama C et al (1993) SREBP-1, a basic-helix-loop-helix-leucine zipper protein that controls transcription of the low density lipoprotein receptor gene. Cell 75:187
Briggs MR, Yokoyama C, Wang X, Brown MS, Goldstein JL (1993) Nuclear protein that binds sterol regulatory element of low density lipoprotein receptor promoter. I. Identification of the protein and delineation of its target nucleotide sequence. J Biol Chem 268:14490
Wang X et al (1993) Nuclear protein that binds sterol regulatory element of low density lipoprotein receptor promoter. II. Purification and characterization. J Biol Chem 268:14497
Shimano H (2001) Sterol regulatory element-binding proteins (SREBPs): transcriptional regulators of lipid synthetic genes. Prog Lipid Res 40:439
Horton JD, Shimomura I (1999) Sterol regulatory element-binding proteins: activators of cholesterol and fatty acid biosynthesis. Curr Opin Lipidol 10:143
Liang G et al (2002) Diminished hepatic response to fasting/refeeding and liver X receptor agonists in mice with selective deficiency of sterol regulatory element-binding protein-1c. J Biol Chem 277:9520
Bene H, Lasky D, Ntambi JM (2001) Cloning and characterization of the human stearoyl-CoA desaturase gene promoter: transcriptional activation by sterol regulatory element binding protein and repression by polyunsaturated fatty acids and cholesterol. Biochem Biophys Res Commun 284:1194
Goldstein JL, DeBose-Boyd RA, Brown MS (2006) Protein sensors for membrane sterols. Cell 124:35
Jeon TI, Osborne TF (2012) SREBPs: metabolic integrators in physiology and metabolism. Trends Endocrinol Metab 23:65
Kim JB et al (1995) Dual DNA binding specificity of ADD1/SREBP1 controlled by a single amino acid in the basic helix-loop-helix domain. Mol Cell Biol 15:2582
Bengoechea-Alonso MT, Ericsson J (2007) SREBP in signal transduction: cholesterol metabolism and beyond. Curr Opin Cell Biol 19:215
Toth JI, Datta S, Athanikar JN, Freedman LP, Osborne TF (2004) Selective coactivator interactions in gene activation by SREBP-1a and -1c. Mol Cell Biol 24:8288
Lin J et al (2005) Hyperlipidemic effects of dietary saturated fats mediated through PGC-1beta coactivation of SREBP. Cell 120:261
Yang F et al (2006) An ARC/mediator subunit required for SREBP control of cholesterol and lipid homeostasis. Nature 442:700
Osborne TF, Espenshade PJ (2009) Evolutionary conservation and adaptation in the mechanism that regulates SREBP action: what a long, strange tRIP it’s been. Genes Dev 23:2578
Taubert S, Van Gilst MR, Hansen M, Yamamoto KR (2006) A mediator subunit, MDT-15, integrates regulation of fatty acid metabolism by NHR-49-dependent and -independent pathways in C. Elegans. Genes Dev 20:1137
Zhao X et al (2012) Regulation of lipogenesis by cyclin-dependent kinase 8-mediated control of SREBP-1. J Clin Inves 122:2417
Punga T, Bengoechea-Alonso MT, Ericsson J (2006) Phosphorylation and ubiquitination of the transcription factor sterol regulatory element-binding protein-1 in response to DNA binding. J Biol Chem 281:25278
Sundqvist A et al (2005) Control of lipid metabolism by phosphorylation-dependent degradation of the SREBP family of transcription factors by SCF(Fbw7). Cell Metab 1:379
Kim KH et al (2004) Regulatory role of glycogen synthase kinase 3 for transcriptional activity of ADD1/SREBP1c. J Biol Chem 279:51999
Tansey WB (2001) Transcriptional activation: risky business. Genes Dev 15:1045
Xu W, Ji JY (2011) Dysregulation of CDK8 and cyclin C in tumorigenesis. J Genet Genomics 38:439
Kampjarvi K et al (2012) Somatic MED12 mutations in uterine leiomyosarcoma and colorectal cancer. Br J Cancer 107:1761
Barbieri CE et al (2012) Exome sequencing identifies recurrent SPOP, FOXA1 and MED12 mutations in prostate cancer. Nat Genet 44:685
McGuire MM, et al (2012) Whole exome sequencing in a random sample of North American women with leiomyomas identifies MED12 mutations in majority of uterine leiomyomas. PLoS One 7, e33251
Makinen N et al (2011) MED12, the mediator complex subunit 12 gene, is mutated at high frequency in uterine leiomyomas. Science 334:252
Knuesel MT, Meyer KD, Donner AJ, Espinosa JM, Taatjes DJ (2009) The human CDK8 subcomplex is a histone kinase that requires Med12 for activity and can function independently of mediator. Mol Cell Biol 29:650
Rosen ED, Spiegelman BM (2001) PPARg: a nuclear regulator of metabolism, differentiation, and cell growth. J Biol Chem 276:37731
Tontonoz P, Hu E, Spiegelman BM (1994) Stimulation of adipogenesis in fibroblasts by PPARg2, a lipid-activated transcription factor. Cell 79:1147
Yu S et al (2003) Adipocyte-specific gene expression and adipogenic steatosis in the mouse liver due to peroxisome proliferator-activated receptor 1 (PPAR1) overexpression. J Biol Chem 278:498
Teruel T, Hernandez R, Rial E, Martin-Hidalgo A, Lorenzo M (2005) Rosiglitazone up-regulates lipoprotein lipase, hormone-sensitive lipase and uncoupling protein-1, and down-regulates insulin-induced fatty acid synthase gene expression in brown adipocytes of Wistar rats. Diabetologia 48:1180
Watkins SM, Reifsnyder PR, Pan HJ, German JB, Leiter EH (2002) Lipid metabolome-wide effects of the PPAR gamma agonist rosiglitazone. J Lipid Res 43:1809
Kubota N et al (1999) PPAR gamma mediates high-fat diet-induced adipocyte hypertrophy and insulin resistance. Mol Cell 4:597
Rosen ED et al (1999) PPAR gamma is required for the differentiation of adipose tissue in vivo and in vitro. Mol Cell 4:611
Poritsanos NJ, Wong D, Vrontakis ME, Mizuno TM (2008) Regulation of hepatic PPARgamma2 and lipogenic gene expression by melanocortin. Biochem Biophys Res Commun 376:384
Schroeder-Gloeckler JM et al (2007) CCAAT/enhancer-binding protein beta deletion reduces adiposity, hepatic steatosis, and diabetes in lepr(db/db) mice. J Biol Chem 282:15717
J. Zhou et al (2009) PPARgamma activation induces autophagy in breast cancer cells. Int J Biochem Cell Biol Nov; 41(11):2334–42
Fajas L et al (1999) Regulation of peroxisome proliferator-activated receptor gamma expression by adipocyte differentiation and determination factor 1/sterol regulatory element binding protein 1: implications for adipocyte differentiation and metabolism. Mol Cell Biol 19:5495
Kast-Woelbern HR et al (2004) Rosiglitazone induction of insig-1 in white adipose tissue reveals a novel interplay of peroxisome proliferator-activated receptor gamma and sterol regulatory element-binding protein in the regulation of adipogenesis. J Biol Chem 279:23908
Brockman JA, Gupta RA, Dubois RN (1998) Activation of PPARgamma leads to inhibition of anchorage-independent growth of human colorectal cancer cells. Gastroenterology 115:1049
Coopman PJ, Do MT, Thompson EW, Mueller SC (1998) Phagocytosis of cross-linked gelatin matrix by human breast carcinoma cells correlates with their invasive capacity. Clin Cancer Res 4:507
Elstner E et al (1998) Ligands for peroxisome proliferator-activated receptorg and retinoic acid receptor inhibit growth and induce apoptosis of human breast cancer cells in vitro and in BNX mice. Proc Natl Acad Sci USA 95:8806
Ricote M et al (1998) Expression of the peroxisome proliferator-activated receptor gamma (PPARgamma) in human atherosclerosis and regulation in macrophages by colony stimulating factors and oxidized low density lipoprotein. Proc Natl Acad Sci USA 95:7614
Sarraf P et al (1998) Differentiation and reversal of malignant changes in colon cancer through PPARgamma. Nat Med 4:1046
Suh N et al (1999) A new ligand for the peroxisome proliferator-activated receptor-gamma (PPAR-gamma), GW7845, inhibits rat mammary carcinogenesis. Cancer Res 59:5671
Mehta RG, Williamson E, Patel MK, Koeffler HP (2000) A ligand of peroxisome proliferator-activated receptor gamma, retinoids, and prevention of preneoplastic mammary lesions. J Natl Cancer Inst 92:418
Pighetti GM et al (2001) Therapeutic treatment of DMBA-induced mammary tumors with PPAR ligands. Anticancer Res 21:825
Nicol CJ et al (2004) PPARgamma influences susceptibility to DMBA-induced mammary, ovarian and skin carcinogenesis. Carcinogenesis 25:1747
Kroll TG et al (2000) PAX8-PPARgamma1 Fusion oncogene in human thyroid carcinoma. Science 289:1357
Wang C et al (2003) Cyclin D1 repression of peroxisome proliferator-activated receptor gamma expression and transactivation. Mol Cell Biol 23:6159
Wang C et al (2001) Inhibition of cellular proliferation through IkappaB kinase-independent and peroxisome proliferator-activated receptor gamma-dependent repression of cyclin D1. Mol Cell Biol 21:3057
Saez E et al (1998) Activators of the nuclear receptor PPARgamma enhance colon polyp formation. Nat Med 4:1058
Sarraf P et al (1999) Loss-of-function mutations in PPAR gamma associated with human colon cancer. Mol Cell 3:799
Saez E et al (2004) PPAR gamma signaling exacerbates mammary gland tumor development. Genes Dev 18:528
Tian L et al (2009) Activating peroxisome proliferator-activated receptor gamma mutant promotes tumor growth in vivo by enhancing angiogenesis. Cancer Res 69:9236
Elstner E et al (1998) Ligands for peroxisome proliferator-activated receptor gamma and retinoic acid receptor inhibit growth and induce apoptosis of human breast cancer cells in vitro and in BNX mice. Proc Natl Acad Sci USA A95:8806
Zhou J et al (2009) PPAR gamma activation induces autophagy in breast cancer cells. Int J Biochem Cell Biol 41:2334
Vamecq J et al (2012) PPARs: interference with Warburg’ effect and clinical anticancer trials. PPAR Res 2012:304760
Mueller E et al (1998) Terminal differentiation of human breast cancer through PPAR gamma. Mol Cell 1:465
Perou CM et al (2000) Molecular portraits of human breast tumours. Nature 406:747
Sorlie T (2004) Molecular portraits of breast cancer: tumour subtypes as distinct disease entities. Eur J Cancer 40:2667
Sorlie T et al (2001) Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc Natl Acad Sci USA A98:10869
Casimiro MC et al (2012) ChIP sequencing of cyclin D1 reveals a transcriptional role in chromosomal instability in mice. J Clin Invest 122:833
Osmak M (2012) Statins and cancer: current and future prospects. Cancer Lett 324:1
Beckers A et al (2007) Chemical inhibition of acetyl-CoA carboxylase induces growth arrest and cytotoxicity selectively in cancer cells. Cancer Res 67:8180
Jump B, Torres-Gonzalez M, Olson LK, Soraphen A (2011) An inhibitor of acetyl CoA carboxylase activity, interferes with fatty acid elongation. Biochem Pharmacol 81:649
Sugimoto Y et al (2007) A novel acetyl-CoA carboxylase inhibitor reduces de novo fatty acid synthesis in HepG2 cells and rat primary hepatocytes. Arch Biochem Biophys 468:44
McCune SA, Harris RA (1979) Mechanism responsible for 5-(tetradecyloxy)-2-furoic acid inhibition of hepatic lipogenesis. J Biol Chem 254:10095
Harwood HJ Jr et al (2003) Isozyme-nonselective N-substituted bipiperidylcarboxamide acetyl-CoA carboxylase inhibitors reduce tissue malonyl-CoA concentrations, inhibit fatty acid synthesis, and increase fatty acid oxidation in cultured cells and in experimental animals. J Biol Chem 278:37099
Kuhajda FP et al (1994) Fatty acid synthesis: a potential selective target for antineoplastic therapy. Proc Natl Acad Sci USA A91:6379
Pizer ES et al (1996) Inhibition of fatty acid synthesis delays disease progression in a xenograft model of ovarian cancer. Cancer Res 56:1189
Pizer ES, Chrest FJ, DiGiuseppe JA, Han WF (1998) Pharmacological inhibitors of mammalian fatty acid synthase suppress DNA replication and induce apoptosis in tumor cell lines. Cancer Res 58:4611
Pizer ES et al (2000) Malonyl-coenzyme-a is a potential mediator of cytotoxicity induced by fatty-acid synthase inhibition in human breast cancer cells and xenografts. Cancer Res 60:213
Zhou W et al (2007) Fatty acid synthase inhibition activates AMP-activated protein kinase in SKOV3 human ovarian cancer cells. Cancer Res 67:2964
Kridel SJ, Axelrod F, Rozenkrantz N, Smith JW (2004) Orlistat is a novel inhibitor of fatty acid synthase with antitumor activity. Cancer Res 64:2070
Brusselmans K, De Schrijver E, Heyns W, Verhoeven G, Swinnen JV (2003) Epigallocatechin-3-gallate is a potent natural inhibitor of fatty acid synthase in intact cells and selectively induces apoptosis in prostate cancer cells. Int J Cancer 106:856
Oliveras G et al (2010) Novel anti-fatty acid synthase compounds with anti-cancer activity in HER2+ breast cancer. Ann N Y Acad Sci 1210:86
Vazquez MJ et al (2008) Discovery of GSK837149A, an inhibitor of human fatty acid synthase targeting the beta-ketoacyl reductase reaction. FEBS J J275:1556
Oballa RM et al (2011) Development of a liver-targeted stearoyl-CoA desaturase (SCD) inhibitor (MK-8245) to establish a therapeutic window for the treatment of diabetes and dyslipidemia. J Med Chem 54:5082
Uto Y et al (2011) Discovery of novel SCD1 inhibitors: 5-alkyl-4,5-dihydro-3H-spiro[1,5-benzoxazepine-2,4′-piperidine] analogs. Eur J Med Chem 46:1892
Kamisuki S et al (2009) A small molecule that blocks fat synthesis by inhibiting the activation of SREBP. Chem Biol 16:882
Kamisuki S et al (2011) Synthesis and evaluation of diarylthiazole derivatives that inhibit activation of sterol regulatory element-binding proteins. J Med Chem 54:4923
Das BC, Zhao X, Tang XY, Yang F (2011) Design, synthesis and biological study of pinacolyl boronate-substituted stilbenes as novel lipogenic inhibitors. Bioorg Med Chem Lett 21:5638
Wahli W, Michalik L (2012) PPARs at the crossroads of lipid signaling and inflammation. Trends Endocrinol Metab 23:351
Acknowledgements
This work was supported in part by R01CA70896, R01CA75503, and R01CA86072 (R.G.P.). The Kimmel Cancer Center was supported by the NIH Cancer Center Core Grant P30CA56036 (R.G.P). This work is also funded in part from the Dr. Ralph and Marian C. Falk Medical Research Trust (R.G.P.), the Breast Cancer Research Foundation (R.G.P) and a grant from the Pennsylvania Department of Health (R.G.P., C.W.), the startup funds from Texas A&M Health Science Center and a grant from the American Heart Association (J.Y.J.).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer Science+Business Media New York
About this chapter
Cite this chapter
Wang, C., Ji, JY., Tian, L., Pestell, R.G. (2014). Transcriptional Regulation of Lipogenesis as a Therapeutic Target for Cancer Treatment. In: Kumar, R. (eds) Nuclear Signaling Pathways and Targeting Transcription in Cancer. Cancer Drug Discovery and Development. Humana Press, New York, NY. https://doi.org/10.1007/978-1-4614-8039-6_10
Download citation
DOI: https://doi.org/10.1007/978-1-4614-8039-6_10
Published:
Publisher Name: Humana Press, New York, NY
Print ISBN: 978-1-4614-8038-9
Online ISBN: 978-1-4614-8039-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)