
Abstract
Short- and long-distance transport in woody plants is performed within two different systems of apoplasm and symplasm. Although the apoplasmic route of transport via dead conductive elements dominates in trees, the presence of interconnected living cells in xylem suggests the involvement of the symplasmic route in the transportation and communication processes occurring in wood. In this chapter, an attempt will be made to demonstrate and review the numerous functions of living xylem cells and the role of symplasmic transport in the secondary xylem of seed plants. The anatomical and ultrastructural characteristics of xylem parenchyma, reflecting the participation of living cells in the accumulation and distribution of stored compounds via symplasmic routes, will be presented. The involvement of living xylem parenchyma cells in the aspects of sugar transport and exchange at the symplasm/apoplasm interface, embolism repair, defense mechanisms against vascular pathogenic infection, and differentiation processes of xylem elements will be reviewed to emphasize the crucial impact of symplasmic transport and communication processes, via plasmodesmata, on integration and proper functioning of trees.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
- Axial parenchyma
- Contact cells
- Embolism repair
- Isolation cells
- Sapwood
- Secondary xylem
- Storage parenchyma
- Sugar influx
- Sugar efflux
- Symplasm/apoplasm interface
- Xylem rays
4.1 Introduction
Secondary xylem is a heterogeneous tissue composed of both dead and living cells. Conductive elements like tracheids and vessels, as well as strengthening dead fibers, form the apoplasmic system, which itself is responsible for the long-distance transport of water and solutes in woody plants. In addition, a counterpart system for short- and long-distance symplasmic transport is formed by the living fibers and parenchymatous cells of the secondary xylem. Both the apoplasmic and symplasmic systems are closely integrated and cannot function alone. Thus, transport in woody plants has to be considered as a whole, as a union of apoplasmic and symplasmic parts. Fast apoplasmic transport prevails in the secondary xylem and thus is studied intensively (Holbrook and Zwieniecki 2005). However, the presence of a spatial and continuous system of different types of living cells in the secondary xylem region enables symplasmic communication in this tissue and provides efficient routes for short- and long-distance symplasmic transport in woody plants. Living xylem cells are involved in different physiological and developmental processes, where more careful control over intercellular transport and communication is necessary (Barnett 2006). For example, they provide routes for the transport of photoassimilate and solutes between living cells of the xylem, phloem, and cambial regions and between living and dead cells at the symplasm/apoplasm interface (Sauter 1982; Sauter and Kloth 1986; Améglio et al. 2004). They produce chemical weapons during vascular pathogen infection (Benhamou 1995), influence the differentiation processes of xylem elements (Lachaud and Maurousset 1996), and participate in the refilling of embolized vessels (Salleo et al. 2004; Améglio et al. 2004). Although many of the processes involving living xylem cells are rather still unknown, we are slowly beginning to understand the significance of symplasm in the regulation and integration of woody plant function.
4.2 Living Cells Are Essential Components of Wood of Seed Plants
All the elements of secondary conductive tissues are produced by cambium, the lateral meristem of woody plants. Cambium is composed of two different types of initials—ray and fusiform—whose division activity gives rise to the radial and axial system of the secondary xylem and phloem elements. Thus, the living cells in wood, as derivatives of cambial cells, are incorporated into both of these two systems of radial and axial secondary conductive tissues (Larson 1994).
Additionally, two different regions can be distinguished within the volume of the secondary xylem. The outer wood region in the vicinity of cambium, where living xylem cells occur, is called sapwood. The inner wood region, close to the center of the trunk, which does not contain living cells and in which reserve storage materials are removed or converted into secondary metabolites, is called heartwood (Carquist 1988; Romberger et al. 1993).
4.2.1 The Living Cells of the Radial System
The radial system in wood is composed of rays—groups of cells that extend radially from the cambium in two directions: to the pith (centripetally) forming xylem rays and towards the cortex (centrifugally) creating phloem rays. Xylem rays are usually formed by living parenchymatous cells, although different specialized cells can additionally occur in some taxa of gymno- and angiosperms. For instance, rays can include ray tracheids (Bannan 1934, 1941), radial vessels (Botosso and Gomes 1982), radial fibers (Lev-Yadun 1994), as well as radially orientated canals or cavities filled with resin, mucilage, and oils (Carquist 1988; IAWA 1989; LaPasha and Wheeler 1990).
Rays can be classified as uniseriate, when they are only one cell wide, or multiseriate, where two or more cells are present in the widest point of the ray viewed at the tangential plane (Fig. 4.1). Another feature describing the rays from an anatomical point of view is their height, which is defined by the number of cells located between both (apical and bottom) ends of a particular ray along vertical line (Fig. 4.1; Carquist 1988; IAWA 1989). The living parenchymatous ray cells of angiosperm can be also subdivided into three different morphological types based on the shape of ray cells at the radial plane: upright when the cells are axially higher than wide, procumbent when the radial dimension of the cell is longer than its height, and square when cells are as tall as wide. If the ray is composed of only one cell type, e.g., exclusively procumbent, it is a homogenous ray. If other morphological types of parenchymatous ray cells or additional specialized elements, such as ray tracheids, are present, the ray is classified as heterogeneous (Fig. 4.1; Metcalfe and Chalk 1983; Carquist 1988; IAWA 1989). Finally, the xylem ray cells neighboring the tracheary elements can be divided into two categories (Fig. 4.2): contact cells which commonly have pits with tracheary elements and isolation cells that lack pit connections with tracheary elements or have sparse pitting (Höll 1975; Czaninski 1977; Gregory 1978; Murakami et al. 1999).

The radial system of xylem parenchyma in seed plants. (a–d) Different types of xylem rays and their arrangement in the wood of gymnosperms. (a–c) Transverse (a), tangential (b), and radial (c) sections of Abies sp. wood. Homogenous rays, one cell wide (uniseriate), are frequent. The cell shape of some ray cells is outlined by solid lines; a double-head solid line indicates the height of the xylem ray. (d) Tangential section of Picea sp. wood. A heterogeneous multiseriate ray, containing a resin canal (indicated by an arrowhead), is visible between homogeneous uniseriate xylem rays. (e–j) Different types of xylem rays and their arrangement in the wood of angiosperms. (e) Diffuse-porous wood, containing multiple vessels with approximately similar dimensions, is shown in a transverse section of Populus sp. Frequent narrow xylem rays are present. (f) Magnification of a microphotograph (e). Uniseriate xylem rays surrounding vessel element are clearly visible. (g) The ring-porous wood, with large vessels produced at the beginning of the season and narrow vessels formed in its second part, is shown in a transverse section of Quercus sp. Frequent uniseriate and characteristic multiseriate rays are presented. (h) Horizontal bands of xylem rays are visible on a radial section of Quercus sp. wood. (i, j) Heterogeneous, multiseriate rays of Cornus sp. are shown in radial (i) and tangential (j) sections of the secondary xylem. Procumbent ray cells are present in the center of xylem rays, upright ray cells are visible at the ray margins, while square ray cells are located between procumbent and upright cells. R xylem ray, Rp procumbent xylem ray, Rs square xylem ray, Ru upright xylem ray, T, tracheid/s, V vessel; dashed lines show the border of annual ring. Microphotographs were taken in white transmitted light; scale bars: 100 μm (a–e, g–j) and 20 μm (f) (g–j courtesy of Dr. E. Myśkow)

Contact ray cells in seed plants. Xylem rays neighboring tracheids (a, b) and vessels (c, d) are presented in radial sections of the secondary xylem of Pinus sp. (a), Picea sp. (b), and Salix sp. (c, d). Large pits of contact cells are clearly visible; they are indicated by arrowheads; the borders of some contact pits are marked by solid lines. In heterogeneous rays of Salix sp., contact ray cells and isolation ray cells are labeled. CC contact cells, IC isolation cells. Microphotographs were taken in white transmitted light, scale bars: 20 μm (a, b courtesy of Dr. E. Myśkow)
All features of ray anatomy, useful for taxonomists, are a direct consequence of the defined roles that particular xylem elements perform in the plant. In this light, the spacing and dimension of xylem rays result from the necessity to transport solutes in radial direction in woody plants. Rays are specialized highways for radial transportation within the xylem region and with neighboring tissues, especially towards to the cambium and phloem. Thus, their distribution has to be precisely regulated by the controlled spacing of ray initials in cambium. The latter is controlled by interactions between regulatory factors, presumably ethylene and auxin (Lev-Yadun and Aloni 1991, 1992, 1995). Formation of new ray initials and the enlargement of existing ones are induced by ethylene, while the presence of ray tracheids, vessels, and fibers within the rays results from radial auxin movement. Similarly the formation of specific morphological types of ray cells is the effect of hormonal interaction between axially transported auxin and radially spreading ethylene. If one of the hormones prevails, the cell shape changes in accordance with the direction of dominant hormone movement. If both of these hormones are balanced, the cells maintain a square shape (Lev-Yadun and Aloni 1991, 1995).
The size of the rays changes during their ontogeny and strongly depends on plant growth rate. Rays become bigger as they age reflecting developmental events occurring in the cambial ray initials (Barghoorn 1941a; Evert 1961; Cumbie 1967; Ghouse and Yunus 1973; Myśkow and Zagórska-Marek 2004, 2008, 2013). The number of xylem rays and their volume in the total mass of the xylem region differ among vascular plants, but usually ranges between 8 and 25 % (Ghouse and Yunus 1974; Šćukanec and Petrić 1977; Gregory 1977), although in some tropical trees, e.g., Dillenia indica, the volume of ray initials can reach up to 75 % of the total volume of the wood (Ghouse and Yunus 1974). Moreover, in fast-growing trees, where radial growth leads to intensive circumference expansion, the ray size and the number of cells in particular rays increase rapidly (Bannan 1965; Gregory 1977). There is also a positive relationship between leaf area and the volume of ray parenchyma cells in outer sapwood (Gartner et al. 2000).
Rays may also affect cambial activity. One exquisite and striking example of this is the impact of rays on the maintenance of the meristematic character of cambial fusiform initials. The fusiform initials that are in direct contact with ray initials, or are in their close vicinity, maintain a meristematic identity. Conversely, the lack of such contacts and their more distant localization may induce the conversion of a fusiform initial into a new ray initial, or the loss of its meristematic character, leading to cell differentiation (Bannan 1965). The nature of signals transported from ray to fusiform initials, securing their meristematic state, is unknown.
4.2.2 Living Cells of the Axial System
Long-living cells of nucleated fibers and axial parenchyma cells can occur in the axial system of sapwood derived from cambial fusiform initials (Metcalfe and Chalk 1983; Carquist 2010). These cells are surrounded by secondary cell walls with numerous pits and can have a transverse septate. Living fibers and axial parenchyma have similar functions in woody plants. They are involved in sugar exchange at the symplasm/apoplasm interface (Sauter 1982; Améglio et al. 2004) and in the process of starch accumulation (Sinnott 1918; Ziegler 1964; Yamada et al. 2011). They also participate in long-distance symplasmic transport, through the formation of vertically orientated pathways for the spread of solutes, being simultaneously in direct contact with oppositely orientated rays (Fig. 4.3; van der Schoot and van Bel 1989; Sokołowska and Zagórska-Marek 2012).

The axial system of xylem parenchyma in angiosperms. (a) Strands of axial terminal parenchyma cells are shown in the tangential section of the secondary xylem of Ulmus sp. The cells have tapered ends and maintain the shape of fusiform cambial initials, from which they derived. (b–f) Different types of apotracheal (b–d, f) and paratracheal (e) axial parenchymas are presented in transverse sections of Juglans sp. (b), Cornus sp. (c), Ficus sp. (d), Acer sp. (e), and Betula sp. (f) wood. Axial parenchyma cells are marked by arrowheads on microphotographs (b), (c), (e), and (f). (b–d) In the apotracheal type, narrow (b, c) or wide (d) strands of axial parenchyma cells are distributed perpendicular (b, d) or parallel (c) to the xylem rays. (e) In the paratracheal type, axial parenchyma cells are closely associated with vessels. (f) Terminal parenchyma, the characteristic type of apotracheal parenchyma, is formed at the end of growing season and it is localized alongside the border of annual ring. Ap axial parenchyma, R xylem ray, Tp terminal parenchyma, V vessel; dashed line shows the border of annual rings. Microphotographs were taken in white transmitted light, scale bars: 50 μm (a, b courtesy of Dr. E. Myśkow)
Specific distribution of the axial parenchyma cells among other xylem elements is important from the taxonomical point of view, while also giving additional information about the function of living cells in wood. Axial parenchyma cells rarely occur in gymnosperms, whereas in angiosperms they are more frequent and can form various patterns, e.g., in Alnus glutinosa or Crataegus sp., where parenchyma cells are sparsely localized, while in Ficus sp., they are abundant and form wide bands of living cells in the xylem region (Fig. 4.3; Metcalfe and Chalk 1983; Carquist 1988; IAWA 1989). Two major types of axial parenchyma—apotracheal and paratracheal—can be distinguished based on the type of cell distribution in regard to vessels (Fig. 4.3). In paratracheal parenchyma, the most common in angiosperms, the living cells surround tracheary elements and form an incomplete or complete sheath around the vessels. In the case of apotracheal types, the cells of axial parenchyma are distributed without relation to the vessels, being usually sparsely localized in the xylem region. When apotracheal parenchyma cells are more frequent, they can aggregate, forming smaller groups or bands among the other xylem elements. Moreover, axial parenchyma cells can be closely associated with growth rings, forming narrow bands at the beginning (initial parenchyma) and at the end (terminal parenchyma) of the growing season (Fig. 4.3). The latter type is clearly more frequent in woody plants (Braun and Wolkinger 1970; Metcalfe and Chalk 1983; Carquist 1988; IAWA 1989).
4.2.3 Xylem Parenchyma Cells: Classification Based on Cell Function
The cells of both axial and radial xylem parenchyma play different functions in sapwood. They can form storage tissue, where accumulation and remobilization of the storage substances take place, or they can serve as specialized cells which are in contact with conducting elements and are involved in exchange at the symplasm/apoplasm interface (Czaninski 1977; Barnett 2006).
The terminology of the latter, specialized xylem parenchyma cells, is diverse. Generally, these cells are described as “accessory tissue” that is formed by parenchymatous cells closely associated through numerous pits with the tracheids or vessels. Accessory tissue includes the contact cells of xylem rays and paratracheal parenchyma cells abutting the tracheids in gymnosperms and vessels in angiosperms (Braun 1984). According to another, more narrow classification, “contact cells” are defined as specialized groups of living axial and radial xylem parenchyma cells that are not only contiguous with tracheary conducting elements but are particularly involved in solute exchange between the symplasm and apoplasm in wood (Gregory 1978). In angiosperms, a narrow term of “vessel-associated cells,” which refers to the xylem parenchyma cells facing the vessels and physiologically linked with them, was proposed by Czaninski (1977). Alternatively, it has been suggested to maintain the term “contact cells” as a broader and more accurate definition, true for all seed plants (Gregory 1978). In recent literature both expressions are used frequently and alternatively, because the biological functions of contact cells in seed plants and vessel-associated cells in angiosperms are similar. In this chapter the term “contact cells” will be used and refer to all seed plants.
4.3 Cellular Organization of Xylem Parenchyma Cells
4.3.1 Metabolic Activity and Vitality
Xylem parenchyma is composed of long-living cells which are metabolically active during the whole year (Fig. 4.4). Judging by cell respiration, the metabolic rate is estimated to be slightly higher in xylem parenchyma cells than in storage tissues, but far below the meristematic cells. The respiration activity of xylem parenchyma is low and varies between tree species; however, it does not decline with cell age (Spicer and Holbrook 2007).

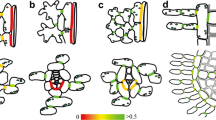
The metabolic activity and symplasmic coupling of xylem parenchyma cells in the sapwood of Acer pseudoplatanus. (a) The metabolic activity of xylem parenchyma cells was confirmed by staining with thiazolyl blue. The dark color of formazan, the product of mitochondrial dehydrogenases activity in thiazolyl blue reaction (marked with arrowheads), is visible in the xylem rays and in the cytoplasm of paratracheal and terminal axial parenchyma cells. (b–d) Symplasmic connectivity between living xylem parenchyma cells visualized by carboxyfluorescein (CF). After loading of carboxyfluorescein diacetate solution into the maple branches through the vascular system and its conversion to fluorescent tracer—CF in the living cells—the dye was transported symplasmically within the spatial system of living cells in the secondary xylem. The strong fluorescent signal of CF in the cytoplasm of xylem rays and paratracheal and terminal axial parenchyma makes visible the routes of symplasmic transport in sapwood. C cambium, Pp paratracheal parenchyma, R xylem ray, Tp terminal parenchyma, V vessel; arrowheads show metabolically active axial parenchyma cells. Microphotographs were taken in white transmitted light (a) and in blue (475 nm) excitation light (b–d) in the epi-fluorescent microscope, scale bars: 50 μm
The vitality of parenchyma cells subsequently lowers with the increase in distance from the cambial region and can be exemplified by decreasing starch content and the number of organelles in the ray cells towards the center of the trunk (Höll 1975; Song et al. 2011), although cell vitality is not a simple function of the distance from the cambial region. Among the population of living xylem parenchyma cells, some of them can die earlier, while others later. This discrepancy can be induced by the process of early cell death that occurs in sapwood when cells have contact pits with short-living conducting elements of the xylem, e.g., ray tracheids in Pinus and Abies (Nakaba et al. 2006, 2008) or vessels in Populus (Nakaba et al. 2012a). The occurrence of early cell death is species dependent. No evidence of natural early cell death, either in ray or in axial parenchyma, was found in the sapwood of Acer rubrum, Fraxinus americana, or Quercus rubra, where all analyzed parenchyma cells were alive (Spicer and Holbrook 2007). Nevertheless, during the process of heartwood formation, all xylem parenchyma cells ultimately die (see Sect. 4.4.3.4). The timing of cell death during heartwood formation varies between different types of xylem parenchyma. It begins earlier in axial parenchyma, compared to the rays, and depends on vicinity to the pith region as was shown in branches of Robinia pseudoacacia (Nakaba et al. 2012b).
4.3.2 Arrangement of the Cytoskeleton Network
Long-living xylem parenchyma cells exhibit a characteristic organization of the cytoskeleton. Microtubules and microfilaments are bundled and show a generally net axial orientation (Chaffey and Barlow 2001, 2002). They spread from one end of the cell to the other and are parallel to the long axis of prosenchymatous xylem parenchyma cells. Thus, according to cell shape and in respect to the main axis of the stem, cytoskeleton components are arranged radially in ray cells and vertically in axial parenchyma, although a meshwork arrangement of microfilaments or helical arrays of microtubules can also be found (Chaffey et al. 1997, 1999, 2000; Chaffey and Barlow 2001, 2002). The mostly axial distribution of the cytoskeleton is disturbed in the pit periphery, where concentric rings of microfilaments and microtubules are present (Chaffey and Barlow 2002; see Sect. 4.3.4). The directional orientation of microtubules and microfilaments may facilitate intracellular transport and assist in the intercellular movement. Additionally, the cytoskeleton network in radial and axial xylem parenchyma cells forms a continuity with similar cytoskeleton elements of cambial and phloem regions, creating a three-dimensional system supporting symplasmic transport in plants (Chaffey and Barlow 2001, 2002).
4.3.3 Characteristics of the Cell Wall
Xylem parenchyma cells are usually surrounded by a thick primary cell wall and a thick, frequently multilayered, secondary cell wall. The latter is generally rich in lignins and pectins (Chafe and Chauret 1974). The lignification process starts earlier in the contact cells than in the isolation cells, with the exception of pit membranes, which are composed of a primary cell wall and are clearly unlignified for a very long time (Chafe and Chauret 1974; Chafe 1974; Murakami et al. 1999; Nakaba et al. 2012a). Formation of the secondary cell wall and deposition of its component, xylan, start much later in ray cells than in fibers and vessels (Kim et al. 2012). Additionally, in some conifers, considerably delayed lignification or the lack of a secondary cell wall is characteristic of ray parenchyma cells that are not in direct contact with ray tracheids. These ray parenchyma cells can maintain the primary nature of the cell wall for more than 30 years, as in Pinus densiflora, while in Pinus rigida a secondary cell wall does not form in these cells at all (Nakaba et al. 2008).
Parenchyma cells neighboring to the tracheary elements have an additional layer of a loose-textured, usually unlignified material that is generally called the “protective layer” (Chafe 1974; Mueller and Beckman 1984; Wiśniewski et al. 1987; Barnett et al. 1993). The protective layer extends between plasmalemma and the entire inner surface of the secondary cell wall of contact cells. It is usually thicker around the pits (Chafe 1974; van Bel and van der Schoot 1988; Barnett et al. 1993). The real role of the protective layer is still a matter of debate. This layer was believed to protect living paratracheal cells against high osmotic pressure and the oscillations present in the conducting elements of the xylem (van Bel and van der Schoot 1988). However, it is not clear why some cells, e.g., ray cells, without this layer can survive in contact with dead xylem elements (Chafe 1974; Chafe and Chauret 1974) or how a layer formed mainly by pectins and cellulose can withstand hydrostatic fluctuations in conducting tracheary elements (Barnett et al. 1993). Nor is the protective layer necessary for formation of tyloses—outgrowths of parenchyma cells into tracheary elements—as it can occur in non-tylose-forming species and be absent in tylose-forming ones (Chafe and Chauret 1974; Barnett et al. 1993). Presently it is suggested that the protective layer is not involved in plant defense but rather in intercellular exchange at the symplasm/apoplasm interface (Barnett et al. 1993). Due to the low permeability of secondary cell walls, demonstrated in experiments with lanthanum (Wiśniewski et al. 1987), the protective layer can increase continuity between the protoplast and the lumen of tracheary elements (Barnett et al. 1993). Its presence increases the surface of the symplasm/apoplasm interface, which is not limited to the pit regions exclusively but expands to the entire plasmalemma surface of the contact cells (Barnett et al. 1993). Thus, the protective layer considerably enhances the efficiency of intercellular exchange in woody plants.
4.3.4 Pitting and Plasmodesmata Distribution
The striking differences in the permeability of pit membranes compared to secondary cell walls suggest that the longitudinal and lateral exchange of solutes in the xylem region occurs via the pit system (Wiśniewski et al. 1987). Pits between xylem parenchyma cells are usually simple, while half-bordered at contact with tracheary elements (Yang 1978). The type of pit—simple or bordered—strongly depends on the thickness of xylem parenchyma walls (Yang 1978; Carquist 2007). Simple pits are localized in relatively thin walls. As the thickness of the secondary cell wall increases, selective pressure rises to form bordered pits. Thus, bordered pits among living parenchyma cells, especially in ray cells, are quite common (Carquist 2007).
The distribution and density of the pits provide important information about the possible direction and intensity of solute flow between the cells. Pits are usually unevenly distributed, e.g., in tangential walls which are more densely pitted compared to radial walls in procumbent ray cells, while in square and upright ray cells, the pits are more or less equally distributed through all cell walls (Carquist 2007). The function of contact cells in the transfer between symplasm and apoplasm is underscored by the presence of extremely large pits between tracheary elements and contact cells (Fig. 4.2; Sauter et al. 1973; Sauter and Kloth 1986; Murakami et al. 1999).
Pits occurring between living parenchyma cells have a characteristic ultrastructure, with concentric rings of both microtubules and microfilaments, as can be seen by the immunolocalization of myosin, tubulin, and actin around a pit area (Chaffey and Barlow 2002). During pit formation, microfilaments probably prevent the deposition of the secondary cell wall at the pit membrane by exclusion of microtubules from the pit periphery (Chaffey and Barlow 2002). Thus, the pit membranes of xylem parenchyma cells are thin and possess the primary structure of a wall, with numerous simple or branched plasmodesmata (Yang 1978; Sauter and Kloth 1986; Wiśniewski et al. 1987; Chaffey and Barlow 2001).
Besides living cells, plasmodesmata are also frequently present in the pseudo-tori, which are thickenings of the primary cell wall located eccentrically in the developing pit membranes of fibers, fiber tracheids, and vessels of some angiosperm families such as Rosaceae and Ericaceae (Barnett 1982, 1987a, b; Lachaud and Maurousset 1996; Rabaey et al. 2006, 2008). Plasmodesmata in pseudo-tori are densely packed, branched, and secondary in origin (Barnett 1982, 1987a, b; Rabaey et al. 2006; Jansen et al. 2007). Just before accomplishing cell differentiation and protoplast autolysis, a secondary cap-like layer is deposited on the primary thickenings of the pseudo-tori (Barnett 1987a; Lachaud and Maurousset 1996; Jansen et al. 2007; Rabaey et al. 2006, 2008). This additional layer can seal intercellular connections, but plasmodesmata can sometimes protrude through its outer part (Lachaud and Maurousset 1996; Rabaey et al. 2008). It is unknown whether or not the plasmodesmata in pseudo-tori are functional. The irregular and amorphous shape of pseudo-tori, as well as an eccentric position that does not match the pit aperture, suggests that pseudo-tori cannot properly seal the pits and thus they are not involved in pit aspiration (Rabaey et al. 2006, 2008). Therefore, it is possible that the pseudo-tori are formed to maintain intercellular communication between various xylem elements (Barnett 1982, 1987b; Lachaud and Maurousset 1996; see Sect. 4.4.3.5).
4.4 Functions of Xylem Parenchyma Cells in the Secondary Xylem
The long-living xylem parenchyma, which is composed of both axial and radial systems, forms a unique, functional continuum for efficient intercellular communication (Fig. 4.5). Thus, the xylem parenchyma is a vital component of the secondary xylem. The whole spatial network of interconnected and integrated cells is an example of a three-dimensional symplasmic field where free symplasmic short- and long-distance transport is continued (Figs. 4.4 and 4.5). Apart from the main function of storing and distributing reserve substances, living xylem parenchyma cells are involved in exchange at the symplasm/apoplasm interface, embolism repairs, defense mechanisms against vascular pathogens, and vascular structure (Chowdhury 1953; Sauter 1982; Benhamou 1995; Hilaire et al. 2001; Améglio et al. 2004; Salleo et al. 2004). They also play an important role in the biomechanics of living deciduous trees and increase radial strength and the transverse elastic anisotropy of trunks (Burgert et al. 1999, 2001; Burgert and Eckstein 2001).

Schematic diagram of living parenchyma cells in the secondary xylem of angiosperms. Xylem rays and different types of axial apotracheal and paratracheal parenchymas form the spatial system of interconnected living cells in sapwood. To simplify the diagram, only vessels are indicated, although other types of nonliving elements (tracheids, fibers, fiber tracheids) are also present in the secondary xylem. Ap apotracheal parenchyma, Pp paratracheal parenchyma, R xylem ray, Rp procumbent xylem ray, Ru upright xylem ray, Tp terminal parenchyma, V vessel
Wood without living cells cannot exist. Even in a species devoid of rays, at least in juvenile wood, living cells of axial parenchyma and nucleated fibers are present (Barghoorn 1941b; Lev-Yadun and Aloni 1991; Carquist 2009, 2010). Over the course of evolution, the volume of living parenchyma cells in the secondary xylem and their specialization have increased. Initially, the diffusive parenchyma cells supported the conductive system by maintaining the appropriate water column in vessels. Then, the tendency to group living axial elements around the vessels, together with the intensive secondary thickening of trunks, forced the integration of axial parenchyma cells with developing rays. At the end, due to the widening of the rays and the predominance of procumbent cells, the rays became a route for photoassimilate transport across the stem (Carquist 2009).
4.4.1 Storage Parenchyma
Storage parenchyma is specialized for seasonal accumulation and mobilization of organic and inorganic materials. Besides starch and lipids (the most frequent storage materials in plants) parenchyma cells can store nitrogen compounds in the form of protein bodies as well as crystals and silica bodies (Sinnott 1918; Ziegler 1964; Höll 1975; Carquist 1988; Harms and Sauter 1992a). Starch grains are accumulated mostly in the apotracheal parenchyma and in isolation xylem ray cells (Gregory 1978), as well as in the living fibers of the outermost ring (Yamada et al. 2011). The amount of accumulated starch grains varies among different types of xylem parenchyma cells and depends on the distance from the cambium, where the largest quantity of starch is deposited in the current xylem year (Nakaba et al. 2012a). Parenchyma cells have also a protein-storing function. In the small vacuoles of temperate-zone trees, polypeptides of 32–36 kDa are accumulated and form specific protein bodies. They appear near the cambium, mainly in the rays and in axial paratracheal parenchyma (Harms and Sauter 1992a, b; Hao and Wu 1993).
The quantity of accumulated storage compounds in xylem parenchyma changes seasonally. It corresponds to plant activity and to alternating periods of nutrient abundance and deficiency. Reserves are usually accumulated at the end of a growing season and then utilized in spring time, when photoassimilates are necessary but are not yet produced (Harms and Sauter 1992a; Sauter and van Cleve 1994; Sauter 2000). For example, the starch content in poplar and willow trees is lowest after budbreak and leaf expansion. Then, the starch amount starts increasing and reaches maximum value in autumn, when the leaves are shed. After that, starch content gradually decreases (Essiamah and Eschrich 1985; Sauter et al. 1996). The lowered level of starch in winter correlates with increased amounts of sugars (Sauter and van Cleve 1994) which are involved in the adaptation of the cells to frost and in the refilling of embolized tracheary elements (Essiamah and Eschrich 1985; Sauter et al. 1996). Starch is resynthesized again from available sugars before spring. Its content rises temporarily, until the budbreak period, when reserves are utilized during plant growth (Sauter 1972; Essiamah and Eschrich 1985; Harms and Sauter 1992b; Sauter and van Cleve 1994; Sauter et al. 1996; Sauter 2000). Similarly, the amounts of other stored compounds can change seasonally. Lipids are intensively accumulated in summer and remain at a prominent level during the entire winter until spring remobilization (Sauter and van Cleve 1994). However, protein bodies are formed mostly in fall, in the time of leaf yellowing, and are almost completely degraded during budbreak (Sauter and van Cleve 1990; Harms and Sauter 1992a). Nitrogen released from protein bodies is used during the onset of new shoot growth (Langheinrich and Tischner 1991).
Due to the seasonally regulated quantity of storage compounds, the ultrastructure of storage parenchyma cells changes dramatically during the year. Before budbreak the protoplasts of storage parenchyma contain numerous amyloplasts, oleosomes, and protein bodies (Harms and Sauter 1992a, b; Sauter and van Cleve 1994; Sauter et al. 1996). After the onset of new growth in spring, the number of starch grains is significantly reduced, oleosomes almost disappear, while protein bodies can no longer be detected (Harms and Sauter 1992a).
4.4.2 Long-Distance Transport Routes in Woody Stems
The seasonally alternating accumulation and mobilization of storage compounds in xylem and phloem parenchyma cells requires precisely controlled translocation of reserves to the sink regions. This has been clearly demonstrated in experiments with dormant cambium, in which local heating of the stem induced division activity in the cambial cells and starch remobilization from the phloem towards the cambium (Oribe et al. 2001, 2003; Begum et al. 2007). In sapwood, the manner of photoassimilate and other solute flow in the living xylem cells is still a matter of debate (van Bel 1990; Sauter 2000). Although the long-distance transport of water and solute minerals in the apoplasmic system via dead xylem elements like tracheids and vessels is unquestionable (Holbrook and Zwieniecki 2005), the structure of long-living axial and radial parenchyma cells (Fig. 4.5) suggests that communication and trafficking via the symplasmic route is likewise important.
The shape of the xylem parenchyma cells and pit distribution localized in their cell walls (see Sect. 4.3.4) reflect the major directions of solute flow. Axial parenchyma cells are involved in vertical transport, procumbent ray cells in radial spreading, while square to upright ray cells represent an intermediate step (Carquist 2007). Among the specific ray cells abutting the vessels, isolation cells are involved in radial transport and function also as a storage region (Sinnott 1918; Sauter 1972; Sauter and Kloth 1986; Murakami et al. 1999). Due to larger pit-field areas and a higher frequency of plasmodesmata at the tangential walls, isolation cells are better specialized for radial symplasmic transport than contact cells (Sauter and Kloth 1986; Sauter 2000). However, intercellular spaces between isolation cells (Murakami et al. 1999) may also suggest the involvement of the apoplasmic transport route. Delayed differentiation of the secondary cell wall in isolation cells, as compared to contact cells, suggests that the unlignified tangential walls can temporarily assure apoplasmic continuity between phloem and xylem (Murakami et al. 1999). However, radial trafficking in apoplasm, as demonstrated using specific tracers like acid fuchsin and safranin, seems to be limited. These apoplasmic dyes loaded to poplar plants through the vascular system were only visible in the close vicinity of vessel elements and did not move further in the cell wall system of ray parenchyma cells (Sano et al. 2005). Additionally, the simultaneous application of apoplasmic (sulforhodamine B, SRB) and symplasmic (carboxyfluorescein, CF) tracers to maple tree branches revealed a difference in efficiency between both routes of transport. Radial apoplasmic transfer of SRB was restricted to the axial xylem elements, especially vessels, while at the same time CF was visible in the symplasm of xylem rays and in the cambial region (Sokołowska and Zagórska-Marek 2012), suggesting a prevailing role of symplasmic route in radial transfer in wood.
4.4.2.1 Radial Symplasmic Movement
Xylem and phloem rays function as living bridges that connect the cambial region and living elements of secondary xylem and phloem. Therefore, they are involved in centripetal and centrifugal radial symplasmic transport across the stem (Langenfeld-Heyser 1987; van Bel 1990; Sokołowska and Zagórska-Marek 2007, 2012). This route of transport is facilitated by abundant plasmodesmata present in the tangential walls of the rays (Barnett 1982; Sauter and Kloth 1986) and was demonstrated in experiments with tracer loading (Fig. 4.4) and membrane potential measurement (van der Schoot and van Bel 1990; Sokołowska and Zagórska-Marek 2007, 2012; Fuchs et al. 2010). Moreover, radial translocation of 14C-labeled assimilates (Langenfeld-Heyser 1987; Korolev et al. 2000b) and high translocation rates of sugars through the tangential area of ray cells (Sauter and Kloth 1986; Sauter 2000) strongly support the relevance of the radial symplasmic pathway across the stem and towards the cambium. Furthermore, localization of storage proteins remobilized during the first divisions of cambial cells in Populus directly confirmed the significance of the radial symplasmic route. Reserves can indeed be transported symplasmically between phloem parenchyma cells, ray cells, and fusiform cambial cells, because proteinaceous particles were clearly localized in the cytoplasmic sleeve of plasmodesmata (Fuchs et al. 2010).
4.4.2.2 Intercellular Spaces: Apoplasmic Route for Gas Exchange
A complex system of intercellular spaces spans along the axial and radial groups of parenchyma cells, from cambium towards the mature xylem, and serves as an additional apoplasmic route of transport in secondary xylem (van der Schoot and van Bel 1989; Sun et al. 2004; Nagai and Utsumi 2012). This system is comprised of short vertical and prominent radial canacules that probably originate from fusion of schizogenous spaces (van der Schoot and van Bel 1989).
Intercellular spaces are separated from living ray cells by blind pits and seem to be filled with water (van der Schoot and van Bel 1989) or with gases (Nagai and Utsumi 2012). Thus, they are considered to be involved in water (van der Schoot and van Bel 1989; van Bel 1990; Sun et al. 2004; Kitin et al. 2009) or gas (Hook and Brown 1972; Spicer and Holbrook 2005; Nagai and Utsumi 2012) trafficking, presumably in a radial direction. However, the transfer of water and solutes via intercellular spaces has recently been questioned (Nagai and Utsumi 2012). Firstly, intercellular spaces are prominent in tree species living in wet habitats and having a higher tolerance to flooding, e.g., in Nyssa aquatica and Fraxinus pennsylvanica, where their presence probably increases plant aeration (Hook and Brown 1972; Spicer and Holbrook 2005). Secondly, in conifers, intercellular spaces stay empty even if they surround living rays and tracheids filled with cytoplasm or water (Nagai and Utsumi 2012). Thus, it seems plausible that intercellular spaces function rather as a gas exchange system in apoplasm (Nagai and Utsumi 2012).
4.4.2.3 The Role of Cambial Cells in Radial Xylem–Phloem Exchange
Cambium serves as a strong sink that draws nutrients due to a high level of division activity and production of derivatives (Krabel 2000; Korolev et al. 2000b). The radial transport of 14C-assimilates (Langenfeld-Heyser 1987; Tomos et al. 2000; Korolev et al. 2000a, b) and fluorescent tracers (van der Schoot and van Bel 1990; Sokołowska and Zagórska-Marek 2007) from the phloem and xylem regions to the cambium occurs symplasmically and seems to be partially ceased at the cambial zone. The symplasmic route of transport towards the cambium corresponds well with the general principle that unloading to the meristematic tissues or growing organs is symplasmic (Patrick and Offler 1996; Tarpley and Vietor 2007). Moreover, it seems that cambial meristem collects assimilates and metabolizes them quickly to obtain energy for functioning and division activity (Krabel 2000; Korolev et al. 2000a), because cambial cells reveal very low concentrations of K+ and sugars (sucrose, glucose, fructose) compared with neighboring cells of the phloem and xylem regions (Korolev et al. 2000a; Tomos et al. 2000). Accordingly, it becomes obvious why dormant cambial cells, experimentally activated by local heating, cannot divide further in the event of deficiency of reserve materials (Oribe et al. 2001, 2003).
4.4.2.4 Symplasmic Transport: Preferential Route of Transport via Parenchyma Cells in Sapwood
To summarize, the anatomical and physiological features of living xylem parenchyma cells, such as plasmodesmata distribution and frequency (Sauter and Kloth 1986; Barnett 2006), cytoskeleton organization (Chaffey and Barlow 2001, 2002), storage substances mobilization (Fuchs et al. 2010), and movement of fluorescent tracers (Fig. 4.4; Sokołowska and Zagórska-Marek 2012), evidence the role of symplasmic trafficking via parenchyma cells in xylem and towards the cambium. Although, the final choice of the route in xylem parenchyma—symplasmic or apoplasmic—depends mostly on the type of substance transported (Sharkey and Pate 1975; Vogelmann et al. 1985). However, bearing in mind the restricted radial trafficking of apoplasmic tracers and the involvement of intercellular spaces in gas exchange, it can be considered that intercellular communication via living xylem cells in sapwood occurs prominently via the symplasmic route (Fig. 4.5).
4.4.3 Contact Cells: Specialized Types of Axial and Radial Parenchyma Cells
Contact cells, described in angiosperms as vessel-associated cells, are usually small, living parenchyma cells abutting tracheary elements. They have a dense cytoplasm with a voluminous nucleus, small vacuoles, and numerous ribosomes and mitochondria (Czaninski 1977; Alves et al. 2001, 2007). Starch is usually absent, even during the winter season (Sauter et al. 1973; Czaninski 1977; Braun 1984). In trees of temperate zones, starch appears in contact cells only temporarily—in early spring—and is quickly remobilized just before and during bud swelling, considerably earlier than in the neighboring cells of typical storage parenchyma (Sauter 1972; Sauter et al. 1973; Braun 1984). Contact cells are involved in varied physiological processes due to their bidirectional transmembrane transport ability at the symplasm/apoplasm interface. Sugar influx and efflux (Sauter 1982), exudation of occluding and toxic substances during vascular pathogen infection (Shi et al. 1992; Benhamou 1995; Cooper et al. 1996) and of phenolic compounds during heartwood formation (Magel et al. 1991; Spicer 2005), require a controlled exchange of substances between contact cells and tracheary elements, via active and passive routes.
Despite the contact cells being directly connected with tracheary elements by large and densely packed contact pits (Fig. 4.2; Sauter 1972; Sauter et al. 1973; Murakami et al. 1999), they are devoid of plasmodesmata in contact pit membranes (Czaninski 1977; Decourteix et al. 2006), and thus they are symplasmically isolated from vessels and tracheids. Therefore, communication and trafficking between the apoplasm, especially the xylem sap of tracheary elements and contact cells as a part of three-dimensional continuum of living xylem parenchyma cells, have to proceed through the plasma membrane. Probably, the interface of symplasm and apoplasm is enhanced by a protective layer in contact cells (see Sect. 4.3.3), which maintains apoplasmic continuity with vessels.
The intensive transmembrane transport at the symplasm/apoplasm interface requires the increased metabolic activity of contact cells (Fig. 4.4). And indeed, contact cells exhibit enhanced levels of acid phosphatases, peroxidase, and dehydrogenases, in comparison to storage parenchyma cells (Sauter 1972; Sauter et al. 1973; Braun 1984; Alves et al. 2001). The enzymatic activities of contact cells change seasonally in trees of the temperate zones. They increase in late fall and winter to maximum in spring time, before and during the budbreak period, then cease rapidly in late spring when leaves are flushing, to finally become undetectable throughout the entire summer (Sauter 1972; Sauter et al. 1973). The high metabolic activity of contact cells is closely related to the low level of starch in these cells, and starch resynthesis coincides with a rapid drop of acid phosphatase activity (Sauter 1972; Sauter et al. 1973). Thus, it seems reasonable that in continuously growing tropical trees, contact cells are free of starch and show permanent acid phosphatase activity (Braun 1984). Regarding the high metabolic activity of contact cells with simultaneous lack of starch, it can be suggested that these cells are an important site of respiratory activity and are involved in intensive transport between apoplasm and symplasm (Sauter 1972; Gregory 1978; Murakami et al. 1999; Alves et al. 2001).
4.4.3.1 Sugar Influx/Efflux Interactions at the Symplasm/Apoplasm Interface
Sucrose content in the tracheary sap of temperate zone trees changes throughout the year, even in winter time when the growth activity of plants in restricted (Sauter 1980, 1982, 1983, 1988; Sauter et al. 1996; Améglio et al. 2001, 2004). Sucrose can be released from starch; thus, the amount of sucrose is inverse to starch content and changes seasonally. Accumulation of sucrose in tracheal sap usually starts at the beginning of winter, reaches its maximum in midwinter, keeps high values before and during budbreak period, and drops sharply when leaves are flushing (Sauter 1982, 1983, 1988; Améglio et al. 2002). The high osmolarity of tracheal sap coincides with drop of starch concentration in storage parenchyma and the high metabolic activity of contact cells, indicating the involvement of the contact cells in sucrose efflux into tracheary elements (Sauter 1972, 1980, 1981, 1982, 1983, 1988; Sauter et al. 1973; Améglio et al. 2004). In winter, sugar efflux is a slow continuous and selective process that depends on many factors such as temperature, respiration, sucrose concentration in xylem parenchyma cells, the intensity of starch hydrolysis and resynthesis, as well as the rate of sugar absorption into the symplasm (Sauter et al. 1973; Sauter 1981, 1982, 1988; Améglio et al. 2001, 2004). Efflux cannot operate as a simple leaking into the apoplasm, but probably occurs by facilitated diffusion mediated by a yet unknown protein, and regulated by metabolic inhibitors (Sauter 1980, 1982, 1988; Améglio et al. 2004). Alternatively, sugar efflux has recently been proposed as an energy-consuming process that is mediated by proton co-transporters (Salleo et al. 2004; Secchi and Zwieniecki 2012). Apart from sugar efflux, contact cells are also involved in an opposite process of active sugar influx from tracheary sap to symplasm (Sauter 1983; Decourteix et al. 2006, 2008; Alves et al. 2007; Bonhomme et al. 2009). Influx is carried out by putative xylem sucrose and hexose transporters (Decourteix et al. 2006, 2008) which depend on the driving force generated by H+-ATPase localized in the plasma membrane of contact cells (Fromard et al. 1995; Arend et al. 2002, 2004; Decourteix et al. 2006; Alves et al. 2007). Which of these two competing biochemical processes dominates, sugar efflux or influx, depends mostly on temperature, and thus it is periodically regulated (Sauter 1982, 1983; Améglio et al. 2004; Decourteix et al. 2006; Alves et al. 2007; Bonhomme et al. 2009). Efflux dominates in midwinter, when temperatures are low (below 5 °C). In that period sugars are converted from starch in the xylem parenchyma cells and then accumulated in tracheary elements (Sauter 1980, 1982, 1983; Sauter et al. 1996; Alves et al. 2007; Salleo et al. 2009). The activity of H+-ATPase is highly reduced at low temperatures (Alves et al. 2001, 2007; Améglio et al. 2004), resulting in the cessation of sugar influx and, in turn, in unlimited sugar efflux to apoplasm. When temperatures are higher (over 15 °C), starch begins to resynthesize. The sucrose concentration gradient lowers at the symplasm/apoplasm interface and decreases the efflux rate (Sauter 1983, 1988; Sauter et al. 1996). Additionally, H+-ATPase is highly active and produces the proton motive force used by transporters for active sugar uptake. Thus, at higher temperatures in winter, sugar influx dominates (Sauter 1982, 1983, 1988; Alves et al. 2001, 2007; Améglio et al. 2004; Bonhomme et al. 2009).
4.4.3.2 The Biological Role of Active Substance Secretion and Uptake by Contact Cells
Contact cells are involved in specialized translocation of water and other materials between symplasm and apoplasm (Gregory 1978; Murakami et al. 1999; Salleo et al. 2004; Decourteix et al. 2006; Secchi and Zwieniecki 2011), although the detailed mechanism of sugar exchange between contact cells and tracheary elements seems to be more complex (Alves et al. 2007; Nardini et al. 2011; Secchi and Zwieniecki 2011, 2012). Sucrose accumulation in the tracheary sap of Acer saccharum (sugar maple) from November to April, which coincides with the high metabolic activity of contact cells in the vicinity of functional, water-conducting elements and a low level of starch content, together suggest that contact cells are able to release sugars and generate high osmotic pressure in vessels (Sauter 1972; Sauter et al. 1973; Braun 1984). Thus, they can facilitate water transport in xylem elements and ascend the sap before leaf flushing in early spring (Braun 1984). This can be particularly important in tropical trees during conditions of lowered transpiration due to high humidity during overcast days (Braun 1984). A dramatic increase in water uptake in early spring was recorded in some trees like Acer pseudoplatanus, Betula pendula, Alnus glutinosa, or Fagus sylvatica, although the origin of the spring sap was not determined (Essiamah and Eschrich 1986). In at least a few tree species, e.g., Juglans regia (walnut), the increased positive xylem pressure in spring and autumn is mainly generated by root pressure (Ewers et al. 2001). Thus, the mechanism responsible for the rising of sap in spring time varies between species and requires further investigation.
Xylem elements are an efficient translocation route for organic compounds to the meristematic tissues in species which flower before leaf appearance, in late winter or early spring, when the phloem strands are still nonfunctional (Sauter 1980, 1983). For instance, sugar influx rate and the activity of H+-ATPase in spring time are considerably higher in apical parts of 1-year-old branches, bearing bursting buds in walnut, compared to basal stem regions with non-bursting buds (Alves et al. 2007). Over 90 % of the total sucrose pool is provided for the opening of apical buds by xylem elements. This means that the xylem system is practically the only source of sucrose in the apical parts of walnut during budbreak (Bonhomme et al. 2009). This emphasizes the importance of contact cells in sugar uptake from apoplasm during growth resumption (Alves et al. 2007). Further translocation of sugars away from contact cells probably occurs via plasmodesmata, due to symplasmic connectivity between contact cells and the neighboring storage parenchyma (see Sect. 4.4.2). Interestingly, meristematic regions and vascular strands are symplasmically coupled (Duckett et al. 1994; Gisel et al. 1999; Banasiak 2011). Therefore, continued transport via living cells and the elements of the primary vascular system towards the apices serve as an interesting hypothesis that requires further examination. The most surprising fact related to sugar influx in walnut stems is the significantly low active uptake, reaching only 50 % of global sugar uptake in the basal parts of branches, where most of buds do not break (Bonhomme et al. 2009). Thus, different pathways are necessary in basal stem regions for sugar uptake from apoplasm. Although facilitated diffusion is possible (Bonhomme et al. 2009), an alternative route via endocytosis seems to be more convincing. It has previously been suggested that uptake of membrane-impermeant fluorescent tracers—Lucifer Yellow and Texas Red-labeled dextran (10 kDa)—by xylem parenchyma cells abutting the vessel in rice leaves occurs via endocytosis (Botha et al. 2008). Thus, contact cells are crucial elements in unloading in the xylem region and serve as an important, regulatory step in solute exchange at the symplasm/apoplasm interface.
4.4.3.3 The Involvement of Xylem Parenchyma Cells in Embolism Repairs
Contact cells are also an important factor in a mechanism leading to embolism repairs enabling proper long-distance apoplasmic transport. The role of contact cells in the refilling of cavitated vessels was intensively studied in many species like Juglans regia (Améglio et al. 2001, 2002, 2004; Sakr et al. 2003; Decourteix et al. 2006, 2008), Populus sp. (Secchi et al. 2011; Secchi and Zwieniecki 2012), Laurus nobilis (Salleo et al. 2004, 2009), or in two savanna trees—Schefflera macrocarpa and Caryocar brasiliense (Bucci et al. 2003).
Although the embolism repair mechanism has not been fully elaborated (Nardini et al. 2011; Secchi and Zwieniecki 2011), the involvement of xylem parenchyma, especially contact cells, in this process is unquestionable (Améglio et al. 2001, 2002; Bucci et al. 2003; Améglio et al. 2004; Salleo et al. 2009; Secchi et al. 2011; Secchi and Zwieniecki 2011, 2012). Firstly, xylem parenchyma cells are a source of sugars and energy. Sucrose, released from stored starch, can directly increase sap osmolarity and xylem pressure (Améglio et al. 2001, 2004) due to its translocation to embolized elements via contact cells (Améglio et al. 2004; Salleo et al. 2009; Secchi and Zwieniecki 2012), although it is unknown as to whether sugar efflux into cavitated conduits is facilitated through diffusion (Améglio et al. 2004) or is an active process (Salleo et al. 2004; Zwieniecki and Holbrook 2009; Secchi and Zwieniecki 2012). Sucrose also acts as a source of energy and indirectly increases sap osmolarity, through ion efflux into apoplasm, via respiration (Secchi et al. 2011; Secchi and Zwieniecki 2012). Regardless of the sucrose translocation pathway and the mechanism of osmotic gradient generation, defoliation in the vegetative season (Améglio et al. 2001) and phloem girdling (Salleo et al. 2004) both lead to reduction of the starch content in xylem parenchyma cells and strongly limit winter embolism repair ability and plant survival (Améglio et al. 2001).
When high sugar concentration is obtained in cavitated elements, water flow from neighboring cells according to the concentration gradient leads to dilution of the gases and complete conduit refilling (Zwieniecki and Holbrook 2009). Because xylem parenchyma cells are in close vicinity to gas-filled tracheary elements, they are regarded as one of the natural sources of water necessary for embolism repair (Améglio et al. 2001, 2002; Sakr et al. 2003; Bucci et al. 2003). Living and metabolically active contact cells are also considered as embolism sensor that detects cavitation and triggers the processes leading to conduit refilling. What is the nature of this signal emitted during cavitation? Possibly it can be connected with either wall vibration after mechanical stimulus (Salleo et al. 2008) and/or changes in osmotic concentrations between parenchyma cells and a neighboring tracheary element after water transport disruption (Zwieniecki and Holbrook 2009; Secchi and Zwieniecki 2011, 2012). It is quite plausible that xylem parenchyma cells, and especially contact cells, are also involved in other steps of the embolism repair mechanism.
4.4.3.4 Defense Mechanisms During Vascular Parasite Infection: Heartwood Formation
Xylem parenchyma cells abutting the vessels also play an important role in plant defense mechanisms against fungal (El Mahjoub et al. 1984; Street et al. 1986; Shi et al. 1992; Benhamou 1995; Cooper et al. 1996; Cooper and Williams 2004) and bacterial (Goodman and White 1981; Hilaire et al. 2001) infections spreading via tracheary elements. Contact cells neighboring infected vessels detect pathogens and trigger cellular defense responses. One of the first visible reactions, also found after injury (Schmitt and Liese 1990; Sun et al. 2008), is the formation of tyloses leading to partial or complete vessel occlusion (El Mahjoub et al. 1984; Clérivet et al. 2000). Besides tyloses, xylem parenchyma cells can also produce and secrete amorphous substances like gels and gums into the vessel lumen (Street et al. 1986; Shi et al. 1992; Benhamou 1995; Clérivet et al. 2000), which physically restrict pathogen movement within the vascular system (Shi et al. 1992). In addition, contact cells participate in chemical defense mechanisms. Living parenchyma cells abutting infected conductive elements have increased cytoplasmic activity (El Mahjoub et al. 1984; Shi et al. 1992). They produce, accumulate, and secrete into the vessels several materials like lipoidal, phenolic, or terpenoid substances (Shi et al. 1992; Benhamou 1995; Cooper et al. 1996; Clérivet et al. 2000), polysaccharides (Clérivet et al. 2000), elemental sulfur (Cooper et al. 1996; Williams et al. 2002; Cooper and Williams 2004), or cationic peroxidase (Hilaire et al. 2001). Some of these are toxic and can serve as chemical inhibitors restricting pathogen growth and reproduction (Shi et al. 1992; Williams et al. 2002). Moreover, xylem parenchyma cells are involved in callose synthesis and deposition along cell walls and pit membranes neighboring the infected vascular elements (Mueller et al. 1994; Benhamou 1995), as well as in the synthesis of pathogenic-related proteins (Benhamou 1995). They also induce a thickening of secondary walls that decreases pit diameter and limits bacteria spreading via pit membranes into living cells (Hilaire et al. 2001). The few examples mentioned clearly illustrate the significance of living xylem parenchyma cells in physical and chemical defense, assuring the high resistance of plants against various pathogens.
Xylem parenchyma cells perform also a crucial role during the process of heartwood formation (Higuchi et al. 1969; Spicer 2005; Nakaba et al. 2012b). These living cells are involved in vessel occlusions, due to tylose formation or gel secretion (Chattaway 1949) as well as in the production and accumulation of secondary metabolites, which are generally known as heartwood substances (Higuchi et al. 1969; Magel et al. 1991; Hillinger et al. 1996; Taylor et al. 2002; Spicer 2005). These compounds are synthesized in situ by parenchyma cells from stored reserves at the sapwood-heartwood boundary (Magel et al. 1991; Yang et al. 2004). Heartwood substances belong mostly to the phenolic compounds (e.g., tannins, terpenes, flavonoids, lignans, and stilbenes) or to lipids (Higuchi et al. 1969; Magel et al. 1991; Hillinger et al. 1996; Taylor et al. 2002). Their accumulation in heartwood leads to wood colorization and enhancement of wood decay resistance (Taylor et al. 2002; Spicer 2005).
4.4.3.5 The Impact of Living Cells on the Growth and Differentiation Processes of the Secondary Xylem
Different types of cells like vessels, tracheids, fiber-tracheids, fibers, and axial and radial parenchymas can be formed and arranged in peculiar and, more usually, in regular spatial patterns in the secondary xylem, with additional variation among the seed plants. The mechanisms responsible for the growth and differentiation of these various xylem cells, and for formation of specific vascular patterns, remain unknown. Possibly, living xylem parenchyma cells can be involved in these processes by acting as a source of signal molecules, which affect the differentiation of xylem elements (Barnett 1982).
Depending on the morphology and distribution of the vessels in the particular growth rings, two major types of wood can be distinguished in angiosperms: diffuse porous or ring porous (Fig. 4.3). In diffuse porous, wood vessels have approximately similar dimensions and are more or less evenly distributed in growth rings. Conversely, the ring-porous wood dimension of the vessels varies considerably during the vegetative season, with large vessels being produced at the beginning of the season, while narrow ones formed in the second part (IAWA 1989). Developmental studies have clearly demonstrated the impact of living xylem cells on the determination of wood architecture. For example, the occurrence of ring-porous wood in Gmelina genus depends on the presence and distribution of living axial parenchyma cells (Chowdhury 1953) that are formed in the beginning of ring growth (initial parenchyma). The lack of initial parenchyma or its scanty distribution results in the diffuse-porous structure of the secondary xylem, whereas abundant initial parenchyma sheathing newly formed vessels leads to the ring-porous structure of the wood (Chowdhury 1953). Such a correlation may suggest that initial parenchyma prolongs the process of vessel formation and influences its radial expansion, resulting in a wider xylem element development of the ring-porous wood. However, the nature of the intercellular signals possibly exchanged between parenchyma and the differentiating xylem elements, as well as their route of transport, are unknown, although the fact that both initial parenchyma cells and differentiating xylem elements are alive suggests the involvement of symplasmic route. This supposition requires further evaluation.
The differentiation processes of xylem elements can be influenced by the plasmodesmata as in the young branches of Sorbus torminalis (Lachaud and Maurousset 1996). In this species, the presence of plasmodesmata in the pit membrane probably maintains symplasmic continuity between differentiating xylem elements and forms a temporary symplasmic domain. It is unknown whether or not plasmodesmata in these pit regions are functional, because no dye-coupling experiments have been done yet. However, it seems highly possible that plasmodesmata may influence wood structure, at least in Sorbus torminalis, because symplasmic continuity maintained between young xylem elements probably hampers their differentiation and leads to the formation a homogenous wood (Lachaud and Maurousset 1996). The development of a homogenous wood structure corresponds well with the general principle that cells, belonging to the same symplasmic domain, differentiate in a similar manner (Ehlers et al. 1999; see Chap. 2). However, the impact of plasmodesmata on the xylem structure appears to be more complex and species dependent due to the fact that the presence of symplasmic communication affects both the growth and the differentiation of xylem cells.
4.5 Future Perspectives
Symplasmic transport in the spatial system of living xylem cells is involved not only in nutrient translocation or substance exchange at the symplasm/apoplasm interface but also in the integration of cell-to-cell communication over long distances in plants. Although our knowledge about the function of living xylem cells and symplasmic transport in wood has recently increased, there are still some questions and problems which need further examination. It cannot be forgotten that transport in seed plants is composed of integrated symplasmic and apoplasmic systems which cooperate to assure proper plant development and function. Thus, the efficiency and velocity of symplasmic transport in wood needs to be elucidated to fully explain transport capacities in woody plants. Although the general axial and radial routes of symplasmic transport in secondary xylem were observed using small molecular tracers, studies of possible macromolecule trafficking between living xylem cells, and their impact on xylem element differentiation and wood structure, are also necessary. Furthermore, symplasmic transport in secondary xylem not only is coordinated at the cellular level, by the mechanisms that act during typical plasmodesmata trafficking, but can also be regulated on the supracellular level. Taking into account the spatial regulation of symplasmic transport in active cambium (Sokołowska and Zagórska-Marek 2012) or the wavy propagation of solutes in the phloem tubes (Thompson and Holbrook 2004), the supracellular coordination of symplasmic transport in wood can be possible and facilitate fast and efficient communication over long distances in trees. Thus, knowledge about the mechanisms of symplasmic transport in wood and the comprehensive role of living xylem cells is indispensable in plant biology, leading to a full understanding of how such long-lived plants as trees can grow and function properly.
References
Alves G, Sauter JJ, Julien J-L, Fleurat-Lessard P, Ameglio T, et al. Plasma membrane H+-ATPase, succinate and isocitrate dehydrogenases activities of vessel-associated cells in walnut trees. J Plant Physiol. 2001;158:1263–71.
Alves G, Decourteix M, Fleurat-Lessard P, Sakr S, Bonhomme M, Améglio T, et al. Spatial activity and expression of plasma membrane H+-ATPase in stem xylem of walnut during dormancy and growth resumption. Tree Physiol. 2007;27:1471–80.
Améglio T, Ewers FW, Cochard H, Martignac M, Vandame M, Bodet C, et al. Winter stem xylem pressure in walnut trees: effects of carbohydrates, cooling and freezing. Tree Physiol. 2001;21:387–94.
Améglio T, Bodet C, Lacointe A, Cochard H. Winter embolism, mechanisms of xylem hydraulic conductivity recovery and springtime growth patterns in walnut and peach trees. Tree Physiol. 2002;22:1211–20.
Améglio T, Decourteix M, Alves G, Valentin V, Sakr S, Julien J-L, et al. Temperature effects on xylem sap osmolarity in walnut trees: evidence for a vitalistic model of winter embolism repair. Tree Physiol. 2004;24:785–93.
Arend M, Weisenseel MH, Brummer M, Osswald W, Fromm JH. Seasonal changes of plasma membrane H+-ATPase and endogenous ion current during cambial growth in poplar plants. Plant Physiol. 2002;129:1651–63.
Arend M, Monshausen G, Wind C, Weisenseel MH, Fromm J. Effect of potassium deficiency on the plasma membrane H+-ATPase of the wood ray parenchyma in poplar. Plant Cell Environ. 2004;27:1288–96.
Banasiak A. Putative dual pathway of auxin transport in organogenesis in Arabidopsis. Planta. 2011;233:49–61.
Bannan MW. Origin and cellular character of xylem rays in gymnosperms. Bot Gaz. 1934;96:260–81.
Bannan MW. Vascular rays and adventitious root formation in Thuja occidentalis L. Am J Bot. 1941;28:457–63.
Bannan MW. Ray contacts and rate of anticlinal division in fusiform cambial cells of some Pinaceae. Can J Bot. 1965;43:487–507.
Barghoorn ES. The ontogenetic development and phylogenetic specialization of rays in the xylem of dicotyledons. II. Modification of the multiseriate and uniseriate rays. Am J Bot. 1941a;28:273–82.
Barghoorn ES. The ontogenetic development and phylogenetic specialization of rays in the xylem of dicotyledons. III. Elimination of rays. Bull Torrey Bot Club. 1941b;68:317–25.
Barnett JR. Plasmodesmata and pit development in secondary xylem elements. Planta. 1982;155:251–60.
Barnett JR. The development of fibre-tracheid pit membranes in Pyrus communis L. IAWA Bull. 1987a;8:134–42.
Barnett JR. Changes in the distribution of plasmodesmata in developing fibre-tracheid pit membranes of Sorbus aucuparia L. Ann Bot. 1987b;59:269–79.
Barnett JR. Cell-cell communication in wood. In: Baluška F, Volkmann D, Barlow PW, editors. Cell-cell channels. New York: Landes Bioscience and Springer Science+Business Media; 2006. p. 135–47.
Barnett JR, Cooper P, Bonner LJ. The protective layer as an extension of the apoplast. IAWA J. 1993;14:163–71.
Begum S, Nakaba S, Oribe Y, Kubo T, Funada R. Induction of cambial reactivation by localized heating in a deciduous hardwood hybrid poplar (Populus sieboldii × P. grandidentata). Ann Bot. 2007;100:439–47.
Benhamou N. Ultrastructural and cytochemical aspects of the response of eggplant parenchyma cells in direct contact with Verticillium – infected xylem vessels. Physiol Mol Plant Pathol. 1995;46:321–38.
Bonhomme M, Peuch M, Ameglio T, Rageau R, Guilliot A, Decourteix M, et al. Carbohydrate uptake from xylem vessels and its distribution among stem tissues and buds in walnut (Juglans regia L.). Tree Physiol. 2009;30:89–102.
Botha CEJ, Aoki N, Scofield GN, Liu L, Furbank RT, White RG. A xylem sap retrieval pathway in rice leaf blades: evidence of a role for endocytosis? J Exp Bot. 2008;59:2945–54.
Botosso PC, Gomes AV. Radial vessels and series of perforated ray cells in Annonaceae. IAWA Bull. 1982;3:39–44.
Braun HJ. The significance of the accessory tissues of the hydrosystem for osmotic water shifting as the second principle of water ascent, with some thoughts concerning the evolution of trees. IAWA Bull. 1984;5:275–94.
Braun HJ, Wolkinger F. Zur funktionellen Anatomie des axialen Holzparenchyms und Vorschläge zur Reform seiner Terminologie. Holzforschung. 1970;24:19–26.
Bucci SJ, Scholz FG, Goldstein G, Meinzer FC, Sternberg LDASL. Dynamic changes in hydraulic conductivity in petioles of two savanna tree species: factors and mechanisms contributing to the refilling of embolized vessels. Plant Cell Environ. 2003;26:1633–45.
Burgert I, Eckstein D. The tensile strength of isolated wood rays of beech (Fagus sylvatica L.) and its significance for the biomechanics of living trees. Trees. 2001;15:168–70.
Burgert I, Bernasconi A, Eckstein D. Evidence for the strength function of rays in living trees. Holz Roh Werkst. 1999;57:397–9.
Burgert I, Bernasconi A, Niklas KJ, Eckstein D. The influence of rays on the transverse elastic anisotropy in green wood of deciduous trees. Holzforschung. 2001;55:449–54.
Carquist S. Comparative wood anatomy. Systematic, ecological and evolutionary aspects of dicotyledon wood. Berlin: Springer-Verlag; 1988.
Carquist S. Bordered pits in ray cells and axial parenchyma: the histology of conduction, storage and strength in living wood cells. Bot J Linn Soc. 2007;153:157–68.
Carquist S. Xylem heterochrony: an unappreciated key to angiosperm origin and diversifications. Bot J Linn Soc. 2009;161:26–65.
Carquist S. Caryophyllales: a key group for understanding wood anatomy character states and their evolution. Bot J Linn Soc. 2010;164:342–93.
Chafe SC. Cell wall formation and “protective layer” development in the xylem parenchyma of trembling aspen. Protoplasma. 1974;80:335–54.
Chafe SC, Chauret G. Cell wall structure in the xylem parenchyma of trembling aspen. Protoplasma. 1974;80:129–47.
Chaffey N, Barlow P. The cytoskeleton facilitates a three-dimensional symplasmic continuum in the long-lived ray and axial parenchyma cells of angiosperms trees. Planta. 2001;213:811–23.
Chaffey N, Barlow P. Myosin, microtubules, and microfilaments: co-operation between cytoskeletal components during cambial cell division and secondary vascular differentiation in trees. Planta. 2002;214:526–36.
Chaffey NJ, Barnett JR, Barlow PW. Visualization of the cytoskeleton within the secondary vascular system of hardwood species. J Microsc. 1997;187:77–84.
Chaffey N, Barnett J, Barlow P. A cytoskeletal basis for wood formation in angiosperm trees: the involvement of cortical microtubules. Planta. 1999;208:19–30.
Chaffey N, Barlow P, Barnett J. A cytoskeletal basis for wood formation in angiosperm trees: the involvement of microfilaments. Planta. 2000;210:890–6.
Chattaway MM. The development of tyloses and secretion of gum in heartwood formation. Aust J Sci Res B. 1949;2:227–40.
Chowdhury KA. The role of initial parenchyma in the transformation of the structure diffuse-porous to ring-porous in the secondary xylem of the genus Gmelina Linn. Proc Natl Inst Sci India. 1953;19:361–9.
Clérivet A, Déon V, Alami I, Lopez F, Geiger J-P, Nicole M. Tyloses and gels associated with cellulose accumulation in vessels are responses of plane tree seedlings (Platanus × acerifolia) to the vascular fungus Ceratocystis fimbriata f. sp platani. Trees. 2000;15:25–31.
Cooper RM, Williams JS. Elemental sulphur as an induced antifungal substance in plant defence. J Exp Bot. 2004;55:1947–53.
Cooper R, Resende MLV, Flood J, Rowan MG, Beale MH, Potter U. Detection and cellular localization of elemental sulphur in disease-resistant genotypes of Theobroma cacao. Nature. 1996;379:159–62.
Cumbie BG. Development and structure of the xylem of Canavalia (Leguminosae). Bull Torrey Bot Club. 1967;94:162–75.
Czaninski Y. Vessel-associated cells. IAWA Bull. 1977;3:51–5.
Decourteix M, Alves G, Brunel N, Améglio T, Guilliot A, Lemoine R, et al. JrSUT1, a putative xylem sucrose transporter, could mediate sucrose influx into xylem parenchyma cells and be up-regulated by freeze-thaw cycles over the autumn-winter period in walnut tree (Juglans regia L.). Plant Cell Environ. 2006;29:36–47.
Decourteix M, Alves G, Bonhomme M, Peuch M, Baaziz KB, Brunel N, et al. Sucrose (JrSUT1) and hexose (JrHT1 and JrHT2) transporters in walnut xylem parenchyma cells: their potential role in early events of growth resumption. Tree Physiol. 2008;28:215–24.
Duckett CM, Oparka KJ, Prior DAM, Dolan L, Roberts K. Dye-coupling in the root epidermis of Arabidopsis is progressively reduced during development. Development. 1994;120:3247–55.
Ehlers K, Binding H, Kollmann R. The formation of symplasmic domains by plugging of plasmodesmata: a general event in plant morphogenesis. Protoplasma. 1999;209:181–92.
El Mahjoub M, Le Picard D, Moreau M. Origin of tyloses in melon (Cucumis melo L.) in response to a vascular fusarium. IAWA Bull. 1984;5:307–11.
Essiamah S, Eschrich W. Changes of starch content in the storage tissues of deciduous trees during winter and spring. IAWA Bull. 1985;6:97–106.
Essiamah S, Eschrich W. Water uptake in deciduous trees during winter and the role of conducting tissues in spring reactivation. IAWA Bull. 1986;7:31–8.
Evert RF. Some aspects of cambial development in Pyrus communis. Am J Bot. 1961;48:479–88.
Ewers FW, Améglio T, Cochard H, Beaujard F, Martignac M, Vandame M, et al. Seasonal variation in xylem pressure of walnut trees: root and stem pressures. Tree Physiol. 2001;21:1123–32.
Fromard L, Babin V, Fleurat-Lessard P, Fromont J-C, Serrano R, Bonnemain J-L. Control of vascular sap pH by the vessel-associated cells in woody species. Plant Physiol. 1995;108:913–8.
Fuchs M, Ehlers K, Will T, van Bel AJE. Immunolocalization indicates plasmodesmal trafficking of storage proteins during cambial reactivation in Populus nigra. Ann Bot. 2010;106:385–94.
Gartner BL, Baker DC, Spicer R. Distribution and vitality of xylem rays in relation to tree leaf area in Douglas-fir. IAWA J. 2000;21:389–401.
Ghouse AKM, Yunus M. Some aspects of cambial development in the shoots of Dalbergia sissoo Roxb. Flora. 1973;162:549–58.
Ghouse AKM, Yunus M. The ratio of ray and fusiform initials in some woody species of the Ranalian complex. Bull Torrey Bot Club. 1974;101:363–6.
Gisel A, Barella S, Hempel FD, Zambryski PC. Temporal and spatial regulation of symplastic trafficking during development in Arabidopsis thaliana apices. Development. 1999;126:1879–89.
Goodman RN, White JA. Xylem parenchyma plasmolysis and vessel wall disorientation caused by Erwinia amylovora. Phytopathology. 1981;71:844–52.
Gregory RA. Cambial activity and ray cell abundance in Acer saccharum. Can J Bot. 1977;55:2559–64.
Gregory RA. Living elements of the conducting secondary xylem of sugar maple (Acer saccharum Marsh.). IAWA Bull. 1978;4:65–9.
Hao B-Z, Wu J-L. Vacuole proteins in parenchyma cells of secondary phloem and xylem of Dalbergia odorifera. Trees. 1993;8:104–9.
Harms U, Sauter JJ. Localization of a storage protein in the wood ray parenchyma cells of Taxodium distichum (L.). L.C. Rich by immunogold labeling. Trees. 1992a;6:37–40.
Harms U, Sauter JJ. Changes in content of starch, protein, fat and sugars in the branchwood of Betula pendula Roth during fall. Holzforschung. 1992b;46:455–61.
Higuchi T, Onda Y, Fujimoto Y. Biochemical aspects of heartwood formation with special reference to the site of biogenesis of heartwood compounds. Wood Res. 1969;48:15–30.
Hilaire E, Young SA, Willard LH, McGee JD, Sweat T, Chittoor JM, et al. Vascular defense responses in rice: peroxidase accumulation in xylem parenchyma cells and xylem wall thickening. Mol Plant Microbe Interact. 2001;14:1411–9.
Hillinger C, Höll W, Ziegler H. Lipids and lipolytic enzymes in the trunkwood of Robinia pseudoacacia L. during heartwood formation. Trees. 1996;10:366–75.
Holbrook NM, Zwieniecki MA, editors. Vascular transport in plants. Amsterdam: Elsevier; 2005.
Höll W. Radial transport in rays. In: Zimmermann MH, Milburn JA, editors. Transport in plants. I. Phloem transport, Encyclopedia of Plant Physiology. New Series, vol. 1. Berlin: Springer-Verlag; 1975. p. 432–50.
Hook DD, Brown CL. Permeability of the cambium to air in trees adapted to wet habitats. Bot Gaz. 1972;133:304–10.
IAWA Committee. IAWA list of microscopic features for hardwood identification. IAWA Bull. 1989;10:219–332.
Jansen S, Sano Y, Choat B, Rabaey D, Lens F, Dute RR. Pit membranes in tracheary elements of Rosaceae and related families: new records of tori and pseudotori. Am J Bot. 2007;94:503–14.
Kim JS, Sandquist D, Sundberg B, Daniel G. Spatial and temporal variability of xylan distribution in differentiating secondary xylem of hybrid aspen. Planta. 2012;235:1315–30.
Kitin P, Fujii T, Abe H, Takata K. Anatomical features that facilitate radial flow across growth rings and from xylem to cambium in Cryptomeria japonica. Ann Bot. 2009;103:1145–57.
Korolev AV, Tomos AD, Bowtell R, Farrar JF. Spatial and temporal distribution of solutes in the developing carrot taproot measured at single-cell resolution. J Exp Bot. 2000a;51:567–77.
Korolev AV, Tomos AD, Farrar JF. The trans-tissue pathway and chemical fate of 14C photoassimilate in carrot taproot. New Phytol. 2000b;147:299–306.
Krabel D. Influence of sucrose on cambial activity. In: Savidge RA, Barnett JR, Napier R, editors. Cell and molecular biology of wood formation. Oxford: BIOS Scientific; 2000. p. 113–25.
Lachaud S, Maurousset L. Occurrence of plasmodesmata between differentiating vessels and other xylem cells of Sorbus torminalis L. Crantz and their fate during xylem maturation. Protoplasma. 1996;191:220–6.
Langenfeld-Heyser R. Distribution of leaf assimilates in the stem of Picea abies L. Trees. 1987;1:102–9.
Langheinrich U, Tischner R. Vegetative storage proteins in poplar. Plant Physiol. 1991;97:1017–25.
LaPasha CA, Wheeler EA. Resin canals in Pinus taeda. Longitudinal canal lengths and interconnections between longitudinal and radial canals. IAWA Bull. 1990;11:227–38.
Larson PR. The vascular cambium. Development and structure. Berlin: Springer-Verlag; 1994.
Lev-Yadun S. Radial fibres in aggregate rays of Quercus calliprinos Webb. – evidence for radial signal flow. New Phytol. 1994;128:45–8.
Lev-Yadun S, Aloni R. Polycentric vascular rays in Suaeda monoica and the control of ray initiation and spacing. Trees. 1991;5:22–9.
Lev-Yadun S, Aloni R. The role of wounding and partial girdling in differentiation of vascular rays. Int J Plant Sci. 1992;153:348–57.
Lev-Yadun S, Aloni R. Differentiation of the ray system in woody plants. Bot Rev. 1995;61:45–84.
Magel EA, Drouet A, Claudot AC, Ziegler H. Formation of heartwood substances in the stem of Robinia pseudoacacia L. Trees. 1991;5:203–7.
Metcalfe CR, Chalk L. Anatomy of dicotyledons. 2nd ed. Wood structure and conclusion of the general introduction. Oxford: Clarendon; 1983.
Mueller WC, Beckman CH. Ultrastructure of the cell wall of vessel contact cells in the xylem of tomato stems. Ann Bot. 1984;53:107–14.
Mueller WC, Morgham AT, Roberts EM. Immunocytochemical localization of callose in the vascular tissue of tomato and cotton plants infected with Fusarium oxysporum. Can J Bot. 1994;72:505–9.
Murakami Y, Funada R, Sano Y, Ohtani J. The differentiation of contact cells and isolation cells in the xylem ray parenchyma of Populus maximowiczii. Ann Bot. 1999;84:429–35.
Myśkow E, Zagórska-Marek B. Ontogenetic development of storied ray pattern in cambium of Hippophaë rhamnoides L. Acta Soc Bot Pol. 2004;73:93–101.
Myśkow E, Zagórska-Marek B. Vertical migration of rays leads to the development of a double-storied phenotype in the cambium of Aesculus turbinata. Botany. 2008;86:36–44.
Myśkow E, Zagórska-Marek B. Dynamics of the ray pattern in cambium of Diospyros lotus L. Dendrobiology. 2013;69:21–30.
Nagai S, Utsumi Y. The function of intercellular spaces along the ray parenchyma in sapwood, intermediate wood, and heartwood of Cryptomeria japonica (Cupressaceae). Am J Bot. 2012;99:1553–61.
Nakaba S, Sano Y, Kubo T, Funada R. The positional distribution of cell death of ray parenchyma in a conifer, Abies sachalinensis. Plant Cell Rep. 2006;25:1143–8.
Nakaba S, Kubo T, Funada R. Differences in patterns of cell death between ray parenchyma cells and ray tracheids in the conifers Pinus densiflora and Pinus rigida. Trees. 2008;22:623–30.
Nakaba S, Begum S, Yamagishi Y, Jin H-O, Kubo T, Funada R. Differences in the timing of cell death, differentiation and function among three different types of ray parenchyma cells in the hardwood Populus sieboldii x P. grandidentata. Trees. 2012a;26:743–50.
Nakaba S, Yamagishi Y, Sano Y, Funada R. Temporally and spatially controlled death of parenchyma cells is involved in heartwood formation in pith regions of branches of Robinia pseudoacacia var. inermis. J Wood Sci. 2012b;58:69–76.
Nardini A, Lo Gullo MA, Salleo S. Refilling embolized xylem conduits: is it a matter of phloem unloading? Plant Sci. 2011;180:604–11.
Oribe Y, Funada R, Shibagaki M, Kubo T. Cambial reactivation in locally heated stems of the evergreen conifer Abies sachalinensis (Schmidt) Masters. Planta. 2001;212:684–91.
Oribe Y, Funada R, Kubo T. Relationships between cambial activity, cell differentiation and the localization of starch in storage tissues around the cambium in locally heated stems of Abies sachalinensis (Schmidt) Masters. Trees. 2003;17:185–92.
Patrick JW, Offler CE. Post-sieve element transport of photoassimilates in sink regions. J Exp Bot. 1996;47:1165–77.
Rabaey D, Lens F, Smets E, Jansen S. The micromorphology of pit membranes in tracheary elements of Ericales: new records of tori or pseudo-tori? Ann Bot. 2006;98:943–51.
Rabaey D, Lens F, Huysmans S, Smets E, Jansen S. A comparative ultrastructural study of pit membranes with plasmodesmata associated thickenings in four angiosperm species. Protoplasma. 2008;233:255–62.
Romberger A, Hejnowicz H, Hill JF. Plant structure: function and development. A treatise on anatomy and vegetative development, with special reference to woody plants. 1st ed. Berlin: Springer-Verlag; 1993.
Sakr S, Alves G, Morillon R, Maurel K, Decourteix M, Guilliot A, et al. Plasma membrane aquaporins are involved in winter embolism recovery in walnut tree. Plant Physiol. 2003;133:630–41.
Salleo S, Lo Gullo MA, Trifilo P, Nardini A. New evidence for a role of vessel-associated cells and phloem in the rapid xylem refilling of cavitated stems of Laurus nobilis L. Plant Cell Environ. 2004;27:1065–76.
Salleo S, Trifilò P, Lo Gullo MA. Vessel wall vibrations: trigger for embolism repair? Funct Plant Biol. 2008;35:289–97.
Salleo S, Trifilò P, Esposito S, Nardini A, Lo Gullo MA. Starch-to-sugar conversion in wood parenchyma of field-growing Laurus nobilis plants: a component of the signal pathway for embolism repair? Funct Plant Biol. 2009;36:815–25.
Sano Y, Okamura Y, Utsumi Y. Visualizing water-conduction pathways of living trees: selection of dyes and tissue preparation methods. Tree Physiol. 2005;25:269–75.
Sauter JJ. Respiratory and phosphatase activities in contact cells of wood rays and their possible role in sugar secretion. Z Pflanzenphysiol Bd. 1972;67:135–45.
Sauter JJ. Seasonal variation of sucrose content in the xylem sap of Salix. Z Pflanzenphysiol Bd. 1980;98:377–91.
Sauter JJ. Evidence for sucrose efflux and hexose uptake in the xylem of Salix. Z Pflanzenphysiol Bd. 1981;103:183–7.
Sauter JJ. Efflux and reabsorption of sugars in the xylem. I. Seasonal changes in sucrose efflux in Salix. Z Pflanzenphysiol Bd. 1982;106:325–36.
Sauter JJ. Efflux and reabsorption of sugars in the xylem. II. Seasonal changes in sucrose uptake in Salix. Z Pflanzenphysiol Bd. 1983;111:425–40.
Sauter JJ. Seasonal changes in the efflux of sugars from parenchyma cells into the apoplast in poplar stems (Populus x canadensis ‘robusta’). Trees. 1988;2:242–9.
Sauter JJ. Photosynthate allocation to the vascular cambium: facts and problems. In: Savidge RA, Barnett JR, Napier R, editors. Cell and molecular biology of wood formation. Oxford: BIOS Scientific; 2000. p. 71–83.
Sauter JJ, Kloth S. Plasmodesmatal frequency and radial translocation rates in ray cells of poplar (Populus x canadensis Moench ‘robusta’). Planta. 1986;168:377–80.
Sauter JJ, van Cleve B. Biochemical, immunochemical, and ultrastructural studies of protein storage in poplar (Populus x canadensis ‘robusta’) wood. Planta. 1990;183:92–100.
Sauter JJ, van Cleve B. Storage, mobilization and interrelations of starch, sugars, protein and fat in the ray storage tissue of poplar trees. Trees. 1994;8:297–304.
Sauter JJ, Iten W, Zimmermann MH. Studies on the release of sugar into the vessels of sugar maple (Acer saccharum). Can J Bot. 1973;51:1–8.
Sauter JJ, Wisniewski M, Witt W. Interrelationships between ultrastructure, sugar levels, and frost hardiness of ray parenchyma cells during frost acclimation and deacclimation in poplar (Populus x canadensis Moench ‘robusta’) wood. J Plant Physiol. 1996;149:451–61.
Schmitt U, Liese W. Wound reaction of the parenchyma in Betula. IAWA Bull. 1990;11:413–20.
Šćukanec V, Petrić B. The relationship between wood ray shape and ray volume percentage in beech. IAWA Bull. 1977;3:57–9.
Secchi F, Zwieniecki MA. Sensing embolism in xylem vessels: the role of sucrose as a trigger for refilling. Plant Cell Environ. 2011;34:514–24.
Secchi F, Zwieniecki MA. Analysis of xylem sap from functional (non-embolized) and non-functional (embolized) vessels of Populus nigra – chemistry of refilling. Plant Physiol. 2012;160:955–64.
Secchi F, Gilbert ME, Zwieniecki MA. Transcriptome response to embolism formation in stems of Populus trichocarpa provides insight into signaling and biology of refilling. Plant Physiol. 2011;157:1419–29.
Sharkey PJ, Pate JS. Selectivity in xylem to phloem transfer of amino acids in fruiting shoots of white lupin (Lupinus albus L). Planta. 1975;127:251–62.
Shi J, Mueller WC, Beckman CH. Vessel occlusion and secretory activities of vessel contact cells in resistant and susceptible cotton plants infected with Fusarium oxysporum f.sp. vasinfectum. Physiol Mol Plant Pathol. 1992;40:133–47.
Sinnott EW. Factors determining character and distribution of food reserve in woody plants. Bot Gaz. 1918;66:162–75.
Sokołowska K, Zagórska-Marek B. Seasonal changes in the degree of symplasmic continuity between the cells in cambial region of Acer pseudoplatanus and Ulmus minor. Acta Soc Bot Pol. 2007;76:277–86.
Sokołowska K, Zagórska-Marek B. Symplasmic, long-distance transport in xylem and cambial regions in branches of Acer pseudoplatanus (Aceraceae) and Populus tremula x P. tremuloides (Salicaceae). Am J Bot. 2012;99:1–11.
Song K, Liu B, Jiang X, Yin Y. Cellular changes of tracheids and ray parenchyma cells from cambium to heartwood in Cunninghamia lanceolata. J Trop For Sci. 2011;23:478–87.
Spicer R. Senescence in secondary xylem: heartwood formation as an active developmental program. In: Holbrook NM, Zwieniecki M, editors. Vascular transport in plants. Oxford: Elsevier; 2005. p. 457–75.
Spicer R, Holbrook NM. Within-stem oxygen concentration and sap flow in four temperature tree species: does long-lived xylem parenchyma experience hypoxia? Plant Cell Environ. 2005;28:192–201.
Spicer R, Holbrook NM. Parenchyma cell respiration and survival in secondary xylem: does metabolic activity decline with cell age? Plant Cell Environ. 2007;30:934–43.
Street PFS, Robb J, Ellis BE. Secretion of vascular coating components by xylem parenchyma cells of tomatoes infected with Verticillium albo-atrum. Protoplasma. 1986;132:1–11.
Sun Q, Kobayashi K, Suzuki M. Intercellular space system in xylem rays of pneumatophores in Sonneratia alba (Sonneratiaceae) and its possible functional significance. IAWA J. 2004;25:141–54.
Sun Q, Rost TL, Matthews MA. Wound-induced vascular occlusions in Vitis vinifera (Vitaceae): tyloses in summer and gels in winter. Am J Bot. 2008;95:1498–505.
Tarpley L, Vietor DM. Compartmentation of sucrose during radial transfer in mature sorghum culm. BMC Plant Biol. 2007;7:33.
Taylor AM, Gartner BL, Morrell JJ. Heartwood formation and natural durability – a review. Wood Fiber Sci. 2002;34:587–611.
Thompson MV, Holbrook NM. Scaling phloem transport: information transmission. Plant Cell Environ. 2004;27:509–19.
Tomos D, Korolev A, Farrar J, Nicolay K, Bowtell R, Köckenberger W. Water and solute relations of the carrot cambium studied at single-cell resolution. In: Savidge RA, Barnett JR, Napier R, editors. Cell and molecular biology of wood formation. Oxford: BIOS Scientific; 2000. p. 101–12.
van Bel AJE. Xylem-phloem exchange via the rays: the undervalued route of transport. J Exp Bot. 1990;41:631–44.
van Bel AJE, van der Schoot C. Primary function of the protective layer in contact cells: buffer against oscillations in hydrostatic pressure in the vessels? IAWA Bull. 1988;9:285–8.
van der Schoot C, van Bel AJE. Architecture of the intermodal xylem of tomato (Solanum lycopersicum) with reference to longitudinal and lateral transfer. Am J Bot. 1989;76:487–503.
van der Schoot C, van Bel AJE. Mapping membrane potential differences and dye-coupling in intermodal tissues of tomato (Solanum lycopersicum L.). Planta. 1990;182:9–21.
Vogelmann TC, Dickson RE, Larson PR. Comparative distribution and metabolism of xylem-borne amino compounds and sucrose in shoots of Populus deltoides. Plant Physiol. 1985;77:418–28.
Williams JS, Hall SA, Hawkesford MJ, Beale MH, Cooper RM. Elemental sulfur and thiol accumulation in tomato and defense against a fungal vascular pathogen. Plant Physiol. 2002;128:150–9.
Wiśniewski M, Ashworth E, Schaffer K. The use of lanthanum to characterize cell wall permeability in relation to deep supercooling and extracellular freezing in woody plants. I. Intergeneric comparison between Prunus, Cornus, and Salix. Protoplasma. 1987;139:105–16.
Yamada Y, Awano T, Fujita M, Takabe K. Living wood fibers act as large-capacity “single-use” starch storage in black locust (Robinia pseudoacacia). Trees. 2011;25:607–16.
Yang K-C. The fine structure of pits in yellow birch (Betula alleghaniensis Britton). IAWA Bull. 1978;4:71–7.
Yang J, Kamdem DP, Keathley DE, Han K-H. Seasonal changes in gene expression at the sapwood-heartwood transition zone of black locust (Robinia pseudoacacia) revealed by cDNA microarray analysis. Tree Physiol. 2004;24:461–74.
Ziegler H. Storage, mobilization and distribution of reserve material in trees. In: Zimmermann MH, editor. The formation of wood in forest trees. New York: Academic; 1964. p. 303–20.
Zwieniecki MA, Holbrook NM. Confronting Maxwell’s demon: biophysics of xylem embolism repair. Trends Plant Sci. 2009;14:530–4.
Acknowledgements
I wish to thank the colleagues from the Department of Plant Developmental Biology, University of Wrocław, Poland, for their discussions and especially Drs. E. Gola, E. Myśkow, and A. Dolzblasz for critical reading of the manuscript. Financial support came from the University of Wrocław, Poland (No. 2776/W/IBR/07).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer Science+Business Media New York
About this chapter
Cite this chapter
Sokołowska, K. (2013). Symplasmic Transport in Wood: The Importance of Living Xylem Cells. In: Sokołowska, K., Sowiński, P. (eds) Symplasmic Transport in Vascular Plants. Springer, New York, NY. https://doi.org/10.1007/978-1-4614-7765-5_4
Download citation
DOI: https://doi.org/10.1007/978-1-4614-7765-5_4
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4614-7764-8
Online ISBN: 978-1-4614-7765-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)