
Abstract
The landscapes of the Mid-Atlantic Region are dissected by networks of rivers, with their associated wetlands and riparian areas. These systems provide important ecosystem services, both ecological functions and societal values, such as floodwater storage, public water supplies, recreational greenbelts, and habitats for a diversity of flora and fauna. Ecologists and hydrologists have increasingly focused on integrating across the four dimensions of these ecosystems: longitudinal, lateral, hyporheic, and time. Here, we use a hydrogeomorphic classification system to describe wetland types, and present a conceptual model of how they connect to other waters through critical components such as hydrologic connectivity, energy flows and sources, and biological integrity. The connectivity of aquatic habitats is described and related in a watershed context. Concepts are supported by a technical review of pertinent literature. We emphasize the flow of water, nutrients, and organisms from headwaters downstream through an interconnected riverine ecosystem.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1.1 Introduction
The landscapes of the eastern USA, including the Mid-Atlantic Region (MAR), are dissected by networks of rivers, with their associated wetlands and riparian areas. These systems historically have provided important ecosystem services, both ecological functions and societal values, to the inhabitants, and continue to provide for floodwater storage, public water supplies, recreational greenbelts, and habitats for a diversity of flora and fauna. Yet 70–90% of the waterways of the eastern USA have been drastically altered by human activities, and significant amounts of the original wetlands of the eastern states, varying from 30 to 70% among the different states, have been lost (Dahl 1990).
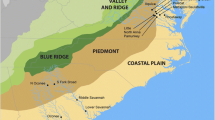
Administratively, the MAR usually includes the states of Pennsylvania, Delaware, Maryland, Virginia, and West Virginia. We define the Mid-Atlantic with somewhat more porous boundaries and an emphasis on the drainage basins of major rivers, ranging in geographic extent east-to-west from the Atlantic beaches and marshes to the top of the Appalachian Mountains, and including the western slopes draining into the Ohio River Basin, approximately to the Pennsylvania–Ohio border. Then, proceeding north-to-south from the most northerly extent of Susquehanna and Delaware river basins in New York and New Jersey, and finally south to the northern portions of North Carolina that drain into the Ablemarle-Pamlico system; this is our region of study (Fig. 1.1).

Extended study area of the Mid-Atlantic Region (MAR) showing boundaries of states, ecoregions, and major river basins
The importance of the headwater portions of watersheds to the overall health of aquatic ecosystems in this region cannot be over emphasized. In the MAR, headwaters typically comprise about 67–75% of the contributing area of any given watershed. That is, the combined areas of terrestrial habitats, wetlands, floodplains, and headwater streams occupy two-thirds to three-quarters of the total area of the drainage basin for larger rivers (Fig. 1.2). Given this influence on downstream portions of large river watersheds, understanding the impacts of human activities on the ecological structure and function of headwater or tributary watersheds is foundational for optimizing their conservation and management. That said, it is also critically important to understand the linkages and contributions of headwaters to the downstream portions of watersheds where large rivers and broader floodplains dominate.

Representations of the relative contributions of stream order to (a) watershed area and (b) flooding and discharge
Increasingly, during the past decade, ecologists and hydrologists have moved toward integrating the traditional studies of the upstream–downstream gradient of rivers (i.e., longitudinal or first dimension, river channel, streambanks) with lateral or second dimension (e.g., floodplains, riparian corridors, wetlands) and vertical or third dimension (e.g., groundwater flows, hyporheic zone) portions to represent a more comprehensive view of all the aquatic components of watersheds, and their interactions with terrestrial areas (e.g., Ward et al. 2002; Naiman et al. 2005) (Fig. 1.3). The separation of constituent aquatic components of watersheds is common in regulatory and management contexts where streams and rivers are treated as separate entities from lakes, wetlands, and estuaries. In fact, such a separation was more for the convenience of defining and managing these units, than it was based on any ecological principles. In this chapter, we discuss the important hydrological and ecological linkages between streams, rivers, and their adjacent floodplains and wetlands, together, comprising a riverine ecosystem. When the interactions with terrestrial components are considered, a holistic aquatic landscape can be visualized.

Longitudinal, vertical and lateral dimensions of riverine ecosystems incorporating riverine, wetland, and riparian components (modified from Ward (2002) by S. Yetter)
Historically, rivers have been variously portrayed as continuous linear (upstream–downstream) gradients (e.g., the River Continuum Concept (RCC), Vannote et al. 1980; Minshall et al. 1985), or as a series of distinct, interconnected habitat patches (e.g., Link Discontinuity Concept, Rice et al. 2001). In addition, alternative conceptual models have evolved seeking to classify stream networks (Frissell et al. 1986; Rosgen 1994), explain the physical heterogeneity of rivers (natural flow regimes, Poff et al. 1997; river discontinua, Poole 2002; network dynamics hypothesis, Benda et al. 2004), describe material cycling (riverine productivity model, Thorp and DeLong 1994, 2002; nutrient spiraling, Newbold et al. 1982; process domains, Montgomery 1999), and perturbations (intermediate disturbance hypothesis applied to rivers, Townsend et al. 1997). Of particular value to this discussion focused on wetlands are the ideas that characterize riverine ecosystems as a series of interconnected hydrogeomorphic (HGM) patches (Church 2002; Poole 2002; Thorp et al. 2006) and the relationship of these dynamic patches to aquatic biodiversity (Townsend et al. 1997; Lake 2000; Ward et al. 2002; Thorp et al. 2006).
Increasingly, these syntheses have begun to move beyond the stream or river channel alone, to incorporating linkages between streams and the landscape in which they flow, thus recognizing longitudinal, lateral, and vertical aspects of the riverine network (e.g., Forman 1995; Ward et al. 2002; Wiens 2002; Naiman et al. 2005). Still missing, however, are attempts to create conceptual models that directly integrate stream, wetland, riparian, and terrestrial components for headwater, tributary, and mainstem portions of watersheds. Within biogeographical constraints, species composition and biological integrity of watersheds are the result of interactions among numerous important instream variables including flow regime, energy source, water quality, instream habitat, and biological interactions. Yet, these variables themselves are largely driven by processes that occur outside of the individual stream channel including weather and climate, geomorphology of the watershed (geology and terrain), and the structure and topology of the surrounding landscape (Karr 1991, 1999). As one moves downstream, instream characteristics are determined by characteristics and processes that occur in upstream areas. Hychka (2010), while exploring the hydrologic responses of wetlands, found that a riverine reach was the spatial unit most appropriate for characterizing the interaction of the stream with the adjacent floodplain and wetlands (Fig. 1.4). It is the magnitude and interplay among these longitudinal, lateral, and vertical processes that form the basis for most conceptual models integrating riverine ecosystems into aquatic landscapes. In this chapter, we will consider these concepts in light of the characteristics of riverine ecosystems that connect to and support freshwater wetlands found in the MAR.

Spatial and temporal scales of relevance for studying wetlands at a reach scale (from Hychka 2010)
1.2 Types of Freshwater Wetlands in the Mid-Atlantic Region
During our studies of MAR wetlands, we have found that starting classification with HGM characteristics (i.e., geomorphic setting, water source, hydrodynamics; Brinson 1993a), combined with portions of the hierarchical classification system used for the National Wetlands Inventory (NWI; Cowardin et al. 1979; Tiner 2003) aids in discriminating among wetland types, particularly during field investigations as opposed to using NWI just for mapping. Early attempts to classify wetlands Cowardin et al. (1979) lumped most vegetated wetlands into a single broadly defined Palustrine system. Using an HGM classification approach, we separate additional types based on their hydrologic characteristics and landscape positions. These HGM classes can be further subdivided by their dominant vegetative life forms; aquatic bed, emergent, shrub, and forest. The additional discrimination of wetland types associated with the HGM approach represents a substantial improvement in wetland classification because it incorporates features that influence wetland function and allows assessment of condition. Our approach to classification and inventory is detailed in Chap. 2 of this book, although terms are introduced here for purposes of introducing concepts and terminology.
The majority (>80%) of freshwater wetlands in the MAR are riverine types, associated with streams and rivers (Brooks et al. 2011a). Some of these riverine wetlands occur as narrow terraces or vegetated islands within the defined channel banks. Most, however, are found in the adjacent floodplain. If a river is free-flowing and it’s channel is not incised, then it interacts with the adjacent floodplain when discharge exceeds channel capacity. Major flood pulses move laterally away from the river channel into the floodplain, and in the process initiate a series of important biogeophysical processes (Junk et al. 1989; Sparks et al. 1990; Bayley 1995).
In headwater portions of a watershed, the most relevant HGM subclasses of wetlands are topographic slopes, depressions, and riverine upper perennial floodplains (which together compose riverine headwater complexes). Perennial and seasonal depressions, some of which are isolated or have subsurface connections, but no surface water connections, and stratigraphic slopes further uphill (e.g., springs and seeps) may contribute to base flow of headwaters. In the lower portions of a watershed, riverine lower perennial types, consisting primarily of the wetland portions of expansive floodplains dominate the system. In some rivers, instream wetlands occur within the defined channel itself, either as low terraces or vegetated islands. Throughout a given watershed, ponds, lakes, and reservoirs (lacustrine types), either naturally formed or impounded, may provide direct hydrologic support or even flow-through connections to streams and rivers.
During the flood pulse as water flows across the floodplain, sediments are deposited and nutrients have an opportunity to be transformed. Pulses of increased flow in the channel that do not overtop most of the riparian banks to inundate the floodplain occur at higher frequencies than overbank flooding events. These are important for replenishing or maintaining sufficient water levels in remnant channels and interconnected depressions. More aptly described as flood flows or minor flood pulses, these events supply water to communities of aquatic macroinvertebrates and wetland plants that occur in the low-lying sections of floodplains (see Chap. 10).
Following the lead of the NWI (Cowardin et al. 1979; Tiner 2003), we divide wetlands associated with riverine reaches of a watershed into upper perennial (i.e., wetlands along smaller tributaries typically with higher gradients; headwaters with or without floodplains) and lower perennial types (i.e., wetlands along rivers with well-developed floodplains, typically with lower gradients) (Brooks et al. 2011a).
Lakes are not common water bodies in the MAR, and, therefore, lacustrine (or fringing) wetlands are not abundant compared to the Great Lakes or New England regions. The greatest density of fringing wetlands occurs along natural and hydrologically altered lakes and ponds in the glaciated ecoregions of northeastern and northwestern Pennsylvania, northern New Jersey, and southern New York. Reservoirs, impounded for water supplies and/or flood control, increase the area of open water and fringing wetlands throughout the remainder of the MAR. Fringing wetlands are found along the edges of these lakes and reservoirs, usually at depths of <2 m, where light penetration is sufficient to promote the growth of rooted aquatic plants. Although most are comprised of emergent plant communities, shrubs and trees can dominate if suitable conditions prevail.
Flats, by definition, occur where water sources are dominated by precipitation and vertical fluctuations of the water table (Brinson 1993a, b). They typically occur in regions of low topographic relief, such as the coastal plains of Delaware, Maryland, Virginia, and North Carolina. There are other ecoregions of the MAR with landscapes that are relatively flat topographically, including parts of the Allegheny Plateau, and the glaciated regions (Fig. 1.1), but hydrologic regimes in these locales are based on a mix of surface flows and soils saturated with groundwater that are more like other wetland types than characteristic flats. Thus, these wetlands continue to be classified as a mixture of shallow depressions, low gradient slopes, and riverine floodplains, rather than flats.
We have identified two major types of slope wetlands (Stein et al. 2004; Brooks et al. 2011a). Topographic slopes, found at the toe of hillslopes, are expressed groundwater discharges. Stratigraphic slopes, located farther upslope, generally represent geologic discontinuities. Both types tend to have unidirectional flows of groundwater from shallow and deep origins.
Wetland depressions are quite diverse, being formed through a variety of geophysical processes, and varying in area, depth, and permanence. Isolated depressions, by definition, have no surface water connections to other water bodies. Due to their separation from navigable waters, isolated depressions have been a contentious topic in the wetlands regulatory arena. Depressions can occur anywhere in the landscape where a low-lying area collects and stores water in a shallow or deep, bowl-shaped feature.
1.3 Conceptual Model of the Structure and Functions of Riverine Ecosystems
A primary goal of this chapter is to synthesize the salient aspects of existing ecological theory as it relates to wetlands closely associated with rivers. Our synthesis attempts to move toward a holistic understanding of the ecology and management of what could be termed “riparia,” the areas of transition between water and land. We believe that it is useful to characterize watersheds as a mosaic of interconnected HGM patches (Ward and Stanford 1983; Townsend 1989; Ward 1989; Brinson 1993b) or settings that contain a set of functional process zones (FPZs, Thorp et al. 2006). FPZs consist not only of a distinguishable stream reach, but also include the geologic and topographic aspects of the surrounding terrestrial landscape. This can lead to considering streams, wetlands, and riparian areas as definable landscape units at a reach scale that support characteristic biota and that respond predictably to a set of anthropogenic stressors.
As the various elements of the system are discussed in subsequent sections, information about potential stressors is included to assist the reader in understanding their influence on ecological integrity (Bryce et al. 1999). The elements of the conceptual model, including anthropogenic drivers and stressors, are summarized in Fig. 1.5 (modified from Karr 1991; Karr and Chu 1999; Brooks et al. 2006a, b). The structure and function of these ecosystems is considered under the following headings: 1.3.1, 1.3.2, 1.3.3, 1.3.4 and 1.3.5. For the purposes of this chapter, the hypothesized interactive relationships among the stream, wetland, riparian, and upland components of watersheds for different stream orders are illustrated in Fig. 1.6a–e. Key features of these illustrations are the relative contributions to the functioning of these systems by upstream portions of the watershed vs. immediately adjacent or lateral components.


(a–e) Conceptual model of elements in riverine ecosystems. (a) Legend. (b) Headwater stream (second order with floodplain). (c) Stream (third/fourth order with floodplain). (d) River (fifth order with floodplain). (e) Tidal River/Subestuary (with floodplain)
1.3.1 Hydrologic Connectivity in Riverine Ecosystems
In the MAR, there is a surplus of precipitation relative to rates of evapotranspiration (i.e., P>ET, see Chap. 3 of this book), thus, many stream and river channels are perennial, receiving sufficient groundwater discharge to sustain base flow throughout the year. Headwater streams (defined here as Strahler (1952, 1957) first through third orders) often occur in proximity to depressions and slopes such that collectively they encompass the type we call a riverine headwater complex. The same phenomenon can contribute to the hydrologic regime of floodplains of larger rivers, referred to as lower perennial wetlands, or a riverine floodplain complex. In these HGM settings, the discharge of groundwater or underlying shallow water table may form a broader band of hydrologically similar conditions such that larger areas of forested wetlands or emergent marshes are formed.
Typically, sustained high flows occur in the late winter and early spring due to either snowmelt or spring rains that often come when water levels are already near peak (Fig. 1.7). There are, of course, exceptions, with singular events like severe thunderstorms or hurricanes precipitating torrential rains over short periods of hours or days. In either case, when flood levels are high, overbank flooding can occur rapidly or over sustained periods of time, and the kinetic energy of these lateral flows is correspondingly strong. This provides the power to erode floodplain soils, alter existing remnant oxbows, cut through sharp meanders of the current river channel, carve new channels, and create vertical banks. The resulting changes in surface flow patterns are influenced by existing floodplain topography (e.g., past channels, areas of dense vegetation, erosion-resistant rock or compact soils) and those interactions modify the location, shape, depth, and size of wetlands within the floodplain. Dams or bank burrows created by beaver, debris piles, or large tree windrows can all influence the directional pathways and force of water flows across the floodplain.

Typical discharge curve for a river in central Pennsylvania showing higher peaks in winter and spring and lower flows in summer and autumn (Data from U.S. Geological Survey Station 01547700, prepared by S. Yetter)
Higher elevation areas, such as portions of the natural levee, distal upland edges, and upland inclusions within the floodplain tend to divert the flows and resist erosive forces. Low-lying depressions, narrow berms, and old and new channels are more likely to shift their shapes and locations dynamically. Through the combination of erosive and depositional forces, existing wetlands may be deepened, filled, or otherwise altered, and new wetlands and channels may be formed. Shallow depressions are scoured out, with and without inflow or outflow channels to the river. Low elevation areas are flooded for extended periods of time, which may shift plant communities toward more hydrophytic species. When the floodwaters recede, infiltrate, or evaporate, then the duration of inundation or saturation, coupled with the interplay of these surface waters with groundwater, will determine whether a wetland is formed, and if so, of what type.
Flow resistance on a floodplain, and hence, its ability to withstand the high kinetic energy of flood pulses and flood flows, is based on the presence and amount of coarse woody debris (CWD), microtopography, and vegetation. The size and density of plant stems (e.g., tree trunks vs. dense grasses) introduces impediments to surface water flow and reduces the energy of storm runoff. Roughness created by vegetation and other physical features (as represented in roughness coefficients such as those in Arcement and Schneider 1989) slows current velocities, causing water to deposit sediment and debris. High vegetation density corresponds to higher effective roughness, flow resistance, and erosion protection of the system. Areas of dense vegetation, including herbaceous and shrub species, and especially mixed age classes of trees, slow flow and create slack water, allowing the water retained by the floodplain to be available for infiltration and potential groundwater recharge. Microtopographic complexity increases the tortuosity of flow pathways, reduces average velocity and increases the variety of moisture conditions present in a site. This increases the diversity of biogeochemical processes occurring in the wetland, especially nitrogen cycling (Brinson et al. 1995), and the presence of abundant and varied microhabitats (Brooks et al. 2004). CWD in the channel, derived from large trees and snags, blocks flows and modifies flow patterns, accelerating the lateral migration of streams and rivers as is expected in a naturally functioning floodplain.
1.3.2 Hydrology/Floodplain and Channel Morphometry
Patterns of discharge and current velocity are the primary hydrologic determinants of species composition in streams and rivers through their influence on carbon and nutrient transport (Newbold et al. 1982), habitat formation and stability (Giberson and Caissie 1998), and direct effects on species mortality patterns resulting from extreme flow events including floods and droughts (Reice 1984). Similarly, wetlands depend on specific hydrologic flows patterns, energies, and duration to maintain their characteristic flora and fauna. While precipitation is the driving force in initiating a flooding event, the physical characteristics of the drainage basin, hydrology, and geomorphology of the stream–floodplain ecosystem are the primary factors controlling the concentration, spatial distribution, and dispersal rate of floodwaters (Staubitz and Sobashinski 1983). Thus, the amount of flooding in a wetland or floodplain is dependent on climate, topography, channel capacity and slope, soil, and lithology (Novitzki 1989; Brinson 1990; Thorne 1998).
Small streams are more directly influenced by precipitation events and thus are “flashier” than larger rivers (Junk and Welcomme 1990; Benke et al. 2000). Although climate and geology are important, they are generally considered to be similar within a given region and wetland type, which holds true for much of the MAR. Differences in landscape-level characteristics, such as upland land uses and stream size are important characteristics to consider. Site-level indicators, however, can be useful for describing these systems with regard to measures of ecological integrity and for functional assessments (Brinson et al. 1995). riparian–wetland areas function properly when site-level indicators such as adequate vegetation, landforms, or large woody debris are present to dissipate stream energy and improve floodwater retention and groundwater recharge.
The physical characteristics of floodplain wetlands determine the potential of an area to store and manage floodwaters. Wetlands, particularly various types of depressions adjacent to streams, reduce the amount of runoff that reaches the streams by storing runoff from adjoining areas (Demissie and Khan 1993). This desynchronizes water delivery to streams, which decreases the frequency and magnitude of flooding downstream (McAllister et al. 2000). Floodplains provide a broad area for floodwaters to dissipate energy through the reductions of water velocities, flood peaks, and erosion. Floodplain vegetation retards water flow through surface roughness (Arcement and Schneider 1989), although slope remains an overarching variable in determining discharge. Topographic depressions, backwater swamp areas, and low elevation areas behind natural levees trap floodwaters as long-term storage, only to be depleted by subsurface seepage to the channel and evapotranspiration (Owen and Wall 1989). In a recent white paper, the Association of State Floodplain Managers (2008) reversed past guidance by stating that conserving the natural hydrodynamics and vegetative structure of floodplains would do more for protecting these critically important systems than imposing engineered solutions.
It is useful to consider the flow of water and materials from the upper reaches of the watershed to lower reaches. Initially, waters at the watershed boundary begin to accumulate in surface and near-surface areas. Precipitation, surface runoff, and near-surface runoff (i.e., interflow) accumulate in narrow, ephemeral, or intermittent channels (sometimes referred to as zero-order streams), or in headwater depressions. A portion of the precipitation component infiltrates into shallow and deep aquifers. The amount is dependent on the areal extent, vertical structure, and composition of vegetation, soil type, topographic gradients, surficial geology, and coverage of human-built structures. Discharges of shallow and deep groundwater may be expressed at the surface as springs, seeps, and slope wetlands, or below the surface entering directly into streams and wetlands, forming a hyporheic zone. Such discharges generally constitute the base flow to these aquatic systems.
As water tables rise to the surface near headwater stream channels, especially in areas of steep topography, the surface runoff of precipitation is nearly instantaneous and contributes to the flashiness of these streams. This “variable source area” expands and contracts depending on antecedent position of the water table as well as duration and intensity of a storm (Hewlett and Nutter 1970). Downstream from the headwater region, eventually, and somewhat dependent on season and the accumulation of base flow, sufficient water accumulates to sustain the flow as a perennial stream. Whereas the zero-order channel tends to dry out seasonally, first-order streams tend to have a persistent base flow in all but the driest of years, usually in a relatively linear channel with little or no floodplain. These relatively small elements are strongly influenced by the characteristics of the adjacent riparian corridor, including the amount of tree cover, type of soil, or range of stressors present. These influences, separated from inputs originating upstream, can be referred to as lateral effects, keeping in mind that these effects include flood flows moving in the opposite direction, connecting the stream channel back to the floodplain. The correspondence between stream order and perennial flow differs in the coastal plain ecoregion. Because of the flat topography, water tables frequently fall below the elevation of low order stream channels during periods of high evapotranspiration during the growing season. For this reason, channelized stream channels, created through deepening and widening actually have more perennial flow than their natural counterparts because the channel is incised deeper into the water table (Hardison et al. 2009).
As discharge increases, energy also increases to the point where physical modifications to the channel can occur. Pool–riffle complexes develop in the widening channels of tributary streams (second to fourth order) (Forman 1995; Naiman et al. 2005). Floodplains continue to widen as the flow transitions from tributary streams to larger rivers (Fig. 1.3). In these stages, the river itself, and to some extent the adjoining floodplain, are tied more closely to the characteristics and periodicity of the flows that have accumulated from upstream reaches, and less by the activities in the riparian corridor and adjacent contributing watershed. Thus, mid-reach and mainstem portions of the river network become uncoupled from upland hill slopes and the sediments eroded from uplands. Sediments, first deposited and then resuspended and redeposited during flood events across the alluvial floodplain, define the channel and its flow path, and determine where and what type of wetland might be formed. This dependency is represented conceptually in Fig. 1.6a–e by the size of the arrows, which represent the strength of influence.
Long-term surface water storage, for weeks or more, helps to maintain the characteristic hydroperiod of wetlands and streams. Hydroperiod affects just about all components of aquatic ecosystems; plant communities, soil processes, nutrient cycling, and faunal communities are all influenced by the duration and frequency of inundation (Gosselink and Turner 1978, Carter 1986; Tiner 1988). Standard gauging stations have long been used to plot the expected hydrographs for streams and rivers throughout the USA. These data are readily available digitally online, although not all streams are gauged. On a smaller scale, Riparia and others have prepared typical hydrographs of the expected hydrologic regime for making comparisons among wetland subclasses (Fig. 1.8) (e.g., Cole and Brooks 2000; Brooks 2004; Cole unpublished, see Chap. 3 of this book). Deviations from this expected pattern can be used to suggest the presence of watershed stressors.

Typical hydrograph for HGM Wetland Subclass, riverine lower perennial wetland in the floodplain of a fifth order river in central Pennsylvania, showing depth to water table in slotted well (0 cm on left side represents ground level) vs. precipitation (along top of graphs, scale in cm on right side) for: (a) drought year, (b) wet year, and (c) across a 12-year period (Data collected by C.A. Cole, graphs prepared by K.C. Hychka)
When one incorporates components outside the stream channel proper into the riverine model, complexity of the ecosystem increases. The accumulation and flow of water across the landscape coupled with the varied microtopography of these areas results in a river mosaic of hydrologically derived gradients and discontinuities across the surface (Forman 1995; Ward et al. 2002). The wetland components of these mosaics can be referred to as headwater and floodplain complexes (see Chap. 2 of this book for details on classification), whereas previously, wetlands were classified primarily on the dominant vegetation and hydrology (Cowardin et al. 1979). Alternatively, the HGM approach (Brinson 1993a; Smith et al. 1995; Cole et al. 2008; Brooks et al. 2011a) emphasizes physical elements for classifying wetlands and for comparing functions and condition across reference sites.
1.3.2.1 Impacts of Human and Beaver Activities and Alterations on Riverine Ecosystem Hydrology
Human activities upstream influence flood frequency and intensity (McAllister et al. 2000). Urbanization creates impervious surfaces and underground sewers, which accelerate the delivery rate of surface water to the stream (Pennsylvania Environmental Council 1973; Paul and Meyer 2001). As little as 3% impervious cover in a contributing area has been shown to negatively impact the ecological integrity of aquatic ecosystems (e.g., May et al. 1997). Serious declines in biological integrity have been observed when urban land exceeds 7% of total watershed area (Snyder et al. 2003). Channelization, levees, and floodwalls, both on-site and upstream, disconnect or destroy wetland and riparian habitat, restrict river flows, decrease water elevations at low flows, and increase water levels at the same locations during floods (Scientific Assessment and Strategy Team 1994). Channelization restricts flow within the stream channel, rather than allowing overbank flow to spread water across wetlands and decrease velocity (Brown 1988). This results in decreases in the ability of wetlands to perform other functions, such as removing sediment and nutrients, and long-term surface water storage (Johnston et al. 1984; Brown 1988; Rheinhardt et al. 1999). Channelization also alters stream morphometry, which leads to scouring and incision. Highway embankments remove vegetation, eliminate natural storage areas, and reduce space available for floodwater storage (Owen and Wall 1989). These and other activities often result in channel degradation, which lessens the depth, frequency, duration, and predictability of flooding. The floodplain frequently becomes increasingly isolated from the stream channel through incision and no longer has the opportunity to perform this function. These activities not only impair performance on-site, but they also increase the flood pulse downstream, a process that places additional pressure on downstream wetlands to dissipate energy and temporarily detain floodwaters. Even in urban areas where intensified peak flows lead to overbank events in incised channels, flooding is too brief to effectively saturate floodplain sediments and substantially raise water tables to permit wetland biogeochemical functioning (Hardison et al. 2009).
Changes in the structure and composition of surrounding landscape, particularly forests, can also have large effects on stream flow. For example, complete removal of forest vegetation associated with logging dramatically increases annual water yields and bank flow flood frequencies (Swank et al. 1988). In addition, human or pest-induced changes to the composition of surrounding forests can alter stream flow. For example, the hemlock woolly adelgid (HWA), an exotic insect forest pest that kills eastern hemlock trees, has been identified in numerous Mid-Atlantic watersheds. The pest is expected to cause significant and perhaps complete hemlock mortality. In a study designed to determine potential effects of HWA-induced hemlock decline on headwater streams in Delaware Water Gap National Recreation Area (DWG), Snyder et al. (2002) found that headwater streams draining hemlock forests were less likely to dry up completely during drought years than similar streams draining mixed hardwood forests. Based on their findings, they predict that HWA will have dramatic effects on headwater stream hydrology, and thus, biodiversity.
Channelization increases the rate of runoff, which increases peak flow, and decreases water storage and the residence time of water (Brown 1988). Studies show that increases in water level fluctuation relate directly to increases in runoff from adjacent uplands (Euliss and Mushet 1996). Human alterations also cause an increase in the amount of sediment transported in a stream and ultimately across a floodplain. Excess sediment can fill critically important interstitial spaces in the substrate of streams, reducing or eliminating aquatic biota (e.g., larvae of aquatic insects, salamanders, and fishes). This same source of sediment may result in the filling of depressions or reduce plant germination rates (Mahaney et al. 2004), and hence, cause a reduction in the storage capacity and topographic complexity of wetlands and on the floodplain in general.
A discussion of headwater wetlands would not be complete without describing the role of beaver. The primary effect of beaver ponds is to expand the area of wetlands in headwater streams, often by several-fold. Several studies in the MAR have demonstrated both the habitat and the water quality consequences of beaver pond establishment in the Adirondack Mountains of New York (Cirmo and Driscoll 1993), the Appalachian Plateau of Pennsylvania and Maryland (Margolis et al. 2001) and the coastal plain of Maryland (Correll et al. 2000) and North Carolina (Bason 2004; Bason and Brinson in preparation). These and studies in other geographical regions demonstrate an expansion of aquatic habitat, conversion of riparian forest and adjacent upland forest to open water and herbaceous wetland vegetation, and the concomitant shift in species composition toward more abundant waterfowl, wading birds, and a host of amphibians and reptiles. Effects on water quality are profound and represent an amplification of the biogeochemical processes discussed below in the section on Water Quality and Biogeochemistry. Principal among these effects is the removal of nitrate, presumably by denitrification, due to the detention of water in the ponds (relative to a stream channel) and the expansion of surface area of organic-rich sediments (Bason and Brinson in preparation).
1.3.3 Energy Flow and Sources
The changes in the relative importance of energy sources and the associated changes in plant and animal species structure and composition is the basis of the RCC. Essentially, the RCC proposes that, in unperturbed watersheds, stream communities change in predictable ways as we move from headwaters to large rivers and these changes are mediated by a continuum of physical gradients that control the amount and sources of energy (Vannote et al. 1980). It is important to understand these aspects of headwater stream to understand how they potentially interact with adjacent wetlands.
Within the headwater stream component of tributary watersheds (zero through second order), the source of detrital energy is mainly from outside the stream channel (i.e., allochthonous inputs), largely in the form of leaf litter, or coarse particulate organic matter (CPOM) (Cummins et al. 1973). The quantity, quality, and timing of leaf litter inputs vary depending on regional climate and the structure and composition of the surrounding forests. In watersheds draining deciduous forests (which predominate in Mid-Atlantic watersheds), the majority of leaf litter inputs to streams occur in autumn when deciduous trees naturally lose their leaves. However, in watersheds comprising mainly coniferous forests, inputs may be more consistent throughout the year. The source of leaves can originate from either upland forests or forested wetlands. Once leaves or conifer needles fall into headwater streams, a large fraction of the associated carbon is rapidly dissolved or leached directly into the water as dissolved organic matter (DOC) and transported to downstream reaches with flow. The CPOM tends to accumulate in pools and stream margins into leaf packs where they are colonized and undergo decomposition by bacteria and aquatic fungi, a process termed conditioning (Cummins and Klug 1979). These incompletely decomposed but conditioned leaves are then available as food for aquatic macroinvertebrates, which in turn supports the production of fish and other secondary consumers. In addition to being used directly by microbial and macroinvertebrate assemblages, a significant fraction of the CPOM component is broken down into smaller particles or fine particulate organic matter (FPOM), by the abrasive forces of stream flow and by the feeding activity of leaf-shredding macroinvertebrates (Boling et al. 1975; Iversen et al. 1982). Subsequently, FPOM is suspended into the water column and exported to stream reaches downstream (e.g., Neatrour et al. 2004).
In forested watersheds, the quantity of leaf litter that enters streams is not limiting. However, the extent to which litter inputs are available to stream communities depends on two factors. The first is the extent to which headwater streams can retain CPOM within headwater reaches in the face of downstream flow. Although increases in flow associated with storms are responsible for most export of CPOM from headwater streams on an annual basis (Schlesinger and Melack 1981), correlations between discharge and transport of organic carbon are weak in headwater streams, especially during non-storm periods indicating the importance of retention. There are many factors that influence organic matter retention including biological uptake. However, the physical structure and complexity of stream channels have been implicated as primary determinants. Specifically, physical features of streams such as boulders and a stream channel that allows floodwaters to overflow their banks, slow the transport of water and materials downstream. Of particular note is the role that CWD plays in retaining particulate organic matter within headwater stream reaches (e.g., Bilby and Likens 1979; Wallace et al. 1995; Brookshire and Dwire 2003). Recent research indicates that undisturbed watersheds contain more CWD and are more retentive of carbon than disturbed watersheds, thus enhancing the availability of organic material to benthic consumers (Wallace et al. 2001; Scott et al. 2002).
The second factor that affects organic matter availability in headwater streams is the species composition of the leaf litter itself. Specifically, leaves of different plant species break down at different rates due in large measure to the chemical characteristics, especially nitrogen and fiber content, of the leaves (Webster and Benfield 1986). In an extensive study of leaf breakdown in streams, Peterson and Cummins (1974) found wide variation in leaf breakdown rates among species and suggested that this variation ensured that carbon was available to secondary consumers throughout the year. Conifer needles break down much slower than deciduous species and the leaves of herbaceous plant break down faster than those of woody plants. Thus, disturbances that change the composition of the surrounding riparian area would be expected to result in changes in the amount and timing of organic carbon available to stream communities. Forest pests such as HWA and gypsy moth are both common in some Mid-Atlantic watersheds and can have dramatic effects on the species composition of riparian forests. In addition, numerous abiotic factors have been shown to affect litter decomposition rates within plant species. For instance, litter breakdown rates are positively correlated with temperature and dissolved nutrients, and negatively correlated with acidity and various toxic effluents. Consequently, factors that reduce water quality are also expected to significantly alter the energy pathway in headwater streams and lead to disruptions in ecological integrity. Water quality of stream and wetland resources is a major concern throughout the various ecoregions of the MAR.
Further downstream in the mid-reaches (third and fourth order), stream channels begin to widen which allows more light to penetrate the forest canopy and reach the stream bottom. At this point, the RCC predicts instream primary production (i.e., autochthonous inputs) becomes an important energy source mainly in the form of benthic diatoms (Molloy 1992). In addition, FPOM derived and exported from upstream reaches also represents a significant energy source to stream biota. Thus, in unperturbed mid-reaches, direct litter inputs from the riparian zone diminish in importance and instream primary production and carbon inputs derived from upstream reaches become more important carbon sources to fuel secondary production. In addition to increased production and diversity of benthic algae in mid-reach streams, the composition of macroinvertebrate assemblages also change in response to changing sources of energy. Therefore, from an energy perspective, the ecological integrity of stream communities in mid-reach streams is determined mostly by factors that affect retention, transport, and the quality of organic matter from headwater areas upstream, and by factors that influence instream primary production within mid-reach areas. In particular, the effects of nonpoint source pollutants associated with agriculture and urban land use in upstream or adjacent landscapes, of significant concern in the MAR, have been shown to affect energy pathways in mid-reach areas. The colonization and movement of macroinvertebrates between streams and the adjoining floodplain and wetlands is discussed further in Chapter 10.
Herbicides and increased sediment inputs have been shown to reduce overall instream primary production with subsequent changes in macroinvertebrate diversity and production (Georgian and Wallace 1983; Guasch et al. 1998). Also, nutrient enrichment from agriculture has been shown to cause a shift in benthic algal composition from an assemblage dominated by diatoms, a preferred food source of many macroinvertebrate species, to an assemblage dominated by filamentous green and blue-green algae that detritivores mostly avoid (Hart and Robinson 1990; Jacoby et al. 1990). Acidification of stream habitats has also been shown to alter primary production in streams (e.g., Planas and Moreau 1986). Brooks et al. (2009) showed that for watershed throughout the MAR, the aforementioned set of stressors occurring collectively in wetlands, the floodplains, and in the stream channel, negatively impact both benthic macroinvertebrate and fish communities.
The RCC also makes specific predictions regarding energy sources in larger rivers and the associated responses of biological communities. Essentially, the RCC predicts that in larger rivers (>4th order) the primary energy source fueling secondary production is derived from terrestrial inputs that were inefficiently processed by consumers in headwaters and mid-reach sections of the drainage network and exported downstream. However, the data from larger rivers show less agreement with the predictions of the RCC than do headwater and mid-reach streams (Thorp et al. 2006). Consequently, other models have been proposed to explain energy pathways and food webs in larger rivers. The two that have received considerable attention are the Flood Pulse Concept (FPC, Junk et al. 1989) and the Riverine Productivity Model (RPM, Thorp and Delong 2002). The FPC emphasizes linkages between the river channel and the floodplain arguing that most secondary production is attributed to organic matter directly or indirectly derived from the periodic flooding of floodplain vegetation and aquatic macrophytes. Thus, the FPC explicitly singles out riparian wetlands as important drivers of large river food webs.
Like the FPC, the RPM recognizes that by the time organic matter, leaked from upstream areas, made it to large rivers (as proposed by the RCC) it is highly refractory and thus probably not sufficient to support secondary production in large rivers. Rather, Thorp and Delong (2002) postulated in the RPM that most large river consumers preferentially assimilate autochthonously derived carbon (i.e., algae and phytoplankton), and to a lesser extent, direct inputs from riparian areas. The RPM also explicitly recognizes the importance of impoundments and reservoirs to large river food webs. There is empirical support for both the FPC (e.g., Bayley 1989; Junk et al. 1989) and the RPM (Bunn et al. 2003; Delong and Thorp 2006), and it appears that the most appropriate of the two models for a given river depends on landscape-scale hydrologic characteristics (Hoeinghaus et al. 2007). Specifically, landscapes that result in low-gradient floodplain rivers are fueled primarily by aquatic macrophyte production associated with wetlands and riparian areas which supports the FPC, whereas landscapes that result in relatively shallow, high-gradient floodplain rivers are disproportionately driven by algae and phytoplankton which conforms to the RPM. Thus, although neither of these models has been tested sufficiently in MAR rivers, we would expect a higher fraction of the food web to be fueled by algae and phytoplankton in those systems located in mountainous regions such as the Ridge and Valley or Allegheny Highlands physiographic provinces, and by macrophytes and riparian vegetation in low-gradient regions such as the Piedmont and Coastal Plain physiographic provinces.
To summarize, recent evidence indicates that the ultimate carbon sources driving large river food webs primarily originate within the river reach or the adjacent floodplain, and not from POM exported from upstream reaches as postulated in the RCC. This does not imply, however, that large river food webs are not influenced by patterns and processes that occur within upstream portions of the watershed. In fact, disruptions to headwater and mid-reaches can have dramatic effects on riverine hydrology and water quality that would ultimately affect large river food webs. For example, urbanization in upstream portions of watersheds increases the frequency of major floods in large rivers (reviewed in Praskievicz and Heejun 2009) that could destroy or degrade riparian wetlands. Likewise, nonpoint source pollution from upstream has been shown to affect instream primary production in large rivers (e.g., Van Nieuwenhuyse and Jones 1996). Rather, predictions from the FPC and the RPM suggest that management and regulatory programs that rely solely on protecting upstream components of watersheds will, by themselves, not be sufficient to maintain biological integrity of large rivers. The protection of riparian areas (especially wetlands) and preserving floodplain–river channel linkages are also required. This new paradigm has significant implications for the conservation of large river biota and further complicates management activities that seek to balance environmental protection with human development. This is because many human activities that routinely take place within or adjacent to large rivers and once thought to be relatively benign to aquatic communities are now believed too highly disruptive. Moreover, many of these activities such as channelization projects to enhance navigation, dikes and levees for flood control, and development within the floodplain are either irreversible or restoration would be prohibitively expensive.
Tributary components of headwater systems are mainly forested under relatively unaltered conditions throughout the MAR. Not only do trees contribute leaf litter and downed wood described below, but forests provide shade, root structure for stream bank stability, and other physical and microclimatic controls to the stream. Wetlands in the riparian zone maintain the organic-rich conditions in the soil that is critical for denitrification in groundwater flowing to stream channels from nitrate sources such as agriculture (Peterjohn and Correll 1984). Forested wetlands support both aquatic and terrestrial food webs; aquatic food webs when they are flooded and terrestrial food webs when they are seasonally dry. As in nearly all terrestrial and aquatic food webs, the detrital food web dominates over grazing pathways (Brinson et al. 1981).
What sets apart headwater streams is the extent to which they are hydrologically connected to riparian wetlands (Fig. 1.6a). In contrast to larger rivers, where large volumes of water flow past floodplain wetlands, both headwater streams and their associate wetlands are minute by comparison, but they form dense dentritic or trellis networks intersecting the terrestrial areas throughout the MAR, often at a density of 1 km of stream per 1 km2 of land surface. The consequence of this proximity and abundance is that they are most exposed to human activities that modify wetland condition. The most pernicious are channelization and ditching (straightening, deepening, and widening), processes that remove most hydrological and biological connections between stream channels and floodplain. The conversion of riparian forest to agriculture, pasture, residential areas, and urban land uses fundamentally changes ecological processes. To the extent that overbank flow during floods connected stream habitat with floodplain habitat, channelization and incision totally disrupts this connection and simplifies the complexity of food webs and the complex pathways of energy. Detrital biomass is an important component of headwater ecosystems and plays a role in nutrient cycling and habitat for plant and animal communities in tributary watersheds. Detrital biomass is represented by snags, down and dead woody debris, organic debris on the forest floor, and organic components of mineral soil. This has been described for wetlands in the national riverine HGM model (Brinson et al. 1995) and regional HGM models (Brooks 2004), and for Mid-Atlantic streams by Barbour et al. (1999) and Boward et al. (1999). Detritus is considered an indicator of the potential decomposition and nutrient cycling rates at a site. Decomposition is generally faster in aquatic than in terrestrial landscapes due to increased leaching, fragmentation, and microbial activity (Shure et al. 1986). Large pieces of CWD derived from adjacent or upstream forests are processed into FPOM and then further processed and incorporated into organic matter (Bilby and Likens 1979; Jones and Smock 1991). Organic material may be transported to channels or respired as CO2 atany stage of the decomposition process (Bilby and Likens 1979; Jones and Smock 1991).
Riverine wetlands are a major source of particulate organic carbon (POC) entering streams. Woody debris is a nutritional substrate, provides habitat for microbes, invertebrates, and vertebrates, is a substrate for seedling growth, and serves as a long-term nutrient reservoir; a consistent source of organic material (Harmon et al. 1986; Brown 1990). POC is a small fraction of total organic carbon (TOC), but ranks disproportionately higher as a food source for fish and invertebrates (Taylor et al. 1990). POC from wetlands contributes substantial amounts of organic matter to stream channels (Mulholland and Kuenzler 1979, Dosskey and Bertsch 1994). In fact, POC comprises between 24 and 46% of the TOC in streams (Dosskey and Bertsch 1994). Detrital inputs to the stream during peak inundation periods support microbial and macroinvertebrate communities in the stream channel (Smock 1990). Although we tend to view movement of materials from uplands and wetlands into headwater streams, there are a significant number of invertebrate and vertebrate taxa moving laterally between streams and their associated floodplains and wetlands. There are macroinvertebrate taxa that are obligate to floodplain wetlands, relying on sufficient flood pulses to create habitats, periodic flood flows to maintain sufficient hydrology, and retention of POC within the wetland itself as a food source (see Chap. 9 of this book, Yetter).
The rate of particulate matter degradation depends on many factors, including soil moisture levels. According to Bilby et al. (1999), when compared to either fully submerged or terrestrial conditions, wood decays at a much faster rate when periodically wetted and dried, conditions typical of many wetlands and floodplains. Floodplains have higher decomposition rates for wood than streams (Cuffney 1988). Forested riparian corridors maintain more benthic habitat, increase channel and bank stability, and provide additional contact area for transforming both nutrients and pesticides than non-forested reaches (Sweeney et al. 2004).
1.3.4 Water Quality and Biogeochemistry
Headwater stream and wetland communities are strongly influenced by the chemistry of the surface and ground waters than those associated with larger rivers (Jones and Mulholland 2000). Natural variation in water hardness, specific conductance, acidity, and dissolved oxygen are all major determinant of species composition, and consequently must be considered when seeking to understand reference conditions and when designing a sampling program to monitoring aquatic resources. This is of interest because human sources of pollution reduce water quality and alter aquatic communities directly by killing or weakening individuals, or by altering energy pathways.
In the headwater portions of watersheds, measures of water chemistry are more reflective of the geologic and topographic characteristics of the landscape than for the lower reaches of larger rivers. The complex geology of the Appalachians, running through several Mid-Atlantic ecoregions, can create circumstances where relatively short stream reaches and individual wetlands can have different water chemistry than their neighbors (USEPA 2000; Snyder et al. 2006). Such variability produces extraordinary biodiversity at a regional scale.
Numerous human sources of water quality degradation have been identified within the region, including urbanization, failing septic systems, agriculture, acid mine drainage (AMD), and acid precipitation. Wetlands and riparian corridors often act as buffers to these water sources due to their ability to filter out and transform contaminants (e.g., Fig.1.6a–e). Of particular importance are nonpoint pollutants, including nutrients such as nitrogen, phosphorus, pesticides, herbicides, and sediments that enter stream and wetland habitats through shallow groundwater and surface runoff.
Eutrophication from excess nutrients (e.g., nitrogen and phosphorus) can be a significant stressor in aquatic ecosystems. Over time, eutrophication typically alters energy pathways by increasing primary production (see section on Energy flow and Sources above), which often results in lower dissolved oxygen concentrations due to excessive organic matter decomposition. These changes usually lead to highly productive, but taxonomically and trophically simple biological communities in both streams and wetlands (Reddy et al. 1999; Sandin and Johnson 2000; Brinson and Malvarez 2002). Herbicides also disrupt energy pathways, but they cause reductions in instream primary production, and pesticides directly affect survival and reproduction of populations. Excess turbidity caused by high levels of suspended sediment decreases oxygen levels and photosynthesis rates, impairs the respiration and feeding of aquatic organisms, destroys fish habitat, and kills benthic organisms (Johnston 1993b). In wetlands, high sedimentation rates decrease the germination of many wetland plant species by eliminating light penetration to seeds, lowering plant productivity by creating stressful conditions, and slowing decomposition rates by burying plant material (Jurik et al. 1994; Vargo et al. 1998; Wardrop and Brooks 1998; Mahaney et al. 2004).
In some instances wetland and riparian habitats can be effective mitigators of nonpoint source pollutants, especially nutrients and sediments, due to their ability to filter and transform contaminants, if their capacities are not exceeded. Because sediments and phosphorus are transported from uplands to streams and wetlands through surface flow, the primary removal mechanisms for phosphorus and metals are the settling of particles out of the water column and adsorption to organic matter and clay. Long-term removal can occur through roots, buried leaves, and sediment deposition (Richardson and Craft 1993). As long as there is sufficient time for transported material to come in contact with surface litter, riparian vegetation can be effective in retaining sediments and nutrients. For example, in a floodplain wetland in Sweden, 95% of phosphorus entering the wetland in surface runoff was removed within 16 m (Vought et al. 1994). In North Carolina, approximately 50% of the phosphorus leaving agricultural fields in runoff was removed in riparian areas (Cooper an. Gilliam 1987). However, during storms and in high-gradient watersheds, sediment retention by riparian zones is less effective (Jordon 1986). Phosphorus is even more sensitive to flow rates because it tends to bind to smaller particles that are less efficiently trapped by surface litter. In contrast, nitrogen moves primarily through ground water as dissolved nitrate, ammonia, or organic nitrogen (Peterjohn and Correll 1984), and thus, its removal is less tied to sediment dynamics than phosphorus.
Most nitrogen is removed from subsurface water through denitrification by soil microbes within wetlands and riparian soils (Davidsson and Stahl 2000). Research has shown that riparian forests are capable of retaining up to 89% as compared to 8% for cropland, and the nitrogen loss from the forest was primarily via groundwater (Peterjohn and Correll 1984; Gilliam 1994; Jordan et al. 1997). But as with sediments and phosphorus, retention of nitrogen is also more efficient at low discharge. During high discharge, relatively more water moves from upland and riparian areas to streams and lowland wetlands through surface flow vs. shallow groundwater flows. Thus, there is less time for vegetative uptake and microbial transformation of nutrients (Pionke et al. 2000). Research has shown a 90% or more reduction in NO3 − concentrations in water as it flows through riparian areas (Groffman et al. 1992; Gilliam 1994; Vidon et al. 2010). Labile organic matter is fundamental to denitrification as it provides the substrate necessary to drive this process for microbes to perform the process of denitrification. Plant uptake is an additional means of nitrogen removal from the system. Channelization and channel incision interferes with both of these processes by driving groundwater flowpaths deeper below the organic-rich alluvium (Phillips et al. 1993).
Sediment retention in wetlands and riparian corridors not only removes phosphorus, but has the additional function of reducing turbidity and contaminants sorbed to sediments, thus benefiting neighboring streams, rivers, and lakes (Oschwald 1972; Boto and Patrick 1978; Cooper an. Gilliam 1987; Hemond and Benoit 1988; Johnston 1991). While wetlands and floodplains have been shown to trap sediment in relatively unaltered settings, accelerated sedimentation can quickly overwhelm the capacity of these habitats to store and process sediments (Jurik et al. 1994; Wardrop and Brooks 1998; Freeland et al. 1999). High sedimentation rates decrease germination of many wetland and riparian plant species by eliminating light penetration to seeds, lower plant productivity by creating stressful conditions, and slow decomposition rates by burying plant material (Jurik et al. 1994; Vargo et al. 1998; Wardrop and Brooks 1998; Mahaney et al. 2004).
Landscape disturbances impact sediment loading and retention within the aquatic components of watersheds, and for the MAR, anthropogenic disturbances have been occurring for several centuries. Walter and Merritts (2008) reported on the sediments stored behind 10,000 s of mill dams in the MAR that as they become derelict are releasing “legacy” sediments to downstream areas. The influence of these sediment releases, mostly unpredictable, on aquatic ecosystems downstream is difficult to discern. In addition, former mill dams and beaver dams may, in fact, be the origins of some wetlands along headwater streams. Hupp et al. (1993) found sedimentation rates to be highest in wetlands located downstream from agricultural and urban areas. Phillips (1989) found that between 14 and 58% of eroded upland sediment is stored in alluvial wetlands and other aquatic environments, and as much as 90% of eroded agricultural soil was retained in a forested floodplain in North Carolina (Gilliam 1994). Eighty-eight percent of the sediment leaving agricultural fields over the last 20 years was retained in the watershed of a North Carolina swamp (Cooper et al. 1986). Approximately 80% of this was retained in riparian areas above the swamp and 22% was retained in the wetland itself. Carline and Walsh (2007) found that streambank fencing restricting livestock access to narrow riparian corridors (3–4 m) along pastures in a central Pennsylvania watershed reduced suspended solids by >50% resulting in an increase in abundance of benthic macroinvertebrates. These studies and others demonstrate the critical need to consider the aquatic and terrestrial mosaic of riverine ecosystems.
Another major threat to water quality in streams and wetlands of higher elevations is increased acidity associated with AMD and acid deposition (AD). As a region, the pH of rainfall in the MAR is among the lowest nationwide (NADP 2003), and, although long-term monitoring have shown wide-spread improvements in air quality and a reduction in acid deposition (Stoddard et al. 1999), aquatic biota in MAR streams and lakes have not shown evidence of recovery to AD (Stoddard et al. 1999). Low pH has a negative impact on the presence and breeding success of pond-breeding amphibians in wetlands, even more so when in combination with high concentrations of metals like aluminum and zinc (Clark 1985; Sadinski and Dunson 1992; Rowe and Dunson 1993; Horne and Dunson 1994). Snyder et al. (2005) found that over one quarter of wetlands in the Delaware Water Gap National Recreation Area along the Pennsylvania–New Jersey border had pH levels <5. In this study area, the lowest pH levels are among depression wetlands that lack permanent stream connections to adjacent water bodies (Julian 2009). This trend threatens amphibian species that breed early in the year because wetlands that lack permanent stream connections are used most frequently by early-breeding species (Julian 2009). Early-breeding species, like ambystomatid salamanders (Family Ambystomatidae), prefer these habitats because the drying regimes of these wetlands reduce aquatic predator populations. In DWG, ambystomatid salamanders bred in half of all wetlands whose maximum flooded areas were more than halved by the start of summer (Julian et al. 2006), yet Snyder et al. (2005) found they were excluded from these wetlands if the pH ≤4.6.
Increased acidity can have dramatic effects on stream and wetland communities, particularly in headwaters that are poorly buffered, chemically speaking. Increased H+ ions directly disrupt ion regulation in most animal species causing death or compromising fitness depending on the level (Gerhardt 1993). Certain metals such as aluminum, which are prevalent but relatively inert in streamside soils and stream sediments, become dissolved, mobilized, and toxic to aquatic species at low pH (Nelson and Campbell 1991). In addition, when acidic waters merge with pH neutral or basic waters at stream junctures, certain metal complexes such as iron hydroxide precipitate out of solution and coat stream substrates thus smothering benthic algae and macroinvertebrates (DeNicola and Stapleton 2002). Consequently, acid effects can extend downstream even in areas where stream pH is relatively high. Finally, leaf litter decomposition rates in headwater streams and wetlands are significantly reduced as streams become acidified (Kittle et al. 1995; Niyogi et al. 2001). The lower reaches of most rivers flow primarily through valleys of the various ecoregions in the MAR. In the Ridge and Valley ecoregion, calcium-rich limestone comprises the underlying bedrock, thus neutralizing the detrimental effects of acidification that arises in the highlands. In other ecoregions, the larger discharges of some rivers tend to override the influence of underlying geologic strata, or alluvial fill for coastal plains.
In the case of AMD, acidity and metal concentrations are frequently so high that the affected stream or wetland may be devoid of all life. In less extreme cases, AMD and AD has been shown to adversely affect the species diversity and productivity of benthic algae (e.g., Verb and Vis 2000), macroinvertebrates (e.g., Rosemund et al. 1992), amphibians (see Freda 1991 and Freda et al. 1991, for reviews mostly concerning wetland and vernal pool-breeding amphibians), and fish (Carline et al. 1992). After completing three separate investigations, which included field sampling as well as in situ and laboratory bioassays, reduced abundance and distribution of most lungless stream salamanders (Family Plethodontidae) was attributed to stream acidification (Rocco 2007).
The pH of water in streams receiving AMD or AD is often poorly correlated with the pH of the sources indicating that some systems are more vulnerable than others to acidification. As mentioned above, the water chemistry of headwater streams are more strongly related to the geology and terrain of the surrounding watershed than for larger rivers (Babb et al. 1997). One important characteristic of headwater stream water chemistry that is an important determinant of sensitivity to AMD and AD is the concentration of base cations (e.g., calcium and magnesium). Streams that have high base cation concentrations typically have high acid neutralizing capacities (ANC), and are, therefore, more able to maintain a stable pH despite AD (Faust 1983). In headwater streams, base cation concentrations are largely a function of the underlying surface geology. Streams underlain by carbonate geologies such as limestone supply considerable ANC to streams compared to geologies with little or no base cations like sandstone. However, the ability of carbonate geologies to buffer acidity associated with AD also depends on the amount of time that streams are exposed to AD. Specifically, in streams exposed to AD, the production of base cations through mineral weathering is slower than the rate they are leached into the stream. Thus, over time, the pool of available cations may become depleted causing a threshold effect whereby the ability of carbonate geologies to buffer AD is compromised (Kirchner 1992).
Wetlands offer some mitigation potential for acidified streams. For example, comparative research studies have shown that beaver ponds generate significant ANC to associated streams resulting in more stable pH (Cirmo et al. 2000; Margolis et al. 2001). Moreover, laboratory experiments using simulated wetlands have demonstrated that wetland soils act as sinks for strong acid anions (nitrates and sulfates), and wetland microbial communities transform toxic metals to less toxic or available forms (Tarutis et al. 1992; Williams et al. 1994). Constructed wetlands have been shown to be an effective mitigation tool for restoring streams affected by AMD by removing up to 99% of the iron and aluminum and up to 30% of the nitrogen loading (Brenner 2000). Although the ability of wetlands to ameliorate AMD was first observed in natural systems, constructed wetlands treatment systems are more likely to be used today, with design and size tailored to match the constituents of specific discharges.
1.3.5 Biological Integrity
The biological diversity of aquatic ecosystems in the MAR has been documented reasonably well. Some taxa pertinent to the region are particularly diverse, notably salamanders (Rocco et al. 2004), freshwater mussels, aquatic insects (Klemm et al. 2003), and breeding neotropical migrant songbirds (e.g., Stein et al. 2000; O’Connell et al. 2003; Tiner 2005). Various investigations have tallied the species and communities that are prevalent in the region (e.g., Majumdar et al. 1989; Croonquist and Brooks 1993; Brooks et al. 1998; Myers et al. 2000; Snyder et al. 2002; Ross et al. 2003) (see Chaps. 6, 7, 8, and 9 of this book for additional information about fauna).
The maintenance of a characteristic plant community is a fundamental property of ecosystems. It is a designated HGM function for wetlands that also relates to a variety of ecological functions in watersheds such as energy dissipation via roughness, stabilization of sediments and soils, detrital production and nutrient cycling, and biodiversity and habitat functions. The composition of vascular plant communities has long been used to characterize and classify wetlands (Cowardin et al. 1979; Tiner 1988; Mitsch and Gosselink 2000; Miller and Wardrop 2006). Plant community composition influences many ecosystem properties, such as primary productivity and nutrient cycling (Hobbie 1992). Plant species composition plays an important role in determining soil fertility through feedbacks attributable to the original potential for site productivity (Wedin and Tilman 1990; Hobbie 1992). Plant community composition also influences the habitat quality for invertebrate, vertebrate, and microbial communities in both wetlands and streams (Gregory et al. 1991; Andreas and Lichvar 1995; Norokorpi 1997; Ainslie et al. 1999).
Plant communities may be highly modified by human alterations that facilitate colonization by invasive and aggressive species. Invasive species change competitive interactions, which result in changes in species composition (Walker and Smith 1997; Woods 1997). A checklist, which includes provisions for invasive plants, has been developed to record any observed stressors on streams, wetlands, and riparian areas in the region (Brooks et al. 2006; Brooks et al. 2009). Streams and riparian systems are particularly vulnerable to exotics because their linear nature exposes them to invasions (Simberloff et al. 2005).
Land use can be considered a major driver of the characteristics and conditions of aquatic landscapes, and activities tied to changing land use can be the source of many stressors. Stream biological integrity is strongly correlated with the extent of agriculture, wetlands, and forests in the surrounding landscape (Roth et al. 1996; Snyder et al. 2003). Of particular importance to aquatic ecosystems are the patterns that arise along riparian corridors (Jordan et al. 1993; Castelle et al. 1994; Sweeney et al. 2004). In the MAR, stream reaches with wider forested riparian corridors, composed of uplands or wetlands, support higher abundance of macroinvertebrates, and process more carbon, nitrogen, and pesticides than narrower reaches. Because of these relationships, attributes of both landscape patterns and riparian corridors can be used to assess condition (King et al. 2005; Brooks et al. 2009).
When considering how various stressors influence aquatic landscapes, it is instructive to consider deviations from reference standard conditions that support the highest levels of biological integrity. In the eastern USA, the best attainable conditions for aquatic systems are usually derived from a landscape dominated by mature forests (or emergent marshes in the portions of some ecoregion such as the Coastal Plain), which produce characteristic inputs of organic matter, shade over wetlands and narrow stream corridors, and habitat for an expected set of species. In the floodplains of larger rivers, microtopographic heterogeneity arises from the interplay of hydrologic forces, vegetative structure, and underlying soil characteristics. The resultant mosaic of wet and dry patches found in natural floodplains and along the interfaces between aquatic and terrestrial systems support a diversity of biological communities adapted to wetting and drying cycles. These physical and biological complexities interact with and upon the materials present through biogeochemical processes to produce the ecological functions and services recognized from these systems.
Reference domains can exist for all major types of land use: forested, natural herbaceous, mixed, agricultural, and urban (Brooks et al. 2006). Most human-caused disturbances set back ecological succession to early stages. That is, for varying lengths of time, mature forests and large trees along riparian corridors will be absent, soil formation may be retarded, and the composition of floral and faunal communities will be different. Although natural processes also retard succession (e.g., severe floods, fire, disease, and insect epidemics), in the MAR these typically create a quilt-like mosaic of recovering habitat patches. As humans continue to transform the landscape of the MAR and elsewhere, forest cover is generally reduced, replaced by agricultural, suburban, and urban land uses linked through transportation and utility corridors, although in some areas forests are stable or increasing, and urbanization replaces agricultural lands (e.g., Brooks et al. 2011b). The spatial extent and pattern of these changes determine the degree of alteration and degradation observed in aquatic landscapes. Additionally, point sources of urban stormwater, agricultural runoff, and other pollutants can severely degrade these systems. Degrees of change can be detected through monitoring if selected attributes are used as indicators or vital signs. If the desired spatial mix and connectivity of natural habitats and human-influenced land uses can be determined, then land use policies and management practices can be focused on achieving those goals. The common thread to consider when planning land use policies and practices is to treat aquatic landscapes holistically rather than as a set of separate, disconnected components. This theme courses through the remaining chapters.
References
Ainslie WB, Smith RD, Pruitt BA, Roberts TH, Sparks EJ, West L, Godshalk GL, Miller MV (1999) A regional guidebook for assessing the functions of low gradient, riverine wetlands in western Kentucky. WRP-DE-17. Army Engineer Waterways Experiment Station, Vicksburg, MS
Andreas BK, Lichvar RW (1995) Floristic index for establishing assessment standards: a case study for northern Ohio. Wetlands Research Program Technical Report WRP-DE-8. U.S. Army Corps of Engineers Waterways Experiment Station, Vicksburg, MS
Arcement GJ Jr, Schneider VR (1989) Guide for selecting Manning’s roughness coefficients for natural channels and flood plains. U.S. Geological Survey Water-Supply Paper 2339, U.S. Government Printing Office, Washington, DC
Association of State Floodplain Managers (2008) Natural and beneficial floodplain functions: Floodplain management—more that flood loss reduction. Position paper, adopted by ASFPM Board 09/16/08, 8pp.
Babb JS, Cole CA, Brooks RP, Rose AW (1997) Hydrogeomorphology, watershed geology, and water quality of wetlands in central Pennsylvania. J PA Acad Sci 71(1):21–28
Barbour MT, Gerritsen J, Snyder BD, Stribling JB (1999) Rapid bioassessment protocols or use in streams and wadeable rivers: periphyton, benthic macroinvertebrates and fish, 2nd edn, EPA 841-B-99-002. U.S. Environmental Protection Agency, Office of Water, Washington, DC
Bason CW (2004) Effects of beaver impoundments on stream water quality and floodplain vegetation in the inner coastal plain of North Carolina. Master’s Thesis, East Carolina University, Greenville, NC
Bayley PB (1989) Aquatic environments in the Amazon basin, with an analysis of carbon sources, fish production, and yield. In: Dodge DP (ed) Proceeding of the international large rivers symposium, Canadian special publication in fisheries and aquatic sciences. NRC, Ottawa, ON, 106:399–408
Bayley PB (1995) Understanding large river-floodplain ecosystems. Bioscience 45:153–158
Bayley PB, Sparks RE (1989) The flood pulse concept in river-floodplain systems. In: Dodge DP (ed) Proceeding of the international large rivers symposium, Canadian special publication in fisheries and aquatic sciences. NRC, Ottawa, ON, 196:110–127
Benda L, Poff NL, Miller D, Dunne T, Reeves G, Pess G, Pollock M (2004) The Network Dynamics Hypothesis: how channel networks structure riverine habitats. BioScience 54:413–427
Benke AC, Chaubey I, Ward GM, Dunn EL (2000) Flood pulse dynamics of an unregulated river floodplain in the southeastern U.S. coastal plain. Ecology 81:2730–2741
Bilby RE, Heffner JT, Frasen BR, Ward JW (1999) Effects of immersion in water on deterioration of wood from five species of trees used for habitat enhancement projects. J N Am Fish Manage 19:687–695
Bilby RE, Likens GE (1979) Effect of hydrologic fluctuations on the transport of fine particulate organic carbon in a small stream. Limnol Oceanogr 24:69–75
Boling RH Jr, Goodman EE, Van Sickle JA, Zimmer OJ (1975) Toward a model of detritus processing in a woodland stream. Ecology 56:141–151
Boto KG, Patrick JWH (1978) Role of wetlands in removal of suspended sediments. In: Greeson PE, Clark JR, Clark JE (eds) Wetland functions and values: the state of our understanding. Proceedings of the National Symposium on Wetlands, American Water Resources Association, Minneapolis, MN, pp 479–489
Boward DM, Kazyak PF, Stranko SA, Hurd MK, Prochaska (1999) From the mountains to the sea: the state of Marylands’ freshwater streams. EPA 903-R-99-023. Maryland Department of Natural Resources, Monitoring and Nontidal Assessment Division, Annapolis, MD, 54pp.
Brenner JJ (2000) Use of constructed wetlands for acid mine drainage abatement and restoration. Water Sci Technol 44:11–12
Brinson MM, Lugo AE, Brown S (1981) Primary productivity, consumer activity, and decomposition in freshwater wetlands. Annu Rev Ecol Syst 12:123–161
Brinson MM (1990) Riverine forests. In: Lugo AE, Brinson MM, Brown S (eds) Forested wetlands: ecosystems of the world. Elsevier, Amsterdam, pp 87–141
Brinson, MM (1993a) A hydrogeomorphic classification for wetlands. Technical Report WRP-DE-4, U.S. Army Engineer Waterways Experiment Station, Vicksburg, MS
Brinson MM (1993b) Changes in the functioning of wetlands along environmental gradients. Wetlands 13:65–74
Brinson MM, Hauer FR, Lee LC, Nutter WL, Rheinhardt RD, Smith RD, Whigham D (1995) A guidebook for application of hydrogeomorphic assessments to riverine wetlands (Operational draft). Wetlands Research Program Technical Report WRP-DE-11, U.S. Army Engineer Waterways Experiment Station, Vicksburg, MS
Brinson MM, Malvarez AI (2002) Temperate freshwater wetlands: types, status, and threats. Environ Cons 29:15–133
Brooks RP, O’Connell TJ, Wardrop DH, Jackson LE (1998) Towards a regional index of biological integrity: the examples of forested riparian ecosystems. Environ Monit Assess 51:131–143
Brooks RP (ed) (2004) Monitoring and assessing pennsylvania wetlands. Final Report for Cooperative Agreement No. X-827157-01, between Penn State Cooperative Wetlands Center, Pennsylvania State University, University Park, PA and U.S. Environmental Protection Agency, Office of Wetlands, Oceans, and Watersheds, Washington, DC. Rep. No. 2004–3 Penn State Cooperative Wetlands Center, University Park, PA
Brooks RP, Wardrop DH, Bishop JA (2004) Assessing wetland condition on a watershed basis in the Mid-Atlantic region using synoptic land cover maps. Environ Monit Assess 94:9–22
Brooks RP, Snyder C, Brinson C (2006a) Structure and functioning of tributary watershed ecosystems in the Eastern Rivers and Mountains Network: Conceptual models and vital signs monitoring. Natural Resources Report NPS/NER/NRR-2006/009. National Park Service, Philadelphia, PA, 88pp.
Brooks RP, Wardrop DH, Thornton KW, Whigham D, Hershner C, Brinson MM, Shortle JS (eds) (2006b) Ecological and socioeconomic indicators of condition for estuaries and watersheds of the Atlantic Slope. Final Report to U.S. Environmental Protection Agency STAR Program, Agreement R-82868401, Washington, DC. Atlantic Slope Consortium, University Park, PA, 96pp. + attachments (CD)
Brooks RP, McKenney-Easterling M, Brinson M, Rheinhardt R, Havens K, O’Brien D, Bishop J, Rubbo J, Armstrong B, Hite J (2009) A Stream-Wetland-Riparian (SWR) Index for assessing condition of aquatic ecosystems in small watersheds along the Atlantic Slope of the Eastern U.S. Environ Monit Assess 150:101–117
Brooks RP, Brinson MM, Havens KJ, Hershner CS, Rheinhardt RD, Wardrop DH, Whigham DF, Jacobs AD, Rubbo JM (2011a) Proposed hydrogeomorphic classification for wetlands of the Mid-Atlantic Region, USA. Wetlands 31(2):207–219
Brooks RP, Yetter SE, Carline RF, Shortle JS, Bishop JA, Ingram H, Weller D, Boomer K, Stedman R, Armstrong A, Mielcarek K, Constantz G, Goslee S, Veith T, Piechnik D (2011b) Analysis of BMP implementation performance and maintenance in Spring Creek, an agriculturally-influenced watershed in Pennsylvania. Final report to U.S. Department of Agriculture, National Institutes of Food and Agriculture, Conservation Effects Assessment Project (CEAP), Washington, DC, 66pp.
Brookshire ENJ, Dwire KA (2003) Controls on patterns of coarse organic particle retention in headwater streams. J N Am Benthol Soc 22:17–34
Brown RG (1988) Effects of wetland channelization on runoff and loading. Wetlands 8:123–133
Brown SL (1990) Structure and dynamics of basin forested wetlands in North America. In: Lugo AE, Brinson MM, Brown S (eds) Forested Wetlands: Ecosystems of the World. Elsevier, Amsterdam, pp 171–199
Bryce SA, Larsen DP, Hughes RM, Kaufmann PR (1999) Assessing relative risks to aquatic ecosystems: a Mid-Appalachian case study. J Am Water Res Assoc 35:23–26
Bunn SE, Davies PM, Winning M (2003) Sources of organic carbon supporting the food web of an arid zone floodplain river. Freshw Biol 48:619–635
Carline RF, Sharpe WE, Gagen CJ (1992) Changes in fish communities and trout management response to acidification of streams in Pennsylvania. Fisheries 17:33–38
Carline RF, Walsh MC (2007) Responses to riparian restoration in the Spring Creek watershed, Central Pennsylvania. Restor Ecol 15:731–742
Carter V (1986) An overview of the hydrologic concerns related to wetlands in the United States. Can J Bot 64:364–374
Castelle AJ, Johnson AW, Conolly C (1994) Wetland and stream buffer size requirements—a review. J Environ Qual 23:878–882
Church M (2002) Geomorphic thresholds in riverine landscapes. Freshw Biol 47:541–557
Cirmo CP, Driscoll CT (1993) Beaver pond biogeochemistry: acid neutralizing capacity generation in a headwater wetland. Wetlands 13:277–292
Cirmo CP, Driscoll CT, Bowes K (2000) Chemical fluxes from sediments in two Adirondack wetlands: Effects of an acid-neutralization experiment. Soil Sci Soc Am J 64:790–799
Clark KL (1985) Responses of spotted salamander Ambystoma maculatum populations in central Ontario to habitat acidity. Can Field Nat 100:463–469
Cole CA, Brooks RP (2000) A comparison of the hydrologic characteristics of natural and created mainstem floodplain wetlands in Pennsylvania. Ecol Eng 14(3):221–231
Cole CA, Cirmo CP, Wardrop DH, Brooks RP, Peterson-Smith J (2008) Transferability of an HGM wetland classification scheme to a longitudinal gradient of the central Appalachian Mountains: initial hydrological results. Wetlands 28(2):439–449
Cooper JR, Gilliam JW, Jacobs TC (1986) Riparian areas as a control of nonpoint pollutants. In: Correll DL (ed) Watershed research perspectives. Smithsonian Institute Press, Washington, DC, pp 166–190
Cooper JR, Gilliam JW (1987) Phosphorus redistribution from cultivated fields into riparian areas. Soil Sci Soc Am J 51:1600–1604
Correll DL, Jordan TE, Weller DE (2000) Beaver pond biogeochemical effects in the Maryland Coastal Plain. Biogeochemistry 49:217–239
Cowardin LM, Carter V, Golet FC, LaRoe ET (1979) Classification of wetlands and deepwater habitats of the United States. FWS/OBS-79/31, U.S. Fish and Wildlife Service, Washington, DC
Croonquist MJ, Brooks RP (1993) Effects of habitat disturbance on bird communities in riparian corridors. J Soil Water Conser 48:65–70
Cuffney TF (1988) Input, movement and exchange of organic matter within a subtropical coastal blackwater river-floodplain system. Freshw Biol 19:305–320
Cummins KW, Petersen RC, Howard FO, Wuycheck JC, Holt VI (1973) The utilization of leaf letter by stream detritivores. Ecology 54:336–345
Cummins KW, Klug MJ (1979) Feeding ecology of stream invertebrates. Annu Rev Ecol Syst 10:147–172
Council PE (1973) Floodplain management: the prospects for Pennsylvania. Pennsylvania Environmental Council, Inc., Philadelphia, PA
Dahl TE (1990) Wetlands: Losses in the United States, 1780’s to 1980’s. U.S. Department of the Interior, Fish and Wildlife Service, FWS/OBS-79/31, Washington, DC
Davidsson TE, Stahl M (2000) The influence of organic carbon on nitrogen transformations in five wetland soils. Soil Sci Soc Am J 64:1129–1136
Delong MD, Thorp JH (2006) Significance of instream autotrophs in trophic dynamics of the Upper Mississippi river. Oecologia 147:76–85
DemissieM, Khan A (1993) Influence of wetlands on streamflow in Illinois. Contract Report 561, Illinois State Water Survey, Champaign, IL
DeNicola DM, Stapleton MG (2002) Impact of acid mine drainage on benthic communities in streams: the relative roles of substratum vs. aqueous effects. Environ Pollut 119:303–315
Dosskey MG, Bertsch PM (1994) Forest sources and pathways of organic matter transport to a blackwater stream: a hydrologic approach. Biogeochemistry 24:1–19
Euliss NH Jr, Mushet DM (1996) Water-level fluctuation in wetlands as a function of landscape condition in the prairie pothole region. Wetlands 16:587–593
Faust SD (1983) Total alkalinity versus buffer value (capacity) as a sensitivity indicator for fresh waters receiving acid rain. J Environ Sci Health 18:701–711
Forman RT (1995) Land mosaics: the ecology of landscapes and regions. Cambridge University Press, Cambridge, 632pp
Freda J (1991) The effects of aluminum and other metals on amphibians. Environ Pollut 71: 305–328
Freda J, Sadinski WJ, Dunson WA (1991) Long term monitoring of amphibian populations with respect to the effects of acidic deposition. Water Air Soil Pollut 55:445–462
Freeland JA, Richardson JL, Foss LA (1999) Soil indicators of agricultural impacts on northern prairie wetlands: Cottonwood Lake Research Area, North Dakota, USA. Wetlands 19:56–64
Frissell CA, Liss WJ, Warren CE, Hurley MD (1986) A hierarchical framework for stream habitat classification: viewing streams in a watershed context. Environ Manage 10:199–214
Georgian T, Wallace JB (1983) Seasonal production dynamics of a guild of periphyton-grazing insects in a southern Appalachian stream. Ecology 64:1236–1248
Gerhardt A (1993) Review of impact of heavy metals on stream invertebrates with special emphasis on acid conditions. Water Air Soil Pollut 66:289–314
Giberson DJ, Caissie D (1998) Stream habitat hydraulics: Interannual variability in three reaches of Catamaran Brook, New Brunswick. Can J Fish Aquat Sci 55:485–494
Gilliam JW (1994) Riparian wetlands and water quality. J Environ Qual 23:896–900
Gosselink JG, Turner RE (1978) The role of hydrology in freshwater wetland ecosystems. In: Good RE, Whigham DF, Simpson RL (eds) Freshwater wetlands ecological processes and management potential. Academic, New York, pp 63–78
Gregory SV, Swanson FJ, McKee WA, Cummins KW (1991) An ecosystem perspective of riparian zones. Bioscience 41:540–551
Groffman PM, Gold AJ, Simmons RC (1992) Nitrate dynamics in riparian forests: microbial studies. J Environ Qual 21:666–671
Guasch H, Ivorra N, Lehmann V, Paulsson M, Real M, Sabater S (1998) Community composition and sensitivity of periphyton to atrazine in waters: the role of environmental factors. J Appl Phycol 10:203–213
Harmon ME, Franklin JF, Swanson FJ, Sollins P, Lattin JD, Anderson NH, Gregory SV, Cline SP, Aumen NG, Sedell JR, Lienkaemper GW, Cromack K Jr, Cummins KW (1986) Ecology of coarse woody debris in temperate ecosystems. Adv Ecol Res 15:133–302
Hardison E, O’Driscoll M, DeLoatch J, Howard R, Brinson M (2009) Urban land use, channel incision, and water table decline along Coastal Plain streams, North Carolina. J Am Water Res Assoc 45:1032–1045
Hart DD, Robinson CT (1990) Resource limitation in a stream community: phosphorus enrichment effects on periphyton and grazers. Ecology 71:1494–1502
Hemond HF, Benoit J (1988) Cumulative impacts on water quality functions of wetlands. Environ Manage 12:639–653
Hewlett JD, Nutter WL (1970) The varying source area of streamflow from upland basins. In: Proceedings of the symposium on interdisciplinary aspects of watershed management. American Society of Civil Engineers, New York, NY, pp. 65–83
Hobbie SE (1992) Effects of plant species on nutrient cycling. Trends Ecol Evol 7:336–339
Hoeinghaus DJ, Winemiller KO, Agostinho AA (2007) Landscape-scale hydrologic characteristics differentiate patterns of carbon flow in large-river food webs. Ecosystems 10:1019–1033
Horne MT, Dunson WA (1994) Behavioral and physiological responses to terrestrial life stages of the Jefferson salamander, Ambystoma jeffersonianum, to low soil pH. Arch Environ Contam Toxicol 27:232–238
Hupp CR, Woodside MD, Yanosky TM (1993) Sediment and trace element trapping in a forested wetland, Chickahominy River, Virginia. Wetlands 13:95–104
Hychka K (2010) Characterizing hydrologic settings and hydrologic regimes of headwater riparian wetlands in the Ridge and Valley of Pennsylvania. Dissertation. Pennsylvania State University, University Park, PA
Iversen TM, Thorup J, Skriver J (1982) Inputs and transformation of allochthonous particulate organic matter in a headwater stream. Holarctic Ecol 5:10–19
Jacoby JM, Seeley MR, Welch EB, Horner RR (1990) Responses of periphyton to changes in current velocity, suspended sediment, and phosphorus concentration. Freshwater Biology 42:215–232
Johnston CA (1991) Sediment and nutrient retention by freshwater wetlands: effects on surface water quality. Crit Rev Environ Control 21:491–565
Johnston CA (1993a) Material fluxes across wetland ecotones in northern landscapes. Ecol Appl 3:424–440
Johnston CA (1993b) Mechanisms of wetland-water quality interaction. In: Moshiri GA (ed) Constructed wetlands for water quality improvement. Lewis, MI, pp 293–299
Johnston CA, Bubenzer GD, Lee GB, Madison FW, McHenry JR (1984) Nutrient trapping by sediment deposition in a seasonally flooded lakeside wetland. J Environ Qual 13:283–290
Jones JB, Mulholland PJ (2000) Streams and ground waters. Academic, San Diego, CA, 425pp
Jones JB Jr, Smock LA (1991) Transport and retention of particulate organic matter in two low-gradient headwater streams. J N Am Benthol Soc 10:115–126
Jordan TE, Correll DL, Weller DE (1993) Nutrient interception by a riparian forest receiving inputs from adjacent cropland. J Environ Qual 22:467–473
Jordan TE, Correll DL, Weller DE (1997) Relating nutrient discharges from watersheds to land use and streamflow variability. Water Resour Res 33(11):2579–2590
Julian JT (2009) Evaluating amphibian occurrence models and the importance of small, isolated wetlands in the Delaware Water Gap National Recreation Area. Ph.D. Dissertation, Ecology. Pennsylvania State University, University Park, PA, 119pp.
Julian JT, Snyder CD, Young JA (2006) The use of artificial impoundments by two amphibian species in the Delaware Water Gap National Recreation Area. Northeast Nat 13:459–468
Junk WJ, Welcomme RL (1990) Floodplains. In: Patten BC et al (eds) Wetlands and shallow continental water bodies. SPB Academic, The Hague, pp 491–524
Jurik TW, Wang SC, van der Valk AG (1994) Effects of sediment load on seedling emergence from wetland seed banks. Wetlands 14:159–165
Karr JR (1991) Biological integrity: a long-neglected aspect of water resource management. Ecol Appl 1:66–84
Karr JR (1999) Defining and measuring river health. Freshw Biol 41:221–234
Karr JR, Chu EW (1999) Restoring life in running waters: better biological monitoring. Island, Washington, D.C., 206pp
King RS, Baker ME, Whigham DF, Weller DE, Jordan TE, Kazyak PF, Hurd MK (2005) Spatial considerations for linking watershed land cover to ecological indicators in streams. Ecol Appl 15:137–153
Kirchner JW (1992) Heterogeneous geochemistry of catchment acidification. Geochim Cosmochim Acta 56:2311–2327
Kittle DL, McGraw JB, Garbutt K (1995) Plant litter decomposition in wetlands receiving acid mine drainage. J Environ Qual 24:301–306
Klemm DJ, Blocksom KA, Fulk FA, Herlihy AT, Hughes RM, Kaufmann PR, Peck DV, Stoddard JL, Thoeny WT, Griffith MB (2003) Development and evaluation of a macroinvertebrate biotic integrity index (MBII) for regionally assessing Mid-Atlantic Highlands Streams. Environ Manage 31(5):656–669
Lake PS (2000) Disturbance, patchiness, and diversity in streams. J N Am Benthol Soc 19:573–592
Mahaney WM, Wardrop DH, Brooks RP (2004) Impacts of sedimentation and nitrogen enrichment on wetland plant community development. Plant Ecol 175:227–243
Majumdar SK, Brooks RP, Brenner FJ, Tiner RW (1989) Wetlands ecology and conservation: emphasis in Pennsylvania. Pennsylvania Academy of Science, Easton, PA, 395pp
Margolis BE, Castro MS, Raesly RL (2001) The impact of beaver impoundments on the water chemistry of two Appalachian streams. Can J Fish Aquat Sci 58:2271–2283
May C, Horner R, Karr J, Mar B, Welch E (1997) Effect of urbanization on small streams in the Puget Sound Lowland Ecoregion. Water Protect Tech 2(4):483–494
McAllister LS, Peniston BE, Leibowitz SG, Abbruzzese B, Hyman JB (2000) A synoptic assessment for prioritizing wetland restoration efforts to optimize flood attenuation. Wetlands 20:70–83
Miller SJ, Wardrop DH (2006) Adapting the floristic quality assessment index to indicate anthropogenic disturbance in central Pennsylvania wetlands. Ecol Indica 6:313–326
Minshall GW, Cummins KW, Petersen RC, Cushing CE, Bruns DA, Sedell JR, Vannote RL (1985) Developments in stream ecosystem theory. Can J Fish Aquat Sci 42:1045–1055
Mitsch WJ, Gosselink JG (2000) Wetlands, 3rd edn. Wiley, New York, NY
Molloy JM (1992) Diatom communities along stream longitudinal gradients. Freshw Biol 28:59–69
Montgomery DR (1999) Process domains and the river continuum concept. J Am Water Resour Assoc 35:397–410
Mulholland PJ, Kuenzler EJ (1979) Organic carbon export from upland and forested wetland watersheds. Limnol Oceanogr 24:960–966
Myers W, Bishop J, Brooks R, O’Connell T, Argent D, Storm G, Stauffer J, Carline R (2000) Pennsylvania Gap Analysis Project. Leading landscapes for collaborative conservation. Final Report to U.S. Geological Survey, Biological Resources Division, Washington, DC, 142pp.+app
NADP (2003) National Acid Deposition Program 2003 annual summary. http://nadp.sws.uiuc.edu/lib/data/2003as.pdf
Naiman RJ, Decamps H, McClain ME (2005) Riparia. Ecology, conservation, and management of streamside communities. Elsevier Academic, Amsterdam, 430pp
Neatrour MA, Webster JR, Benfield EF (2004) The role of floods in particulate organic matter dynamics of a southern Appalachian river-floodplain ecosystem. J N Am Benthol Soc 23:198–213
Nelson WO, Campbell PGC (1991) The effects of acidification on the geochemistry of Al, Cd, Pb, and Hg in freshwater environments: A literature review. Environ Pollut 71:91–130
Newbold JD, Elwood JW, O’Neill RV, Van Winkle W (1982) Nutrient spiraling in streams: Implications for nutrient limitation and invertebrate activity. Am Nat 120:628–652
Niyogi DK, Lewis WM Jr, McKnight DM (2001) Litter breakdown in mountain streams affected by mine drainage: biotic mediation of abiotic controls. Ecol Appl 11:506–516
Norokorpi YEA (1997) Stand structure, dynamics, and diversity of virgin forests on northern peatlands. In: Trettin CC, Jurgensen MF, Grigal DF, Gale MR, Jeglum JK (eds) Northern forested wetlands: ecology and management. Lewis, New York, NY
Novitzki RP (1989) Wetland Hydrology. In: Majumdar SK, Brooks RP, Brenner FJ, Tiner JRW (eds) Wetlands ecology and conservation: emphasis in Pennsylvania. The Pennsylvania Academy of Science, Easton, PA, pp 47–64
O’Connell TJ, Brooks RP, Laubscher SE, Mulvihill RS, Master TL (2003) Using bioindicators to develop a calibrated index of regional ecological integrity for forested headwater ecosystems. Report No. 2003–01, Penn State Cooperative Wetlands Center, Final Report to U.S. Environmental Protection Agency, STAR Grants Program, Washington, DC, 87pp.+app
Oschwald WR (1972) Sediment-water interactions. J Environ Qual 1:360–366
Owen HJ, Wall GR (1989) Floodplain management handbook. Contract No. WR18745467, U.S. Water Resources Council, Flood Loss Reduction Associates, Washington DC
Paul MJ, Meyer JL (2001) Streams in the urban landscape. Ann Rev Ecol System 32:333–365
Peterjohn WT, Correll DL (1984) Nutrient dynamics in an agricultural watershed: observations on the role of a riparian forest. Ecology 65:1466–1475
Peterson RC, Cummins KW (1974) Leaf processing in a woodland stream. Freshw Biol 4:343–368
Phillips JD (1989) Fluvial sediment storage in wetlands. Water Res Bull 25:867–873
Phillips PJ, Denver JM, Shedlock RJ, Hamilton PA (1993) Effect of forested wetlands on nitrate concentrations in ground water and surface water on the Delmarva Peninsula. Wetlands 13:75–83
Pionke HB, Gburek WJ, Sharpley AN (2000) Critical source area controls on water quality in an agricultural watershed located in the Chesapeake Basin. Ecol Eng 14:325–335
Planas D, Moreau G (1986) Reaction of lotic periphyton to experimental acidification. Water Air Soil Pollut 30:681–686
Poff NL, Allan JD, Bain MB, Karr JR, Prestegaard KL, Richter BD, Sparks RE, Stromberg JC (1997) The natural flow regime: a paradigm for river conservation and restoration. Bioscience 47:769–784
Poole GC (2002) Fluvial landscape ecology: addressing uniqueness within the river discontinuum. Freshwater Biology 47:641–660
Praskievicz S, Heejun C (2009) A review of hydrological modeling of basin-scale climate change and urban development impacts. Prog Phys Geogr 33:650–671
Reddy KR, Connor GAO, Flaig E, Gale PM (1999) Phosphorus retention in streams and wetlands: a review. Crit Rev Environ Sci Technol 29:83–146
Reice SR (1984) The impact of disturbance frequency on the structure of a lotic riffle community. Verh Int Ver Theor Angew Limnol 22:1906–1910
Rheinhardt RD, Rheinhardt MC, Brinson MM, Faser KE Jr (1999) Application of reference data for assessing and restoring headwater ecosystems. Restor Ecol 7:241–251
Richardson CJ, Craft CB (1993) Effective phosphorus retention in wetlands: Fact or fiction? In: Moshiri GA (ed) Constructed wetlands for water quality improvement. Lewis, Ann Arbor, MI, pp 271–282
Rocco GL (2007) Responses of Plethodontid salamanders to stream acidification and acid mine drainage in the Pennsylvania Central Appalachians. Ph.D. Thesis. Pennsylvania State University, University Park, PA
Rocco GL, Brooks RP, Hite JT (2004) Stream plethodontid assemblage response (SPAR) index: development, application, and verification in the MAHA. Final Report. U.S. Environmental Protection Agency, STAR Grants Program, Washington, DC. Rep. No. 2004–01. Penn State Cooperative Wetlands Center, University Park, PA, 33pp.+figs+app.
Rosemund AD, Reice SR, Elwood JW, Mulholland PJ (1992) The effects of stream water acidity on benthic invertebrate communities in southeastern United States. Freshwater Biology 27:193–209
Rosgen DL (1994) A classification of natural rivers. Catena 22:169–199
Ross RM, Bennett RM, Snyder CD, Young JA, Smith DR, Lemarie DP (2003) Influence of eastern hemlock (Tsuga Canadensis L.) on fish community structure and function in headwater streams of the Delaware River basin. Ecol Freshw Fish 12:600–665
Roth NE, Allan JD, Erickson DL (1996) Landscape influence on stream biological integrity assessment at multi-spatial scales. Landsc Ecol 47:141–156
Rowe CL, Dunson WA (1993) Relationships among abiotic parameters and breeding effort by three amphibians in temporary wetlands of Central Pennsylvania. Wetlands 13:237–246
Sadinski WJ, Dunson WA (1992) A multilevel study of effects of low pH on amphibians of temporary ponds. J Herpetol 26:413–422
Sandin L, Johnson RK (2000) The statistical power of selected indicator metrics using macroinvertebrates for assessing acidification and eutrophication of running waters. Hydrobiologia 422:233–243
Schlesinger WH (1997) Biogeochemistry: an analysis of global change. Academic, New York, NY
Schlesinger WH, Melack JM (1981) Transport of organic carbon in the worlds rivers. Tellus 33:172–187
Schmidt SR, Breen DB, Parsons KC (2008) Assessing abundance of colonially-nesting waterbirds. Technical Report 1, Draft. Manomet Center for Conservation Sciences, Plymouth, MA
Scientific Assessment and Strategy Team (1994) Science for floodplain management into the 21st century. Part V, Report of the Interagency Floodplain Management Review Committee to the Administration Floodplain Management Task Force. Washington, DC
Scott MC, Helfman GS, McTammany ME, Benfield EF, Bolstad PV (2002) Multiscale influences on physical and chemical stream conditions across Blue Ridge landscapes. J Am Water Resour Assoc 38:1379–1392
Shure DJ, Gottschalk MR, Parsons KA (1986) Litter decomposition processes in a floodplain forest. Am Midl Nat 115:314–327
Simberloff D, Parker IM, Windle PN (2005) Introduced species policy, management, and future research needs. Front Ecol Environ 3(1):12–20
Smith RD, Ammann A, Bartoldus C, Brinson MM (1995) An approach for assessing wetland functions using hydrogeomorphic classification, reference wetlands, and functional indices. Technical Report WRP-DE-9. US Army Corps of Engineers, Waterways Experiment Station, Washington, DC
Smock LA (1990) Spatial and temporal variation in organic matter storage in low-gradient, headwater streams. Arch Hydrobiol 118:169–184
Snyder CD, Young JA, Lemarie DP, Smith DR (2002) Influences of eastern hemlock (Tsuga canadensis) forests on aquatic invertebrate assemblages in headwater streams. Can J Fish Aquat Sci 59:262–275
Snyder CD, Young JA, Villella R (2003) Influences of upland and riparian land use patterns on stream biotic integrity. Landsc Ecol 18:647–664
Snyder CD, Julian JT, Young JA, King TL (2005) Assessment of ambystomatid salamander populations and their breeding habitats in Delaware Water Gap National Recreation Area. Natural Resources Preservation Program Final Report submitted to the National Park Service, Delaware Water Gap National Recreation Area, Milford, PA, 51pp
Snyder CD, Young JA, Stout BM III (2006) Aquatic habitats of Canaan Valley, West Virginia: diversity and environmental threats. Northeast Nat 13:333–352
Sparks RE, Bayley PB, Kohler SL, Osborne LL (1990) Disturbance and recovery of large floodplain rivers. Environ Manage 14:699–709
Staubitz WW, Sobashinski JR (1983) Hydrology of Area 6: Eastern Coal Province, Maryland, West Virginia, and Pennsylvania. Open-file Report 83–33, U.S. Geological Survey, Water Resources Investigations, Towson, MD
Stein BA, Kutner LS, Adams JS (2000) Precious heritage. The status of biodiversity in the United States. Oxford University Press, New York, NY, 399pp
Stein ED, Mattson M, Fetscher AE, Halama KJ (2004) Influence of geologic setting on slope wetland hydrodynamics. Wetlands 24:244–260
Stoddard JL, Jeffries DS, Lukewille A, Clair TA, Dillon PJ, Driscoll CT, Forsius M, Johannessen M, Kahl JS, Kellogg JH, Kemp A, Mannio J, Monteith DT, Murdoch PS, Patrick S, Rebsdorf A, Skjelkvale BL, Stainton MP, Traaen T, van Dam H, Webster KE, Wietling J, Wilander A (1999) Regional trends in aquatic recovery from acidification in North America and Europe. Nature 401:575–578
Strahler AN (1952) Hypsometric (area-latitude) analysis of erosional topography. Bull Geol Soc Am 63:1117–1142
Strahler AN (1957) Quantitative analysis of watershed geomorphology. Am Geophys Union Trans 8:913–920
Swank WT, Swift JLW, Douglass JE (1988) Streamflow changes associated with forest cutting, species conversions, and natural disturbances. In: Swank WT, Crossley DA (eds) Forest hydrology and ecology at Coweeta. Springer, New York, NY, pp 297–312
Sweeney BW, Bott TL, Jackson JK, Kaplan LA, Newbold JD, Standley LJ, Hession WC, Horwitz RJ (2004) Riparian deforestation, stream narrowing and loss of stream ecosystem services. Proc Natl Acad Sci 101(39):14132–14137
Tarutis WJ Jr, Unz RF, Brooks RP (1992) Behavior of sedimentary Fe and Mn in a natural wetland receiving acidic mine drainage, Pennsylvania, U.S.A. Appl Geochem 7:77–85
Taylor JR, Cardamone MA, Mitsch WJ (1990) Bottomland hardwood forests: their functions and values. In: Gosselink JG, Lee LC, Muir TA (eds). Ecological processes and cumulative impacts: illustrated by bottomland hardwood wetland ecosystems. Chelsea: Lewis, pp. 13–86
Thorne DR (1998) Streamside reconnaissance handbook: Geomorphological investigation and analysis of river channels. Wiley, West Sussex
Thorp JH, Delong MD (2002) Dominance of autochthonous autotrophic carbon in food webs of heterotrophic rivers. Oikos 96:543–550
Thorp JH, Thoms MC, Delong MD (2006) The riverine ecosystem synthesis: biocomplexity in river networks across space and time. River Res Appl 22:123–147
Tiner RW (1988) Field guide to non-tidal wetland identification. Maryland Department of Natural Resources, Annapolis, MD, 283pp. + app.
Tiner RW (2003) Dichotomous keys and mapping codes for wetland landscape position, landform, water flow path, and waterbody type descriptors. National Wetlands Inventory Program, Northeast Region, U.S. Fish and Wildlife Service, Hadley, MA, 44pp
Tiner RW (2005) In search of swampland, 2nd edn. Rutgers University Press, New Brunswick, NJ, 330pp
Townsend CR (1989) The patch dynamics concept of stream community ecology. J N Am Benthol Soc 8:36–50
Townsend CR, Scarsbrook MR, Dolédec S (1997) The intermediate disturbance hypothesis, refugia, and biodiversity in streams. Limnol Oceanogr 42:938–949
U.S. Environmental Protection Agency (2000) Mid-Atlantic Highlands Streams Assessment. EPA/903/R-00/015. U.D. Environmental Protection Agency Region 3, Philadelphia, PA, 64pp.
Van Nieuwenhuyse EE, Jones JR (1996) Phosphorus-chlorophyll relationship in temperate streams and its variation with stream catchment area. Can J Fish Aquat Sci 53:99–105
Vannote RL, Minshall GW, Cummins KW, Sedell JR, Cushing CE (1980) The river continuum concept. Can J Fish Aquat Sci 37:130–137
Vargo SM, Neely RK, Kirkwood SM (1998) Emergent plant decomposition and sedimentation: response to sediments varying in texture, phosphorus content and frequency of deposition. Environ Exp Botany 40:43–58
Verb RG, Vis ML (2000) Comparison of benthic diatom assemblages from streams draining abandoned and reclaimed coal mines and nonimpacted sites. J N Am Benthol Soc 19:274–288
Vidon P, Allan C, Burns D, Duval TP, Gurwick N, Inamdar S, Lowrance R, Okay J, Scott D, Sebestyen S (2010) Hot spots and hot moments in riparian zones: potential for improved water quality management. J Am Water Resour Assoc 46(2):278–298. doi:10.1111/j.1752-1688.2010.00420.x
Vought LBM, Dahl J, Pedersen CL, Lacoursiere JO (1994) Nutrient retention in riparian ecotones. Ambio 23:342–348
Walker LR, Smith SD (1997) Community response to plant invasion. In: Luken JO, Thieret JW (eds) Assessment and management of plant invasions. Springer, New York, NY, pp 69–86
Wallace JB, Webster JR, Eggert SL, Meyer JL, Siler ER (2001) Large woody debris in a headwater stream: long-term legacies of forest disturbance. Int Rev Hydrobiol 86:501–513
Wallace JB, Webster JR, Meyer JL (1995) Influence of log additions on physical and biotic characteristics of a mountain stream. Can J Fish Aquat Sci 52:2120–2137
Walter R, Merritts D (2008) Natural streams and the legacy water-powered milling. Science 319(5861):229–304
Ward JV (1989) The four-dimensional nature of lotic ecosystems. J N Am Benthol Soc 8:2–8
Ward JV, Stanford JA (1983) The intermediate disturbance hypothesis: An explanation for biotic diversity patterns in lotic ecosystems. In: Fontaine TD, Bartell SM (eds) Dynamics of lotic ecosystems. Ann Arbor Science, Ann Arbor, MI, pp 347–356
Ward JV, Tockner K, Arscott DB, Claret C (2002) Riverine landscape diversity. Freshwater Biology 47:517–539
Wardrop DH, Brooks RP (1998) The occurrence and impact of sedimentation in central Pennsylvania wetlands. Environ Monit Assess 51:119–130
Webster JR, Benfield EF (1986) Vascular plant breakdown in freshwater ecosystems. Annu Rev Ecol Syst 17:567–594
Wedin DA, Tilman D (1990) Species effects on nitrogen cycling: a test with perennial grasses. Oecologia 84:433–441
Wiens JA (2002) Riverine landscapes: taking landscape ecology into the water. Freshw Biol 47:501–515
Williams DJ, Bland D, Williams MC, Patrzalek RW, Banks L, Logue B, Price MT (1994) Role of wetlands in water quality maintenance: microcosm investigation of factors influencing the buffering capacity of urban riparian wetlands. Lake Reserv Manage 9:124
Woods KD (1997) Community response to plant invasion. In: Luken JO, Thieret JW (eds) Assessment and management of plant invasions. Springer, New York, NY, pp 56–68
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer Science+Business Media New York
About this chapter
Cite this chapter
Brooks, R.P., Snyder, C., Brinson, M.M. (2013). Aquatic Landscapes: The Importance of Integrating Waters. In: Brooks, R., Wardrop, D. (eds) Mid-Atlantic Freshwater Wetlands: Advances in Wetlands Science, Management, Policy, and Practice. Springer, New York, NY. https://doi.org/10.1007/978-1-4614-5596-7_1
Download citation
DOI: https://doi.org/10.1007/978-1-4614-5596-7_1
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4614-5595-0
Online ISBN: 978-1-4614-5596-7
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)