
Abstract
The exploration, conservation, and use of agricultural biodiversity are essential components of efficient transdisciplinary research for a sustainable agriculture and food sector. Most recent advances on plant biotechnology and crop genomics must be complemented with a holistic management of plant genetic resources. Plant breeding programs aimed at improving agricultural productivity and food security can benefit from the systematic exploitation and conservation of genetic diversity to meet the demands of a growing population facing climate change. The genetic diversity of staple small grains, including rice, maize, wheat, millets, and more recently quinoa, have been surveyed to encourage utilization and prioritization of areas for germplasm conservation. Geographic information system technologies and spatial analysis are now being used as powerful tools to elucidate genetic and ecological patterns in the distribution of cultivated and wild species to establish coherent programs for the management of plant genetic resources for food and agriculture.
Graphical Abstract

Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
1 Introduction
Agricultural biodiversity is the key to successful biotechnological approaches for crop improvement. Plant breeding programs aimed at increasing crop productivity and improving food security rely on traits that must be efficiently managed and exploited for the sustainable delivery of cultivars without compromising genetic diversity. Exploration of new biodiversity in the face of climate change and the threat of genetic erosion must complement current trends in crop diversity. Enhanced conservation strategies of important agricultural biodiversity need to be deployed to take advantage of cutting-edge technologies on spacial analysis to monitor the patterns of plant diversity and distribution and prioritize areas for conservation. Most recent advances in the genetic modification of crop plants (i.e. transgenics) are also an important issue for agricultural production. Next-generation crops need to be created using transdiciplinary strategies, with the goal of making farming more productive and more profitable. Major staple crops, such as rice, maize, wheat, millets, and most recently quinoa, play an important role for millions of people worldwide because these crops sustain the lives of the poorest rural farmers. Therefore, diversity and conservation issues involving the utilization of these staple crops will significantly contribute to a household’s food security and nutrition of smallholders, mainly in the tropics.
2 Agricultural Biodiversity: Concept, Importance and Scope
Agricultural biodiversity, also known as agrobiodiversity, can be defined as all of the components of biological diversity that are relevant to food and agriculture, including agricultural ecosystems [1]. From a pragmatic perspective, agrobiodiversity is the result of the interaction between the environment, genetic resources, and management systems and the practices used by people from diverse cultural backgrounds.
Agrobiodiversity is an integral part of overall biodiversity; it comprises the variety and variability of animals, plants, and microorganisms at the genetic, species, and ecosystem levels that are used for food and agriculture, including crops, livestock, forestry, and fisheries. Culture and local knowledge are regarded as essential parts of agrobiodiversity because it is the human activity of agriculture that affects and shapes this biodiversity. In other words, agrobiodiversity is the result of natural selection and human intervention over millennia, and it plays a key role in sustainable development, including processes for, and in support of, food production and food security [2, 3]. Some part of this biodiversity is directly managed to supply the goods and services that people need; however, most of it is not directly intended for production purposes and remains important as a source of materials for its contributions to ecosystem services, such as pollination, control of greenhouse gas emissions, and soil dynamics [4].
The diversity in crops consists of the crops, landraces, and cultivars grown by farmers [5]. The world is currently facing serious environmental problems due to loss of biological diversity at alarming rates. Scientists have estimated that by 2025, 60,000 plant species could be lost. The Food and Agriculture Organization of the United Nations (FAO) has also stated that, since 1900, approximately three quarters of the genetic diversity of domestic agricultural crops has already been lost [6].
It is well known that modern, intensive agriculture reduces agricultural biodiversity [4, 5]; this loss of genetic diversity in agriculture is known as genetic erosion [7]. It is commonly assumed that plant breeding efforts are an important cause of genetic erosion of crops. However, the effects of urbanization and modern agricultural practices are important factors as well. Climate change and environmental degradation can also contribute to changes in cropping patterns and the disappearance of traditional varieties [5].
The spread and adoption of modern crop varieties has implications for genetic erosion and a decline in crop genetic diversity. Conservation of crop genetic resources is therefore a prerequisite for future generations. Organized and well-focused exploitation and conservation strategies of biodiversity will allow users to breed crop varieties for improved food security and face new challenges in the era of climate change.
2.1 Conservation of Agrobiodiversity in Small Grains: Status and Applications
It is generally accepted that the modernization of agriculture and land use changes negatively affect economic plant diversity, both on farmers’ fields and in home gardens [5]. This might eventually lead to the genetic erosion of cultivars and crop wild relatives with potentially useful traits for current and future human use. Therefore, many collecting missions have been organized in the past decades to establish extensive international and national genebank collections for important food crops, including cereals [8]. More than 3 million cereal accessions (i.e. samples of living plant material collected from particular locations) are stored ex situ worldwide [9]. They account for almost half of all genetic materials conserved globally in genebank collections [9]. This confirms the importance of cereals for global food security and agricultural production. The three crops with the most accessions conserved ex situ at a global level are the cereals rice (Oryza spp.), wheat (Triticum spp.), and barley (Hordeum vulgare subsp. vulgare) [9]. Of other cereal crops, such as maize (Zea mays L.), sorghum (Sorghum spp.), oat (Avena sativa L.), and millets (e.g. Pennisetum glaucum), less but still huge amounts of materials are being conserved ex situ [9]. To facilitate access to genetic material for evaluation, breeding, and direct use, some of these collections are put in the Multilateral System (MLS) under the conditions defined in the International Treaty on Plant Genetic Resources for Food and Agriculture (www.planttreaty.org). Several of the collections that are in the open domain, such as those held in trust by the Consultative Group on International Agricultural Research (CGIAR), can be consulted through the GENESYS Web portal (http://www.genesys-pgr.org), developed by Bioversity International.
The genetic integrity of accessions is maintained as much as possible in ex situ gene bank collections to conserve the specific characteristics of each material for evaluation, breeding, and direct use. For example, most accessions of the cereal collections held in trust by CGIAR have been characterized (88 %) [9]. This percentage is higher than all other types of crops conserved by CGIAR centers and The World Vegetable Center [9]. However, considering all reported national collections around the world, the amount of characterized cereal genetic resources is lower and similar to levels of genetic resource characterization of other crop types [9]. As a drawback, ex situ collections do not maintain the continued process of interactions between plants, humans, and environmental factors that take place on farms and between farmers [10]. In situ conservation of crops such as cereals is thought to be important to maintain the adaptive genetic variation of crop populations through interactions with their environment, including human management and selection. Several newly sampled barley materials in Morocco, for example, had more disease resistance than accessions collected several decades ago at the same location [11]. This supports the need of cereal genetic resources in situ conservation to allow evolution of adaptive genetic variation to important diseases for overall crop production. On the other hand, some historic ex situ accessions included rare genes to resistance that were extinct in the current species populations due to genetic erosion [11]. This highlights the importance of ex situ conservation as well as the necessity to develop complementary strategies of ex situ and in situ conservation.
Therefore, there is a need to assess the diversity status and dynamics of in situ plant genetic resources and develop complementary ex situ and in situ conservation strategies [9, 12–14]. At the same time, these types of analyses are useful to identify remaining gaps of diversity that are missing in existing genebank collections and that should therefore be targeted for germplasm collecting [9, 13]. Of course, periodic monitoring activities are required to measure the effectiveness of in situ conservation over time and to check the viability of seed material in ex situ collections.
The main purpose of in situ conservation is to maintain genetic variation in cultivated crop populations for phenotypic selection by farmers and/or natural processes [15]. This allows maintenance of the processes of microevolution and continuous adaptation of crop populations to their environments, including farmer management. The genetic structure of populations can change when phenotypic traits are heritable and selection is sufficiently strong. Following Darwin’s concepts of selection, this allows cumulative directional genetic response over generations—that is, microevolution of these populations to natural and human selection [15, 16]. Microevolution in plant populations is further driven by factors such as random mutation, recombination, and genetic drift [17, 18]. As an additional factor, many smallholders in all parts of the world periodically introduce new materials from neighbors and other localities into their systems to sustain productivity [19]. These factors and activities together make on-farm plant genetic resources management a dynamic system of crop genetic diversity use. Farmers may select for changing preferences as well choose to maintain desired phenotypic traits [15]. The variety of traits that is maintained and evolving under farmers’ care is often unknown to conventional breeders, entrepreneurs, and consumers. This makes on-farm conservation areas potential sources of untapped diversity for the development of new crop varieties for local and wider use. Even genetic diversity itself in cultivated populations may be a trait of farmers’ selection for ecosystem services, such as pest and disease control [20].
The status and trends of intraspecific crop diversity are traditionally being assessed and monitored through varietal diversity, either through farmer interviews or morphological or botanical classifications. For example, classification on the basis of traits that are important for farmers suggests that sorghum and pearl millet varietal diversity in Niger remained at similar levels during the last three decades of the twentieth century [21]. A reason for the maintenance of varietal diversity could be that the areas under study are marginal terrains where the cultivation of traditional crop and varieties outperforms cash crops and/or crop genetic diversity is being maintained as a risk management strategy [21]. Taxonomic keys have been used to monitor traditional maize varieties (www.biodiversidad.gob.mx/genes/proyectoMaices.html). Taxonomic studies are the basis for understanding variation in plant genetic resources. However, assessment of varietal diversity according to morphological or botanical characterization may still lead to a substantial degree of misidentification [22], and taxonomic keys may not exist for botanical varieties of specific crops or from particular geographic areas. This limits intraspecific diversity studies.
Molecular tools that identify polymorphisms have created novel opportunities for assessing crop genetic diversity, particularly when these markers are linked to adaptive traits and applied in combination with new geospatial methods of geographic and environmental analysis [23–25]. In addition, geographic information systems (GIS) can contribute significantly to improving the understanding and monitoring of spatial and temporal patterns of crop diversity [26]. Application of spatial analyses on georeferenced diversity data allows the formulation and implementation of better-targeted, and hence more effective, conservation strategies of inter and intra-specific plant diversity. For example, geospatial analyses combined with molecular marker characterization data have been used to support conservation strategies for African rice (O. glaberrima Steud.) genetic resources [27]. This study found the highest African rice genetic diversity in intermediate humid conditions and a decrease of genetic diversity under semiarid conditions and humid conditions [27]. This may have implications for the genetic resource conservation of this crop under changing climate conditions [27].
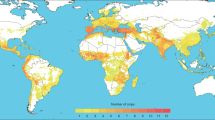
As an example of how to apply geospatial analysis to support plant genetic resources conservation, maize microsatellite diversity was mapped in the Americas. A dataset freely available by the Genetic Architecture of Maize and Teosinte project at www.panzea.org was used. It consists of molecularly characterized maize accessions from different genebanks. This dataset has been used by Vigouroux et al. [22] to understand the genetic structure (beta diversity) in America’s maize distribution. Four big genetic groups can be distinguished, consisting of material from (1) temperate zones in the United States and South America; (2) Mexican highlands; (3) tropical lowland; and (4) the Andes [22]. This data was used to map maize alpha diversity. A total of 1,145 georeferenced accessions that had microsatellite data for 92 markers were selected. Following van Zonneveld et al. [28], a 20 min grid layer (corresponding to approximately 33 km in the study area) was constructed for all genetic parameters, applying a circular neighborhood with a diameter of two degrees (corresponding to approximately 222 km) to improve visualization and group geographically isolated germplasm accessions. Calculations were done in the R program version 2.15.1 with the packages Raster [29] and Adegenet [30]. To standardize comparability of these parameters between cells, sample bias was corrected through resampling without replacement after Leberg [31] to a sample size of six trees. Per parameter, the average value was calculated from six subsamples following the bootstrap method developed by Thomas et al. [32].
The highest allelic richness is found in the central highlands of Mexico (Fig. 1). This indicates a high variation of maize genetic resources, and this area therefore should be prioritized for conservation actions. Nevertheless, the center of maize domestication is thought to be located in the southern lowlands, which thus is also an important area for conservation [33]. Also, a high number of alleles is found in these areas (Fig. 1). In a few scattered areas in northern Mexico, high levels of diversity are also observed.

Maize microsatellite diversity in the Americas constructed from geospatial analysis as a tool for genetic resources conservation. The average number of alleles per locus is shown
Allelic richness is lower in South America, where maize was introduced later. However, high levels of diversity can be found in Ecuador in the Andean region, Venezuela, northern Colombia, and Bolivia in the tropical lowlands (Fig. 1). These materials belong to different genetic groups than the material from the Mexican highlands [22]. These clusters may reflect different groups of evolution and adaptation to different environments. To maximize conservation of plant genetic resources, areas from these clusters that harbor high genetic variation should be prioritized for conservation.
The high genetic maize diversity in the Mexican highlands can be explained by the high levels of introgression between crop wild relatives and maize in that area [33, 34]. Indeed, gene flow between cultivated plants and their relatives in overlapping areas of distribution can cause elevated levels of intraspecific cultivated plant diversity. Such insights allow a better understanding of the role of evolutionary processes in the development of current species distributions and, where relevant, their domestication [35]. Similar phenomena have also been observed in other areas. Higher levels of molecular diversity of domesticated emmer wheat (Triticum turgidum L. subsp. dicoccon (Schrank) Thell.) and bread wheat (T. aestivum L.) have been found in the eastern Mediterranean and Turkey, respectively, and are located south and west of their putative centers of domestication due to crossing between domesticates and their wild ancestors [36].
In addition to studies across crop distribution ranges to target areas for in situ conservation and germplasm collecting, local spatial studies in prioritized traditional rural communities are important to provide input to the development of appropriate on-farm PGR management strategies [25]. These studies help to (1) increase the understanding of how farmers manage and conserve crop diversity within a community and; (2) identify the optimal geographic scale for interventions and crop diversity monitoring, as well as the social context in which in situ conservation should be implemented [37]. Estimates of distribution and levels of crop diversity in rural communities also help to determine the need to introduce new varieties into local seed systems and improve seed distribution systems accordingly [38].
Several case studies illustrate how genetic analysis helps to support in situ conservation interventions. A microsatellite marker study of diversity and structure of local rice varieties (Oryza sativa L., O. glaberrima Steud.) in the Republic of Guinea revealed genetic differences between two different agro-ecosystems, but no differentiation was shown between villages or farms within each of the contrasting agro-ecosystems [37]. This suggests that most genetic diversity can be conserved within just a few farms of a village [37]. Simple sequence repeat (SSR) markers also detected high seed exchange within villages of a traditional rice variety in Thailand [39]. In addition, high genetic differentiation between villages was found [39]. This indicates long periods of local adaptation and selection. Also, low genetic differentiation between seed lots of different farmers of the specific Mexican maize variety, Jala, suggests high seed exchange between farmers [40]. This implies that within communities only a few farmer fields would be necessary to target for in situ conservation and that collection for ex situ conservation of many individuals in a few farmer fields is preferred to collection of a limited number of individuals in many fields [40].
Climate change will certainly impact landraces conserved in situ, such as the native maize races in Mexico [18]. Nevertheless, it remains difficult to predict whether a local landrace goes extinct or can adapt [18]. This depends partly on the magnitude of the climate alterations, the novelty of new climates, and the amount of genetic variation present in landrace populations [18]. Seed exchange between farmers remains an important means to adapt local seed material to changing climates. Ecogeographic analysis of traditional maize systems in Mexico shows that mid-elevation communities can adapt fairly easy their production systems to climate change through seed exchange with farmers within a 10 km radius, where a wide range of different micro-climates can be found [41]. In contrast, highland and lowland systems that have less local micro-climate diversity require seed material from geographically more distant locations. The latter would require active support from governmental and non-governmental organizations [41].
During the domestication process of crops, overall genetic diversity is generally reduced, whereas phenotypic diversity at specific parts of the genome related to traits of interest are increased. However, many unknown traits of interest may have been lost during that domestication process. Crop wild relatives and progenitors may still include many potentially interesting traits, such as adaptive traits to heat stress and other climate change–related traits [42]. Therefore, recently more emphasis has been placed on the conservation of crop wild relatives. Microsatellite studies on rice in Vietnam confirm that wild rice populations contain much more diversity than cultivated populations [43]. A good example is mapping the genetic diversity of the wild subspecies of barley (Hordeum vulgare subsp. spontaneum) in Eastern Mediterranean and Middle East (Israel, Palestine, Syria, and Turkey), and Central Asia [44]. In these locations, barley here has higher genetic diversity in the eastern Mediterranean than elsewhere in its distribution. This area should therefore be a focus of conservation activities for barley genetic resources.
2.2 Genetic Diversity of Pearl Millet and Its Wild Relatives: Distribution, Conservation, and Use
2.2.1 Background
Pearl millet [Pennisetum glaucum (L.) R. Br.] is the fifth most important cereal crop in the world after rice, wheat, maize, and sorghum. It is a widely grown rainfed cereal crop in the arid and semi-arid regions of Africa and Southern Asia, and it can be grown in areas where rainfall is not sufficient (200 to 600 mm/year) for the cultivation of maize and sorghum. In other countries, it is grown under intensive cultivation as a forage crop. Pearl millet accounts for almost half of global millet production, with 60 % of the cultivation areas in Africa, followed by 35 % in Asian countries. European countries represent 4 % of millet cultivation and North America only 1 %, mainly for forage. Today, millet is a staple for more than 500 million people. Areas planted with pearl millet are estimated at 15 million ha annually in Africa and 14 million ha in Asia. Global production exceeds 10 million tons a year [45]. In sub-Saharan Africa, pearl millet is the third major crop, with the major producing countries being Nigeria, Niger, Burkina Faso, Chad, Mali, Mauritania, and Senegal in the West and Sudan and Uganda in the East. In Southern Africa, maize has partially or completely displaced millet cultivation because of commercial farming. India is the largest producer of pearl millet, both in terms of area (9.3 m ha) and production (9.3 mt), with an average productivity of 1044 kg/ha during the last 5 years (2007–2011).
The trends in area, production, and productivity of pearl millet suggest that area has increased marginally (2 %) during the last 2 years (2010–2011) and productivity has gone up by 19 % [46]. The major pearl millet growing states in India are Rajasthan, Maharashtra, Gujarat, Uttar Pradesh, Haryana, Karnataka, Madhya Pradesh, Tamil Nadu and Andhra Pradesh. However, productivity is the highest in Haryana, followed by Gujarat, Uttar Pradesh, Madhya Pradesh, Tamil Nadu, Andhra Pradesh, Rajasthan, Maharashtra, and Karnataka. It is mainly cultivated during the Kharif (rainy) season across the country. However, it is also grown to a lesser extent during the Rabi (post-rainy) season in Andhra Pradesh, producing high yields and excellent grain quality. Outside Africa and India, millets are also grown in Australia, China, Canada, Mexico, Russia, and the United States, primarily grown as a forage crop for livestock production [45]. Pearl millet is endowed with enormous genetic variability for various morphological traits, yield components, adaptation, and quality traits. Pearl millet is also nutritionally superior compared to maize and rice. The protein content of pearl millet is higher than maize and it has a relatively high vitamin A content.
2.2.2 Taxonomy, Origin, and Distribution
Pearl millet, Pennisetum glaucum, is an annual, allogamous, cross-pollinated, diploid cereal belonging to the Poaceae family, subfamily Panicoideae, tribe Paniceae, subtribe Panicinae, section Penicillaria, and genus Pennisetum. The genus Pennisetum contains about 140 species. The important wild relatives of cultivated pearl millet include the progenitor, Pennisetum glaucum subsp. monodii Maire, P. purpureum K. Schumach, P. pedicellatum Trin., P. orientale Rich, P. mezianum Leeke, and P. squamulatum Fresen. Previous names are P. typhoideum L.C. Rich andP. americanum (L.) Leeke. The four cultivated forms of pearl millet are typhoides (found mainly in India and Africa), nigritarum (dominant in eastern Sahel), globosum (dominant in the western Sahel, probably originating in Sahelian Africa in a diffuse belt stretching from western Sudan to Senegal), and leonis (dominant on the West African coast) [47–49].
Linnaeus [50] originally placed pearl millet cultigens into two separate species (P. glaucum and P. americanum) of the genus Panicum. Later, he moved several of these elements to the genus Holcus [51]. Rechard [52] grouped pearl millet along with a number of species previously listed under both Panicum L. and Cenchrus L. in a new genus, Pennisetum. Willdenow [53], however, established the genus Penicillaria to include pearl millet, but Steudel [54] reduced it to its present status as a section in Pennisetum. He merged many variants of pearl millet into a single polymorphic species, recognized as P. typhoideum L. Rich. The limits of the section were expanded by Leeke [55] to include all those wild species of Pennisetum having penicillate anther tips and involucral bristles. The generic name Pennisetum has been derived from two Latin words, Penna and Seta, meaning feather and bristles (i.e., feathery bristles). The most extensive treatment of the genus Pennisetum was contributed by Stapf and Hubbard [56], who divided the genus into five sections: Gymnothrix, Brevuvalvula, Penicillaria, Heterostachya, and Eu-pennisetum. Cultivated pearl millet and its wild and weedy relatives were included in section Penicillaria, which included 14 cultivated, 6 wild, and 13 intermediate species. Brunken [47] further reduced the number of species in section Penicillaria to two, on morphological and cytological grounds; P. purpureum was maintained as a separate species because of its tetraploid chromosome number and perennial lifecycle.
All the diploid cultivated, weedy, and wild taxa that frequently hybridize without genetic barriers are classified under a single species, P. americanum. Based on the morphology and adaptive strategies to domestication, P. americanum was further divided into three subspecies: americanum, including the cultivated forms; monodii, including the wild forms; and stenostachyum, with the weedy forms. Clayton and Renvoize [57] demonstrated that the taxonomically correct name for cultivated pearl millet is P. glaucum. They recognized the weedy forms (colloquially called shibra) as P. sieberanum and their wild progenitor as P. violaceum. P. violaceum differs from pearl millet in having involucres that are sessile, deciduous at maturity, and always contain a single spikelet.
The geographical origin and the center of domestication of pearl millet are situated in western Africa. The plant was subsequently introduced into India, where the earliest archaeological records date back to 2000 BC [48, 58–60]. Records exist for cultivation of pearl millet in the United States in the 1850s, and the crop was introduced into Brazil in the 1960s. The oldest findings of wild and domesticated pearl millet were recorded at about 3500 BC in Dhar Tichitt, a Saharan site in Mauritania [61]. Birimi in northern Ghana has laid claim to one of the earliest findings of domesticated pearl millet, dated at about 1459 BC [62, 63].
These archaeobotanical findings in the Sahara and Sahel confirm the hypothesis of original distribution and widespread utilization of wild and cultivated pearl millet across sub-Saharan Africa [61, 62]. However, there is dispute among scholars as to whether pearl millet has a single center of origin or more than one place of origin (the so-called non-centers), which would have resulted from domestication processes occurring independently in several regions. According to the latter hypothesis, the whole Sahel, from Mauritania to western Sudan, was originally covered with these non-centers [60, 64–67]. Whether domestication took place as multiple parallel processes in the non-centers in several places along the Sahelian distribution belt of the wild progenitors or at one specific place [60, 68], the ultimate center of origin of the wild progenitors, P. monodii and P. mollissimum, is most likely to be situated in the Sahara desert [62, 63, 69].
Based on the distribution of pearl millet throughout the continent, the uniform cradle of domestication is likely to be the regions of Mauritania, Senegal, and western Mali [48, 68, 70]. Today’s cultivated forms developed out of this domestication cradle [71]. Next, these first early-maturing forms of domesticated pearl millet were carried eastwards, facilitated by their efficient adaptation to arid conditions [62]. About 3,000 years BC, the first translocation carried the crop to eastern Africa [68, 71] and then to India, where 3000-year-old carbonized pearl millet was detected at a site on the eastern coast [63, 69].
Another diffusion took place in the region near Lake Chad, more precisely on the Nigerian side [72], where a secondary center of diversity developed at about 2010 BC [68, 73]. There, photoperiod-sensitive cultivars were selected, which adapted to the more humid conditions in the southern Sudanian zone [62, 71]. These late-maturing cultivars were transported further towards the Sudanian zone of southwestern Africa, from northern Nigeria to southern Senegal, as evidenced by the above-mentioned findings in northern Ghana [68, 73]. The third and last major translocation took pearl millet towards the plateaus of southern Africa, across Uganda and towards Namibia, at about 1000 BC [68, 71].
2.2.3 Overview of Pearl Millet Collections
The International Crops Research Institute for the Semi-Arid Tropics (ICRISAT) has the single largest consolidated collection of pearl millet in the world, which comprises a total of 22,211 accessions. Of these, 750 are of wild species (24 species), 19,377 are landraces, 132 are improved cultivars, 1,943 are breeding/research materials, and 25 are others [74]. India contributed a significant number of pearl millet accessions to the global collection maintained at ICRISAT (6,647 accessions). The remaining accessions were collected from about 51 countries. The major diversity centers of pearl millet are considered to be relatively well represented in the collection at ICRISAT. In addition to ICRISAT, the Institut de Recherche pour le Développement in France also maintains 3,968 accessions of pearl millet from 16 countries. Collection of these accessions was supported by Bioversity International and Office de la Recherche Scientifique et Technique d’Outre-Mer. The Canadian Genetic Resources Programme in Saskatoon, Canada maintains 3,821 accessions covering a few species, with emphasis on Pennisetum glaucum (3390 accessions). Accessions of other species include: P. violaceum (221), P. macrourum (1), P. purpureum (12), P. orientale (1), P. pedicellatum (11), P. polystachion (8), P. ramosum (3), P. unisetum (1), and other species (14). The number of seeds maintained in these collections is moderate for long-term conservation; it is intended as safety duplication but not for distribution. In addition to these global collections, the Agricultural Research Station of the U.S. Department of Agriculture (USDA) at Griffin, Georgia maintains 1,314 accessions from 32 countries, of which only 1 is a wild relative, 290 are of breeding/research material, and 552 are for other purposes.
Among the national collections, the largest was recorded from the Indian genebank at the National Bureau of Plant Genetic Resources (NBPGR), which maintains 8,913 accessions under long-term conservation. Most of the accessions are indigenous (8,827), with only 168 accessions from other countries. The Indian collection also includes 221 advanced improved varieties and 272 accessions of breeding/research material. No other countries in South Asia, except Pakistan (193 accessions), have reported pearl millet collections. Among African countries, the collections were reported from gene banks based in Algeria, Benin, Botswana, Burkina Faso, Burundi, Cameroon, Central African Republic, Chad, Eritrea, Ethiopia, Ghana, Kenya, Malawi, Mali, Mauritania, Mozambique, Namibia, Niger, Nigeria, Senegal, Sierra Leone, Sudan, Democratic Republic of the Congo, Uganda, Zambia, and Zimbabwe. In total, 56,580 pearl millet accessions were recorded from various sources. Landraces represent the largest proportion of pearl millet germplasm conserved in gene banks worldwide (49,973 accessions). Of these, only 3 % are wild relatives collections (1,630), 0.80 % are advanced improved varieties (452), 6 % are breeding/research materials (3,600), and 2 % are of unknown description (947).
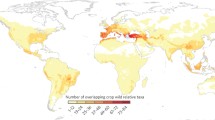
Some progress has been made in the recent past in mapping the pearl millet diversity collected worldwide. Global databases show that georeference data have been assigned to 16,855 accessions. These collections are being maintained at ICRISAT (13542 accessions), USDA Agricultural Research Service (ARS) (472 accessions) and the International Livestock Research Institute (13 accessions). With support from Bioversity International, 2,828 accessions were collected and became part of the global collections being maintained by ICRISAT and USDA-ARS. Not much information is available for any of the national collections. Based on the georeference information from the global database, the distribution pattern of these accessions is shown in Fig. 2, whereas the mapping of Pennisetum species (excluding Pennisetum glaucum) is shown in Fig. 3.

Mapping of pearl millet accessions based on information available from the global database

Mapping of Pennisetum species (excluding Pennisetum glaucum) accession collecting sites based on information obtained from the global accession level information portal GENESYS, developed by Bioversity International (http://www.genesys-pgr.org/)
2.2.4 Characterization and Evaluation
From a global survey undertaken by Bioversity International, it appears that although a modest number of accessions have been assembled and maintained in many countries, systematic characterization and evaluation activities are not sufficient. One reason provided by most gene banks is lack of adequate human and financial resources. Evaluation activities, especially in Africa, have been fewer than hoped. In the ICRISAT gene bank, all cultivated accessions have been characterized and evaluated for 23 morphoagronomic characters following the descriptors for pearl millet [75]. Selected pearl millet germplasm accessions of Indian and African origin were evaluated by NBPGR for important agronomic characters at different locations in India in collaboration with ICRISAT and catalogues were published [76, 77]. Considerable phenotypic diversity was observed for almost all quantitative traits. Distribution of qualitative traits indicates occurrence of nine panicle shapes (cylindrical, conical, spindle, club, candle, dumb bell, lanceolate, oblanceolate, and globose), five seed shapes (obovate, oblanceolate, elliptical,hexagonal, globular) and ten seed colors (ivory, cream, yellow, grey, dark grey, grey-brown, brown, purple, purplish-black, and mixture of white and grey) in the entire collection. Accessions with candle-shaped panicles, short bristled panicles, globular seed shape, grey seed color, and seeds with partly corneous endosperm texture are predominant in the collection maintained at ICRISAT [74].
Based on the characterization and evaluation information contained in the database, sources of resistance to biotic and abiotic stresses, adaptation, and nutritional qualitative traits have been identified. It was also reported that some of the landraces have wide adaptation and are therefore very useful in light of the changing climate scenario and for use in crop improvement programs. Much of this diversity is still available in areas of early cultivation in Africa and regions of early introduction in Asia.
2.2.5 Utilization
Identification of useful germplasm for crop improvement is the first step in encouraging utilization. From the information obtained through the Bioversity International survey questionnaire, it was difficult to obtain a good comparison of utilization activities being carried out in various genebanks, especially in Africa. However, in India, modest efforts have been undertaken in the last three decades to exploit pearl millet germplasm with useful genes for crop improvement, especially in developing composites. The iniadi germplasm from the Togo-Ghana-Burkina Faso-Benin region of Western Africa is most commonly used in pearl millet breeding programs worldwide [78]. At ICRISAT, a small seed sample of each accession is available on request to all research workers under the Standard Material Transfer Agreement of the International Treaty on Plant Genetic Resources for Food and Agriculture [74]. To further enhance the utilization of pearl millet germplasm, ICRISAT evaluated sets of selected germplasm accessions at different locations in India and several other countries in Africa; trait-specific genepools (early maturing, high tillering, large panicle, and large grain) were developed to provide partially conserved genotypes to the breeders. There has been a general lack of interest in using wild species because of the large genetic variability in pearl millet landraces. However, some wild species are very useful in pearl millet improvement programs, notably P. glaucum subsp. monodii for new source of cytoplasmic-nuclear male sterility (CMS); P. purpureum for forage, stiff stalk, and restorer genes of the A1 CMS system; P. orientale for drought tolerance and forage; P. schweinfurthii for large seed; P. pedicellatum and P. polystachion for downy mildew resistance, and P. squamulatum for apomictic gene [48].
In general, the Indian pearl millet landraces have contributed to earliness, high tillering, high harvest index, and local adaptation, whereas African materials have been a good source of high head volume, large seed size, and disease resistance. In order to enhance the use of these genetic resources, Bioversity International, in consultation with various partners, has developed a comprehensive list of descriptors for pearl millet [79]. This strategic set of descriptors, together with passport data, are an integral part of the information available through the global accession-level information portal GENESYS. It will facilitate access to and utilization of pearl millet accessions held in gene banks worldwide.
2.2.6 Distribution of Pennisetum Species and Gaps in World Collection
A study of the distribution of Pennisetum species and of gaps in world collections was undertaken by Bioversity International with support from the Global Crop Diversity Trust (GCDT) and the World Bank, using datasets of herbarium collections, as well as the germplasm collections available from Global Biodiversity Information Facility and the System-wide Information Network for Genetic Resources and the climate database available at WorldClim. Based on the available records, 53 wild species and 2 infraspecific taxa have been identified, accounting for a total of 55 taxa for the genus Pennisetum. These different taxa are classified as follows, according to their closeness to the cropped species P. glaucum, using the Maxted and Kell [80] model. The analysis dataset contained 4,326 observations (http://gisweb.ciat.cgiar.org/GapAnalysis/) with 3,364 (78 %) being herbarium specimens and 962 (22 %) being gene bank accessions. The average number of total samples per taxon was 79, indicating that available data is not particularly limited, although it is concentrated in certain taxa (i.e. P. ciliare, P. polystachion, P. purpureum, P. violaceum, P. clandestinum, P. villosum). Other taxa such as P. domingense, P. lanatum, and P. sieberianum present a very limited sampling and/or data availability and thus need further characterization and sampling in order to obtain a reliable ecogeographic evaluation.
The gap analysis of the Pennisetum genepool showed that there are 47 out of 55 taxa under analysis that are either underrepresented or not represented in gene banks; these were therefore flagged as high-priority species. Twenty-six of these taxa presented only 10 data points, which indicates that these species in particular need to be further collected. Only species P. violaceum was found to be adequately represented in gene banks, while P. ciliare, P. flaccidum, P. orientale, and P. pedicellatum were found to be relatively underrepresented and thus flagged as medium-priority species. Based on the analysis, all these species have been identified as high priority for conservation.
2.3 Quinoa Diversity and Its Potential in the New Millennium
Quinoa (Chenopodium quinoa Willd.) is an ancient native Andean grain that was extensively cultivated in the Andean region by pre-Columbian cultures, such as the Tiahuanacota and Inca, from around 5,000 years ago. This grain was used in the diet of the settlers from the inter-Andean valleys and the high plateaus along with other native species such as potato (Solanum tuberosum L.), oca (Oxalis tuberosa Molina), amaranth (Amaranthus caudatus L.), chili peppers (Capsicum sp.), among others [81, 82]. Quinoa is an herbaceous annual Chenopod that has played not only a vital role in family food security and farmer livelihoods, but also in economic, social, ecological, nutritional, and cultural contexts [83, 84]. Quinoa plants are part of various Andean ecosystems; the grains and leaves of this crop are used for food while its subproducts are used for forage, fuel wood, as well as in rituals and handicrafts [85].
This crop, however, began to be marginalized in the sixteenth century with the introduction of cereals such as barley and wheat [81, 86], leading to a significant decrease of quinoa-cultivated areas in the Andean countries [86]. Nevertheless, quinoa cultivation continued in marginal areas. For many decades, quinoa has been considered as a food for the poor and the peasants, but now it is considered to be today’s “golden grain”. In fact, quinoa was catalogued by FAO in 1996 as “one of the most promissory crops for humanity not only due to its high nutritional value and versatility, but also because it offers alternatives to solve the increasingly serious problems of human nutrition” [86].
Quinoa′s key asset lies in its potential as a high-quality source food. Quinoa has high concentrations of some essential amino acids (lysine, methionine, threonine, and tryptophan) that usually limit the quality of human diet. Quinoa is also rich in oligo elements, vitamins, and is gluten free, just to mention some of its nutritional properties [87]. The high content of these amino acids plus the high-quality protein found in its grain can cover the nutritional requirements for schoolchildren and even adults [88], which are serious problems in some populations around the world.
The Andean region is one of eight centers of origin and diversity of cultivated plants in the world described by Vavilov in 1953; that is, it is one of the regions that has the highest diversity of cultivated crops and their wild relatives. In this region and specifically around Lake Titicaca, a high plateau region in Bolivia and Peru, the highest genetic diversity of wild and cultivated quinoa genotypes are found in farmer fields [81, 89, 90].
Agriculture in the Andean highlands is characterized by a high degree of risk due to a range of harsh climatic factors such as frost, hail, wind, drought, high radiation, and poor and saline soils [91]. It is highly probable that during domestication, Andean farmers selected certain genotypes based on their use and their tolerance to adverse biotic and abiotic factors, thereby obtaining current plants and ecotypes that possess high diversity of traits. Some of these ecotypes have been strictly selected as a source of food (finding up to hundred different recipes), while others have been selected based upon their tolerance to salinity, poor soils, cold climate, frost and hail, drought, and flooding. It is important to remark that some genotypes have also been selected due to their high yield potential and precocity [89, 92, 93].
During the domestication of quinoa by Andean farmers, a wide range of morphological modifications have occurred to the plant, such as the condensation of the inflorescence in the highest part of the plant, an increase in plant and seed size, reduction of the testa, loss of seed dormancy and dispersion mechanisms, and high variation in levels of pigmentation, among others. Today, we can find quinoa plants with high production of larger seeds and clear colors; these traits reflect the long time man has been using, selecting, and cultivating this species [93]. Although this species has been completely domesticated, the seeds still contain saponin, which must be extracted before consumption [86]. Quinoa ancestors and relatives still exhibit the wild traits of the crop [93].
The genetic variability of quinoa is huge [91]. For example, one of the most popular varieties grown today in Bolivia (Quinua real) has at least 73 ecotypes. This variety is extremely well adapted to saline environments and has big grains. In addition, 47 local landraces and approximately 10 improved varieties are also found (Table 1).
In other quinoa-producing countries within the Andean region, a considerable diversity of local and improved varieties can also be found [86, 92]. The existence of this vast amount of ecotypes, improved and local varieties, and/or landraces are the result of the intense breeding processes, domestication, and use and conservation practices adopted by ancient and contemporary Andean farmer communities based on their traditional knowledge [86].
Currently, the demand for quinoa in the national and international markets has increased; quinoa production areas have also shown this tendency due to high market prices. However, this demand has preference for larger seed size, uniform color and white color, and saponin-free seeds [93, 94]. This has progressively caused the domination of a group of cultivated varieties with these traits, such as various ecotypes of Quinua real [89, 94]. Although markets have preferred these varieties, Bonifacio et al. [94] reported that, in the last couple of years, those with red and brown seeds have also been accepted now in some markets, although not to the same extent. Nevertheless, this market preference can be risky as it can further lead to genetic erosion and the loss of some varieties. Unfortunately, this process is in progress, and it has been stated that three varieties of quinoa and a wild relative have already been included in the International Union for Conservation of Nature red list categories as endangered (EN), near threatened (NT) and least concern (LC) [95].
Apaza et al. [96] reported that the highest diversity of quinoa is found in aynokas, mandas, and laymes, a group of traditional organization systems or communal fields maintained by farmer communities in field borders and sacred places (all called Gentil wasi or Phiru) using local cultivation methods and traditional knowledge. If these places continue being eroded and the traditional practices and knowledge become lost by the adoption of modern culture practices, it is highly probable that the diversity present in these sites will disappear.
To conserve the enormous genetic diversity of quinoa and some of its wild relatives, various germplasm banks were created in the 1960s throughout the South American region. These banks are being managed by public or private entities interested in the ex situ conservation of plant genetic resources, thereby finding more facilities in countries with higher diversity (Table 2). Collections of quinoa germplasm began in this period with the help and sponsorship of various private and public organizations, donors, and projects that still subsist today.
Although these banks keep an important number of quinoa accessions, the variability stored in these collections do not represent all the Andean diversity [86], especially when it comes to wild populations and wild relatives [97]. There are at least ten wild species or wild relatives and a couple of subspecies of the Chenopodium genus, known as wild quinoa or ajara, that are used for certain recipes by Andean farmers. These wild species are not being properly conserved.
Since 2001, some quinoa varieties and wild relatives, such as cañahua (Chenopodium pallidicaule Aellen), have been evaluated for yield, harvest index, postharvest, market, industrialization potential, new products and uses, among other fields, within the framework of the International Fund for Agricultural Development Neglected and Underutilized Species–Bioversity International Project 2001–2014. Additionally, other wild relatives are being evaluated for their potential to improve the tolerance to biotic and abiotic stresses as part of a United Nations Environment Programme/Global Environment Facility project for nutritive properties [90].
To date, results show that there is a huge potential in the conservation and use of quinoa diversity and its wild relatives. Studies also show that there is considerable variation among cultivars for a wide range of traits. This fact allows users to take advantage of quinoa’s versatility for use in more than 100 different preparations and products. Other than its cosmetic and industrial and pharmaceutical uses, quinoa can also be used as forage, medicine, pesticide, ornament, and fuel wood [91, 93, 98]. In addition, quinoa has a promising market potential for high-value grain, subproducts, and byproducts that still remain unexploited [99].
Both quinoa and its wild relatives possess an enormous reservoir of genetic variation that can be exploited through plant breeding. They are also an essential resource to meet the challenge to improve food security, enhance agricultural production, and sustain productivity in the context of a growing world population and the threats of climate change [90, 100]. Likewise, quinoa diversity can be greatly exploited through biotechnological approaches to the benefit of agricultural industries [98].
2.4 A Practical Use of Agrobiodiversity in Cereal Crops to Mitigate Climate Change through Regulation of Soil Nitrification
It has been shown repeatedly how the exploration of the natural diversity in cereals -and in crops in general- has been the solution for multiple challenges that naturally arise as a result of agricultural practices (tolerance to biotic and abiotic stresses, yield increments, adaptation to different conditions other than the ones in the center of origin, etc.). Along with the need of yield increments in the main staples cereals -as the global population is exponentially growing-, climate change is a prominent issue that is challenging agriculture and even mankind. Agriculture is an important source of anthropogenic emissions of the greenhouse gases (GHG) methane (CH4) and nitrous oxide (N2O) associated with nitrogen (N) fertilizer production [101]. It is now known that N2O has a higher ozone-depletion potential than any other reactive chemical, including carbon dioxide [102], and also that agricultural practices are the major source of N2O emissions to the atmosphere [103]. Under this scenario, a disjunctive stands, either to continue practicing agriculture for food and feed in the way we are currently doing or to look into alternatives to implement a “climate-smart” agriculture, where we as humans not only aim to feed the world but also care for the environment. Once again, researchers have found that natural diversity in certain crops confer characteristics to some lines that are “eco-friendly” and that, for example, reduce the emission of GHG as CH4 and N2O [104].
Much of the N2O produced by agriculture is generated from the use of N fertilizers that, after application to soil, feed the nitrification reaction; in this process, nitrifying microorganisms take ammonium and convert it into nitrite and finally into nitrate, a compound susceptible to leaching, thereby contaminating bodies of water. Nitrate is eventually reduced by denitrification, releasing N2O gas as a byproduct [105]. Researchers from the International Center for Tropical Agriculture and the Japan International Research Center for Agricultural Sciences observed that the tropical pasture Brachiaria spp. has the ability to inhibit the soil nitrification process by releasing chemical compounds from its root system to the soil; a compound with major inhibition capacity was identified and named brachialactone [106]. This phenomenon was termed biological nitrification inhibition (BNI) [107]. The BNI trait would decrease costs for the farmer because the N applied will stay longer in the soil and the plants would have a better chance to intake it before it is converted into forms that are prone to leaching and losses via gaseous forms, thereby improving the ecoefficiency of agricultural practices by diminishing the amount of N2O emissions that are released to the atmosphere. Subbarao et al. [106] demonstrated under field conditions that plots of Brachiaria pastures significantly reduced N2O emissions when compared to bare soil, soybean, and guinea grass plots.
After the BNI concept was brought to light, researchers identified the great potential that this phenomenon has in agricultural practices by reducing the N contaminants (i.e., nitrate and N2O); therefore, this strategy can be applied to mitigate global warming largely occasioned by GHG. For that reason, Subbarao et al. [108] achieved and attempt to identify in which other plant species the BNI potential was presented, as well as to what extent there was genetic variability of the BNI potential in cereal crops. The outcome of this investigation showed that BNI is widely existent in pasture grasses, but the investigation also reported that the roots of two cereal crops Sorghum bicolor (L.) Moench var. hybrid sorgo and Pennisetum glaucum (L.) R. Br. var. CIVT (Pearl millet) release chemicals to the soil that reduce the population of nitrifier microorganism and therefore inhibit the nitrification rates. Other cereals, such as rice, maize, barley, and wheat, were also tested in this preliminary screening with negative results (no BNI activity detected), but in most cases using single lines; therefore, it is recommended to test a diversity panel for each species to really inspect the BNI trait on each cereal crop. Tanaka et al. [109] tested the BNI activity in the root exudates of 36 different rice genotypes and found substantial genotypic variation for BNI, with the upland cultivar IAC25 expressing high BNI activity, whereas lowland varieties such as Nipponbare or IR64 exhibited low BNI activity.
Although initial evaluations in wheat showed that the roots of this cereal do not exudate chemical compounds to the soil that successfully control the nitrification process, further analysis of Leymus racemosus (mammoth wild rye, a wild wheat relative), resulted in the discovery of a high BNI capacity that efficiently suppressed soil nitrification [110]. The exploration of genetic diversity of wild predecessors of wheat showed how the BNI trait has been lost in the course of decades of breeding and selection of “desired” (at a given time and condition) agronomic conditions that inevitable lead to accidental loss of other valuable traits. As a result, the chromosomal location of the genes conferring the BNI trait has been identified and wheat varieties expressing the BNI trait have been formed by the production of wheat–Leymus racemosus chromosome addition lines [111].
The cereal crop in which significant BNI research has been conducted is sorghum, and both hydrophilic and hydrophobic root exudates with BNI activity have been identified [112, 113]. An ample natural diversity has also been identified among sorghum lines for the release of sorgoleone (a major hydrophobic root exudate with the highest BNI capacity). Nimbal et al. [114] evaluated sorgoleone production among 25 sorghum lines, finding nearly a 30-fold variation. Current efforts will focus on the exploration of sorghum diversity to exploit the BNI trait and to identify sorghum genetic stocks with high potential to release chemicals to the soil that suppress nitrification, reduce N2O emissions, and improve nitrogen use efficiency in sorghum-based production systems [113].
Cereal production uses the most of N-fertilizers; wheat itself accounts for a third of the global production [115] to cope with the food demand of a growing population. The indiscriminate use of N fertilizers results in N contaminants (e.g. nitrate and nitrite) that are causing major problems to bodies of water and are also contributing to global warming. BNI function represents a novel opportunity to naturally establish agricultural systems with improved N use efficiency and reduced N-contaminants. Therefore, the exploration of diversity of major cereal crops is imperative to promote the ecoefficiency of agricultural systems.
2.5 Impact of Nitrogen Pollutants on Biodiversity
Biodiversity is declining at an exceptional rate and on a worldwide scale. Indeed, loss of ecosystem functions and services associated with such declines has generated international debate [116–118]. Agricultural crops can be injured when exposed to high concentrations of various air, water, and soil pollutants. Air pollution affects plants in many ways, which have implications for overall biodiversity and ecology. There is evidence that air pollution can reduce some plants’ ability to reproduce, thus causing long-term changes to population ecology [119]. Of the different kinds of pollutants damaging the environment, nitrate pollution is a major problem along with the pollution of the atmosphere by ammonia and oxides of nitrogen. Nitrogen is a beneficial plant fertilizer in small amounts, but large amounts cause negative impacts on ecosystems and serious threat to biodiversity of many groups of organisms, including diversity of plants [120–122]. Nitrogen deposition refers to the input of reactive nitrogen species from the atmosphere to the biosphere. At the global scale, current N emission scenarios project that most regions will have increased rates of atmospheric N deposition in 2030 [123], which is causing concern about significant impacts on global plant biodiversity [116, 117, 124].
Even though low-to-medium levels of N addition (≤100 kg N/ha/year) generally did not alter plant diversity through time, high levels of N application significantly reduced species diversity [125]. The declines of diversity appeared to arise from N-related changes in soil properties, such as significant decreases in pH and extractable calcium (Ca) and increases in extractable aluminum (Al). Research reports revealed that N deposition may have shifted plant communities towards species composition typical of high-N availability. This shift has often been associated with a loss in diversity of plant species, particularly in areas with high deposition rates [126]. Mechanisms underlying the declines of diversity include competitive exclusion of more N-efficient dominant species by relatively fast-growing nitrophilic species, as a result of high-N availability induced by N deposition. Other such mechanisms include increased susceptibility to secondary stress and disturbance factors and species invasions [127, 128]. The research review of N-addition experiments across the tropics and subtropics have shown that N deposition may potentially affect plant diversity in some ecosystems more than originally thought, and because atmospheric N loads are gradually increasing in some tropical areas such as Asia, research on this topic is now urgently required [127].
Although improved agronomic approaches are one way to enhance nitrogen use efficiency (NUE) and reduce the N inputs, there is a growing interest in understanding the genetics of NUE in crop plants. Integration of these approaches may reduce both N deposition and the rapid loss of biodiversity.
2.6 Transgenic Crops and Food Security
The development of technologies for plant transformation started in the early 1980s following the production of chimeric genes [129–132], transformation vectors [133, 134], and DNA delivery systems [135–139], combined with plant tissue culture regeneration systems, which were pioneered by Murashige and Skoog [140]. This continued over the decades with the development of tissue culture for the three most important cereals: rice [141, 142], maize [143, 144], and wheat [145, 146]. The transformation of cereals since the 1990s was achieved with Agrobacterium-based protocols for rice [147], maize [148], and wheat [149], usually with a low-copy insertion of transgenes [150–152] and the gus reporter gene [153] with antibiotic selectable marker genes [154, 155]. These techniques were the most widely used in earlier transgenic plant research and crop development. Currently, more than 50 selectable marker genes are available for transgenic research and commercialization, and they can be divided into several categories depending on the mode of action and substrates used [156–158].
An important milestone in plant transgenic technology is the ability to generate plants free of the selectable marker gene using co-transformation with multiple T-DNAs [159, 160]. Among the major advances in plant transformation technology, we can consider the understanding of the plant cell–Agrobacterium relationship [161] and the molecular mechanisms of T-DNA transfer [162, 163] together with integration into the plant genome [164, 165].
Vain [166, 167] reviewed publication trends in and scientific knowledge on plant transgenic science and technologies available worldwide. In addition, the security of transgenic plants and products has been scrutinized [168–172], including the economic, environmental, and social dimensions [173].
The newest discoveries describing gene silencing [174], gene targeting [175], and RNA interference [176, 177], with alteration of the transcriptional activity of genes by using zinc-fingers nucleases [178, 179], together with the possibility of genome editing by TALENs (transcription activator-like effector nucleases) [180, 181], including the technology of using site-directed nucleases, are providing an opportunity to develop a new generation of transgenic crops with both improved traits and minimized potential for unintended effects that can impact safety [182].
It is very well known that agricultural production in the twenty-first century is going to face a number of new challenges [183–186]. A staple crop such as rice, for which more than half of the global population already depends on its production, will be under more and more pressure to increase its yield steadily; this has happened during the last four decades in some countries, where rice yield doubled or tripled [187]. New and multi-integrated strategies, including functional genomics, phenomics, and transgenics, coupled with conventional rice breeding, can help to develop new rice cultivars (referred to by Zhang [188] as Green Super Rice) and can meet future demand for world rice production. Take into consideration the fact that, from 1996 to 2012, millions of farmers in 30 countries worldwide adopted biotech/genetically modified organism (GMO) crops at an unprecedented level, going from 1.7 million ha in 1996 to 170 million ha in 2012 [189], a 100-fold increase. In addition, Golden Rice [190] is expected to be released in the Philippines in 2014, together with drought-tolerant sugarcane in Indonesia, biotech maize [191], and rice [192] in China. As a result, up to a billion poor people in rice households in Asia could benefit. Beyond 2015, it is difficult to predict what will come to the market, but expectations are also high in Africa, where the private/public partnership Water Efficient Maize for Africa [193] will release the first biotech drought-tolerant maize in the sub-Saharan region, where the need for drought-tolerant crops is greatest.
Commercialized crops could help to reduce the impacts of agriculture on biodiversity by alleviating pressure to convert additional land into agricultural use [194].
3 Summary and Conclusions
The management of plant genetic resources for food and agriculture is becoming a central issue in the context of sustainable agriculture, climate change, and food security. As the world faces unprecedented challenges, the exploration, use, and conservation of agricultural biodiversity will play a major role to address the most critical socioeconomical and environmental issues concerning agricultural research and food security strategies. The agricultural biodiversity of staple small grains, such as rice, maize, wheat, millets, and quinoa, is currently being studied from several perspectives to offer information to researchers and breeders worldwide.
Only a few accessions of pearl millets have been maintained so far in several countries. In-depth characterization and evaluation of these accessions remain insufficient due to lack of human and financial resources. Nevertheless, significant phenotypic diversity has been observed for most quantitative traits in a set of selected pearl millet germplasm accessions. This diversity is therefore very useful for use in crop improvement programs. Although the genetic variability in pearl millet landraces is large, some wild species have also been considered as a source of genes controlling agronomically important traits.
In the light of land change uses, modernization of agricultural practices, and climate change, which eventually affect plant diversity patterns, transnational efforts have focus on the establishment of extensive genebank collections for cereal crops. Both in situ and ex situ conservation of plant genetic resources are complementary strategies for the preservation of agricultural biodiversity. Geospatial analysis tools can be applied to map microsatellite diversity of cereal accessions characterized molecularly in order to identify priority geographic areas for conservation.
Although quinoa and its wild relatives are a reservoir of rich genetic variation that can be used by breeders for further improvement of the species, quinoa diversity is being lost. Market demand has encouraged the cultivation of commercially valuable varieties with certain characteristics, neglecting the immense variation found within the species. Some local quinoa genebanks have been created, containing many known quinoa varieties with a wide range of diverse agronomic and nutritional properties. However, current quinoa genebanks still do not represent the Andean diversity of the cultivated and wild relatives.
Agrobiodiversity can provide additional sources of new traits. Some cereal crops have the ability to inhibit soil nitrification and reduce the emissions of greenhouse gases (BNI activity) by releasing root exudates. The screening of some cereal accessions revealed genotypic variation for BNI. The exploration of agrobiodiversity for the BNI trait offers a novel strategy to improve the nitrogen use efficiency and increase crop productivity while reducing the environmental impacts of agriculture.
Chemical pollutants that are released into the environment are posing a threat to plant biodiversity. Nitrogen pollution is a major problem worldwide due to the indiscriminate use of synthetic nitrogen fertilizers in agricultural systems. This practice affects the natural balance of soil ecosystems, leading to significant losses of plant biodiversity as a result of changes in soil properties and plant community structure. Research addressing the genetics of nitrogen use efficiency in crop plants plus the adoption of sustainable agronomic practices offer an integrated approach to minimize the negative impact of reactive nitrogen on plant biodiversity and the environment.
Integration of transgenics, genomics, proteomics, and conventional plant breeding strategies can greatly accelerate the development of new cereal cultivars. Other than increasing crop productivity, commercialized genetically modified crops could help to reduce the impacts of agriculture on biodiversity by alleviating pressure to convert additional land into agricultural use.
Abbreviations
- AVRDC:
-
The World Vegetable Center
- BNI:
-
Biological Nitrification Inhibition
- CIAT:
-
International Center for Tropical Agriculture
- CGIAR:
-
Consultative Group on International Agricultural Research
- CGRP:
-
Canadian Genetic Resources Programme
- DNA:
-
Deoxyribonucleic acid
- FAO:
-
Food and Agriculture Organization of the United Nations
- GBIF:
-
Global Biodiversity Information Facility
- GCDT:
-
Global Crop Diversity Trust
- GEF:
-
Global Environment Facility
- GHG:
-
Greenhouse gas
- GM:
-
Genetically modified
- GMO:
-
Genetically modified organism
- IBPGR:
-
International Board for Plant Genetic Resources
- ICRISAT:
-
International Crops Research Institute for the Semi-Arid Tropics
- IFAD:
-
International Fund for Agricultural Development
- ILRI:
-
International Livestock Research Institute
- IRD:
-
Institut de Recherche pour le Développement
- ITPGRFA:
-
International Treaty on Plant Genetic Resources for Food and Agriculture
- IUCN:
-
International Union for Conservation of Nature
- JIRCAS:
-
Japan International Research Center for Agricultural Sciences
- MLS:
-
Multilateral system
- NBPGR:
-
National Bureau of Plant Genetic Resources (India)
- NUS:
-
Neglected and underutilized species
- ORSTOM:
-
Office de la Recherche Scientifique et Technique d’Outre-Mer
- SINGER:
-
System-wide Information Network for Genetic Resources
- SMTA:
-
Standard Material Transfer Agreement
- T-DNA:
-
Transfer-deoxyribonucleic acid
- TALENs:
-
Transcription activator-like effector nucleases
- UNEP:
-
United Nations Environment Programme
- UNU:
-
United Nations University
- USDA-ARS:
-
United States Department of Agriculture, Agricultural Research Service
- WEMA:
-
Water Efficient Maize for Africa
- WHO:
-
World Health Organization
References
CBD (2000) Agricultural biological diversity: review of phase I of the programme of work and adoption of a multi-year programme. http://www.cbd.int/decision/cop/?id=7147. Accessed 25 Feb 2013
Bioversity International (2009) Learning agrobiodiversity the importance of agricultural biodiversity and the role of universities. http://www.bioversityinternational.org/fileadmin/bioversityDocs/Training/Agrobiodiversity_Education/Learning_agrobiodiversity.pdf. Accessed 20 Feb 2013
FAO (1999) What is agrobiodiversity? http://www.fao.org/docrep/007/y5609e/y5609e01.htm#TopOfPage. Accessed 25 Feb 2013
Frison EA, Cherfas J, Hodgkin T (2011) Agricultural biodiversity is essential for a sustainable improvement in food and nutrition security. Sustainability 3:238–253. doi:10.3390/su3010238
van de Wouw M, Kik C, van Hintum T et al (2009) Genetic erosion in crops: concept, results and challenges. Plant Genet Res: Charact Utilization 8:1–15. doi:10.1017/S1479262109990062
Singh A, Singh HN, Singh J (2008) Rice biodiversity and its social implication. Int J Rural Stud 15(2)
Pistorius R (1997) Scientists, plants and politics—A history of the plant genetic resources movement. International Plant Genetic Resources Institute, Rome
Thormann I, Gaisberger H, Mattei F et al (2012) Digitization and online availability of original collecting mission data to improve data quality and enhance the conservation and use of plant genetic resources. Genet Resour Crop Evol 59:635–644
FAO (2010) The second report on the state of the world’s plant genetic resources for food and agriculture. http://www.fao.org/agriculture/seed/sow2/en/. Accessed 17 Apr 2013
Altieri MA, Merrick LC (1987) In situ conservation of crop genetic resources through maintenance of traditional farming. Econ Bot 41:86–96
Jensen HR, Dreiseitl A, Sadiki M et al (2012) The Red Queen and the seed bank: pathogen resistance of ex situ and in situ conserved barley. Evol Appl 5:353–367
Dulloo ME, Hunter D, Borelli T (2010) Ex situ and in situ conservation of agricultural biodiversity: major advances and research needs. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 38:123–135
FAO (2011a) Draft updated global plan of action for the conservation and sustainable utilization of plant genetic resources for food and agriculture. In: Fifth session of the Intergovernmental Technical Working Group on Plant Genetic Resources for Food and Agriculture, Rome, 27–29 Apr 2011
Pereira HM, Ferrier S, Walters M et al (2013) Essential biodiversity variables. Science 339:227–278
Cleveland DA, Soleri D (2007) Extending Darwin’s analogy: bridging differences in concepts of selection between farmers, biologists, and plant breeders. Econ Bot 61:121–136
Thomas M, Dawson JC, Goldringer I et al (2011) Seed exchanges, a key to analyze crop diversity dynamics in farmer-led on-farm conservation. Genet Resour Crop Evol 58:321–338
Briggs D, Walters M (1999) Evolution: some general considerations. Plant variation and evolution. 3rd edn. Cambridge University Press, Cambridge, pp 367–398
Mercer KL, Perales HR (2010) Evolutionary response of landraces to climate change in centers of crop diversity. Evol Appl 3:480–493
Zeven AC (1999) The traditional inexplicable replacement of seed and seed ware of landraces and cultivars: a review. Euphytica 110:181–191
Hajjar R, Jarvis DI, Gemmill-Herren B (2008) The utility of crop genetic diversity in maintaining ecosystem services. Agric Ecosyst Environ 123:261–270
Bezançon G, Pham JL, Deu M et al (2009) Changes in the diversity and geographic distribution of cultivated millet (Pennisetum glaucum (L.) R. Br.) and sorghum (Sorghum bicolor (L.) Moench) varieties in Niger between 1976 and 2003. Genet Resour Crop Evol 56:223–236
Vigouroux Y, Glaubitz JC, Matsuoka Y et al (2008) Population structure and genetic diversity of new world maize races assessed by DNA microsatellites. Am J Bot 95:1240–1253
Chan LM, Brown JL, Yoder AD (2011) Integrating statistical genetic and geospatial methods brings new power to phylogeography. Mol Phylogenet Evol 59:523–537
Kozak KH, Graham CH, Wiens JJ (2008) Integrating GIS-based environmental data into evolutionary biology. Trends Ecol Evol 23:141–148
van Zonneveld M, Dawson I, Thomas E, et al (2013) Application of molecular markers in spatial analysis to optimize in situ conservation of plant genetic resources. In: Tuberosa. R, Adler A, Frison E (eds) Advances in genomics of plant genetic resources. Springer, New York
Guarino L, Jarvis A, Hijmans RJ et al (2002) Geographic information systems (GIS) and the conservation and use of plant genetic resources. In: Engels JMM, Ramanatha Rao V, Brown AHD, Jackson MT (eds) Managing plant genetic diversity. International Plant Genetic Resources Institute (IPGRI), Rome, pp 387–404
Kiambi DK, Newbury HJ, Maxted N et al (2008) Molecular genetic variation in the African wild rice Oryza longistaminata A. Chev. et Roehr. and its association with environmental variables. Afr J Biotechnol 7:1446–1460
van Zonneveld M, Scheldeman X, Escribano P et al (2012) Mapping genetic diversity of cherimoya (Annona cherimola Mill.): application of spatial analysis for conservation and use of plant genetic resources. PloS ONE 7:e29845
Hijmans RJ, van Etten J (2012) Geographic analysis and modelling with raster data. R package “Raster”. http://cran.r-project.org/web/packages/raster/raster.pdf. Accessed 17 Apr 2013
Jombart T (2008) Adegenet: a R package for the multivariate analysis of genetic markers. Bioinformatics 24:1403–1405
Leberg PL (2002) Estimating allelic richness: effects of sample size and bottlenecks. Mol Ecol 11:2445–2449
Thomas E, van Zonneveld M, Loo J et al (2012) Present spatial diversity patterns of Theobroma cacao L. in the Neotropics reflect genetic differentiation in Pleistocene refugia followed by human-influenced dispersal. PLoS ONE 7:e47676
van Heerwaarden J, Doebley J, Briggs WH et al (2010) Genetic signals of origin, spread, and introgression in a large sample of maize landraces. Proc Natl Acad Sci. doi:10.1073/pnas.1013011108
Hufford MB, Lubinksy P, Pyhäjärvi T et al (2013) The genomic signature of crop-wild introgression in maize. PLoS Genet 9:e1003477
Russell J, Dawson IK, Flavell AJ et al (2011) Analysis of >1000 single nucleotide polymorphisms in geographically matched samples of landrace and wild barley indicates secondary contact and chromosome-level differences in diversity around domestication genes. New Phytol 191:564–578
Dvorak J, Luo M-Ch, Akhunov ED (2011) N.I. Vavilov’s theory of centres of diversity in the light of current understanding of wheat diversity, domestication and evolution. Czech J Genet Plant Breed 47:S20–S27
Barry MB, Pham JL, Courtois B et al (2007) Rice genetic diversity at farm and village levels and genetic structure of local varieties reveal need for in situ conservation. Genet Resour Crop Evol 54:1675–1690
Jarvis D, Hodgkin T, Sthapit BR et al (2011) An heuristic framework for identifying multiple ways of supporting the conservation and use of traditional crop varieties within the agricultural production system. Crit Rev Plant Sci 30:125–176
Pusadee T, Jamjoda S, Chiang Y-C et al (2009) Genetic structure and isolation by distance in a landrace of Thai rice. Proc Nat Acad Sci 106:13880–13885
Rice EB, Smith ME, Mitchell SE et al (2006) Conservation and change: a comparison of in situ and ex situ conservation of Jala maize germplasm. Crop Sci 46:428–436
Bellon MR, Hodson D, Hellin J (2011) Assessing the vulnerability of traditional maize seed systems in Mexico to climate change. Proc Natl Acad Sci 108:13432–13437
Tanksley SD, McCouch SR (1997) Seed banks and molecular maps: unlocking genetic potential from the wild. Science 227:1063–1066
Ishii T, Hiraoka T, Kanzaki T et al (2011) Evaluation of genetic variation among wild populations and local varieties of rice. Rice 4:170–177
Russell J, van Zonneveld M, Dawson IK, et al (2013) Genetic diversity and ecological niche modeling of wild barley: refugia, large-scale post-LGM range expansion and limited mid-future climate threats? PloS ONE (accepted)
National Research Council (NRC) (1996) Pearl millet. Lost Crops of Africa, vol 1, Grains. National Academy Press, Washington, pp 77–126
Yadav OP (2011) Project Coordinator’s Review (2010-11) on Pearl Millet Research. http://www.aicpmip.res.in/pcr2011.pdf. Accessed 02 May 2013
Brunken JN, de Wet JMJ, Harlan JR (1977) The morphology and domestication of pearl millet. Econ Botan 31:163–174
Rai KN, AppaRao S, Reddy KN (1997) Pearl Millet In: Fuccillo D, Sears L, Stapleton P (eds) Biodiversity in Trust. Cambridge University Press, Cambridge, pp 243–258
The Syngenta Foundation for Sustainable Agriculture (2006) Harnessing Modern Science in Africa to Sustain Sorghum and Pearl Millet Production for Resource poor Farmers. www.syngentafoundation.com/millet.html. Accessed 02 May 2013
Linnaeus C (1753) Species plantarum. Laurentius Salvius, Stockholm
Linnaeus C (1759) Systema naturae, 10th edn, vol 2. Laurentius Salvius, Stockholm
Rechard L (1805) Pennisetum. In: Person CH (ed) Synopsis plantarum,vol l. Cotta, Tubingen
Willdenow KL (1809) Enumeratio plantarum horti regni botanici Berolinensis. Berolini, Berlin
Steudel EG (1855) Synopsis plantarum glumaceum. Stuttgart. B65.10.01. M. inscription vol. 1: Ferd Muller, M.D., Ph.D. 1987
Leeke P (1907) Untersuchunguber Abstammung and Heimat der Negerhirse (Pennisetum americanum (L.)L schum.). Z Naturwissenchaften 79:1–108
Stapf O, Hubbard CE (1934) Pennisetum. In: Prain D (ed) Flora of tropical Africa, Part 9. Crown Agents, London
Clayton WD, Renvoize SA (1982) Gramineae. In: Polhill RM (ed) Flora of tropical East Africa, Part 3. Balkema, Rotterdam
Gari JA (2002) Review of the African millet diversity. In: Paper for the International workshop on fonio, food security and livelihood among the rural poor in West Africa, IPGRI/IFAD, Bamako, Mali. Programme for Neglected and Underutilized Species. IPGRI, Rome, Italy, 19–22 Nov 2001
Hanna WW (1987) Utilization of wild relatives of pearl millet. In: Proceedings of the international pearl millet wokshop, ICRISAT Center, India. ICRISAT, Patancheru, Andhra Pradesh, India, 7–11 Apr 1986
Oumar I, Mariac C, Pham JL et al (2008) Phylogeny and origin of pearl millet (Pennisetum glaucum [L.] R. Br) as revealed by microsatellite loci. Theor Appl Genet 117:489–497
Amblard S, Pernes J (1989) The identification of the cultivated pearl millet (Pennisetum) amongst plant impressions on pottery from Oued Chebbi (Dhar Oualata, Mauritania). Afr Archaeol Rev 7:117–126
D’Andrea AC, Casey J (2002) Pearl millet and kintampo subsistence. Afr Archaeol Rev 19:147–173
D’Andrea AC, Klee M, Casey J (2001) Archaeological evidence for pearl millet (Pennisetum glaucum) in sub-saharan West Africa. Antiquity 75:341–348
Clark JD (1962) The spread of food production in sub-Saharan Africa. J Afr Hist III 2:211–228
Harlan JR (1971) Agricultural origins: centres and non-centres. Science 174:468–474
Marchais L (1994) Wild pearl millet population (Pennisetum glaucum, Poaceae) integrity in agricultural Sahelian areas. An example from Keita (Niger). Plant Syst Evol 189:233–245
Portères R (1962) Berceaux agricolos primaries sur le continent africain. J Afr Hist III 2:195–210
Tostain S, Marchais L (1993) Wild pearl millet population (Pennisetum glaucum, Poaceae) integrity in agricultural Sahelian areas. Plant Syst Evol 189:233–245
Khairwal IS, Rai KN, Diwakar B et al (2007) Pearl millet: crop management and seed production manual. ICRISAT, Patancheru, Andhra Pradesh, India, pp 108
Tostain S (1992) Enzyme diversity in pearl millet (Pennisetum glaucum L.). Theor Appl Genet 83:736–742
Tostain S (1998) Le mil, une longue histoire: hypothèses sur sa domestication et ses migrations. In: Chastenet M (ed) Plantes et Paysages d’Afrique: Une Histoire à Explorer. Karthala and Centre de Recherches Africaines, Paris, pp 461–490
Klee M, Zach B, Stika HP (2004) Four thousand years of plant exploitation in the Lake Chad basin (Nigeria), part III: plant impressions in postherds from the final stone age Gajiganna culture. Veg Hist Archaeobotony 13:131–142
Tostain S, Riandey MF, Marchais L (1987) Enzyme diversity in pearl millet (Pennisetum glaucum) I. West Africa. Theor Appl Genet 74:188–193
Upadhyaya HD, Reddy KN, Gowda CLL (2007) Pearl millet germplasm at ICRISAT genebank—status and impact. International Crops Research Institute for the Semi-Arid Tropics (ICRISAT), Patancheru 502 324, Andhra Pradesh, India
IBPGR and ICRISAT (1993) Descriptors for Pearl Millet. International Plant genetic resources Institute, Rome
Mathur PN, Rana RS, Aggarwal RC (1993) Evaluation of pearl millet germplasm part I. National Bureau of Plant Genetic Resources, New Delhi
Mathur PN, Rana RS, Aggarwal RC (1993) Evaluation of pearl millet germplasm part II. National Bureau of Plant Genetic Resources, New Delhi
Andrews DJ, Anand Kumar K (1996) Use of the West African pearl millet landrace Iniadi in cultivar development. Plant Genet Resour Newsl 105:15–22
Bioversity International (2010) Key access and utilization descriptors for pearl millet genetic resources. Bioversity International, Rome, Italy. http://www.bioversityinternational.org/index.php?id=19&user_bioversitypublications_pi1%5BshowUid%5D=3376. Accessed 02 May 2013
Maxted N, Kell SP (2009) Establishment of a global network for the In Situ conservation of crop wild relatives: status and needs. FAO Commission on Genetic Resources for Food and Agriculture, Rome, p 266
Mujica A (1992) Granos y leguminosas andinas. In: Hernandez J, Bermejo J, Leon J (eds) Cultivos marginados: otra perspectiva de 1492. Organización de la Naciones Unidas para la Agricultura y la Alimentación (FAO), Roma
Rojas W, Pinto M, Soto JL (2010a) Distribución geográfica y variabilidad genética de los granos andinos. In: Rojas W, Soto JL, Pinto M, Jäger M, Padulosi S (eds) Granos Andinos. Avances, logros y experiencias desarrolladas en quinua, cañahua y amaranto en Bolivia. Bioversity International, Roma
Aroni G, Pinto M, Rojas W (2012) Small-scale quinoa processing technology in the southernAltiplano of Bolivia. In: Giuliani A, Hintermann F, Rojas W, Padulosi S (eds) Biodiversity of Andean grains: balancing market potential and sustainable livelihoods. Bioversity International, Rome
Carrasco E, Soto JL (2010) Importancia de los granos andinos. In: Rojas W, Soto JL, Pinto M, Jäger M, Padulosi S(eds) Granos Andinos. Avances, logros y experiencias desarrolladas en quinua, cañahua y amaranto en Bolivia. Bioversity International, Roma
Pinto M, Alarcón V, Soto JL, Rojas W (2010a) Usos tradicionales, no tradicionales e innovaciones agroindustriales de los granos andinos. In: Rojas W, Soto JL, Pinto M, Jäger M, Padulosi S (eds) Granos Andinos. Avances, logros y experiencias desarrolladas en quinua, cañahua y amaranto en Bolivia. Bioversity International, Roma
FAO (2011b) La Quinua: cultivo milenario para contribuir a la seguridad alimentaria mundial. FAO, Oficina Regional para América Latina y el Caribe, La Paz
Rojas W, Pinto M, Soto JL, Alcocer E (2010b) Valor nutricional, agroindustrial y funcional de los granos andinos. In: Rojas W, Soto JL, Pinto M, Jäger M, Padulosi S (eds) Granos Andinos. Avances, logros y experiencias desarrolladas en quinua, cañahua y amaranto en Bolivia. Bioversity International, Roma
FAO, WHO, UNU (1985) Necesidades de energía y proteínas. Informes Técnicos. OMS, Ginebra
Astudillo D (2012) Livelihoods of quinoa producers in southern Bolivia. In: Giuliani A, Hintermann F, Rojas W, Padulosi S (eds) Biodiversity of Andean grains: balancing market potential and sustainable livelihoods. Bioversity International, Italy
Hunter D, Heywood V (eds) (2011) Crop wild relatives: a manual of in situ conservation, 1st edn. Earthscan, London
Jacobsen SE (2003) The Worldwide Potential for Quinoa (Chenopodium quinoa Willd.). Food Rev Int 19:167–177
Apaza V, Estrada R, Quispe MG (2010b) Identificación y selección participativa de material genético promisorio. In: Bravo R, Valdivia R, Andrade K, Padulosi S, Jäger M (eds) Granos Andinos. Avances, logros y experiencias desarrolladas en quinua, cañihua y kiwicha en Perú. Bioversity International, Roma
Mujica A, Izquierdo J, Marathee JP (2001) Origen y Descripción de la Quinua. In: Mujica A, Jacobsen SE, Izquierdo J, Marathee JP. Quinua (Chenopodium quinoa Willd.). Ancestral Cultivo Andino, Alimento del Presente y Futuro. FAO. Santiago
Bonifacio A, Aroni G, Villca M (2012) Catálogo Etnobotánico de la Quinua Real. PROINPA, Cochabamba
Ministerio de Medio Ambiente y Agua (MMAyA), Viceministerio de Medio Ambiente, Biodiversidad y Cambios Climáticos (VMABCC), Bioversity International (2009) Libro Rojo de Parientes Silvestres de Cultivos de Bolivia. VMABCC-Bioversity International, La Paz
Apaza V, Catacora P, Quispe MG (2010a) Distribución geográfica y variabilidad genética de los granos andinos. In: Bravo R, Valdivia R, Andrade K, Padulosi S, Jäger M (eds). Granos Andinos. Avances, logros y experiencias desarrolladas en quinua, cañihua y kiwicha en Perú. Bioversity International, Roma
Rojas W, Pinto M (2010) Colecta de germoplasma. In: Rojas W, Soto JL, Pinto M, Jäger M, Padulosi S (eds) Granos Andinos. Avances, logros y experiencias desarrolladas en quinua, cañahua y amaranto en Bolivia. Bioversity International, Roma
Bravo R, Valdivia R, Andrade K, Padulosi S, Jäger M (eds) (2010) Granos Andinos. Avances, logros y experiencias desarrolladas en quinua, cañihua y kiwicha en Perú. Bioversity International, Roma
Jaeger M (2012) Novel products, markets and partnerships in value chains for Andean grains in Peru and Bolivia. In: Giuliani A, Hintermann F, Rojas W, Padulosi S (eds) Biodiversity of Andean grains: balancing market potential and sustainable livelihoods. Bioversity International, Rome
Bravo R, Catacora P (2010) Situación actual de los bancos nacionales de germoplasma. In: Bravo R, Valdivia R, Andrade K, Padulosi S, Jäger M (eds) Granos Andinos. Avances, logros y experiencias desarrolladas en quinua, cañihua y kiwicha en Perú. Bioversity International, Roma
Reay SD, Davidson EA, Smith KA et al (2012) Global agriculture and nitrous oxide emissions. Nat Clim Chang 2:410–416. doi:10.1038/nclimate1458
Ravishankara AR, Daniel JS, Portmann RW (2009) Nitrous oxide (N2O): the dominant ozone-depleting substance emitted in the 21st century. Science 326:123–125
Wuebbles DJ (2009) Nitrous oxide: no laughing matter. Science 326:56–57
Philippot L, Hallin S (2011) Towards food, feed and energy crops mitigating climate change. Trends Plant Sci 16:476–480
Canfield DE, Glazer AN, Falkowski PG (2010) The evolution and future of Earth’s nitrogen cycle. Science 330:192–196
Subbarao GV, Nakahara K, Hurtado MP et al (2009) Evidence for biological nitrification inhibition in Brachiaria pastures. Proc Natl Acad Sci 106:17302–17307
Subbarao GV, Sahrawat KL, Nakahara K et al (2012) A paradigm shift towards low-nitrifying production systems: the role of biological nitrification inhibition (BNI). Ann Bot. doi:10.1093/aob/mcs230
Subbarao GV, Rondon M, Ito O et al (2007) Biological nitrification inhibition (BNI)—Is it a widespread phenomenon? Plant Soil 294:5–18
Tanaka JP, Nardi P, Wissuwa M (2010) Nitrification inhibition activity, a novel trait in root exudates of rice. AoB Plants 2010:1–11
Subbarao GV, Ban T, Masahiro K et al (2007) Can biological nitrification inhibition (BNI) genes from perennial Leymus racemosus (Triticeae) combat nitrification in wheat farming? Plant Soil 299:55–64
Zahn LM (2007) A boost from wild wheat. Science 318:171. doi:10.1126/science.318.5848.171c
Hossain AKMZ, Subbarao GV, Pearse SJ et al (2008) Detection, isolation and characterization of a root-exuded compound, methyl 3-(4-hydroxyphenyl) propionate, responsible for biological nitrification inhibition by sorghum (Sorghum bicolor). New Phytol 180:442–451
Subbarao GV, Nakahara K, Ishikawa T et al (2013) Biological nitrification inhibition (BNI) activity in sorghum and its characterization. Plant Soil 366:243–259
Nimbal CI, Pedersen JF, Yerkes CN et al (1996) Phytotoxicity and distribution of sorgoleone in grain sorghum. J Agric Food Chem 44:1343–1347
Dobermann A, Cassman KG (2005) Cereal area and nitrogen use efficiency are drivers of future nitrogen fertilizer consumption. Sci China Life Sci 48:745–758
Phoenix GK, Hicks WK, Cinderby S et al (2006) Atmospheric nitrogen deposition in world biodiversity hotspots: the need for a greater global perspective in assessing N deposition impacts. Glob Change Biol 12:470–476
Sala OE, Chapin FS III, Armesto JJ et al (2000) Global biodiversity scenarios for the year 2100. Science 287:1770–1774
Zhou Z, Sun OJ, Huang J et al (2006) Land-use affects the relationship between species diversity and productivity at the local scale in a semi-arid steppe ecosystem. Funct Ecol 20:753–762
Cox RM (1992) Air pollution effects on plant reproductive processes and possible consequences to their population ecology. In: Barker JR, Tingey DT (eds) Air pollution effects on biodiversity. Springer, New York, pp 131–158
Clark CM, Tilman D (2008) Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature 451:712–715
Nordin A, Strengbom J, Ericson L (2006) Responses to ammonium and nitrate additions by boreal plants and their natural enemies. Environ Pollut 41:167–174
Strengbom J, Nordin A, Näsholm T, Ericson L (2002) Parasitic fungus mediates change in nitrogen-exposed boreal forest vegetation. J Ecol 90:61–67
Dentener F, Drevet J, Lamarque JF et al (2006) Nitrogen and sulfur deposition on regional and global scales: a multi-model evaluation. Glob Biogeochem Cycles 20:GB4003. doi: 10.1029/2005GB002672
Vitousek PM, Aber JD, Howarth RW et al (1997) Human alteration of the global nitrogen cycle: sources and consequences. Ecol Appl 7:737–750
Lu X, Mo J, Gilliam FS, Zhou G et al (2010) Effects of experimental nitrogen additions on plant diversity in an old-growth tropical forest. Glob Chang Biol 16:2688–2700
Bobbink R, Hornung M, Roelofs JGM (1998) The effects of air-borne nitrogen pollutants on species diversity in natural and semi-natural European vegetation. J Ecol 86:717–738
Bobbink R, Hicks K, Galloway J et al (2010) Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis. Ecol Appl 20:30–59
Gilliam FS, Hockenberry AW, Adams MB (2006) Effects of atmospheric nitrogen deposition on the herbaceouslayer of a central Appalachian hardwood forest. J Torrey Bot Soc 133:240–254
Bevan MW, Flavell RB, Chilton MD (1983) A chimeric antibiotic-resistance gene as a selectable marker for plant cell transformation. Nature 304:184–187
Fraley RT, Rogers SG, Horsch RB et al (1983) Expression of bacterial genes in plant cells. Proc Natl Acad Sci 80:4803–4807
Herrera-Estrella L, Depicker A, Van Montagu M et al (1983) Expression of chimaeric genes transferred into plant cells using a Ti-plasmid-derived vector. Nature 303:209–213
Horsch RB, Fry JE, Hoffmann NL et al (1985) A simple and general method for transferring genes into plants. Science 227:1229–1231
Bevan M (1984) Binary Agrobacterium vectors for plant transformation. Nucleic Acids Res 12:8711–8721
Hoekema A, Hirsch PR, Hooykaas PJJ et al (1983) A binary plant vector strategy based on separation of Vir-region and T-region of the Agrobacterium tumefaciens Ti-plasmid. Nature 303:179–180
Chilton MD, Tepfer DA, Petit A et al (1982) Agrobacterium rhizogenes inserts T-DNA into the genomes of the host plant root cells. Nature 295:432–434
Draper J, Davey MR, Freeman JP et al (1982) Ti plasmid homologous sequences present in tissues from Agrobacterium plasmid transformed petunia protoplasts. Plant Cell Physiol 23:451–458
Krens FA, Molendijk L, Wullems GJ et al (1982) In vitro transformation of plant protoplasts with Ti-plasmid DNA. Nature 296:72–74
Paszkowski J, Shillito RD, Saul M et al (1984) Direct gene transfer to plants. EMBO J 3:2717–2722
Sanford JC, Klein TM, Wolf ED et al (1987) Delivery of substances into cells and tissues using a particle bombardment process. J Part Sci Technol 5:27–37
Murashige T, Skoog F (1962) A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol Plant 15:473–497
Bajaj YPS (1991) Biotechnology in rice improvement. In: Bajaj YPS (ed) Biotechnology in Agriculture and Forestry, vol 14. Rice. Springer, Berlin, pp 1–15
Shimammoto K, Tareda R, Izawa H (1989) Fertile transgenic rice plants regenerated from transformed protoplast. Nature 338:274–276
Bajaj YPS (1994) Biotechnology in maize improvement. In: Bajaj YPS (ed) Biotechnology in Agriculture and Forestry vol. 25. Maize. Springer, Berlin, pp 1–17
Gordon-Kamm WJ, Spencer TM, Mangano ML et al (1990) Transformation of maize cells and regeneration of fertile transgenic plants. Plant Cell 2:603–618
Bajaj YPS (ed) (1990) Biotechnology in agriculture and forestry, vol. 13. Wheat. Springer, Berlin
Vasil V, Castillo AM, From ME et al (1992) Herbicide resistant fertile transgenic wheat plants obtained by microprojectil bombardment of regenerable embryogenic callus. Biotechnol 10:667–674
Hiei Y, Ohta S, Komari T et al (1994) Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J 6:271–282
Ishida V, Saito H, Ohta O et al (1996) High efficiency transformation of maize (Zea mays L.) mediated by Agrobacterium tumefaciens. Nat Biotechnol 6:745–750
Cheng M, Fry JE, Pang S et al (1997) Genetic Transformation of Wheat Mediated by Agrobacterium tumefaciens. Plant Physiol 115:971–980
Afolabi AS, Worland B, Snape JW et al (2004) A large-scale study of rice plants transformed with different T-DNAs provides new insights into locus composition and T-DNA linkage configurations. Theor Appl Genet 109:815–826
Ingham DJ, Beer S, Money S et al (2001) Quantitative Real-Time PCR assay for determining transgene copy number in transformed plants. Biotechniques 31:132–140
Vain P, Afolabi AS, Worland B et al (2003) Transgene behavior in populations of rice plants transformed using a new dual binary vector system: pGreen/pSoup. Theor Appl Genet 107:210–217
Jefferson RA, Burgess SM, Hirsh D (1986) Beta-glucuronidase from Escherichia coli as a gene-fusion marker. Proc Natl Acad Sci 83:8447–8451
Carrer H, Hockenberry TN, Svab Z et al (1993) Kanamycin resistance as a selectable marker for plastid transformation in tobacco. Mol Gen Genet 241:49–56
Waldron C, Murphy EB, Roberts JL et al (1985) Resistance to hygromycin B. Plant Mol Biol 5:103–108
Miki B, McHugh S (2003) Selectable marker genes in transgenic plants: application, alternatives and biosafety. J Biotechnol 107:193–232
Padilla IMG, Burgos L (2010) Aminoglycoside antibiotics: structure, functions and effects on in vitro plant culture and genetic transformation protocols. Plant Cell Rep 29:1203–1213
Sundar IK, Sakthitel N (2008) Advances in selectable marker genes for plant transformation. J Plant Physiol 165:1698–1716
Komari T, Hiei Y, Saito Y et al (1996) Vectors carrying two separate T-DNAs for co-transformation of higher plants mediated by Agrobacterium tumefaciens and segregation of transformants free from selection markers. Plant J 10:165–174
Miller M, Tagliani L, Wang N et al (2002) High efficiency transgene segregation in co-transformed maize plants using an Agrobacterium tumefaciens 2 T-DNA binary system. Transgenic Res 11:381–396
Gelvin SB (2003) Improving plant genetic engineering by manipulating the host. Trends Biotechnol 21:95–98
Tzfira T, Citovsky V (2006) Agrobacterium-mediated genetic transformation of plants: biology and biotechnology. Curr Opin Biotechnol 17:147–154
Zambryski P (1992) Chronicles from the Agrobacterium–plant cell DNA transfer story. Ann Rev Plant Physiol Mol Biol 43:4645–4690
Gelvin SB (2010) Plant proteins involved in Agrobacterium mediated genetic transformation. Ann Rev Phytopatol 48:45–68
Gelvin SB (2010b) Finding the way to the nucleus.Curr Opin Microbiol 13:53–58
Vain P (2005) Plant transgenic science knowledge. Nat Biotechnol 23:1348–1349
Vain P (2007) Thirty years of plant transformation technology development. Plant Biotechnol J 5:221–229
Areal FJ, Riesgo L, Rodríguez-Cerezo E (2011) Attitudes of European farmers towards GM crop adoption. Plant Biotechnol J 9:945–957
EU Commission (2010) A decade of EU funded GMO research (2001–2010). ftp://cordis.europa.eu/pub/fp7/kbbe/docs/a-decade-of-eu-funded-gmo-research_en.pdf. Accessed 02 May 2013
European Policy Evaluation Consortium (EPEC) (2011) The Evaluation of the EU legislative framework in the field of cultivation of GMOs under Directive 2001/18/EC. http://ec.europa.eu/food/food/biotechnology/evaluation/docs/gmo_cultivation_report_en.pdf. Accessed 02 May 2013
Lusser M, Rodríguez-Cerezo E (2012) Comparative regulatory approaches for new plant breeding techniques. Workshop proceedings European Commission, JRC Technical Report EUR 25237 EN (2012)
Phillips PWB (2002) Biotechnology in the global agri-food systems. Trends Biotechnol 20:376–381
Park JR, McFarlane I, Phipps RH et al (2011) The role of transgenic crops in sustainable development. Plant Biotechol J 9:2–21
Dunoyer P, Himber C, Voinnet O (2006) Induction, suppression and requirement of RNA silencing pathways in virulent Agrobacterium tumefaciens infections. Nat Genet 38:258–263
Shaked H, Melamed-Bessudo C, Levy AA (2005) High frequency gene targeting in Arabidopsis plants expressing the yeast RAD54 gene. Proc Natl Acad Sci 12:265–269
Miki D, Itoh R, Shimamoto K (2005) RNA silencing of single and multiple members in a gene family of rice. Plant Physiol 138:1903–1913
Reynolds A, Leake D, Boese Q et al (2004) Rational sRNAi design for RNA interterference. Nat Biotechnol 22:326–330
Carroll D (2011) Genome engineering with zinc-finger nucleases. Genetics 188:773–782
Shukla VK, Doyon Y, Miller JC et al (2009) Precise genome modification in the crop species Zea mays using zinc-finger nucleases. Nature 459:437–441
Boch J, Scholze H, Schornack S (2009) Breaking the code of DNA binding specificity of TAL-type III effectors. Science 326:1509–1512
Bogdanove AJ, Voytas JF (2011) TAL effectors: customizable proteins for DNA targeting. Science 333:1843–1846
Podevin N, Davies HV, Hartung F (2013) Site-directed nucleases: a paradigm shift in predictable, knowledge-based plant breeding. Trends Biotechnol. doi:10.1016/j.tibtech.2013.03.004
Chen H, Lin Y, Zhang Q (2010) Rice. In: Kempker F, Jung Ch (eds) Genetic modification of plants. Biotechnology in Agriculture and Forestry, vol 64. Springer, Berlin, p 423–451
Hirano HY, Hirai A, Sano Y, Sasaki T (eds) (2008) Rice Biology in the Genomics era. Biotechnol in Agriculture and Forestry, vol 62. Springer, Berlin
Serraj R, Bennett J, Hardy B (eds) (2008) Drought frontiers in rice: crop improvement for increased rainfed production. World Scientific Publishing, Singapore
Sheehy JE, Mitchell PL, Hardy B (eds) (2007) Charting new pathways to C4 rice. IRRI, World Scientific, Los Banos
FAOSTAT (2013) http://faostat.fao.org. Accessed 05 Mar 2013
Zhang Q (2007) Strategies for developing Green Super Rice. Proc Natl Acad Sci 16:16402–16409
James C (2012) Global status of commercialized Biotech/GMO crops: 2012. ISAAA Briefs No. 44. ISAAA, Ithaca, NY
Datta SK, Datta K, Parkhi V et al (2007) Golden rice: introgression, breeding, and field evaluation. Euphytica 154:271–278
D’Halluin K, Vanderstraeten Ch, Stals E et al (2008) Homologous recombination: a basis for targeted genome optimization in crop species such as maize. Plant Biotechnol J 6:93–102
Duan Y, Zhai Ch, Li H (2012) An efficient and high-throughput protocol for Agrobacterium mediated transformation based on phosphomannose isomerase positive selection in Japonica rice (Oryza sativa L.). Plant Cell Rep 31:1611–1624
Water Efficient Maize for Africa (2011) Water efficient maize for Africa: pushing GMO crops onto Africa. The African Center for Biosafety. http://www.monsanto.com/improvingagriculture/Pages/water-efficient-maize-for-africa.aspx. Accessed 10 May 2013
Carpenter JE (2011) Impact of GM crops on biodiversity. GM Crops 2:7–23
Aroni J, Aroni G, Quispe R, Bonifacio A (2003) Catálogo de Quinoa Real. Fundación PROINPA, La Paz
Pinto M, Marin W, Rojas W (2010b) Estrategias para la conservación y promoción de los granos andinos: ferias y concursos. In: Rojas W, Soto JL, Pinto M, Jäger M, Padulosi S (eds) Granos Andinos. Avances, logros y experiencias desarrolladas en quinua, cañahua y amaranto en Bolivia. Bioversity International, Roma
Soto JL (2010) Tecnología del cultivo de granos andinos. In: Rojas W, Soto JL, Pinto M, Jäger M, Padulosi S (eds) Granos Andinos. Avances, logros y experiencias desarrolladas en quinua, cañahua y amaranto en Bolivia. Bioversity International, Roma
Knudsen H (ed) (2000) Directorio de Colecciones de Germoplasma en América Latina y el Caribe, 1st edn. International Plant Genetic Resources Institute (IPGRI), Roma
Mazón N, Rivera M, Peralta E, Estrella J, Tapia C (2002) Catálogo del banco de germoplasma de quinua (Chenopodium quinoa Willd.) del INIAP—Ecuador. Programa Nacional de Leguminosas y Granos Andinos, Departamento Nacional de Recursos Fitogenéticos y Biotecnología, Estación Experimental Santa Catalina. Quito
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Moreta, D.E. et al. (2013). Current Issues in Cereal Crop Biodiversity. In: Mukherjee, J. (eds) Biotechnological Applications of Biodiversity. Advances in Biochemical Engineering/Biotechnology, vol 147. Springer, Berlin, Heidelberg. https://doi.org/10.1007/10_2013_263
Download citation
DOI: https://doi.org/10.1007/10_2013_263
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-662-45096-3
Online ISBN: 978-3-662-45097-0
eBook Packages: Chemistry and Materials ScienceChemistry and Material Science (R0)